The Roles of Sphingosine Kinase 1 and 2 in Regulating the Metabolome and Survival of Prostate Cancer Cells

 ,
,  ,
,
Abstract
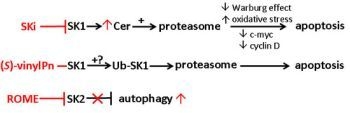
:
1. Introduction
2. Results and Discussion
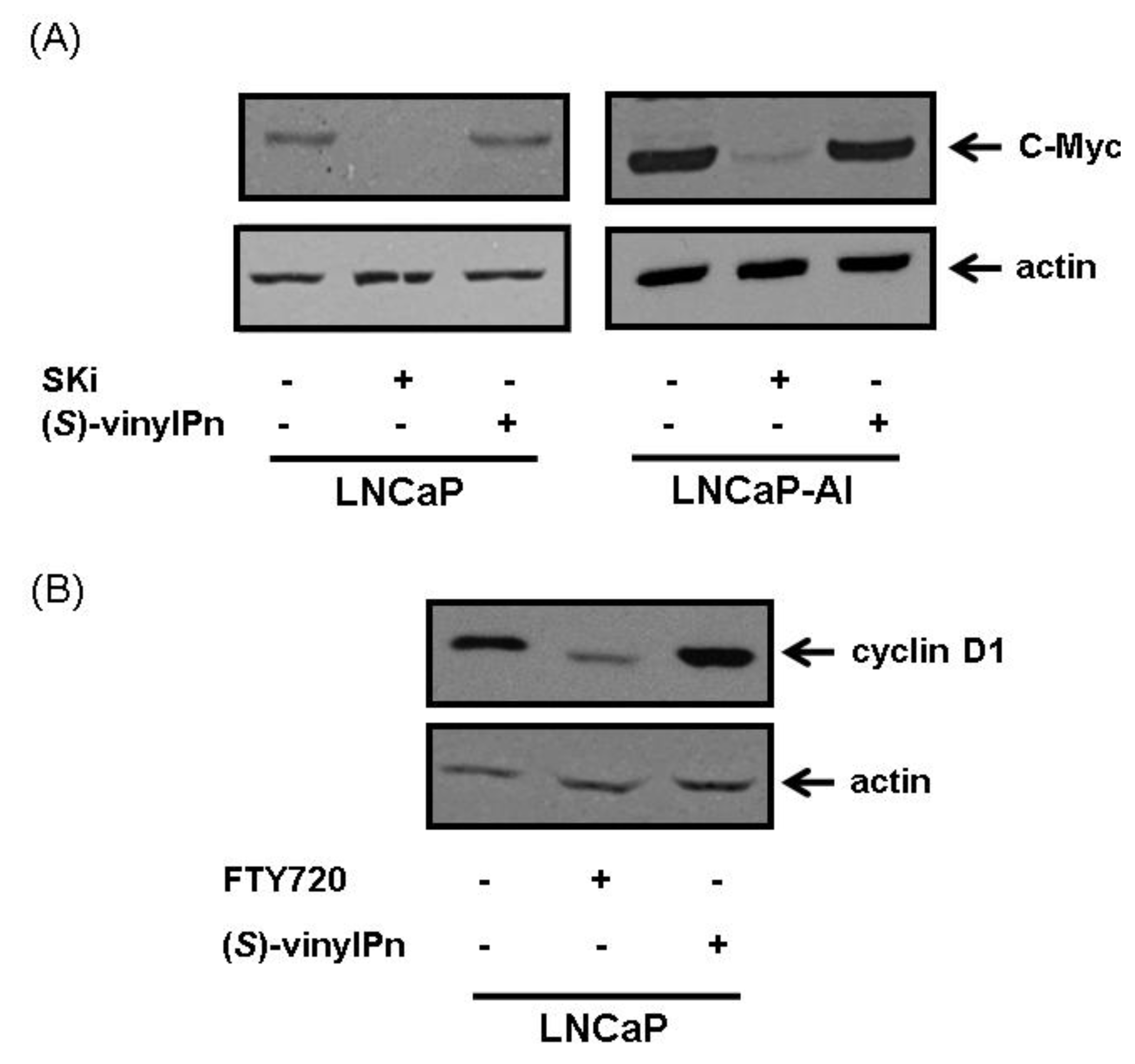
2.1. Effect of SKi on the Metabolome of LNCaP-AI Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ratio of SKi : control | P value n = 6 | |
|---|---|---|
| Oxidative Stress | ||
| GSSG | 12.4 | <0.001 |
| NADP+ | 5.9 | <0.001 |
| NADPH | 0.24 | <0.0001 |
| Phosphogluconate | 4.4 | <0.05 |
| Ribulose phosphate | 3.7 | <0.01 |
| Glycolysis/TCA cycle | ||
| Dihydroxyacetone phosphate | 7.6 | <0.005 |
| D-Glyceraldehyde 3-phosphate | 23.4 | <0.01 |
| 3-Phosphoglycerate | 2.7 | <0.05 |
| Fructose 1,6-bisphosphate | 5.2 | <0.0001 |
| S-D-Lactoylglutathione | 28.3 | <0.001 |
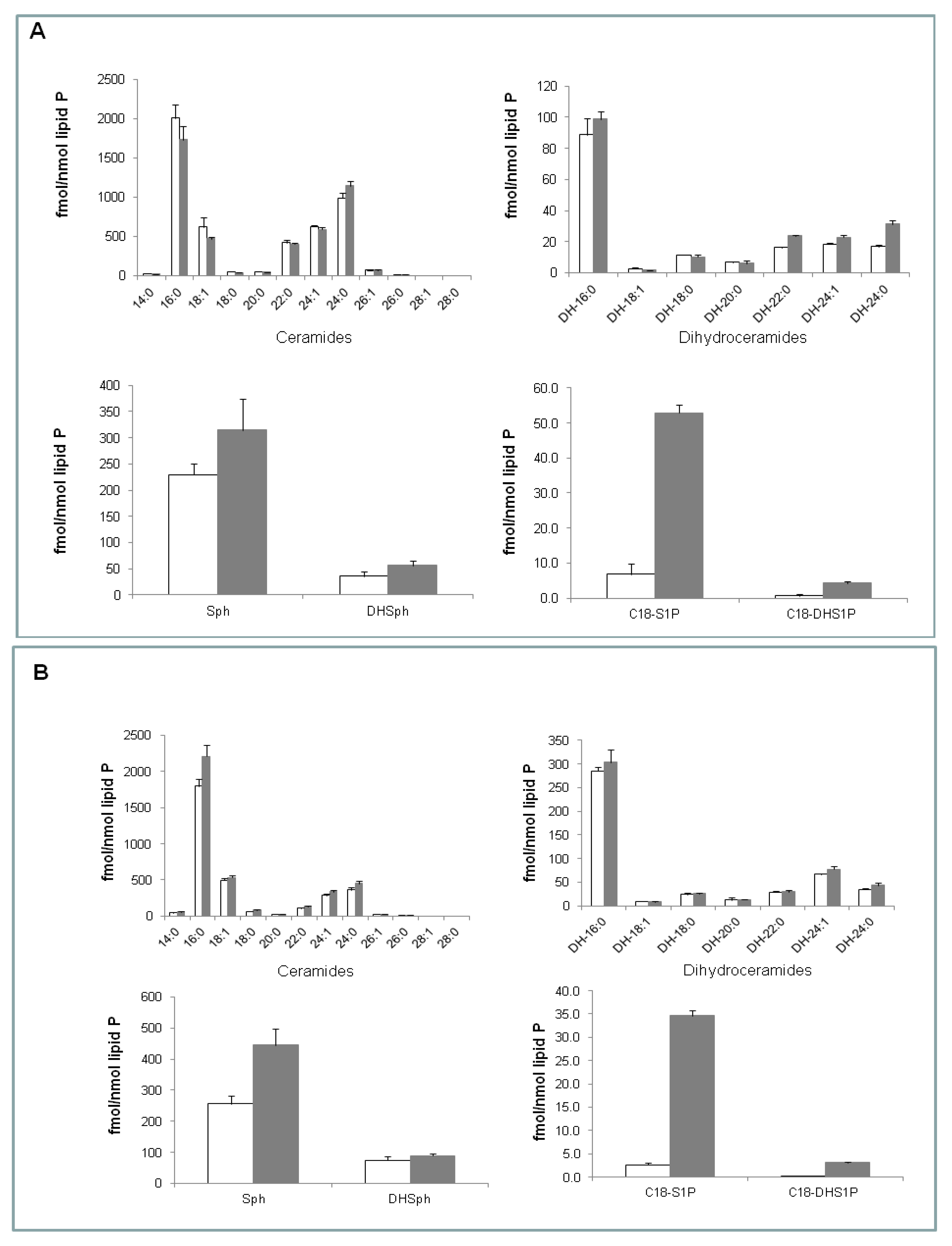
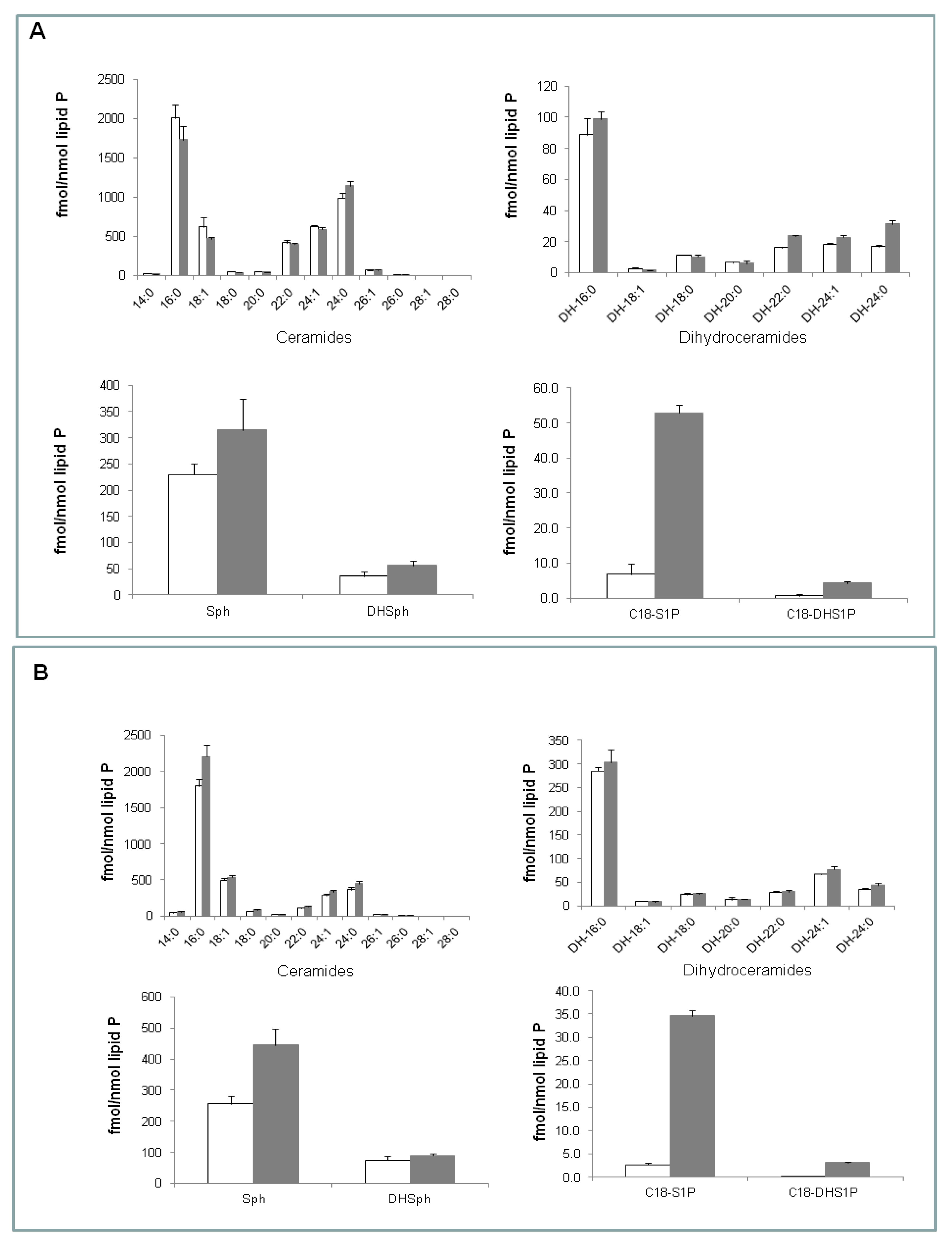
2.2. Effect of (S)-FTY720 Vinylphosphonate on Sphingolipids in LNCaP and LNCaP-AI Cells
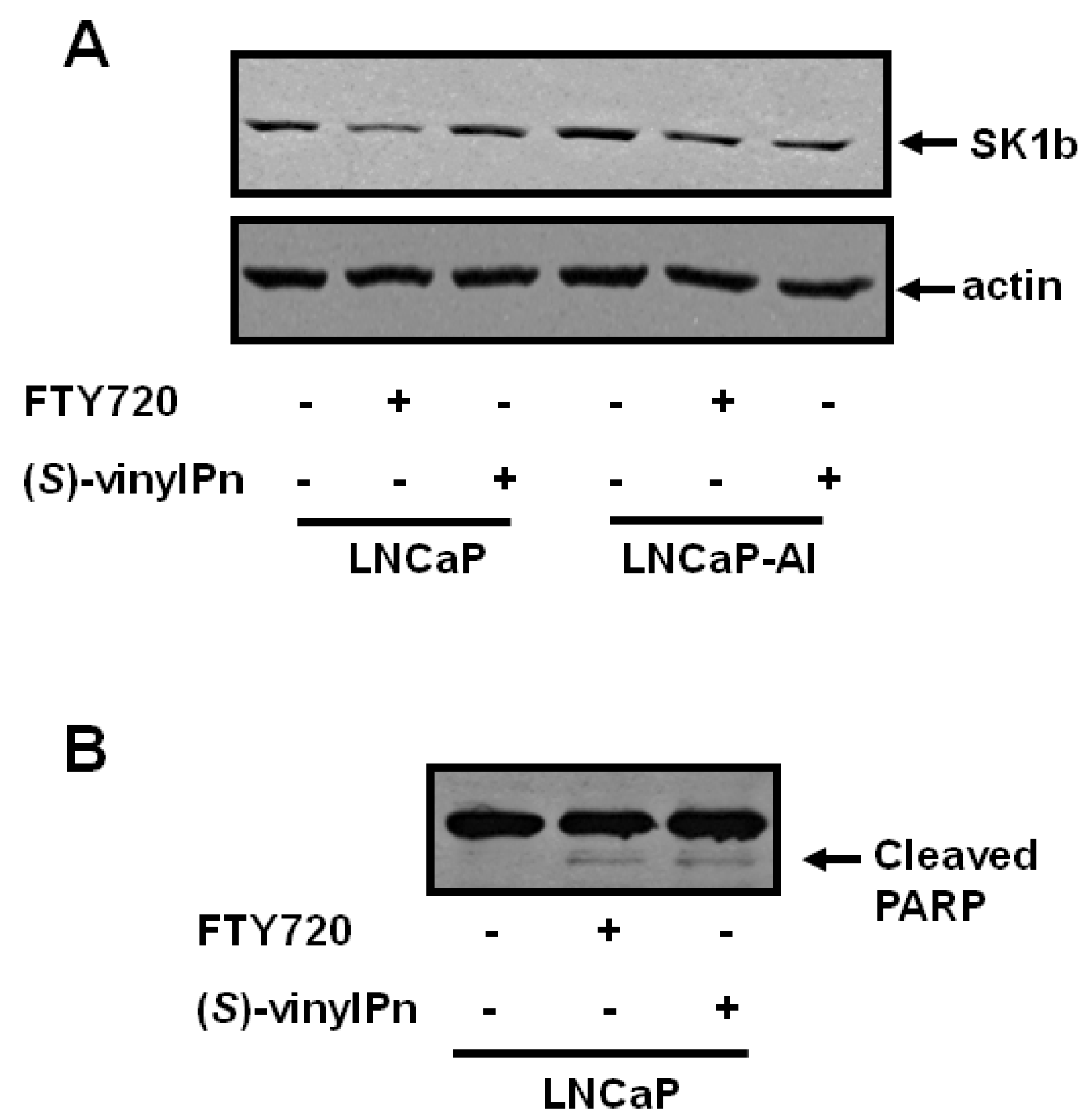
2.3. Differential Effect of (R)- and (S)-FTY720 Vinylphosphonate on S1P Levels in Human Lung Endothelial Cells
| (A) | [32P] S1P formed (dpm) |
| Vehicle | 878 +/− 6 |
| Sphingosine | 4025 +/− 47 |
| (S)-vinylPn + Vehicle | 1124 +/− 58 |
| (S)-vinylPn + Sphingosine | 5150 +/− 18 |
| (R)-vinylPn + Vehicle | 640 +/− 40 |
| (R)-vinylPn + Sphingosine | 2326 +/− 32 |
| (B) | [32P] S1P formed (dpm) |
| Vehicle | 1015 +/− 69 |
| Sphingosine | 3665 +/− 264 |
| (S)-vinylPn + Vehicle | 1011 +/− 35 |
| (S)-vinylPn + Sphingosine | 5530 +/− 358 |
| (R)-vinylPn + Vehicle | 724 +/− 84 |
| (R)-vinylPn + Sphingosine | 2627 +/− 178 |
2.4. Effect of (S)-FTY720 Vinylphosphonate on the Metabolome of LNCaP and LNCaP-AI Cells
| Compound | Ratio (S)-vinylPn- treated/controlLNCaP-AI | P value | Ratio (S)-vinylPn- treated/controlLNCaP | P value |
|---|---|---|---|---|
| Lipids | ||||
| PA 32:1 | 2.39 | < 0.01 | 1.35 | 0.36 |
| PA 32:2 | 2.50 | < 0.001 | 2.07 | < 0.05 |
| PA 34:2 | 2.33 | < 0.001 | 1.54 | 0.11 |
| PS 32:1 | 2.04 | < 0.02 | ||
| PS 38:6 | 2.16 | < 0.01 | ND | |
| Lyso-PE 18:1 | 2.18 | < 0.01 | 0.92 | 0.61 |
| Lyso-PE 20:1 | 2.10 | < 0.01 | 1.02 | 0.85 |
| Lyso-PE 20:2 | 2.10 | < 0.01 | 0.97 | 0.78 |
| Oxidative stress | ||||
| GSH | 0.98 | 0.76 | 0.75 | < 0.01 |
| S-D-Lactoylglutathione | 0.97 | 0.83 | 0.61 | < 0.01 |
| GSSG | 0.88 | 0.39 | 0.58 | 0.06 |
| Compound | Ratio ROME- treated/control LNCaP | P value |
|---|---|---|
| Lyso-PE 18:0 | 1.53 | < 0.05 |
| Lyso-PE 18:1 | 1.36 | < 0.05 |
| Lyso-PE 20:1 | 1.61 | < 0.05 |
| Compound | Ratio SKi-treated/control LNCaP | P value |
|---|---|---|
| Lyso-PE 18:0 | 3.0 | < 0.01 |
| Lyso-PE 18:1 | 1.8 | < 0.01 |
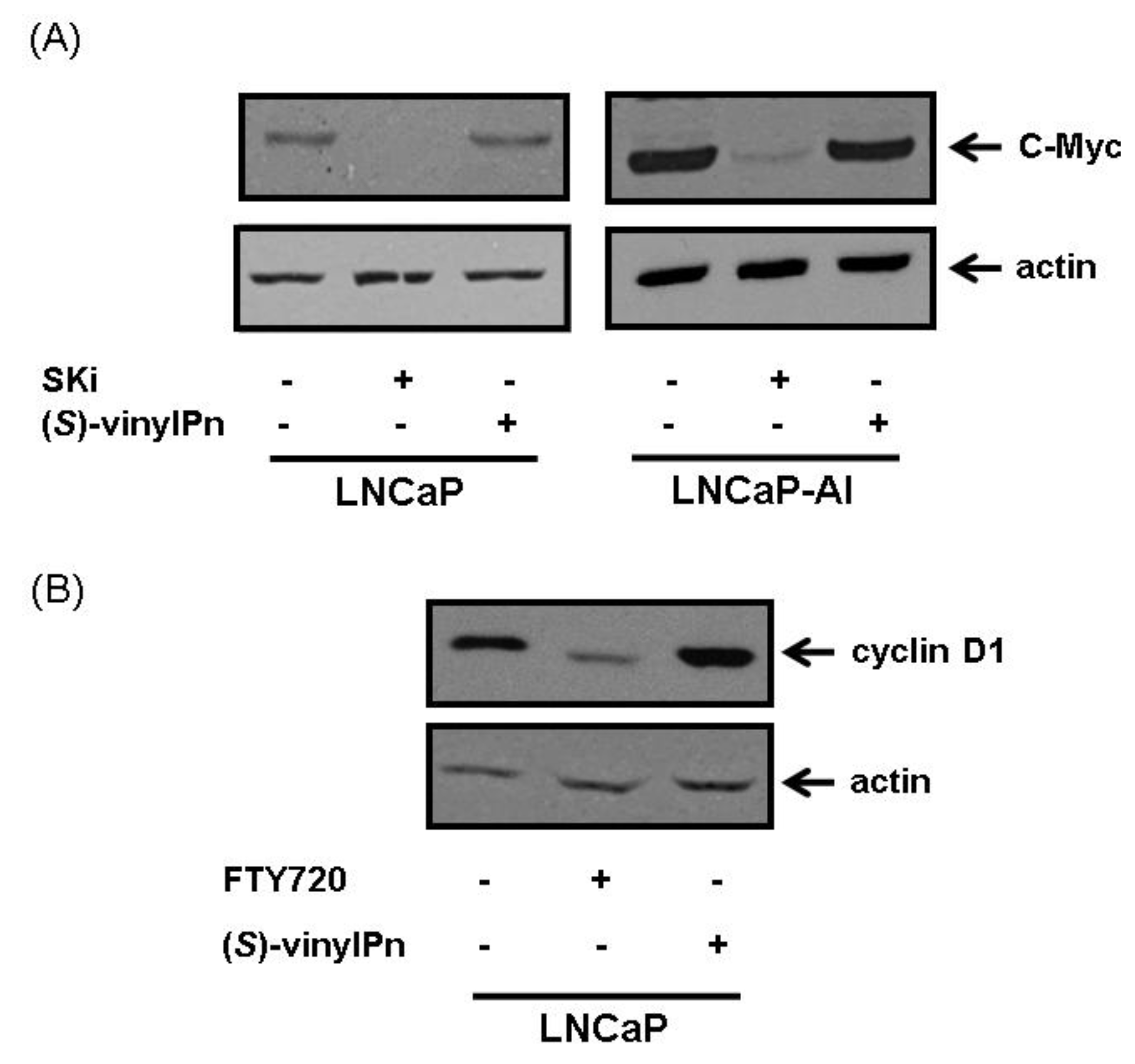
2.5. Effect of ROME on the Metabolome of LNCaP Cells
| Ratio of ROME : control | P value n = 3 | |
|---|---|---|
| Lyso-PI 16:0 | 218.2 | < 0.005 |
| Lyso-PI 18:0 | 30.0 | < 0.005 |
| Lyso-PI 18:1 | 22.2 | < 0.005 |
| Palmityl glucuronide | 11.2 | < 0.01 |
| LPA 16:0 | 2.4 | < 0.05 |
| LPA 16:1 | 1.4 | < 0.01 |
| LPA 18:0 | 3.6 | < 0.05 |
3. Experimental Section
3.1. Materials
3.2. Cell Culture
3.3. Preparation of Whole Cell Extracts
3.4. Measurement of Intracellular [32P]S1P Generation
3.5. Analysis of Sphingoid Base 1-Phosphates and Ceramides
3.6. Isolation of Cell Extracts and Liquid Chromatography Mass Spectrometry
3.7. Data Extraction
4. Conclusions
| Compound | SKi | (S)-FTY720 -vinylPn) | ROME |
|---|---|---|---|
| Inhibitory specificity | SK1/2 | SK1 selective | SK2 selective |
| Proteasome | Activator | No effect | No effect |
| SK1 expression | Proteasomal degradation of SK1a/SK1b in LNCaP but only SK1a in LNCaP-AI | Proteasomal degradation of SK1a/SK1b in LNCaP-AI | No effect on SK1 expression |
| Apoptosis | Inducer in LNCaP No effect in LNCaP-AI | Inducer in LNCaP and LNCaP-AI | No effect |
| Authophagy | Inhibition | ND | Stimulation |
| Oxidative stress | Induction | No effect | No effect |
| Glycolysis | Increased glycolytic metabolites; reduced flux? | No effect | Reduced glycolytic metabolites: increased flux? |
| c-Myc expression | Reduced | No effect | No effect |
| Androgen receptor expression | Reduced | Reduced | No effect |
| Lipid metabolism | Lyso-PE modestly increased in LNCaP | Lyso-PE modestly increased in LNCaP-AI | Lyso-PI substantially increased in LNCaP |
Acknowledgments
Conflict of Interest
References
- Pyne, N.J.; Pyne, S. Sphingosine 1-phosphate and cancer. Nat. Rev. Cancer 2010, 10, 489–503. [Google Scholar] [CrossRef]
- Hait, N.C.; Allegood, J.; Maceyka, M.; Strub, G.M.; Harikumar, K.B.; Singh, S.K.; Luo, C.; Marmorstein, R.; Kordula, T.; Milstien, S.; et al. Regulation of histone acetylation in the nucleus by sphingosine-1-phosphate. Science 2009, 325, 1254–1257. [Google Scholar] [CrossRef]
- Alvarez, S.E.; Harikumar, K.B.; Hait, N.C.; Allegood, J.; Strub, G.M.; Kim, E.Y.; Maceyka, M.; Jiang, H.; Luo, C.; Kordula, T.; et al. Sphingosine-1-phosphate is a missing cofactor for the E3 ubiquitin ligase TRAF2. Nature 2010, 465, 1084–1088. [Google Scholar] [CrossRef]
- Strub, G.M.; Paillard, M.; Liang, J.; Gomez, L.; Allegood, J.C.; Hait, N.C.; Maceyka, M.; Price, M.M.; Chen, Q.; Simpson, D.C.; et al. Sphingosine-1 phosphate produced by sphingosine kinase 2 in mitochondria interacts with prohibitin 2 to regulate complex IV assembly and respiration. FASEB J. 2011, 25, 600–612. [Google Scholar] [CrossRef]
- Sankala, H.M.; Hait, N.C.; Paugh, S.W.; Shida, D.; Lépine, S.; Elmore, L.W.; Dent, P.; Milstien, S.; Spiegel, S. Involvement of sphingosine kinase 2 in p53-independent induction of p21 by the chemotherapeutic drug doxorubicin. Cancer Res. 2007, 67, 10466–10474. [Google Scholar] [CrossRef]
- Gao, P.; Smith, C.D. Ablation of sphingosine kinase-2 inhibits tumor cell proliferation and migration. Mol. Cancer Res. 2011, 9, 1509–1519. [Google Scholar] [CrossRef]
- Loveridge, C.; Tonelli, F.; Leclercq, T.; Lim, K.G.; Long, S.; Berdyshev, E.; Tate, R.J.; Natarajan, V.; Pitson, S.M.; Pyne, N.J.; et al. The sphingosine kinase 1 inhibitor 2-(p-hydroxyanilino)-4-(p-chlorophenyl)thiazole induces proteosomal degradation of sphingosine kinase 1 in mammalian cells. J. Biol. Chem. 2010, 285, 38841–38852. [Google Scholar] [CrossRef]
- Tonelli, F.; Lim, K.G.; Loveridge, C.; Long, J.; Pitson, S.M.; Tigyi, G.; Bittman, R.; Pyne, S.; Pyne, N.J. FTY720 and (S)-FTY720 vinylphosphonate inhibit sphingosine kinase 1 and promotes its proteasomal degradation in human pulmonary artery smooth muscle, breast cancer and androgen-independent prostate cancer cells. Cell. Signal. 2010, 22, 1536–1542. [Google Scholar] [CrossRef]
- Lim, K.G.; Tonelli, F.; Berdyshev, E.V.; Gorshkova, I.; Leclercq, T.; Pitson, S.M.; Bittman, R.; Pyne, S.; Pyne, N.J. Inhibition kinetics and regulation of sphingosine kinase 1 expression in prostate cancer cells: Functional differences between sphingosine kinase 1A and 1B. Int. J. Biochem. Cell Biol. 2012, 44, 1457–1464. [Google Scholar] [CrossRef]
- Watson, D.; Tonelli, F.; Al-Osaimi, M.; Williamson, L.; Chan, E.; Gorshkova, I.; Berdyshev, E.; Bittman, R.; Pyne, N.J.; Pyne, S. The role of sphingosine kinase 1 and 2 in regulating the Warburg effect in prostate cancer cells. Cell. Signal. 2013, 25, 1011–1017. [Google Scholar] [CrossRef]
- Lim, K.G.; Tonelli, F.; Li, Z.; Lu, X.; Bittman, R.; Pyne, S.; Pyne, N.J. FTY720 analogues as sphingosine kinase 1 inhibitors: Enzyme inhibition kinetics, allosterism, proteasomal degradation and actin rearrangement in MCF-7 breast cancer cells. J. Biol. Chem. 2011, 286, 18633–18640. [Google Scholar]
- Edwards, L.G.; Adesida, A.; Thornalley, P.J. Inhibition of human leukaemia 60 cell growth by S-D-lactoylglutathione in vitro. Mediation by metabolism to N-D-lactoylcysteine and induction of apoptosis. Leuk. Res. 1996, 20, 17–26. [Google Scholar] [CrossRef]
- Lim, K.G.; Sun, C.; Bittman, R.; Pyne, N.J.; Pyne, S. (R)-FTY720 methyl ether is a specific sphingosine kinase 2 inhibitor: Effect on sphingosine kinase 2 expression in HEK 293 cells and actin rearrangement and survival of MCF-7 breast cancer cells. Cell. Signal. 2011, 23, 1590–1595. [Google Scholar] [CrossRef]
- French, K.J.; Zhuang, Y.; Maines, L.W.; Gao, P.; Wang, W.; Beljanski, V.; Upson, J.J.; Green, C.L.; Keller, S.N.; Smith, C.D. Pharmacology and antitumor activity of ABC294640, a selective inhibitor of sphingosine kinase-2. J. Pharmacol. Exp. Ther. 2010, 333, 129–139. [Google Scholar] [CrossRef]
- Tonelli, F.; Osaimi, M.A.; Williamson, L.; Tate, R.J.; Watson, D.G.; Chan, E.; Bittman, R.; Pyne, N.J.; Pyne, S. The sphingosine kinase inhibitor 2-(p-hydroxyanilino)-4-(p-chlorophenyl)thiazole reduces androgen receptor expression via an oxidative stress-dependent mechanism. Br. J. Pharmacol. 2013, 168, 1497–1505. [Google Scholar] [CrossRef]
- Griffin, J.L.; Nicholls, A.W.; Daykin, C.A.; Heald, S.; Keun, H.C.; Schuppe-Koistinen, I.; Griffiths, J.R.; Cheng, L.L.; Rocca-Serra, P.; Rubtsov, D.V.; Robertson, D. Standard reporting requirements for biological samples in metabolomics experiments: Mammalian/in vivo experiments. Metabolomics 2007, 3, 179–188. [Google Scholar] [CrossRef]
- Ji, L.; Fang, B.; Yen, N.; Fong, K.; Minna, J.D.; Roth, J.A. Induction of apoptosis and inhibition of tumorigenicity and tumor growth by adenovirus vector-mediated Fragile Histidine Triad (FHIT) gene overexpression. Cancer Res. 1999, 59, 3333–3339. [Google Scholar]
- Pichiorri, F.; Palumbo, T.; Suh, S-S.; Okamura, H.; Trepasso, F.; Ishii, H.; Huebner, K.; Coce, C.M. Fhit tumor suppressor: Guardian of the preneoplastic genome. Fut. Oncol. 2008, 4, 815–824. [Google Scholar]
- Deng, W.G.; Nishizaki, M.; Fang, B.; Roth, J.A.; Ji, L. Induction of apoptosis by tumor suppressor FHIT via death receptor signaling pathway in human lung cancer cells. Biochem. Biophys. Res. Commun. 2007, 355, 993–999. [Google Scholar] [CrossRef]
- Bandhuvula, P.; Tam, Y.Y.; Oskouian, B.; Saba, J.D. The immune modulator FTY720 inhibits sphingosine-1-phosphate lyase activity. J. Biol. Chem. 2005, 280, 33697–33700. [Google Scholar] [CrossRef]
- Limaye, V.; Vadas, M.A.; Pitson, S.M.; Gamble, J.R. The effects of markedly raised intracellular sphingosine kinase-1 activity in endothelial cells. Cell Mol. Biol. Lett. 2009, 14, 411–423. [Google Scholar] [CrossRef]
- Chipuk, J.E.; McStay, G.P.; Bharti, A.; Kuwana, T.; Clarke, C.J.; Siskind, L.J.; Obeid, L.M.; Green, D.R. Sphingolipid metabolism cooperates with BAK and BAX to promote the mitochondrial pathway of apoptosis. Cell 2012, 148, 988–1000. [Google Scholar] [CrossRef]
- Anavi-Goffer, S.; Baillie, G.; Irving, A.J.; Gertsch, J.; Greig, I.R.; Pertwee, R.G.; Ross, R.A. Modulation of L-α-lysophosphatidylinositol/GPR55 mitogen-activated protein kinase (MAPK) signaling by cannabinoids. J. Biol. Chem. 2012, 287, 91–104. [Google Scholar]
- Tigyi, G. Aiming drug discovery at lysophosphatidic acid targets. Br. J. Pharmacol. 2010, 161, 241–270. [Google Scholar] [CrossRef]
- Siow, D.L.; Anderson, C.D.; Berdyshev, E.V.; Skobeleva, A.; Pitson, S.M.; Wattenberg, B.W. Intracellular localization of sphingosine kinase 1 alters access to substrate pools but does not affect the degradative fate of sphingosine-1-phosphate. J. Lipid Res. 2011, 51, 2546–2559. [Google Scholar]
- Halkidou, K.; Gnanapragasam, V.J.; Mehta, P.B.; Logan, I.R.; Brady, M.E.; Cook, S.; Leung, H.Y.; Neal, D.E.; Robson, C.N. Expression of Tip60, an androgen receptor coactivator, and its role in prostate cancer development. Oncogene 2003, 22, 2466–2477. [Google Scholar] [CrossRef]
- Gorshkova, I.; He, D.; Berdyshev, E.; Usatuyk, P.; Burns, M.; Kalari, S.; Zhao, Y.; Pendyala, S.; Garcia, J.G.; Pyne, N.J.; Brindley, D.N.; Natarajan, V. Protein kinase C-epsilon regulates sphingosine 1-phosphate-mediated migration of human lung endothelial cells through activation of phospholipase D2, protein kinase-zeta, and Racl. J. Biol. Chem. 2008, 283, 11794–11806. [Google Scholar] [CrossRef]
- Zhao, Y.; Kalari, S.K.; Usatyuk, P.V.; Gorshkova, I.; He, D.; Watkins, T.; Brindley, D.N.; Sun, C.; Bittman, R.; Garcia, J.G.; et al. Intracellular generation of sphingosine 1-phosphate in human lung endothelial cells: Role of lipid phosphate phosphatase-1 and sphingosine kinase 1. J. Biol. Chem. 2007, 282, 14165–14177. [Google Scholar] [CrossRef]
- Berdyshev, E.V.; Gorshkova, I.A.; Garcia, J.G.; Natarajan, V.; Hubbard, W.C. Quantitative analysis of sphingoid base-1-phosphates as bisacetylated derivatives by liquid chromatography-tandem mass spectrometry. Anal. Biochem. 2005, 339, 129–136. [Google Scholar] [CrossRef]
- Berdyshev, E.V.; Gorshkova, I.A.; Usatyuk, P.; Zhao, Y.; Saatian, B.; Hubbard, W.; Natarajan, V. De novo biosynthesis of dihydrosphingosine-1-phosphate by sphingosine kinase 1 in mammalian cells. Cell. Signal. 2006, 18, 1779–1792. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Tonelli, F.; Alossaimi, M.; Natarajan, V.; Gorshkova, I.; Berdyshev, E.; Bittman, R.; Watson, D.G.; Pyne, S.; Pyne, N.J. The Roles of Sphingosine Kinase 1 and 2 in Regulating the Metabolome and Survival of Prostate Cancer Cells. Biomolecules 2013, 3, 316-333. https://doi.org/10.3390/biom3020316
Tonelli F, Alossaimi M, Natarajan V, Gorshkova I, Berdyshev E, Bittman R, Watson DG, Pyne S, Pyne NJ. The Roles of Sphingosine Kinase 1 and 2 in Regulating the Metabolome and Survival of Prostate Cancer Cells. Biomolecules. 2013; 3(2):316-333. https://doi.org/10.3390/biom3020316
Chicago/Turabian StyleTonelli, Francesca, Manal Alossaimi, Viswanathan Natarajan, Irina Gorshkova, Evgeny Berdyshev, Robert Bittman, David G. Watson, Susan Pyne, and Nigel J. Pyne. 2013. "The Roles of Sphingosine Kinase 1 and 2 in Regulating the Metabolome and Survival of Prostate Cancer Cells" Biomolecules 3, no. 2: 316-333. https://doi.org/10.3390/biom3020316