PA28αβ: The Enigmatic Magic Ring of the Proteasome?

{kind=link}
{kind=link}
Abstract
:1. MHC Class I Antigen Presentation
2. Proteasome Activator PA28αβ
2.1. Structure
2.2. Role in MHC Class I Antigen Presentation
2.3. Biochemical Properties
2.3.1. Hydrolysis of Fluorogenic Peptide Substrates
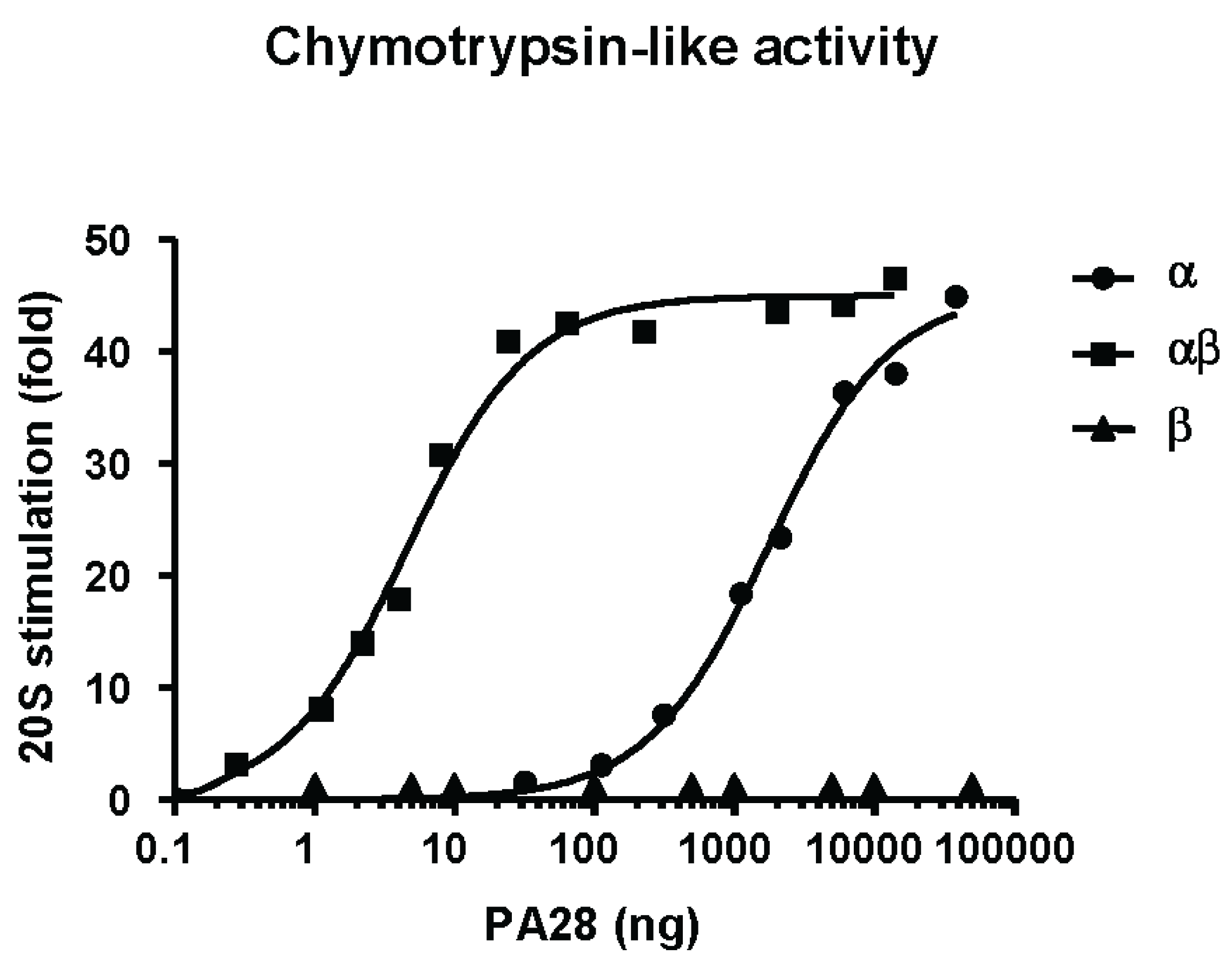
2.3.2. Gate Opening Mechanism
2.3.3. Effects of PA28αβ on Protein Degradation
2.3.4. Effects of PA28αβ on Peptide Products Generation
2.3.5. PA28 as a Smart Sieve
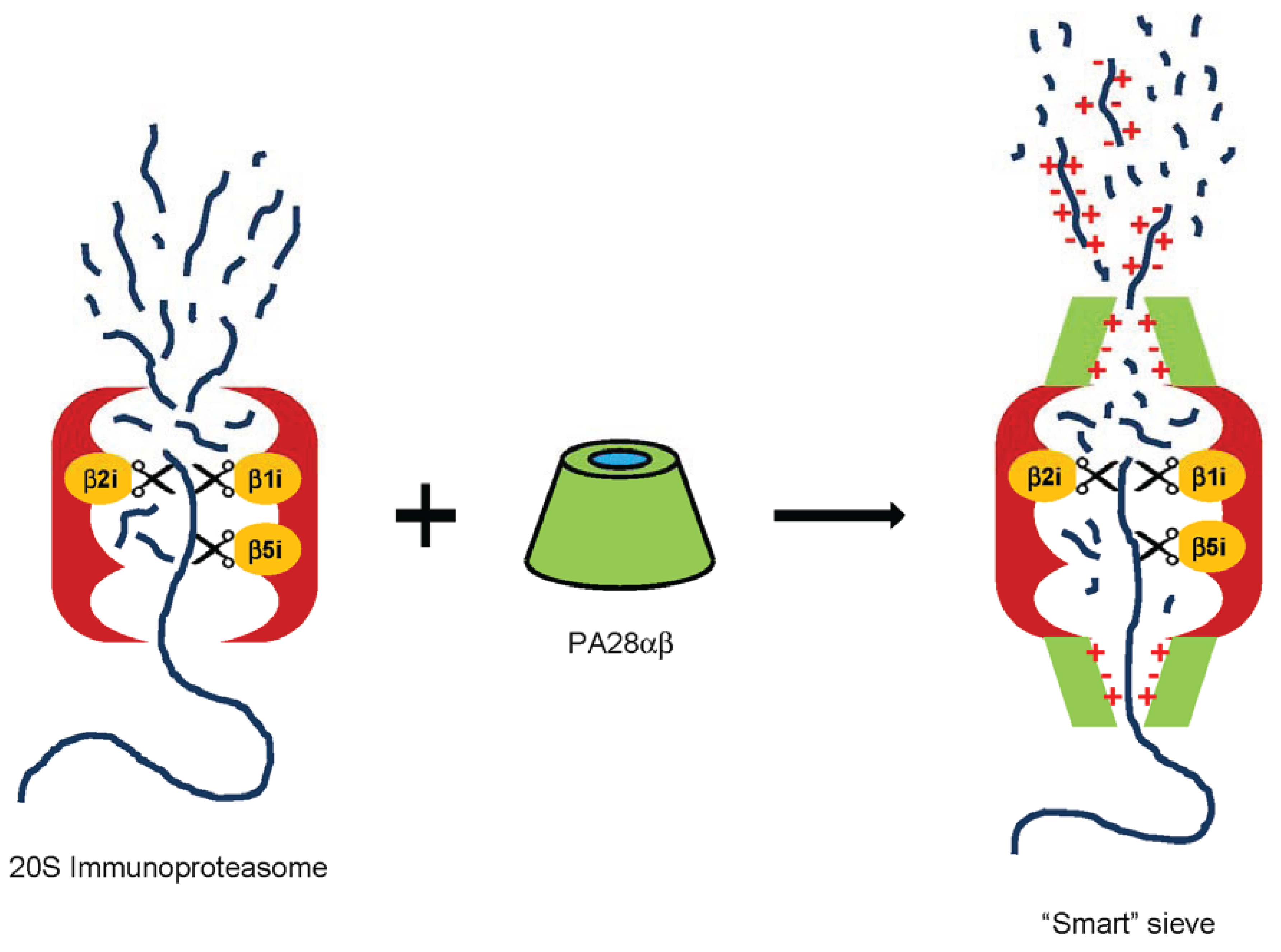
2.4. Other Potential Biological Functions
3. Conclusions
Acknowledgments
Conflicts of Interest
References
- Pamer, E.; Cresswell, P. Mechanisms of MHC class I—Restricted antigen processing. Annu. Rev. Immunol. 1998, 16, 323–358. [Google Scholar] [CrossRef]
- Rock, K.L.; Goldberg, A.L. Degradation of cell proteins and the generation of MHC class I—Presented peptides. Annu. Rev. Immunol. 1999, 17, 739–779. [Google Scholar]
- Goldberg, A.L.; Cascio, P.; Saric, T.; Rock, K.L. The importance of the proteasome and subsequent proteolytic steps in the generation of antigenic peptides. Mol. Immunol. 2002, 39, 147–164. [Google Scholar] [CrossRef]
- Yewdell, J.W. Not such a dismal science: The economics of protein synthesis, folding, degradation and antigen processing. Trends Cell Biol. 2001, 11, 294–297. [Google Scholar] [CrossRef]
- Abele, R.; Tampe, R. The ABCs of immunology: Structure and function of TAP, the transporter associated with antigen processing. Physiology 2004, 19, 216–224. [Google Scholar] [CrossRef]
- Kloetzel, P.M. Antigen processing by the proteasome. Nat. Rev. Mol. Cell Biol. 2001, 2, 179–187. [Google Scholar] [CrossRef]
- Baumeister, W.; Walz, J.; Zuhl, F.; Seemuller, E. The proteasome: Paradigm of a self-compartmentalizing protease. Cell 1998, 92, 367–380. [Google Scholar] [CrossRef]
- Voges, D.; Zwickl, P.; Baumeister, W. The 26S proteasome: A molecular machine designed for controlled proteolysis. Annu. Rev. Biochem. 1999, 68, 1015–1068. [Google Scholar] [CrossRef]
- Glickman, M.H.; Ciechanover, A. The ubiquitin-proteasome proteolytic pathway: Destruction for the sake of construction. Physiol. Rev. 2002, 82, 373–428. [Google Scholar]
- Coux, O.; Tanaka, K.; Goldberg, A.L. Structure and functions of the 20S and 26S proteasomes. Annu. Rev. Biochem. 1996, 65, 801–847. [Google Scholar]
- Tanaka, K.; Kasahara, M. The MHC class I ligand-generating system: Roles of immunoproteasomes and the interferon-gamma-inducible proteasome activator PA28. Immunol. Rev. 1998, 163, 161–176. [Google Scholar] [CrossRef]
- Sijts, E.J.; Kloetzel, P.M. The role of the proteasome in the generation of MHC class I ligands and immune responses. Cell. Mol. Life Sci. 2011, 68, 1491–1502. [Google Scholar] [CrossRef]
- Kincaid, E.Z.; Che, J.W.; York, I.; Escobar, H.; Reyes-Vargas, E.; Delgado, J.C.; Welsh, R.M.; Karow, M.L.; Murphy, A.J.; Valenzuela, D.M.; et al. Mice completely lacking immunoproteasomes show major changes in antigen presentation. Nat. Immunol. 2012, 13, 129–135. [Google Scholar]
- Groettrup, M.; Kirk, C.J.; Basler, M. Proteasomes in immune cells: More than peptide producers? Nat. Rev. Immunol. 2010, 10, 73–78. [Google Scholar] [CrossRef]
- Cerruti, F.; Martano, M.; Petterino, C.; Bollo, E.; Morello, E.; Bruno, R.; Buracco, P.; Cascio, P. Enhanced expression of interferon-gamma-induced antigen-processing machinery components in a spontaneously occurring cancer. Neoplasia 2007, 9, 960–969. [Google Scholar] [CrossRef]
- Egerer, T.; Martinez-Gamboa, L.; Dankof, A.; Stuhlmuller, B.; Dorner, T.; Krenn, V.; Egerer, K.; Rudolph, P.E.; Burmester, G.R.; Feist, E. Tissue-specific up-regulation of the proteasome subunit beta5i (LMP7) in Sjogren’s syndrome. Arthritis Rheum. 2006, 54, 1501–1508. [Google Scholar] [CrossRef]
- Mishto, M.; Bellavista, E.; Santoro, A.; Stolzing, A.; Ligorio, C.; Nacmias, B.; Spazzafumo, L.; Chiappelli, M.; Licastro, F.; Sorbi, S.; et al. Immunoproteasome and LMP2 polymorphism in aged and Alzheimer’s disease brains. Neurobiol. Aging 2006, 27, 54–66. [Google Scholar] [CrossRef]
- Raule, M.; Cerruti, F.; Cascio, P. Enhanced rate of degradation of basic proteins by 26S immunoproteasomes. BBA Cell Res. 2014. [Google Scholar] [CrossRef]
- Dubiel, W.; Pratt, G.; Ferrell, K.; Rechsteiner, M. Purification of an 11S regulator of the multicatalytic protease. J. Biol. Chem. 1992, 267, 22369–22377. [Google Scholar]
- Ma, C.P.; Slaughter, C.A.; DeMartino, G.N. Identification, purification, and characterization of a protein activator (PA28) of the 20S proteasome (macropain). J. Biol. Chem. 1992, 267, 10515–10523. [Google Scholar]
- Cascio, P.; Call, M.; Petre, B.M.; Walz, T.; Goldberg, A.L. Properties of the hybrid form of the 26S proteasome containing both 19S and PA28 complexes. EMBO J. 2002, 21, 2636–2645. [Google Scholar] [CrossRef]
- Hendil, K.B.; Khan, S.; Tanaka, K. Simultaneous binding of PA28 and PA700 activators to 20S proteasomes. Biochem. J. 1998, 332, 749–754. [Google Scholar]
- Kopp, F.; Dahlmann, B.; Kuehn, L. Reconstitution of hybrid proteasomes from purified PA700–20S complexes and PA28alphabeta activator: Ultrastructure and peptidase activities. J. Mol. Biol. 2001, 313, 465–471. [Google Scholar] [CrossRef]
- Tanahashi, N.; Murakami, Y.; Minami, Y.; Shimbara, N.; Hendil, K.B.; Tanaka, K. Hybrid proteasomes. Induction by interferon-gamma and contribution to ATP-dependent proteolysis. J. Biol. Chem. 2000, 275, 14336–14345. [Google Scholar]
- Ahn, J.Y.; Tanahashi, N.; Akiyama, K.; Hisamatsu, H.; Noda, C.; Tanaka, K.; Chung, C.H.; Shibmara, N.; Willy, P.J.; Mott, J.D.; et al. Primary structures of two homologous subunits of PA28, a gamma-interferon-inducible protein activator of the 20S proteasome. FEBS Lett. 1995, 366, 37–42. [Google Scholar] [CrossRef]
- Ahn, K.; Erlander, M.; Leturcq, D.; Peterson, P.A.; Fruh, K.; Yang, Y. In vivo characterization of the proteasome regulator PA28. J. Biol. Chem. 1996, 271, 18237–18242. [Google Scholar]
- Honore, B.; Leffers, H.; Madsen, P.; Celis, J.E. Interferon-gamma up-regulates a unique set of proteins in human keratinocytes. Molecular cloning and expression of the cDNA encoding the RGD-sequence-containing protein IGUP I-5111. Eur. J. Biochem. 1993, 218, 421–430. [Google Scholar] [CrossRef]
- Jiang, H.; Monaco, J.J. Sequence and expression of mouse proteasome activator PA28 and the related autoantigen Ki. Immunogenetics 1997, 46, 93–98. [Google Scholar] [CrossRef]
- Realini, C.; Dubiel, W.; Pratt, G.; Ferrell, K.; Rechsteiner, M. Molecular cloning and expression of a gamma-interferon-inducible activator of the multicatalytic protease. J. Biol. Chem. 1994, 269, 20727–20732. [Google Scholar]
- Rechsteiner, M.; Realini, C.; Ustrell, V. The proteasome activator 11S REG (PA28) and class I antigen presentation. Biochem. J. 2000, 345, 1–15. [Google Scholar] [CrossRef]
- Tanahashi, N.; Yokota, K.; Ahn, J.Y.; Chung, C.H.; Fujiwara, T.; Takahashi, E.; DeMartino, G.N.; Slaughter, C.A.; Toyonaga, T.; Yamamura, K.; et al. Molecular properties of the proteasome activator PA28 family proteins and gamma-interferon regulation. Genes Cells 1997, 2, 195–211. [Google Scholar]
- Mao, I.; Liu, J.; Li, X.; Luo, H. REGgamma, a proteasome activator and beyond? Cell. Mol. Life Sci. 2008, 65, 3971–3980. [Google Scholar] [CrossRef]
- Mott, J.D.; Pramanik, B.C.; Moomaw, C.R.; Afendis, S.J.; DeMartino, G.N.; Slaughter, C.A. PA28, an activator of the 20S proteasome, is composed of two nonidentical but homologous subunits. J. Biol. Chem. 1994, 269, 31466–31471. [Google Scholar]
- Knowlton, J.R.; Johnston, S.C.; Whitby, F.G.; Realini, C.; Zhang, Z.; Rechsteiner, M.; Hill, C.P. Structure of the proteasome activator REGalpha (PA28alpha). Nature 1997, 390, 639–643. [Google Scholar] [CrossRef]
- Zhang, Z.; Clawson, A.; Realini, C.; Jensen, C.C.; Knowlton, J.R.; Hill, C.P.; Rechsteiner, M. Identification of an activation region in the proteasome activator REGalpha. Proc. Natl. Acad. Sci. USA 1998, 95, 2807–2811. [Google Scholar]
- Li, J.; Gao, X.; Joss, L.; Rechsteiner, M. The proteasome activator 11S REG or PA28: Chimeras implicate carboxyl-terminal sequences in oligomerization and proteasome binding but not in the activation of specific proteasome catalytic subunits. J. Mol. Biol. 2000, 299, 641–654. [Google Scholar] [CrossRef]
- Ma, C.P.; Willy, P.J.; Slaughter, C.A.; DeMartino, G.N. PA28, an activator of the 20S proteasome, is inactivated by proteolytic modification at its carboxyl terminus. J. Biol. Chem. 1993, 268, 22514–22519. [Google Scholar]
- Song, X.; Mott, J.D.; von Kampen, J.; Pramanik, B.; Tanaka, K.; Slaughter, C.A.; DeMartino, G.N. A model for the quaternary structure of the proteasome activator PA28. J. Biol. Chem. 1996, 271, 26410–26417. [Google Scholar]
- Realini, C.; Jensen, C.C.; Zhang, Z.; Johnston, S.C.; Knowlton, J.R.; Hill, C.P.; Rechsteiner, M. Characterization of recombinant REGalpha, REGbeta, and REGgamma proteasome activators. J. Biol. Chem. 1997, 272, 25483–25492. [Google Scholar]
- Bose, S.; Mason, G.G.; Rivett, A.J. Phosphorylation of proteasomes in mammalian cells. Mol. Biol. Rep. 1999, 26, 11–14. [Google Scholar] [CrossRef]
- Li, N.; Lerea, K.M.; Etlinger, J.D. Phosphorylation of the proteasome activator PA28 is required for proteasome activation. Biochem. Biophys. Res. Commun. 1996, 225, 855–860. [Google Scholar] [CrossRef]
- Realini, C.; Rechsteiner, M. A proteasome activator subunit binds calcium. J. Biol. Chem. 1995, 270, 29664–29667. [Google Scholar] [CrossRef]
- Johnston, S.C.; Whitby, F.G.; Realini, C.; Rechsteiner, M.; Hill, C.P. The proteasome 11S regulator subunit REG alpha (PA28 alpha) is a heptamer. Protein Sci. 1997, 6, 2469–2473. [Google Scholar]
- Sugiyama, M.; Sahashi, H.; Kurimoto, E.; Takata, S.; Yagi, H.; Kanai, K.; Sakata, E.; Minami, Y.; Tanaka, K.; Kato, K. Spatial arrangement and functional role of alpha subunits of proteasome activator PA28 in hetero-oligomeric form. Biochem. Biophys. Res. Commun. 2013, 432, 141–145. [Google Scholar] [CrossRef]
- Zhang, Z.; Krutchinsky, A.; Endicott, S.; Realini, C.; Rechsteiner, M.; Standing, K.G. Proteasome activator 11S REG or PA28: Recombinant REG alpha/REG beta hetero-oligomers are heptamers. Biochemistry 1999, 38, 5651–5658. [Google Scholar]
- Gray, C.W.; Slaughter, C.A.; DeMartino, G.N. PA28 activator protein forms regulatory caps on proteasome stacked rings. J. Mol. Biol. 1994, 236, 7–15. [Google Scholar] [CrossRef]
- Macagno, A.; Gilliet, M.; Sallusto, F.; Lanzavecchia, A.; Nestle, F.O.; Groettrup, M. Dendritic cells up-regulate immunoproteasomes and the proteasome regulator PA28 during maturation. Eur. J. Immunol. 1999, 29, 4037–4042. [Google Scholar] [CrossRef]
- Ossendorp, F.; Fu, N.; Camps, M.; Granucci, F.; Gobin, S.J.; van den Elsen, P.J.; Schuurhuis, D.; Adema, G.J.; Lipford, G.B.; Chiba, T.; et al. Differential expression regulation of the alpha and beta subunits of the PA28 proteasome activator in mature dendritic cells. J. Immunol. 2005, 174, 7815–7822. [Google Scholar] [CrossRef]
- Groettrup, M.; Soza, A.; Eggers, M.; Kuehn, L.; Dick, T.P.; Schild, H.; Rammensee, H.G.; Koszinowski, U.H.; Kloetzel, P.M. A role for the proteasome regulator PA28alpha in antigen presentation. Nature 1996, 381, 166–168. [Google Scholar] [CrossRef]
- Yamano, T.; Murata, S.; Shimbara, N.; Tanaka, N.; Chiba, T.; Tanaka, K.; Yui, K.; Udono, H. Two distinct pathways mediated by PA28 and hsp90 in major histocompatibility complex class I antigen processing. J. Exp. Med. 2002, 196, 185–196. [Google Scholar] [CrossRef]
- Schwarz, K.; van den Broek, M.; Kostka, S.; Kraft, R.; Soza, A.; Schmidtke, G.; Kloetzel, P.M.; Groettrup, M. Overexpression of the proteasome subunits LMP2, LMP7, and MECL-1, but not PA28 alpha/beta, enhances the presentation of an immunodominant lymphocytic choriomeningitis virus T cell epitope. J. Immunol. 2000, 165, 768–778. [Google Scholar] [CrossRef]
- Stohwasser, R.; Salzmann, U.; Giesebrecht, J.; Kloetzel, P.M.; Holzhutter, H.G. Kinetic evidences for facilitation of peptide channelling by the proteasome activator PA28. Eur. J. Biochem. 2000, 267, 6221–6230. [Google Scholar] [CrossRef]
- Sun, Y.; Sijts, A.J.; Song, M.; Janek, K.; Nussbaum, A.K.; Kral, S.; Schirle, M.; Stevanovic, S.; Paschen, A.; Schild, H.; et al. Expression of the proteasome activator PA28 rescues the presentation of a cytotoxic T lymphocyte epitope on melanoma cells. Cancer Res. 2002, 62, 2875–2882. [Google Scholar]
- Van Hall, T.; Sijts, A.; Camps, M.; Offringa, R.; Melief, C.; Kloetzel, P.M.; Ossendorp, F. Differential influence on cytotoxic T lymphocyte epitope presentation by controlled expression of either proteasome immunosubunits or PA28. J. Exp. Med. 2000, 192, 483–494. [Google Scholar]
- Murata, S.; Udono, H.; Tanahashi, N.; Hamada, N.; Watanabe, K.; Adachi, K.; Yamano, T.; Yui, K.; Kobayashi, N.; Kasahara, M.; et al. Immunoproteasome assembly and antigen presentation in mice lacking both PA28alpha and PA28beta. EMBO J. 2001, 20, 5898–5907. [Google Scholar] [CrossRef]
- De Graaf, N.; van Helden, M.J.; Textoris-Taube, K.; Chiba, T.; Topham, D.J.; Kloetzel, P.M.; Zaiss, D.M.; Sijts, A.J. PA28 and the proteasome immunosubunits play a central and independentrole in the production of MHC class I-binding peptides in vivo. Eur. J. Immunol. 2011, 41, 926–935. [Google Scholar] [CrossRef]
- Yamano, T.; Sugahara, H.; Mizukami, S.; Murata, S.; Chiba, T.; Tanaka, K.; Yui, K.; Udono, H. Allele-selective effect of PA28 in MHC class I antigen processing. J. Immunol. 2008, 181, 1655–1664. [Google Scholar]
- Gomes, A.V.; Zong, C.; Edmondson, R.D.; Li, X.; Stefani, E.; Zhang, J.; Jones, R.C.; Thyparambil, S.; Wang, G.W.; Qiao, X.; et al. Mapping the murine cardiac 26S proteasome complexes. Circ. Res. 2006, 99, 362–371. [Google Scholar] [CrossRef]
- Husom, A.D.; Peters, E.A.; Kolling, E.A.; Fugere, N.A.; Thompson, L.V.; Ferrington, D.A. Altered proteasome function and subunit composition in aged muscle. Arch. Biochem. Biophys. 2004, 421, 67–76. [Google Scholar] [CrossRef]
- Powell, S.R.; Samuel, S.M.; Wang, P.; Divald, A.; Thirunavukkarasu, M.; Koneru, S.; Wang, X.; Maulik, N. Upregulation of myocardial 11S-activated proteasome in experimental hyperglycemia. J. Mol. Cell. Cardiol. 2008, 44, 618–621. [Google Scholar] [CrossRef]
- Yang, Y.; Fruh, K.; Ahn, K.; Peterson, P.A. In vivo assembly of the proteasomal complexes, implications for antigen processing. J. Biol. Chem. 1995, 270, 27687–27694. [Google Scholar]
- Bose, S.; Brooks, P.; Mason, G.G.; Rivett, A.J. gamma-Interferon decreases the level of 26S proteasomes and changes the pattern of phosphorylation. Biochem. J. 2001, 353, 291–297. [Google Scholar] [CrossRef]
- Barton, L.F.; Cruz, M.; Rangwala, R.; Deepe, G.S., Jr.; Monaco, J.J. Regulation of immunoproteasome subunit expression in vivo following pathogenic fungal infection. J. Immunol. 2002, 169, 3046–3052. [Google Scholar] [CrossRef]
- Khan, S.; van den Broek, M.; Schwarz, K.; de Giuli, R.; Diener, P.A.; Groettrup, M. Immunoproteasomes largely replace constitutive proteasomes during an antiviral and antibacterial immune response in the liver. J. Immunol. 2001, 167, 6859–6868. [Google Scholar] [CrossRef]
- Kuehn, L.; Dahlmann, B. Proteasome activator PA28 and its interaction with 20S proteasomes. Arch. Biochem. Biophys. 1996, 329, 87–96. [Google Scholar] [CrossRef]
- Soza, A.; Knuehl, C.; Groettrup, M.; Henklein, P.; Tanaka, K.; Kloetzel, P.M. Expression and subcellular localization of mouse 20S proteasome activator complex PA28. FEBS Lett. 1997, 413, 27–34. [Google Scholar]
- Song, X.; von Kampen, J.; Slaughter, C.A.; DeMartino, G.N. Relative functions of the alpha and beta subunits of the proteasome activator, PA28. J. Biol. Chem. 1997, 272, 27994–28000. [Google Scholar]
- Wilk, S.; Chen, W.E.; Magnusson, R.P. Properties of the beta subunit of the proteasome activator PA28 (11S REG). Arch. Biochem. Biophys. 2000, 384, 174–180. [Google Scholar] [CrossRef]
- Li, J.; Rechsteiner, M. Molecular dissection of the 11S REG (PA28) proteasome activators. Biochimie 2001, 83, 373–383. [Google Scholar] [CrossRef]
- Kohler, A.; Cascio, P.; Leggett, D.S.; Woo, K.M.; Goldberg, A.L.; Finley, D. The axial channel of the proteasome core particle is gated by the Rpt2 ATPase and controls both substrate entry and product release. Mol. Cell 2001, 7, 1143–1152. [Google Scholar] [CrossRef]
- Whitby, F.G.; Masters, E.I.; Kramer, L.; Knowlton, J.R.; Yao, Y.; Wang, C.C.; Hill, C.P. Structural basis for the activation of 20S proteasomes by 11S regulators. Nature 2000, 408, 115–120. [Google Scholar]
- Harris, J.L.; Alper, P.B.; Li, J.; Rechsteiner, M.; Backes, B.J. Substrate specificity of the human proteasome. Chem. Biol. 2001, 8, 1131–1141. [Google Scholar] [CrossRef]
- Kisselev, A.F.; Akopian, T.N.; Goldberg, A.L. Range of sizes of peptide products generated during degradation of different proteins by archaeal proteasomes. J. Biol. Chem. 1998, 273, 1982–1989. [Google Scholar] [CrossRef]
- Kleijnen, M.F.; Roelofs, J.; Park, S.; Hathaway, N.A.; Glickman, M.; King, R.W.; Finley, D. Stability of the proteasome can be regulated allosterically through engagement of its proteolytic active sites. Nat. Struct. Mol. Biol. 2007, 14, 1180–1188. [Google Scholar] [CrossRef]
- Li, J.; Gao, X.; Ortega, J.; Nazif, T.; Joss, L.; Bogyo, M.; Steven, A.C.; Rechsteiner, M. Lysine 188 substitutions convert the pattern of proteasome activation by REGgamma to that of REGs alpha and beta. EMBO J. 2001, 20, 3359–3369. [Google Scholar] [CrossRef]
- Osmulski, P.A.; Gaczynska, M. Atomic force microscopy reveals two conformations of the 20S proteasome from fission yeast. J. Biol. Chem. 2000, 275, 13171–13174. [Google Scholar] [CrossRef]
- Osmulski, P.A.; Gaczynska, M. Nanoenzymology of the 20S proteasome: Proteasomal actions are controlled by the allosteric transition. Biochemistry 2002, 41, 7047–7053. [Google Scholar]
- Osmulski, P.A.; Hochstrasser, M.; Gaczynska, M. A tetrahedral transition state at the active sites of the 20S proteasome is coupled to opening of the alpha-ring channel. Structure 2009, 17, 1137–1147. [Google Scholar] [CrossRef]
- Ruschak, A.M.; Kay, L.E. Proteasome allostery as a population shift between interchanging conformers. Proc. Natl. Acad. Sci. USA 2012, 109, E3454–E3462. [Google Scholar] [CrossRef]
- Da Fonseca, P.C.; He, J.; Morris, E.P. Molecular model of the human 26S proteasome. Mol. Cell 2012, 46, 54–66. [Google Scholar] [CrossRef]
- Da Fonseca, P.C.; Morris, E.P. Structure of the human 26S proteasome: Subunit radial displacements open the gate into the proteolytic core. J. Biol. Chem. 2008, 283, 23305–23314. [Google Scholar] [CrossRef]
- Dick, T.P.; Ruppert, T.; Groettrup, M.; Kloetzel, P.M.; Kuehn, L.; Koszinowski, U.H.; Stevanovic, S.; Schild, H.; Rammensee, H.G. Coordinated dual cleavages induced by the proteasome regulator PA28 lead to dominant MHC ligands. Cell 1996, 86, 253–262. [Google Scholar] [CrossRef]
- Shimbara, N.; Nakajima, H.; Tanahashi, N.; Ogawa, K.; Niwa, S.; Uenaka, A.; Nakayama, E.; Tanaka, K. Double-cleavage production of the CTL epitope by proteasomes and PA28: Role of the flanking region. Genes Cells 1997, 2, 785–800. [Google Scholar] [CrossRef]
- Cascio, P.; Hilton, C.; Kisselev, A.F.; Rock, K.L.; Goldberg, A.L. 26S proteasomes and immunoproteasomes produce mainly N-extended versions of an antigenic peptide. EMBO J. 2001, 20, 2357–2366. [Google Scholar] [CrossRef]
- Mo, X.Y.; Cascio, P.; Lemerise, K.; Goldberg, A.L.; Rock, K. Distinct proteolytic processes generate the C and N termini of MHC class I-binding peptides. J. Immunol. 1999, 163, 5851–5859. [Google Scholar]
- Saric, T.; Chang, S.C.; Hattori, A.; York, I.A.; Markant, S.; Rock, K.L.; Tsujimoto, M.; Goldberg, A.L. An IFN-gamma-induced aminopeptidase in the ER, ERAP1, trims precursors to MHC class I-presented peptides. Nat. Immunol. 2002, 3, 1169–1176. [Google Scholar] [CrossRef]
- Serwold, T.; Gonzalez, F.; Kim, J.; Jacob, R.; Shastri, N. ERAAP customizes peptides for MHC class I molecules in the endoplasmic reticulum. Nature 2002, 419, 480–483. [Google Scholar] [CrossRef]
- Snyder, H.L.; Yewdell, J.W.; Bennink, J.R. Trimming of antigenic peptides in an early secretory compartment. J. Exp. Med. 1994, 180, 2389–2394. [Google Scholar] [CrossRef]
- Stoltze, L.; Schirle, M.; Schwarz, G.; Schroter, C.; Thompson, M.W.; Hersh, L.B.; Kalbacher, H.; Stevanovic, S.; Rammensee, H.G.; Schild, H. Two new proteases in the MHC class I processing pathway. Nat. Immunol. 2000, 1, 413–418. [Google Scholar] [CrossRef]
- York, I.A.; Mo, A.X.; Lemerise, K.; Zeng, W.; Shen, Y.; Abraham, C.R.; Saric, T.; Goldberg, A.L.; Rock, K.L. The cytosolic endopeptidase, thimet oligopeptidase, destroys antigenic peptides and limits the extent of MHC class I antigen presentation. Immunity 2003, 18, 429–440. [Google Scholar] [CrossRef]
- Beninga, J.; Rock, K.L.; Goldberg, A.L. Interferon-gamma can stimulate post-proteasomal trimming of the N terminus of an antigenic peptide by inducing leucine aminopeptidase. J. Biol. Chem. 1998, 273, 18734–18742. [Google Scholar] [CrossRef]
- Preckel, T.; Fung-Leung, W.P.; Cai, Z.; Vitiello, A.; Salter-Cid, L.; Winqvist, O.; Wolfe, T.G.; von Herrath, M.; Angulo, A.; Ghazal, P.; et al. Impaired immunoproteasome assembly and immune responses in PA28−/− mice. Science 1999, 286, 2162–2165. [Google Scholar] [CrossRef]
- Schwarz, K.; Eggers, M.; Soza, A.; Koszinowski, U.H.; Kloetzel, P.M.; Groettrup, M. The proteasome regulator PA28alpha/beta can enhance antigen presentation without affecting 20S proteasome subunit composition. Eur. J. Immunol. 2000, 30, 3672–3679. [Google Scholar] [CrossRef]
- Groll, M.; Bajorek, M.; Kohler, A.; Moroder, L.; Rubin, D.M.; Huber, R.; Glickman, M.H.; Finley, D. A gated channel into the proteasome core particle. Nat. Struct. Biol. 2000, 7, 1062–1067. [Google Scholar] [CrossRef]
- Forster, A.; Masters, E.I.; Whitby, F.G.; Robinson, H.; Hill, C.P. The 1.9 Å structure of a proteasome-11S activator complex and implications for proteasome-PAN/PA700 interactions. Mol. Cell 2005, 18, 589–599. [Google Scholar] [CrossRef]
- Forster, A.; Whitby, F.G.; Hill, C.P. The pore of activated 20S proteasomes has an ordered 7-fold symmetric conformation. EMBO J. 2003, 22, 4356–4364. [Google Scholar] [CrossRef]
- Stadtmueller, B.M.; Hill, C.P. Proteasome activators. Mol. Cell 2011, 41, 8–19. [Google Scholar] [CrossRef]
- Berko, D.; Tabachnick-Cherny, S.; Shental-Bechor, D.; Cascio, P.; Mioletti, S.; Levy, Y.; Admon, A.; Ziv, T.; Tirosh, B.; Goldberg, A.L.; et al. The direction of protein entry into the proteasome determines the variety of products and depends on the force needed to unfold its two termini. Mol. Cell 2012, 48, 601–611. [Google Scholar] [CrossRef]
- Kisselev, A.F.; Akopian, T.N.; Woo, K.M.; Goldberg, A.L. The sizes of peptides generated from protein by mammalian 26 and 20S proteasomes. Implications for understanding the degradative mechanism and antigen presentation. J. Biol. Chem. 1999, 274, 3363–3371. [Google Scholar]
- Nussbaum, A.K.; Dick, T.P.; Keilholz, W.; Schirle, M.; Stevanovic, S.; Dietz, K.; Heinemeyer, W.; Groll, M.; Wolf, D.H.; Huber, R.; et al. Cleavage motifs of the yeast 20S proteasome beta subunits deduced from digests of Enolase 1. Proc. Natl. Acad. Sci. USA 1998, 95, 12504–12509. [Google Scholar] [CrossRef]
- Raule, M.; Cerruti, F.; Benaroudj, N.; Migotti, R.; Kikuchi, J.; Bachi, A.; Navon, A.; Dittmar, G.; Cascio, P. PA28alphabeta reduces size and increases hydrophilicity of 20S immunoproteasome peptide products. Chem. Biol. 2014, 21, 470–480. [Google Scholar] [CrossRef]
- Mo, A.X.; van Lelyveld, S.F.; Craiu, A.; Rock, K.L. Sequences that flank subdominant and cryptic epitopes influence the proteolytic generation of MHC class I-presented peptides. J. Immunol. 2000, 164, 4003–4010. [Google Scholar] [CrossRef]
- Craiu, A.; Akopian, T.; Goldberg, A.; Rock, K.L. Two distinct proteolytic processes in the generation of a major histocompatibility complex class I-presented peptide. Proc. Natl. Acad. Sci. USA 1997, 94, 10850–10855. [Google Scholar] [CrossRef]
- Gillette, T.G.; Kumar, B.; Thompson, D.; Slaughter, C.A.; DeMartino, G.N. Differential roles of the COOH termini of AAA subunits of PA700 (19S regulator) in asymmetric assembly and activation of the 26S proteasome. J. Biol. Chem. 2008, 283, 31813–31822. [Google Scholar]
- Rabl, J.; Smith, D.M.; Yu, Y.; Chang, S.C.; Goldberg, A.L.; Cheng, Y. Mechanism of gate opening in the 20S proteasome by the proteasomal ATPases. Mol. Cell 2008, 30, 360–368. [Google Scholar] [CrossRef]
- Bajorek, M.; Finley, D.; Glickman, M.H. Proteasome disassembly and downregulation is correlated with viability during stationary phase. Curr. Biol. 2003, 13, 1140–1144. [Google Scholar] [CrossRef]
- Benaroudj, N.; Zwickl, P.; Seemuller, E.; Baumeister, W.; Goldberg, A.L. ATP hydrolysis by the proteasome regulatory complex PAN serves multiple functions in protein degradation. Mol. Cell 2003, 11, 69–78. [Google Scholar] [CrossRef]
- Matouschek, A. Protein unfolding—An important process in vivo? Curr. Opin. Struct. Biol. 2003, 13, 98–109. [Google Scholar] [CrossRef]
- Smith, D.M.; Kafri, G.; Cheng, Y.; Ng, D.; Walz, T.; Goldberg, A.L. ATP binding to PAN or the 26S ATPases causes association with the 20S proteasome, gate opening, and translocation of unfolded proteins. Mol. Cell 2005, 20, 687–698. [Google Scholar] [CrossRef]
- Chen, X.; Barton, L.F.; Chi, Y.; Clurman, B.E.; Roberts, J.M. Ubiquitin-independent degradation of cell-cycle inhibitors by the REGgamma proteasome. Mol. Cell 2007, 26, 843–852. [Google Scholar] [CrossRef]
- Li, X.; Amazit, L.; Long, W.; Lonard, D.M.; Monaco, J.J.; O’Malley, B.W. Ubiquitin- and ATP-independent proteolytic turnover of p21 by the REGgamma-proteasome pathway. Mol. Cell 2007, 26, 831–842. [Google Scholar] [CrossRef]
- Li, X.; Lonard, D.M.; Jung, S.Y.; Malovannaya, A.; Feng, Q.; Qin, J.; Tsai, S.Y.; Tsai, M.J.; O’Malley, B.W. The SRC-3/AIB1 coactivator is degraded in a ubiquitin- and ATP-independent manner by the REGgamma proteasome. Cell 2006, 124, 381–392. [Google Scholar] [CrossRef]
- Moriishi, K.; Okabayashi, T.; Nakai, K.; Moriya, K.; Koike, K.; Murata, S.; Chiba, T.; Tanaka, K.; Suzuki, R.; Suzuki, T.; et al. Proteasome activator PA28gamma-dependent nuclear retention and degradation of hepatitis C virus core protein. J. Virol. 2003, 77, 10237–10249. [Google Scholar]
- Benaroudj, N.; Tarcsa, E.; Cascio, P.; Goldberg, A.L. The unfolding of substrates and ubiquitin-independent protein degradation by proteasomes. Biochimie 2001, 83, 311–318. [Google Scholar] [CrossRef]
- Mishto, M.; Luciani, F.; Holzhutter, H.G.; Bellavista, E.; Santoro, A.; Textoris-Taube, K.; Franceschi, C.; Kloetzel, P.M.; Zaikin, A. Modeling the in vitro 20S proteasome activity: The effect of PA28-alphabeta and of the sequence and length of polypeptides on the degradation kinetics. J. Mol. Biol. 2008, 377, 1607–1617. [Google Scholar] [CrossRef]
- Saric, T.; Beninga, J.; Graef, C.I.; Akopian, T.N.; Rock, K.L.; Goldberg, A.L. Major histocompatibility complex class I-presented antigenic peptides are degraded in cytosolic extracts primarily by thimet oligopeptidase. J. Biol. Chem. 2001, 276, 36474–36481. [Google Scholar]
- Saric, T.; Graef, C.I.; Goldberg, A.L. Pathway for degradation of peptides generated by proteasomes: A key role for thimet oligopeptidase and other metallopeptidases. J. Biol. Chem. 2004, 279, 46723–46732. [Google Scholar] [CrossRef]
- Dolenc, I.; Seemuller, E.; Baumeister, W. Decelerated degradation of short peptides by the 20S proteasome. FEBS Lett. 1998, 434, 357–361. [Google Scholar] [CrossRef]
- Shabek, N.; Ciechanover, A. Degradation of ubiquitin: The fate of the cellular reaper. Cell Cycle 2010, 9, 523–530. [Google Scholar] [CrossRef]
- Verma, R.; Aravind, L.; Oania, R.; McDonald, W.H.; Yates, J.R., 3rd; Koonin, E.V.; Deshaies, R.J. Role of Rpn11 metalloprotease in deubiquitination and degradation by the 26S proteasome. Science 2002, 298, 611–615. [Google Scholar] [CrossRef]
- Yao, T.; Cohen, R.E. A cryptic protease couples deubiquitination and degradation by the proteasome. Nature 2002, 419, 403–407. [Google Scholar]
- Qian, S.B.; Reits, E.; Neefjes, J.; Deslich, J.M.; Bennink, J.R.; Yewdell, J.W. Tight linkage between translation and MHC class I peptide ligand generation implies specialized antigen processing for defective ribosomal products. J. Immunol. 2006, 177, 227–233. [Google Scholar] [CrossRef]
- Reits, E.A.; Vos, J.C.; Gromme, M.; Neefjes, J. The major substrates for TAP in vivo are derived from newly synthesized proteins. Nature 2000, 404, 774–778. [Google Scholar] [CrossRef]
- Schubert, U.; Anton, L.C.; Gibbs, J.; Norbury, C.C.; Yewdell, J.W.; Bennink, J.R. Rapid degradation of a large fraction of newly synthesized proteins by proteasomes. Nature 2000, 404, 770–774. [Google Scholar] [CrossRef]
- Anton, L.C.; Schubert, U.; Bacik, I.; Princiotta, M.F.; Wearsch, P.A.; Gibbs, J.; Day, P.M.; Realini, C.; Rechsteiner, M.C.; Bennink, J.R.; et al. Intracellular localization of proteasomal degradation of a viral antigen. J. Cell Biol. 1999, 146, 113–124. [Google Scholar] [CrossRef]
- Qian, S.B.; Princiotta, M.F.; Bennink, J.R.; Yewdell, J.W. Characterization of rapidly degraded polypeptides in mammalian cells reveals a novel layer of nascent protein quality control. J. Biol. Chem. 2006, 281, 392–400. [Google Scholar]
- Asher, G.; Tsvetkov, P.; Kahana, C.; Shaul, Y. A mechanism of ubiquitin-independent proteasomal degradation of the tumor suppressors p53 and p73. Genes Dev. 2005, 19, 316–321. [Google Scholar] [CrossRef]
- Davies, K.J. Degradation of oxidized proteins by the 20S proteasome. Biochimie 2001, 83, 301–310. [Google Scholar] [CrossRef]
- Moorthy, A.K.; Savinova, O.V.; Ho, J.Q.; Wang, V.Y.; Vu, D.; Ghosh, G. The 20S proteasome processes NF-kappaB1 p105 into p50 in a translation-independent manner. EMBO J. 2006, 25, 1945–1956. [Google Scholar] [CrossRef]
- Orlowski, M.; Wilk, S. Ubiquitin-independent proteolytic functions of the proteasome. Arch. Biochem. Biophys. 2003, 415, 1–5. [Google Scholar] [CrossRef]
- Shringarpure, R.; Grune, T.; Mehlhase, J.; Davies, K.J. Ubiquitin conjugation is not required for the degradation of oxidized proteins by proteasome. J. Biol. Chem. 2003, 278, 311–318. [Google Scholar]
- Yuksek, K.; Chen, W.L.; Chien, D.; Ou, J.H. Ubiquitin-independent degradation of hepatitis C virus F protein. J. Virol. 2009, 83, 612–621. [Google Scholar] [CrossRef]
- Whittier, J.E.; Xiong, Y.; Rechsteiner, M.C.; Squier, T.C. Hsp90 enhances degradation of oxidized calmodulin by the 20S proteasome. J. Biol. Chem. 2004, 279, 46135–46142. [Google Scholar]
- Seifert, U.; Bialy, L.P.; Ebstein, F.; Bech-Otschir, D.; Voigt, A.; Schroter, F.; Prozorovski, T.; Lange, N.; Steffen, J.; Rieger, M.; et al. Immunoproteasomes preserve protein homeostasis upon interferon-induced oxidative stress. Cell 2010, 142, 613–624. [Google Scholar] [CrossRef]
- Li, J.; Powell, S.R.; Wang, X. Enhancement of proteasome function by PA28α overexpression protects against oxidative stress. FASEB J. 2011, 25, 883–893. [Google Scholar] [CrossRef]
- Pickering, A.M.; Davies, K.J. Differential roles of proteasome and immunoproteasome regulators Pa28alphabeta, Pa28gamma and Pa200 in the degradation of oxidized proteins. Arch. Biochem. Biophys. 2012, 523, 181–190. [Google Scholar] [CrossRef]
- Pickering, A.M.; Koop, A.L.; Teoh, C.Y.; Ermak, G.; Grune, T.; Davies, K.J. The immunoproteasome, the 20S proteasome and the PA28alphabeta proteasome regulator are oxidative-stress-adaptive proteolytic complexes. Biochem. J. 2010, 432, 585–594. [Google Scholar] [CrossRef]
- Klaunig, J.E.; Kamendulis, L.M. The role of oxidative stress in carcinogenesis. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 239–267. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cascio, P. PA28αβ: The Enigmatic Magic Ring of the Proteasome? Biomolecules 2014, 4, 566-584. https://doi.org/10.3390/biom4020566
Cascio P. PA28αβ: The Enigmatic Magic Ring of the Proteasome? Biomolecules. 2014; 4(2):566-584. https://doi.org/10.3390/biom4020566
Chicago/Turabian StyleCascio, Paolo. 2014. "PA28αβ: The Enigmatic Magic Ring of the Proteasome?" Biomolecules 4, no. 2: 566-584. https://doi.org/10.3390/biom4020566
APA StyleCascio, P. (2014). PA28αβ: The Enigmatic Magic Ring of the Proteasome? Biomolecules, 4(2), 566-584. https://doi.org/10.3390/biom4020566