Comparative Ultrastructure and Carbohydrate Composition of Gastroliths from Astacidae, Cambaridae and Parastacidae Freshwater Crayfish (Crustacea, Decapoda)


Abstract
:
1. Introduction
2. Results and Discussion
2.1. Microscopic Analysis of the Gastrolith Structure
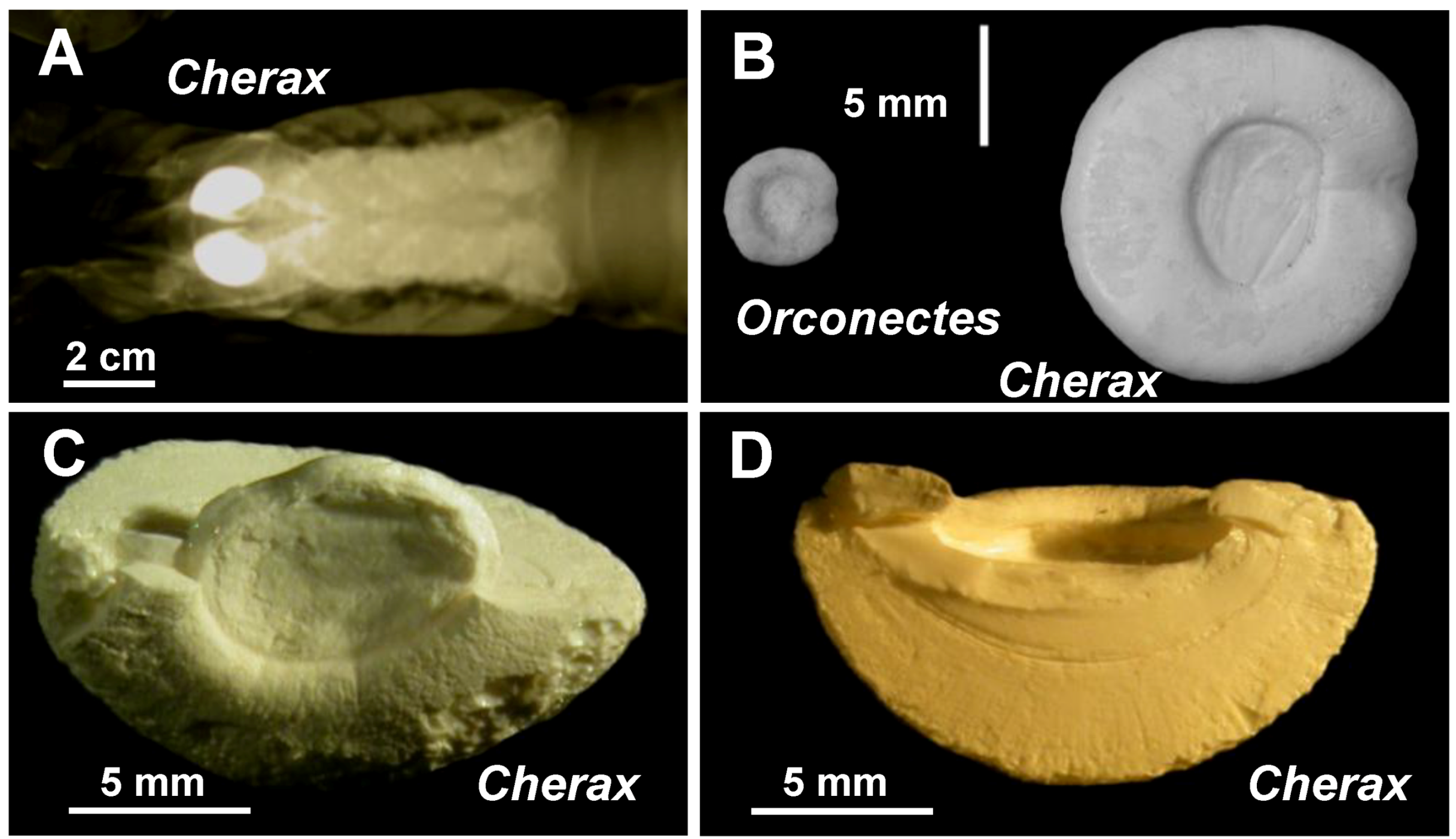
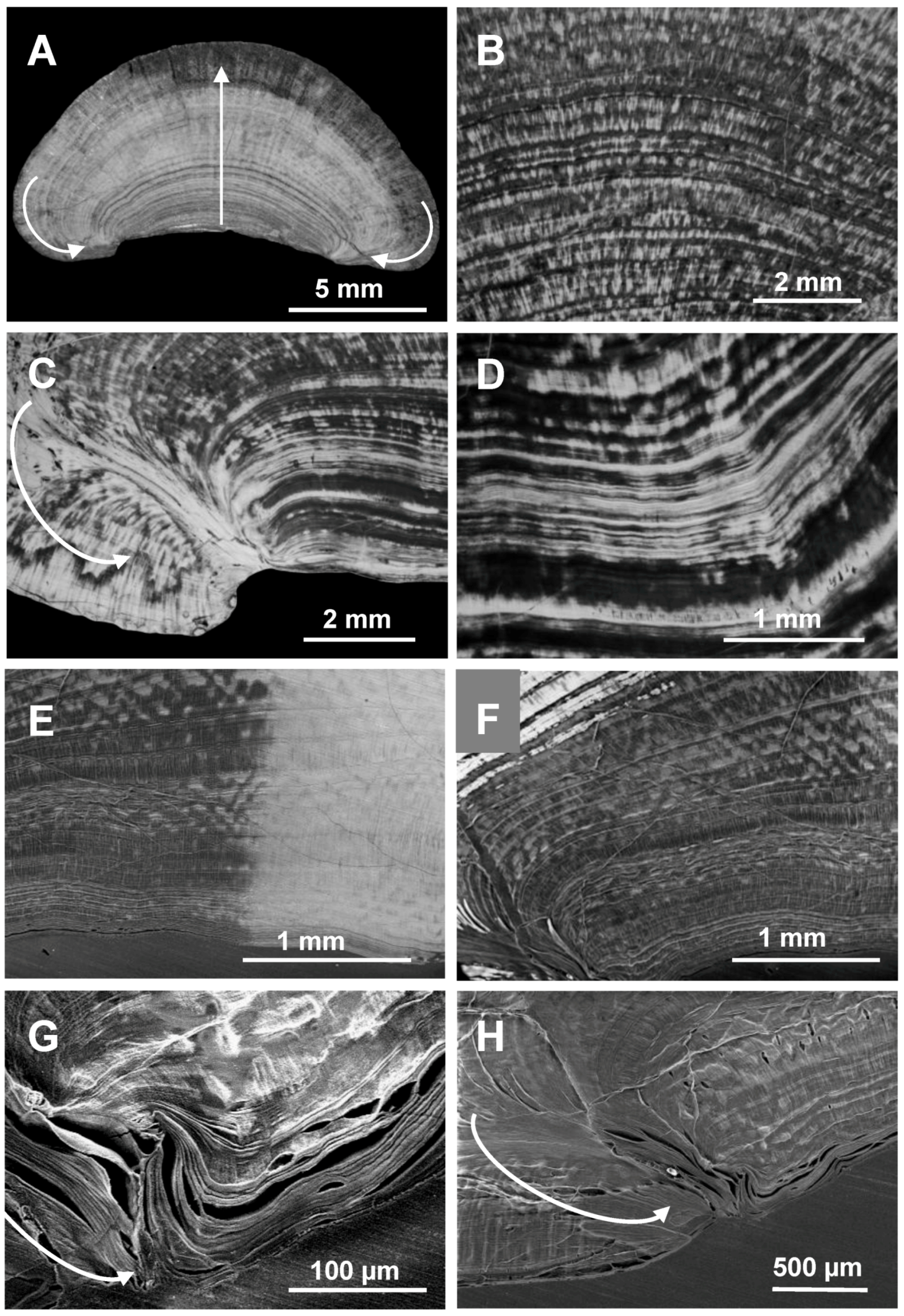


2.2. Mineralogical Composition
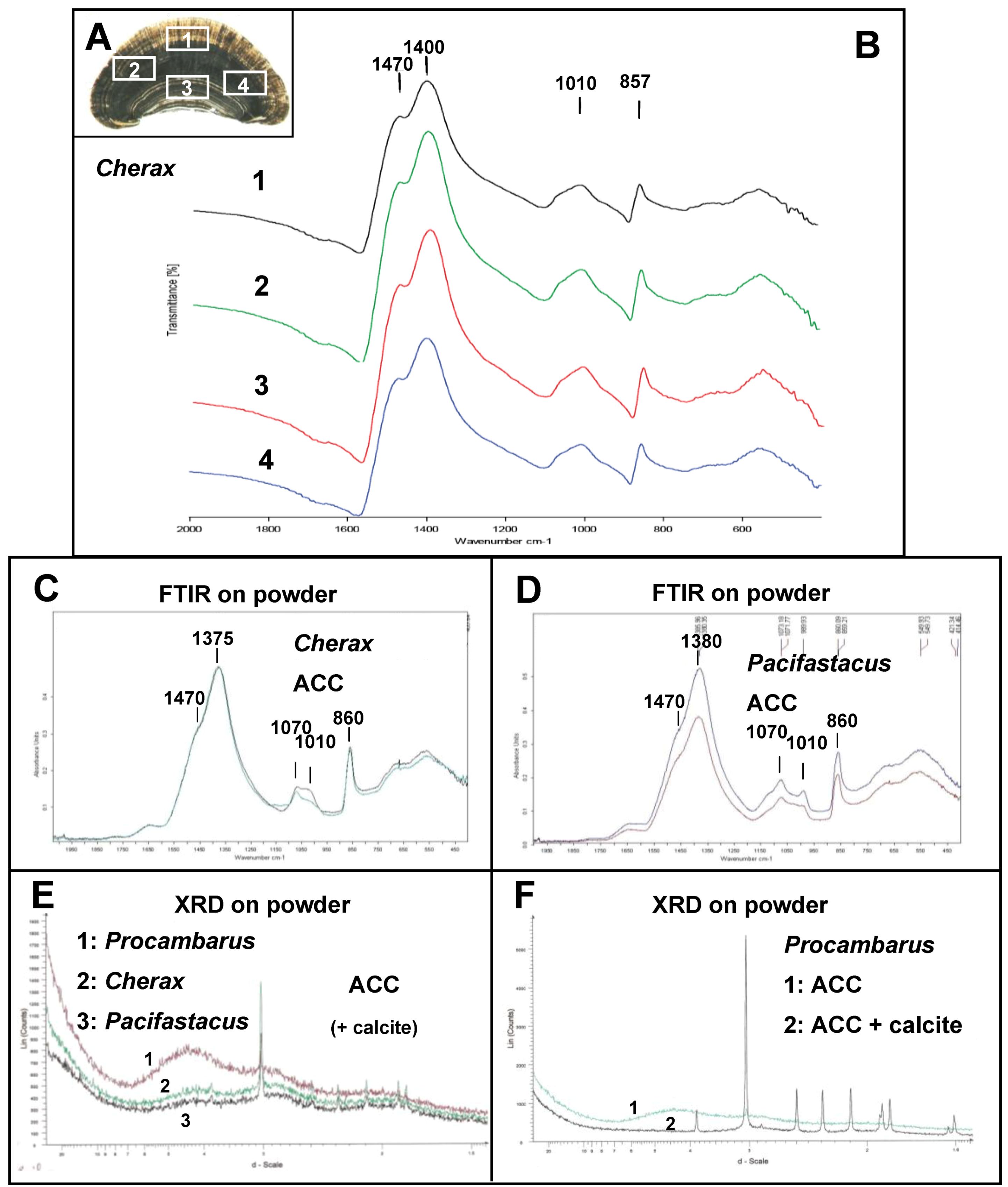
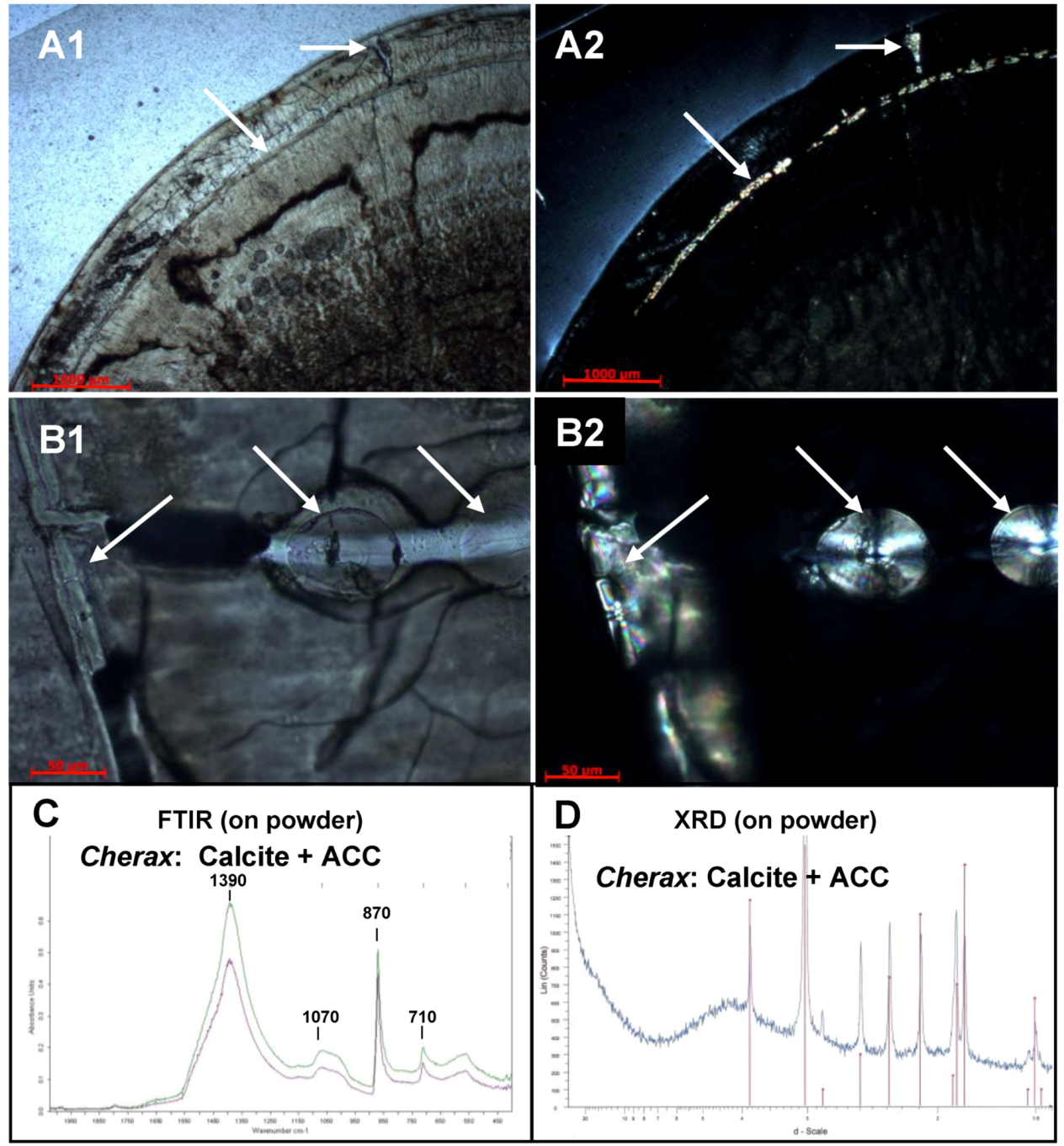
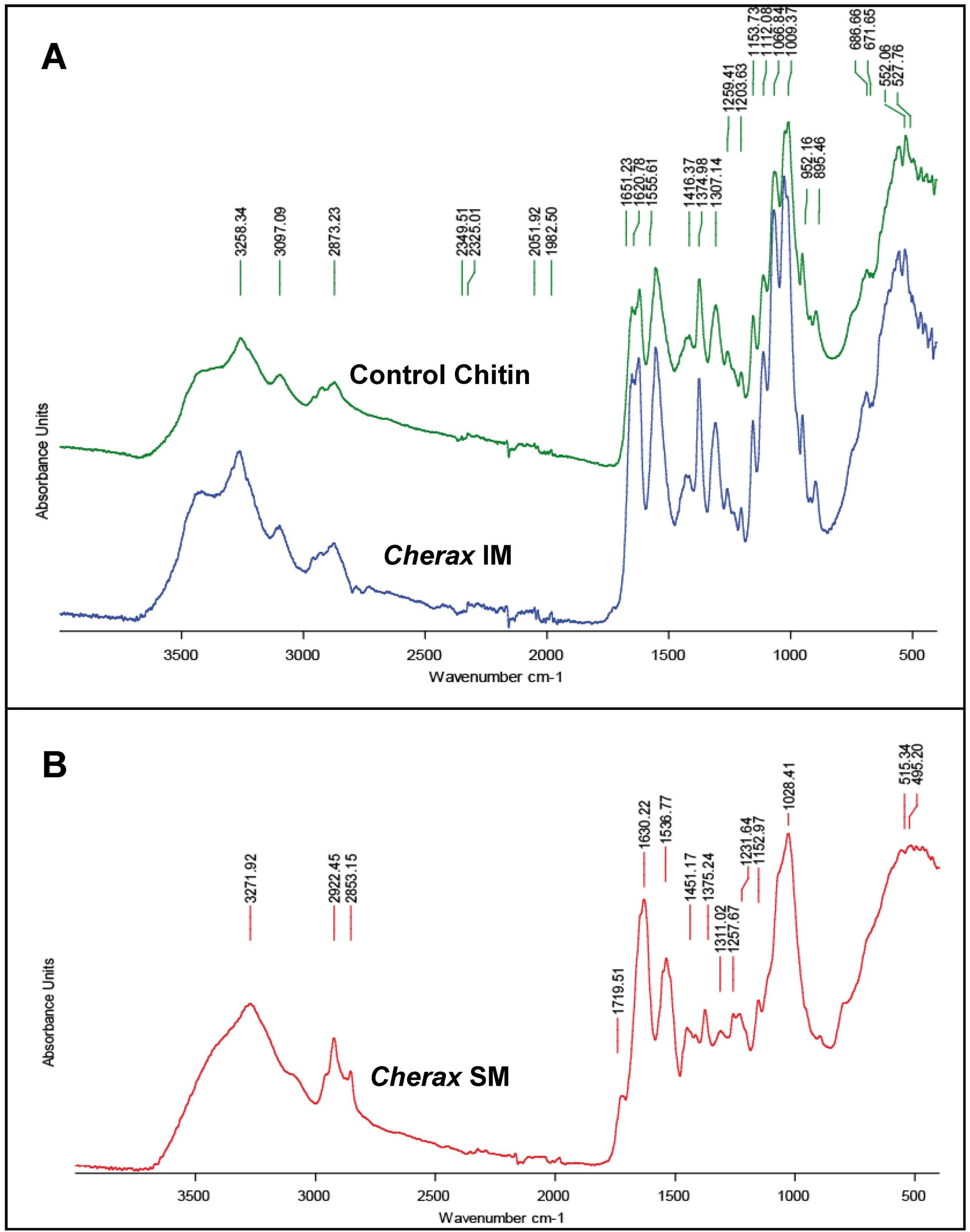
2.3. Sugar Analysis and FTIR Matrix Analysis

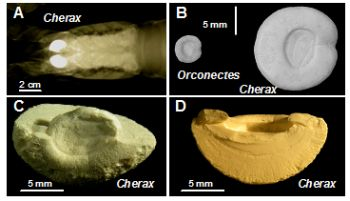{kind=link}
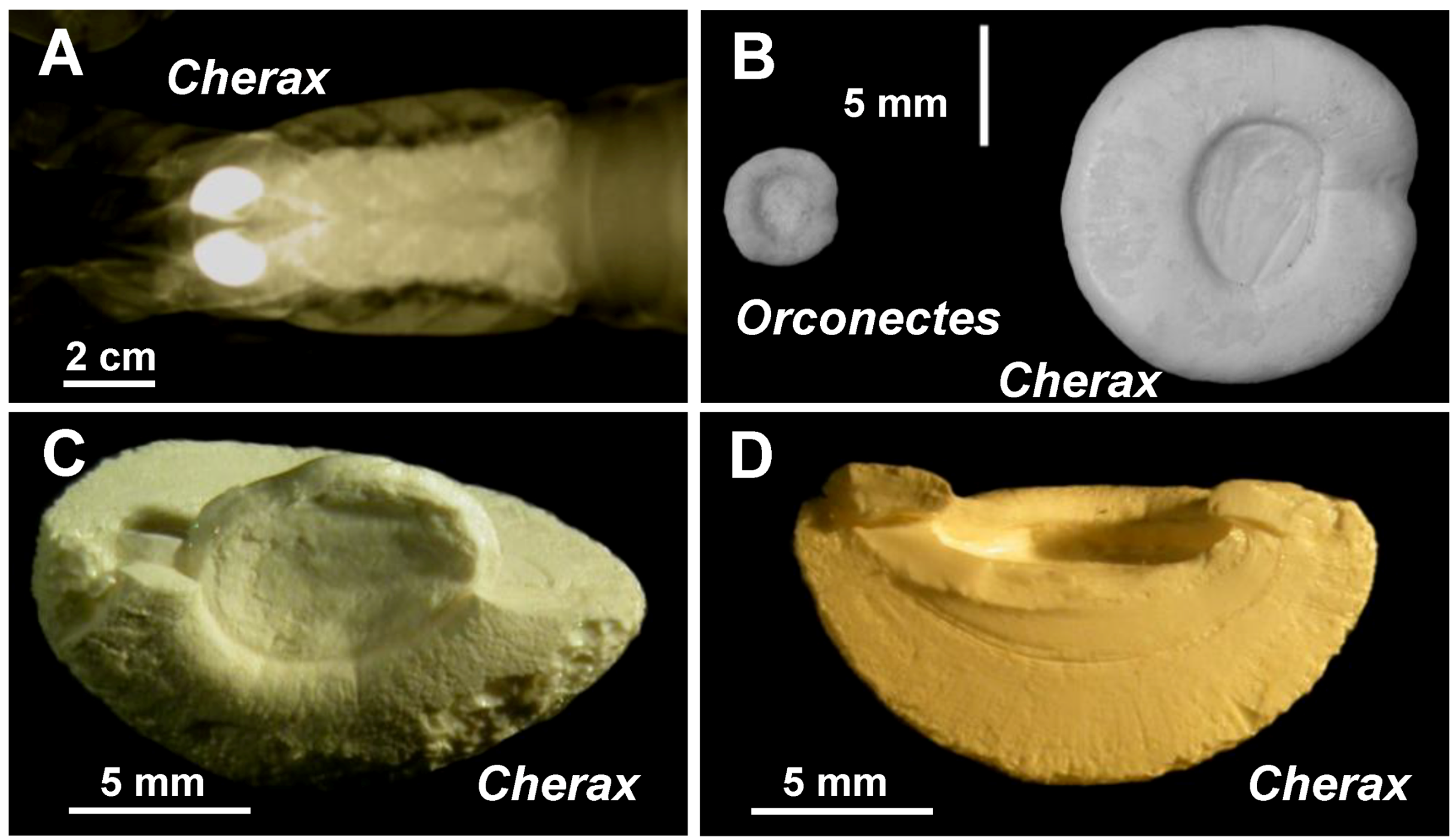{kind=link}
{kind=link}
{kind=link}
{kind=link}
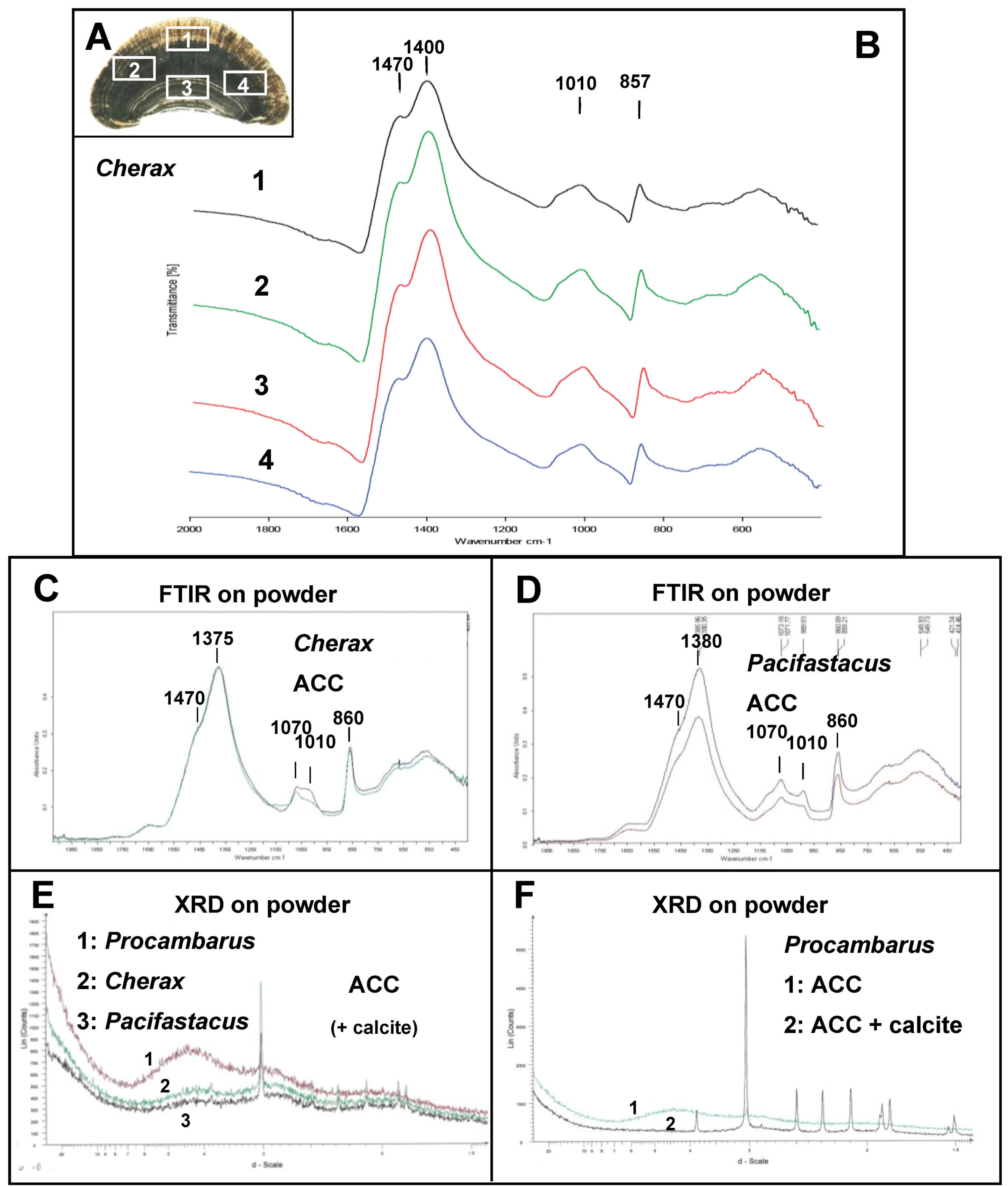{kind=link}
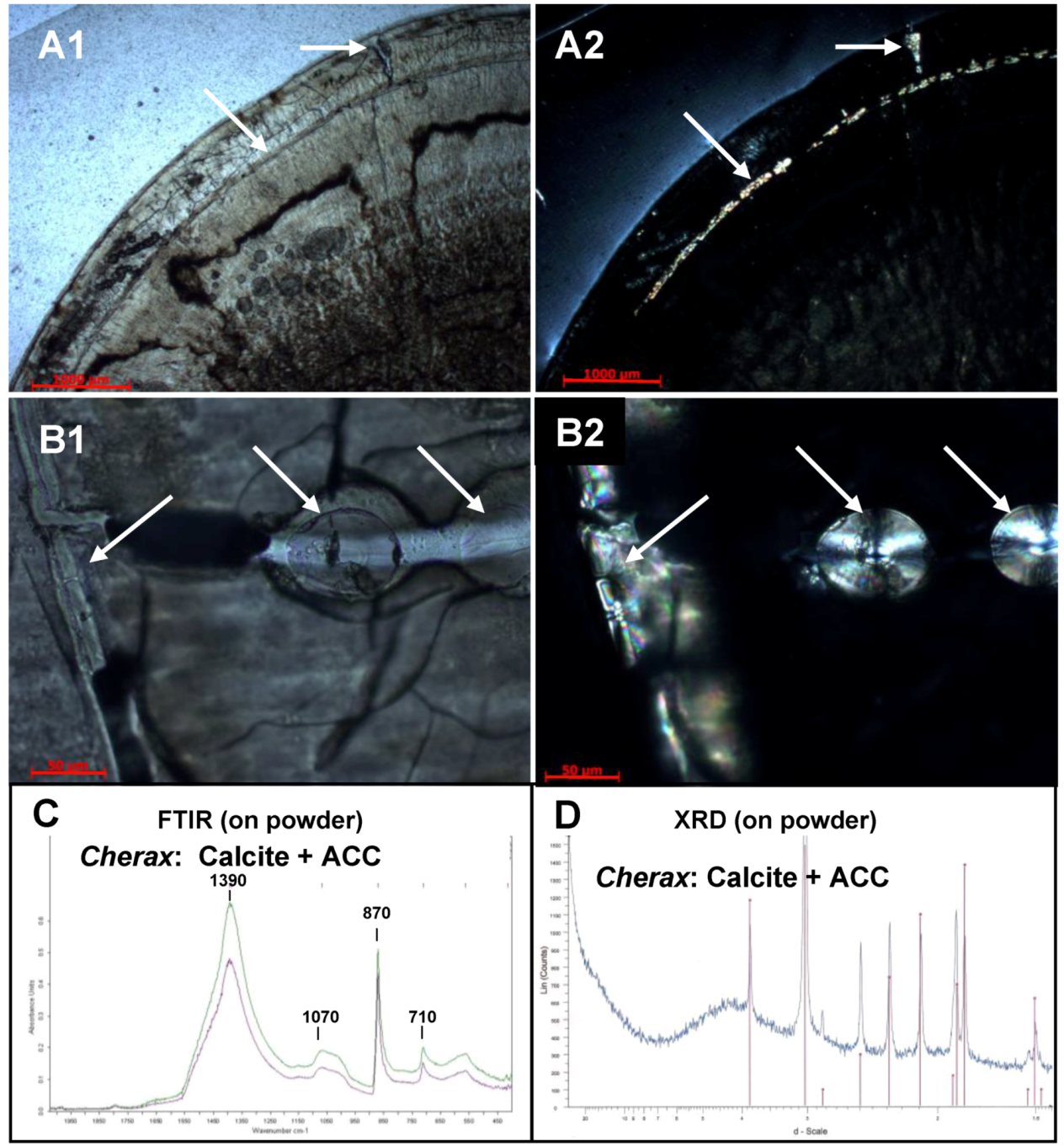{kind=link}
{kind=link}
 |
3. Experimental Section
3.1. Animals
3.2. Samples Preparation and Chemical Treatments
3.3. Light Microscopy, Scanning Electron Microscopy and Polarized Light Microscopy
3.4. X-ray Diffraction
3.5. Fourier transform infrared spectroscopy
3.6. Sugar Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Graf, F. Les sources de calcium pour les crustacés venant de muer. Arch. Zool. Exp. Gen. 1978, 119, 143–161. [Google Scholar]
- Greenaway, P. Calcium balance and molting in the Crustacea. Biol. Rev. 1985, 60, 425–454. [Google Scholar] [CrossRef]
- Wheatly, M.G. Calcium homeostasis in Crustacea: the evolving role of branchial, renal, digestive and hypodermal epithelia. J. Exp. Zool. 1999, 283, 620–64. [Google Scholar] [CrossRef]
- Luquet, G.; Marin, F. Biomineralization in crustaceans: Storage strategies. C. R. Palevol. 2004, 3, 515–534. [Google Scholar] [CrossRef]
- Luquet, G. Biomineralizations: insights and prospects from crustaceans. Zookeys 2012, 176, 103–121. [Google Scholar] [CrossRef]
- Travis, D.F. The deposition of skeletal structures in the crustacea. I. The histology of the gastrolith skeletal tissue complex and the gastrolith in the crayfish, Orconectes (Cambarus) virilis Hagen - Decapoda. Biol. Bull. 1960, 18, 137–149. [Google Scholar] [CrossRef]
- Travis, D.F. Structural features of mineralization from tissue to macromolecular levels of organization in the decapod Crustacea. Ann. N. Y. Acad. Sci. 1963, 109, 177–245. [Google Scholar] [CrossRef]
- Hikida, T.; Nagasawa, H.; Kogure, T. Characterization of amorphous calcium carbonate in the gastrolith of crayfish, Procambarus clarkii. In Biomineralization: formation, diversity, evolution and application; Kobayashi, I., Ozawa, H., Eds.; Tokai University Press: Kanagawa, Japan, 2003; pp. 81–84. [Google Scholar]
- Shechter, A.; Berman, A.; Singer, A.; Freiman, A.; Grinstein, M.; Erez, J.; Aflalo, E.D.; Sagi, A. Reciprocal changes in calcification of the gastrolith and cuticle during the molt cycle of the red claw crayfish Cherax quadricarinatus. Biol. Bull. 2008, 214, 122–134. [Google Scholar] [CrossRef]
- Scott, D.; Duncan, K.W. The function of freshwater crayfish and their occurrence in perch, trout and shag stomachs. New Zeal. J. Mar. Fresh. Res. 1967, 1, 99–104. [Google Scholar] [CrossRef]
- Mann, S. Mineralization in biological systems. Struct. Bond. 1983, 54, 125–174. [Google Scholar] [CrossRef]
- Lowenstam, H.A.; Weiner, S. On biomineralization; Oxford University Press: NewYork, USA, 1989; p. 324. [Google Scholar]
- Bentov, S.; Weil, S.; Glazer, L.; Sagi, A.; Berman, A. Stabilization of amorphous calcium carbonate by phosphate rich organic matrix proteins and by single phosphoamino acids. J. Struct. Biol. 2010, 17, 207–215. [Google Scholar]
- Ishii, K.; Yanagisawa, T.; Nagasawa, H. Characterization of a matrix protein in the gastroliths of the crayfish Procambarus clarkii. Biosci. Biotechnol. Biochem. 1996, 60, 1479–1482. [Google Scholar] [CrossRef]
- Ishii, K.; Tsutsui, N.; Watanabe, T.; Yanagisawa, T.; Nagasawa, H. Solubilization and chemical characterization of an insoluble matrix protein in the gastroliths of a crayfish, Procambarus clarkii. Biosci. Biotech. Biochem. 1998, 62, 291–296. [Google Scholar] [CrossRef]
- Tsutsui, N.; Ishii, K.; Takagi, Y.; Watanabe, T.; Nagasawa., H. Cloning and expression of a cDNA encoding an insoluble matrix protein in the gastroliths of a crayfish, Procambarus clarkii. Zool. Sci. 1999, 16, 619–628. [Google Scholar] [CrossRef]
- Takagi, Y.; Ishii, K.; Ozaki, N.; Nagasawa, H. Immunolocalization of gastrolith matrix protein (GAMP) in the gastroliths and exoskeleton of crayfish, Procambarus clarkii. Zool. Sci. 2000, 17, 179–184. [Google Scholar] [CrossRef]
- Shechter, A.; Glazer, L.; Cheled, S.; Mor, E.; Weil, S.; Berman, A.; Bentov, S.; Aflalo, E.D.; Khalaila, I.; Sagi, A. A gastrolith protein serving a dual role in the formation of an amorphous mineral containing extracellular matrix. Proc. Natl. Acad. Sci. USA 2008, 105, 7129–7134. [Google Scholar]
- Glazer, L.; Shechter, A.; Tom, M.; Yudkovski, Y.; Well, S.; Aflalo, E.D.; Pamuru, R.R.; Khalaila, I.; Bentov, S.; Berman, A.; Sagi, A. A protein involved in the assembly of an extracellular calcium storage matrix. J. Biol. Chem. 2010, 285, 12831–12839. [Google Scholar]
- Luquet, G.; Le Roy, N.; Zanella-Cléon, I.; Becchi, M.; Bucarey, S.; Fernandez, M.S.; Arias, J.L.; Guichard, N.; Marie, B.; Marin, F. Characterization of crustacyanin-A2 subunit as a component of the organic matrix of gastroliths from the crayfish Cherax quadricarinatus. In Structure-Property Relationships in Biomineralized and Biomimetic Composites; Kisalius, D., Estroff, L., Landis, W., Zavattieri, P., Gupta, H.S., Eds.; Warrendale, PA, USA, Mater. Res. Soc. Symp. Proc. 2009, 1187, 69–75.
- Yudkovski, Y.; Glazer, L.; Shechter, A.; Reinhardt, R.; Chalifa-Caspi, V.; Sagi, A.; Tom, M. Multi-transcript expression patterns in the gastrolith disk and the hypodermis of the crayfish Cherax quadricarinatus at premolt. Comp. Biochem. Physiol. D 2010, 5, 171–177. [Google Scholar]
- Glazer, L.; Sagi, A. On the involvement of proteins in the assembly of the crayfish gastrolith extracellular matrix. Invert. Rep. Dev. 2012, 56, 57–65. [Google Scholar] [CrossRef]
- Luquet, G.; Fernandez, M.S.; Navarrete, M.J.; Arias, J.L.; Guichard, N.; Marie, B.; Marin, F. Biochemical characterization of the soluble organic matrix of gastroliths from decapods. In Biomineralization, from Paleontology to Materials Science; Arias, J.L., Fernandez, M.S., Eds.; Editorial Universitaria: Santiago, Chile, 2007; pp. 319–328. [Google Scholar]
- Akiva-Tal, A.; Kabaya, S.; Balazs, Y.S.; Glazer, L.; Berman, A.; Sagi, A.; Schmidt, A. In situ molecular NMR picture of bioavailable calcium stabilized as amorphous CaCO3 biomineral in crayfish gastroliths. Proc. Natl. Acad. Sci. USA 2011, 108, 14763–14768. [Google Scholar]
- Sato, A.; Nagasaka, S.; Fuhirata, K.; Nagata, S.; Arai, S.; Saruwatari, K.; Kogure, T.; Sakuda, S.; Nagasawa, H. Glycolytic intermediates induce amorphous calcium carbonate formation in crustaceans. Nat. Chem. Biol. 2011, 7, 197–199. [Google Scholar] [CrossRef]
- Arias, J.L.; Fernandez, M.S. Biomimetic processes through the study of mineralized shells. Mater. Charact. 2003, 50, 189–195. [Google Scholar] [CrossRef]
- Rodríguez-Navarro, A.; Cabral de Melo, C.; Batista, N.; Morimoto, N.; Alvarez-Lloret, P.; Ortega-Huertas, M.; Fuenzalida, V.M.; Arias, J.I.; Wiff, J.P.; Arias, J.L. Microstructure and crystallographic-texture of giant barnacle (Austromegabalanus psittacus) shell. J. Struct. Biol. 2006, 156, 355–362. [Google Scholar] [CrossRef]
- Fernandez, M.S.; Bustos, C.; Luquet, G.; Saez, D.; Neira-Carrillo, A.; Corneillat, M.; Alcaraz, G.; Arias, J.L. Proteoglycan occurrence in gastrolith of the crayfish Cherax quadricarinatus (Crustacea, Malacostraca, Decapoda). J. Crust. Biol. 2012, 32, 802–815. [Google Scholar] [CrossRef]
- Westbroek, P.; de Jong, E.W.; Dam, W.; Bosch, L. Soluble intracrystalline polysaccharides from coccoliths of Coccolithus huxleyi (Lohmann) Kamptner (I). Calcif. Tissue Res. 1973, 12, 227–238. [Google Scholar] [CrossRef]
- de Jong, E.W.; Bosch, L.; Westbroek, P. Isolation and characterization of a Ca2+-binding polysaccharide associated with coccoliths of Emiliania huxleyi (Lohmann) Kamptner. Eur. J. Biochem. 1976, 70, 611–621. [Google Scholar] [CrossRef]
- Borman, A.H.; de Jong, E.W.; Huizinga, M.; Kok, D.J.; Westbroek, P.; Bosch, L. The role in CaCO3 crystallization of an acid Ca2+-binding polysaccharide associated with coccoliths of Emiliania huxleyi. Eur. J. Biochem. 1982, 129, 179–183. [Google Scholar] [CrossRef]
- Yang, M.; Stipp, S.L.S.; Harding, J. Biological control on calcite crystallization by polysaccharides. Cryst. Growth Des. 2008, 8, 4066–4074. [Google Scholar] [CrossRef]
- Kingsley, R.J.; Watabe, N. Analysis of proteinaceous components of the organic matrices of spicules from the gorgonian Leptogorgia virgulata. Comp. Biochem. Physiol. B 1983, 76, 443–447. [Google Scholar] [CrossRef]
- Cuif, J.P.; Gautret, P. Glucides et protéines de la matrice soluble des biocristaux de Scléractinaires Acroporidés. C. R. Acad. Sci. Paris 1995, 320, 273–278. [Google Scholar]
- Collins, M.J.; Muyzer, G.; Curry, G.B.; Sandberg, P.; Westbroek, P. Macromolecules in brachiopod shells: characterization and diagenesis. Lethaia 1991, 24, 387–97. [Google Scholar] [CrossRef]
- Gaspard, D.; Marin, F.; Guichard, N.; Morel, S.; Alcaraz, G.; Luquet, G. Shell matrices of recent Rhynchonelliform Brachiopods: microstructures and glycosylation studies. R. Soc. Edinburgh Trans. 2008, 98, 415–424. [Google Scholar]
- Benson, S.C.; Benson, N.C.; Wilt, F. The organic matrix of the skeletal spicule of sea urchin embryos. J. Cell. Biol. 1986, 102, 1878–7886. [Google Scholar] [CrossRef]
- Albeck, S.; Addadi, L.; Weiner, S. Regulation of calcite crystal morphology by intracrystalline acidic proteins and glycoproteins. Connect. Tissue Res. 1996, 35, 365–370. [Google Scholar] [CrossRef]
- Albeck, S.; Weiner, S.; Addadi, L. Polysaccharides of intracrystallline glycoproteins modulate calcite crystal growth in vitro. Chem. Eur. J. 1996, 2, 278–284. [Google Scholar] [CrossRef]
- Wheeler, A.P.; George, J.W.; Evans, C.A. Control of calcium carbonate nucleation and crystal growth by soluble matrix of oyster shell. Science 1981, 212, 1397–1398. [Google Scholar]
- Marie, B.; Luquet, G.; Pais De Barros, J.P.; Guichard, N.; Morel, S.; Alcaraz, G.; Bollache, L.; Marin, F. The shell matrix of the freshwater mussel Unio pictorum (Paleoheterodonta, Unionoida). Involvement of acidic polysaccharides from glycoproteins in nacre mineralization. FEBS J. 2007, 274, 2933–2945. [Google Scholar] [CrossRef]
- Marie, B.; Marin, F.; Marie, A.; Bédouet, L.; Dubost, L.; Alcaraz, G.; Milet, C.; Luquet, G. Evolution of nacre: biochemistry and proteomics of the shell organic matrix of the cephalopod Nautilus macromphalus. ChemBioChem. 2009, 26, 1495–1506. [Google Scholar]
- Marie, B.; Zanella-Cléon, I.; Corneillat, M.; Becchi, M.; Alcaraz, G.; Plasseraud, L.; Luquet, G.; Marin, F. Nautilin-63, a novel acidic glycoprotein from the shell nacre of Nautilus macromphal. FEBS J. 2011, 278, 2117–2130. [Google Scholar] [CrossRef]
- Pavat, C.; Zanella-Cléon, I.; Becchi, M.; Medakovic, D.; Luquet, G.; Guichard, N.; Alcaraz, G.; Dommergues, J-L.; Serpentini, A.; Lebel, J.-M.; Marin, F. The shell matrix of the pulmonate land snail Helix aspersa maxima. Comp. Biochem. Physiol. B 2012, 161, 303–314. [Google Scholar] [CrossRef]
- Addadi, L.; Moradian, J.; Shay, E.; Maroudas, M.G.; Weiner, S. A chemical model for the cooperation of sulfates and carboxylates in calcite crystal nucleation: relevance to biomineralization. Proc. Natl. Acad. Sci. USA 1987, 84, 2732–2736. [Google Scholar] [CrossRef]
- Zhong, C.; Chu, C.C. Acid polysaccharide-induced amorphous calcium carbonate (ACC) films: colloidal nanoparticle self-organization process. Langmuir 2009, 25, 3045–3049. [Google Scholar] [CrossRef]
- Dillaman, R.; Hequembourg, S.; Gay, M. Early pattern of calcification in the dorsal carapace of the blue crab, Callinectes sapidus. J. Morphol. 2005, 263, 356–374. [Google Scholar] [CrossRef]
- Romano, P.; Fabritius, H.; Raabe, D. The exoskeleton of the lobster, Homarus americanus, as an example of a smart anisotropic biological material. Acta Biomater. 2007, 3, 301–309. [Google Scholar] [CrossRef]
- Soejoko, D.S.; Tjia, M.O. Infrared spectroscopy and X-ray diffraction study on the morphological variations of carbonate and phosphate compounds in giant prawn (Macrobrachium rosenbergii) skeletons during its moulting period. J. Mater. Sci. 2003, 38, 2087–2093. [Google Scholar] [CrossRef]
- Dauphin, Y. Infrared spectra and elemental composition in recent biogenic calcites: Relationships between the ν4 bandwavelength and Sr and Mg concentrations. Appl. Spectrosc. 1997, 51, 253–258. [Google Scholar] [CrossRef]
- Dauphin, Y. Infrared spectra and elemental composition in recent carbonate skeletons: Relationships between the ν2 band wavenumber and Sr and Mg concentrations. Appl. Spectrosc. 1999, 53, 184–190. [Google Scholar] [CrossRef]
- Auzoux-Bordenave, S.; Badou, A.; Gaume, B.; Berland, S.; Helléouet, M.-N.; Milet, C.; Huchette, S. Ultrastructure, chemistry and mineralogy of the growing shell of the European abalone Haliotis tuberculata. J. Struct. Biol. 2010, 171, 277–290. [Google Scholar] [CrossRef]
- Günther, C.; Becker, A.; Wolf, G.; Epple, M. In vitro synthesis and structural characterization of Amorphous Calcium Carbonate. Z. Anorg. Allg. Chem. 2005, 631, 2830–2835. [Google Scholar] [CrossRef]
- Nebel, H.; Neumann, M.; Mayer, C.; Epple, M. On the structure of Amorphous Calcium Carbonate - A detailed study by solid-state NMR spectroscopy. Inorg. Chem. 2008, 47, 7874–7879. [Google Scholar] [CrossRef]
- Senorale-Pose, M.; Chalar, C.; Dauphin, Y.; Massard, P.; Pradel, P.; Marin, M. Monohydrocalcite in calcareous corpuscles of Mesocestoides corti. Exp. Parasitol. 2008, 118, 54–58. [Google Scholar] [CrossRef]
- Raz, S.; Testenière, O.; Hecker, A.; Weiner, S.; Luquet., G. Stable amorphous calcium carbonate is the main components of the calcium storage structures of the crustacean Orchestia cavimana. Biol. Bull. 2002, 203, 269–274. [Google Scholar] [CrossRef]
- Funderburgh, J.L. Keratan sulfate: structure, biosynthesis, and function. Glycobiology 2000, 10, 951–958. [Google Scholar] [CrossRef]
- Arias, J.L.; Fernandez, M.S. Polysaccharides and proteoglycans in calcium carbonate-based biomineralization. Chem. Rev. 2008, 108, 4475–4482. [Google Scholar] [CrossRef]
- Addadi, L.; Raz, S.; Weiner, S. Taking advantage of disorder: amorphous calcium carbonate and its role in biomineralization. Adv. Mater. 2003, 15, 959–970. [Google Scholar] [CrossRef]
- Aizenberg, J.; Lambert, G.; Addadi, L.; Weiner, S. Stabilization of amorphous calcium carbonate by specialized macromolecules in biological and synthetic precipitates. Adv. Mater. 1996, 8, 222–226. [Google Scholar] [CrossRef]
- Aizenberg, J.; Lambert, G.; Weiner, S.; Addadi, L. Factors involved in the formation of amorphous and crystalline calcium carbonate: A study of an ascidian skeleton. J. Amer. Chem. Soc. 2002, 124, 32–39. [Google Scholar] [CrossRef]
- Ajikumar, P.K.; Wong, L.G.; Subramanyam, G.; Lakshminarayanan, R.; Valiyaveettil, S. Synthesis and characterization of monodispersed spheres of amorphous calcium carbonate and calcite spherules. Cryst. Growth Des. 2005, 5, 1129–1134. [Google Scholar] [CrossRef]
- Al-Sawalmih, A.; Li, C.; Siegel, S.; Fratzl, P.; Paris, O. On the stability of amorphous minerals in lobster cuticle. Adv. Mater. 2009, 21, 391–400. [Google Scholar] [CrossRef]
- Loste, E.; Wilson, R.W.; Seshadri, R.; Meldrum, F. The role of magnesium in stabilizing amorphous calcium carbonate and controlling calcite morphologies. J. Cryst. Growth 2003, 254, 206–2178. [Google Scholar] [CrossRef]
- Liu, Y.; Cui, Y.; Guo, R. Amphiphilic phosphoprotein-controlled formation of amorphous calcium carbonate with hierarchical superstructure. Langmuir 2012, 28, 6097–6105. [Google Scholar]
- Raz, S.; Weiner, S.; Addadi, L. Formation of high-magnesian calcites via an amorphous precursor phase: possible biological implications. Adv. Mater. 2000, 12, 38–42. [Google Scholar] [CrossRef]
- Raz, S.; Hamilton, P.C.; Wilt, F.H.; Weiner, S.; Addadi, L. The transient phase of amorphous calcium carbonate in sea urchin larval spicules: The involvement of proteins and magnesium ions in its formation and stabilization. Adv. Funct. Mater. 2003, 13, 480–486. [Google Scholar] [CrossRef]
- Fröhlich, F.; Gendron-Badou, A. La spectroscopie infrarourge, un outil polyvalent. In Géologie de la Préhistoire; Miskovsky, J.C., Ed.; AEEGP éditeur: Paris, France, 2002; pp. 662–677. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Luquet, G.; Fernández, M.S.; Badou, A.; Guichard, N.; Roy, N.L.; Corneillat, M.; Alcaraz, G.; Arias, J.L. Comparative Ultrastructure and Carbohydrate Composition of Gastroliths from Astacidae, Cambaridae and Parastacidae Freshwater Crayfish (Crustacea, Decapoda). Biomolecules 2013, 3, 18-38. https://doi.org/10.3390/biom3010018
Luquet G, Fernández MS, Badou A, Guichard N, Roy NL, Corneillat M, Alcaraz G, Arias JL. Comparative Ultrastructure and Carbohydrate Composition of Gastroliths from Astacidae, Cambaridae and Parastacidae Freshwater Crayfish (Crustacea, Decapoda). Biomolecules. 2013; 3(1):18-38. https://doi.org/10.3390/biom3010018
Chicago/Turabian StyleLuquet, Gilles, María S. Fernández, Aïcha Badou, Nathalie Guichard, Nathalie Le Roy, Marion Corneillat, Gérard Alcaraz, and José L. Arias. 2013. "Comparative Ultrastructure and Carbohydrate Composition of Gastroliths from Astacidae, Cambaridae and Parastacidae Freshwater Crayfish (Crustacea, Decapoda)" Biomolecules 3, no. 1: 18-38. https://doi.org/10.3390/biom3010018
APA StyleLuquet, G., Fernández, M. S., Badou, A., Guichard, N., Roy, N. L., Corneillat, M., Alcaraz, G., & Arias, J. L. (2013). Comparative Ultrastructure and Carbohydrate Composition of Gastroliths from Astacidae, Cambaridae and Parastacidae Freshwater Crayfish (Crustacea, Decapoda). Biomolecules, 3(1), 18-38. https://doi.org/10.3390/biom3010018