
New Variants of the Cytochrome P450 2R1 (CYP2R1) Gene in Individuals with Severe Vitamin D-Activating Enzyme 25(OH)D Deficiency

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Biochemical Analyses
2.3. CYP2R1 Gene Sequencing
2.3.1. DNA Isolation
2.3.2. PCR of Selected Fragments of the CYP2R1 Gene
2.3.3. Purification of the CYP2R1 PCR Products
2.3.4. Fluorescence-Based Cycle Sequencing Reaction
2.3.5. Purification of Templates after Cycle Sequencing Reaction
2.3.6. Capillary Electrophoresis
2.4. Bioinformatic Analysis
2.5. Statistical Methods
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sirajudeen, S.; Shah, I.; Al Menhali, A. A Narrative Role of Vitamin D and Its Receptor: With Current Evidence on the Gastric Tissues. Int. J. Mol. Sci. 2019, 20, 3832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jäpelt, R.B.; Jakobsen, J. Vitamin D in plants: A review of occurrence, analysis, and biosynthesis. Front. Plant Sci. 2013, 4, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, R.; Maseeh, A. Vitamin D: The sunshine vitamin. J. Pharmacol. Pharmacother. 2012, 3, 118–126. [Google Scholar] [CrossRef]
- Sahota, O. Understanding vitamin D deficiency. Age Ageing 2014, 43, 589–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dziedzic, E.A.; Gąsior, J.S.; Pawłowski, M.; Wodejko-Kucharska, B.; Saniewski, T.; Marcisz, A.; Dąbrowski, M.J. Vitamin D level is associated with severity of coronary artery atherosclerosis and incidence of acute coronary syndromes in non-diabetic cardiac patients. Arch. Med. Sci. 2019, 15, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Mozos, I.; Marginean, O. Links between Vitamin D Deficiency and Cardiovascular Diseases. BioMed Res. Int. 2015, 2015, 1–12. [Google Scholar] [CrossRef]
- Judd, S.E.; Tangpricha, V. Vitamin D Deficiency and Risk for Cardiovascular Disease. Am. J. Med. Sci. 2009, 338, 40–44. [Google Scholar] [CrossRef] [Green Version]
- Płudowski, P.; Karczmarewicz, E.; Bayer, M.; Carter, G.; Chlebna-Sokół, D.; Czech-Kowalska, J.; Dębski, R.; Decsi, T.; Dobrzanska, A.; Franek, E.; et al. Practical guidelines for the supplementation of vitamin D and the treatment of deficits in Central Europe—Recommended vitamin D intakes in the general population and groups at risk of vitamin D deficiency. Endokrynol. Pol. 2013, 64, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Rusińska, A.; Pludowski, P.; Walczak, M.; Borszewska-Kornacka, M.K.; Bossowski, A.; Chlebna-Sokół, D.; Czech-Kowalska, J.; Dobrzanska, A.; Franek, E.; Helwich, E.; et al. Vitamin D Supplementation Guidelines for General Population and Groups at Risk of Vitamin D Deficiency in Poland—Recommendations of the Polish Society of Pediatric Endocrinology and Diabetes and the Expert Panel With Participation of National Specialist Consultants and Representatives of Scientific Societies—2018 Update. Front. Endocrinol. 2018, 9, 246. [Google Scholar] [CrossRef]
- Bikle, D. Vitamin D. Production, Metabolism, and Mechanisms of Action; Endotext: Dartmouth, MA, USA, 2017. [Google Scholar]
- HUGO Gene Nomenclature Committee. CNR2. Available online: https://www.genenames.org/data/gene-symbol-report/#!/hgnc_id/HGNC:2160 (accessed on 15 July 2020).
- Thacher, T.D.; Fischer, P.R.; Singh, R.J.; Roizen, J.; Levine, M.A. CYP2R1Mutations Impair Generation of 25-hydroxyvitamin D and Cause an Atypical Form of Vitamin D Deficiency. J. Clin. Endocrinol. Metab. 2015, 100, E1005–E1013. [Google Scholar] [CrossRef] [Green Version]
- Strushkevich, N.; Usanov, S.A.; Plotnikov, A.N.; Jones, G.; Park, H.-W. Structural Analysis of CYP2R1 in Complex with Vitamin D3. J. Mol. Biol. 2008, 380, 95–106. [Google Scholar] [CrossRef]
- Fetahu, I.S.; Höbaus, J.; Kállay, E. Vitamin D and the epigenome. Front. Physiol. 2014, 5, 164. [Google Scholar] [CrossRef] [Green Version]
- Kämpe, A.; Enlund-Cerullo, M.; Valkama, S.; Holmlund-Suila, E.; Rosendahl, J.; Hauta-Alus, H.; Pekkinen, M.; Andersson, S.; Mäkitie, O. Genetic variation in GC and CYP2R1 affects 25-hydroxyvitamin D concentration and skeletal parameters: A genome-wide association study in 24-month-old Finnish children. PLoS Genet. 2019, 15, e1008530. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.M.; Sandler, D.P.; Shi, M.; Harmon, Q.E.; Taylor, J.; Weinberg, C. Genome-Wide Association Study of Serum 25-Hydroxyvitamin D in US Women. Front. Genet. 2018, 9, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, J.B.; Levine, M.; Bell, N.H.; Mangelsdorf, D.; Russell, D. Genetic evidence that the human CYP2R1 enzyme is a key vitamin D 25-hydroxylase. Proc. Natl. Acad. Sci. USA 2004, 101, 7711–7715. [Google Scholar] [CrossRef] [Green Version]
- CYP2R1 Cytochrome P450 Family 2 Subfamily R Member 1 Homo Sapiens (Human)-Gene-NCBI. Available online: https://www.ncbi.nlm.nih.gov/gene?Db=gene&Cmd=DetailsSearch&Term=120227 (accessed on 12 August 2021).
- Roizen, J.D.; Levine, M.A. The role of genetic variation in CYP2R1, the principal vitamin D 25-hydroxylase, in vitamin D homeostasis. In Vitamin D: Volume 2: Health, Disease and Therapeutics; Martin, H., Roger, B., Edward, G., David, G., Eds.; Academic Press: Cambridge, UK, 2017; Volume 2, pp. 303–315. [Google Scholar]
- Osadnik, T.; Osadnik, K.; Pawlas, N.; Strzelczyk, J.; Kasperczyk, J.; Poloński, L.; Gąsior, M. Metabolic and genetic profiling of young adults with and without a family history of premature coronary heart disease (MAGNETIC). Study design and methodology. Arch. Med. Sci. 2019, 15, 590–597. [Google Scholar] [CrossRef]
- Rausch, T.; Fritz, M.H.-Y.; Untergasser, A.; Benes, V. Tracy: Basecalling, alignment, assembly and deconvolution of sanger chromatogram trace files. BMC Genom. 2020, 21, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bcftools by Samtools. Available online: https://samtools.github.io/bcftools/ (accessed on 12 August 2021).
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.S.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol. 2016, 17, 1–14. [Google Scholar] [CrossRef] [Green Version]
- GeneStudio Pro. Available online: https://genestudio-pro.software.informer.com/ (accessed on 12 August 2021).
- Ng, P.C. SIFT: Predicting amino acid changes that affect protein function. Nucleic Acids Res. 2003, 31, 3812–3814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pathogenicity Predictions. Available online: https://m.ensembl.org/info/genome/variation/prediction/protein_function.html (accessed on 24 September 2021).
- Adzhubei, I.A.; Schmidt, S.; Peshkin, L.; Ramensky, V.E.; Gerasimova, A.; Bork, P.; Kondrashov, A.S.; Sunyaev, S.R. A method and server for predicting damaging missense mutations. Nat. Methods 2010, 7, 248–249. [Google Scholar] [CrossRef] [Green Version]
- Jaganathan, K.; Panagiotopoulou, S.K.; McRae, J.F.; Darbandi, S.F.; Knowles, D.; Li, Y.I.; Kosmicki, J.A.; Arbelaez, J.; Cui, W.; Schwartz, G.B.; et al. Predicting Splicing from Primary Sequence with Deep Learning. Cell 2019, 176, 535–548.e24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.-H.; Yang, S.; Lim, K.-H.; Ko, E.; Jang, H.-J.; Kang, M.; Suh, P.-G.; Joo, J.-Y. Prediction of Alzheimer’s disease-specific phospholipase c gamma-1 SNV by deep learning-based approach for high-throughput screening. Proc. Natl. Acad. Sci. USA 2021, 118, e2011250118. [Google Scholar] [CrossRef] [PubMed]
- Chr11: 14.88M-14.89M-Genome Data Viewer-NCBI. Available online: https://www.ncbi.nlm.nih.gov/genome/gdv/browser/nucleotide/?id=NM_024514.5 (accessed on 3 December 2021).
- PDBe-KB Protein Pages. Available online: https://www.ebi.ac.uk/pdbe/pdbe-kb/proteins/Q6VVX0 (accessed on 12 September 2021).
- Amino Acids-Arginine R (Arg). Available online: http://www.biology.arizona.edu/biochemistry/problem_sets/aa/Arginine.html (accessed on 12 September 2021).
- Berg, J.M.; Tymoczko, J.L.; Stryer, L. Amino Acids Are Encoded by Groups of Three Bases Starting from a Fixed Point; W. H. Freeman and Company: New York, NY, USA, 2002. [Google Scholar]
- Schneider, F. Die Funktion des Arginins in den Enzymen. Naturwissenschaften 1978, 65, 376–381. [Google Scholar] [CrossRef]
- Amino Acids–Lysine. Available online: http://www.biology.arizona.edu/biochemistry/problem_sets/aa/Lysine.html (accessed on 12 September 2021).
- Li, L.; Vorobyov, I.; Allen, T.W. The Different Interactions of Lysine and Arginine Side Chains with Lipid Membranes. J. Phys. Chem. B 2013, 117, 11906–11920. [Google Scholar] [CrossRef]
- Sokalingam, S.; Raghunathan, G.; Soundrarajan, N.; Lee, S.-G. A Study on the Effect of Surface Lysine to Arginine Mutagenesis on Protein Stability and Structure Using Green Fluorescent Protein. PLoS ONE 2012, 7, e40410. [Google Scholar] [CrossRef] [Green Version]
- Musafia, B.; Buchner, V.; Arad, D. Complex Salt Bridges in Proteins: Statistical Analysis of Structure and Function. J. Mol. Biol. 1995, 254, 761–770. [Google Scholar] [CrossRef]
- Matsutani, M.; Hirakawa, H.; Nishikura, M.; Soemphol, W.; Ali, I.A.I.; Yakushi, T.; Matsushita, K. Increased number of Arginine-based salt bridges contributes to the thermotolerance of thermotolerant acetic acid bacteria, Acetobacter tropicalis SKU1100. Biochem. Biophys. Res. Commun. 2011, 409, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Nussinov, R. Salt bridge stability in monomeric proteins. J. Mol. Biol. 1999, 293, 1241–1255. [Google Scholar] [CrossRef]
- Strub, C.; Alies, C.; Lougarre, A.; Ladurantie, C.; Czaplicki, J.; Fournier, D. Mutation of exposed hydrophobic amino acids to arginine to increase protein stability. BMC Biochem. 2004, 5, 9. [Google Scholar] [CrossRef] [Green Version]
- Chan, C.-H.; Yu, T.-H.; Wong, K.-B. Stabilizing Salt-Bridge Enhances Protein Thermostability by Reducing the Heat Capacity Change of Unfolding. PLoS ONE 2011, 6, e21624. [Google Scholar] [CrossRef] [Green Version]
- Dong, A.N.; Tan, B.H.; Pan, Y.; Ong, C.E. The CYP2R1 Enzyme: Structure, Function, Enzymatic Properties and Genetic Polymorphism. J. Pharm. Pharm. Sci. 2021, 24, 94–112. [Google Scholar] [CrossRef]
- Al Mutair, A.N.; Nasrat, G.H.; Russell, D. Mutation of the CYP2R1 Vitamin D 25-Hydroxylase in a Saudi Arabian Family with Severe Vitamin D Deficiency. J. Clin. Endocrinol. Metab. 2012, 97, E2022–E2025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molin, A.; Wiedemann, A.; Demers, N.; Kaufmann, M.; Cao, J.D.; Mainard, L.; Dousset, B.; Journeau, P.; Abeguile, G.; Coudray, N.; et al. Vitamin D-Dependent Rickets Type 1B (25-Hydroxylase Deficiency): A Rare Condition or a Misdiagnosed Condition? J. Bone Miner. Res. 2017, 32, 1893–1899. [Google Scholar] [CrossRef]
- Ramos-Lopez, E.; Brück, P.; Jansen, T.; Herwig, J.; Badenhoop, K. CYP2R1 (vitamin D 25-hydroxylase) gene is associated with susceptibility to type 1 diabetes and vitamin D levels in Germans. Diabetes/Metab. Res. Rev. 2007, 23, 631–636. [Google Scholar] [CrossRef]
- Duan, L.; Xue, Z.; Ji, H.; Zhang, D.; Wang, Y. Effects of CYP2R1 gene variants on vitamin D levels and status: A systematic review and meta-analysis. Gene 2018, 678, 361–369. [Google Scholar] [CrossRef]
- Lafi, Z.M.; Irshaid, Y.M.; El-Khateeb, M.; Ajlouni, K.M.; Hyassat, D. Association of rs7041 and rs4588 Polymorphisms of the Vitamin D Binding Protein and the rs10741657 Polymorphism of CYP2R1 with Vitamin D Status Among Jordanian Patients. Genet. Test. Mol. Biomark. 2015, 19, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Slater, N.A.; Rager, M.L.; Havrda, D.E.; Harralson, A.F. Genetic Variation in CYP2R1 and GC Genes Associated With Vitamin D Deficiency Status. J. Pharm. Pr. 2017, 30, 31–36. [Google Scholar] [CrossRef]
- Nissen, J.; Rasmussen, L.B.; Ravn-Haren, G.; Andersen, E.W.; Hansen, B.; Andersen, R.; Mejborn, H.; Madsen, K.H.; Vogel, U. Common Variants in CYP2R1 and GC Genes Predict Vitamin D Concentrations in Healthy Danish Children and Adults. PLoS ONE 2014, 9, e89907. [Google Scholar] [CrossRef] [Green Version]
- Hassanein, S.I.; Abu El Maaty, M.A.; Sleem, H.M.; Gad, M.Z. Triangular relationship between single nucleotide polymorphisms in the CYP2R1 gene (rs10741657 and rs12794714), 25-hydroxyvitamin d levels, and coronary artery disease incidence. Biomarkers 2014, 19, 488–492. [Google Scholar] [CrossRef]
- Saponaro, F.; Saba, A.; Zucchi, R. An Update on Vitamin D Metabolism. Int. J. Mol. Sci. 2020, 21, 6573. [Google Scholar] [CrossRef]
- Morris, H.A. Vitamin D Metabolism and Molecular Modes of Action: New Insights into Vitamin D Activities. Med. Res. J. 2014, 2, 1–5. [Google Scholar]
- Jones, G.; Prosser, D.E.; Kaufmann, M. Cytochrome P450-mediated metabolism of vitamin D. J. Lipid Res. 2014, 55, 13–31. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; DeLuca, H.F. Vitamin D 25-hydroxylase–Four decades of searching, are we there yet? Arch. Biochem. Biophys. 2012, 523, 30–36. [Google Scholar] [CrossRef]
- Gupta, R.P.; He, Y.A.; Patrick, K.S.; Halpert, J.R.; Bell, N.H. CYP3A4 Is a Vitamin D-24- and 25-Hydroxylase: Analysis of Structure Function by Site-Directed Mutagenesis. J. Clin. Endocrinol. Metab. 2005, 90, 1210–1219. [Google Scholar] [CrossRef] [Green Version]
- Aiba, I.; Yamasaki, T.; Shinki, T.; Izumi, S.; Yamamoto, K.; Yamada, S.; Terato, H.; Ide, H.; Ohyama, Y. Characterization of rat and human CYP2J enzymes as Vitamin D 25-hydroxylases. Steroids 2006, 71, 849–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergadà, L.; Pallares, J.; Arcidiacono, M.V.; Cardus, A.; Santacana, M.; Valls, J.; Cao, G.; Fernàndez, E.; Dolcet, X.; Dusso, A.S.; et al. Role of local bioactivation of vitamin D by CYP27A1 and CYP2R1 in the control of cell growth in normal endometrium and endometrial carcinoma. Lab. Investig. 2014, 94, 608–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.G.; Ochalek, J.T.; Kaufmann, M.; Jones, G.; DeLuca, H.F. CYP2R1 is a major, but not exclusive, contributor to 25-hydroxyvitamin D production in vivo. Proc. Natl. Acad. Sci. USA 2013, 110, 15650–15655. [Google Scholar] [CrossRef] [Green Version]
- Putscher, E.; Hecker, M.; Fitzner, B.; Lorenz, P.; Zettl, U. Principles and Practical Considerations for the Analysis of Disease-Associated Alternative Splicing Events Using the Gateway Cloning-Based Minigene Vectors pDESTsplice and pSpliceExpress. Int. J. Mol. Sci. 2021, 22, 5154. [Google Scholar] [CrossRef] [PubMed]
- Sanz, D.J.; Hollywood, J.A.; Scallan, M.F.; Harrison, P.T. Cas9/gRNA targeted excision of cystic fibrosis-causing deep-intronic splicing mutations restores normal splicing of CFTR mRNA. PLoS ONE 2017, 12, e0184009. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Parameters | 25(OH)D > 30 ng/mL n (%) 43 (100%) | 25(OH)D < 10 ng/mL n (%) 46 (100) | p-Value U Mann–Whitney/2 |
|---|---|---|---|
| Physical activity low moderate high | 11 (25.58) 20 (46.51) 12 (27.91) | 17 (36.96) 22 (47.82) 7 (15.22) | 0.273 |
| BMI (kg/m2) | 23.42 (22.20; 25.15) | 24.62 (22.43; 26.20) | 0.440 |
| FH of DM2 | 10 (23.26) | 7 (15.22) | 0.335 |
| FH of P-CAD | 27 (62.79) | 29 (63.04) | 0.980 |
| WC (cm) | 81.25 (75.00; 89.00) | 86.00 (76.00; 92.00) | 0.535 |
| WHR | 0.86 (0.82; 0.90) | 0.85 (0.82; 0.89) | 0.932 |
| VAI | 0.81 (0.65; 1.11) | 1.12 (0.84; 1.33) | 0.024 |
| SBP (mmHg) | 123.00 (120.00; 128.00) | 126.71 (124.00; 135.00) | 0.044 |
| DBP (mmHg) | 75.00 (72.00; 79.00) | 81.50 (77.46; 90.00) | 0.002 |
| Smoking, n (%) smoker non-smoker smoked in the past | 9 (20.93) 27 (62.79) 7 (16.28) | 12 (26.09) 28 (60.87) 6 (13.04) | 0.876 |
| IMT (mm) right (average) left (average) | 0.54 (0.52; 0.57) 0.55 (0.52; 0.60) | 0.54 (0.50; 0.57) 0.54 (0.50; 0.59) | 0.813 0.576 |
| Cholesterol (mmol/L) | 4.80 (4.23; 5.04) | 4.56 (4.08; 4.95) | 0.812 |
| LDL (mmol/L) | 2.73 (2.50; 3.22) | 2.71 (2.39; 3.02) | 0.921 |
| HDL (mmol/L) | 1.52 (1.33; 1.73) | 1.34 (1.16; 1.48) | 0.004 |
| HDL (%) | 34.00 (29.00; 39.00) | 29.00 (25.00; 33.00) | 0.047 |
| TG (mmol/L) | 0.83 (0.67; 1.08) | 1.04 (0.77; 1.23) | 0.063 |
| Lp(a) (nmol/L) | 10.00 (5.00; 15.00) | 15.00 (7.00; 23.00) | 0.579 |
| Apo A (g/L) | 1.60 (1.48; 1.72) | 1.49 (1.42; 1.63) | 0.071 |
| Apo B (g/L) | 0.88 (0.76; 0.94) | 0.87 (0.77; 0.95) | 0.850 |
| hsCRP (mg/dl) | 0.99 (0.64; 1.24) | 1.15 (0.88; 1.40) | 0.278 |
| FG (mmol/L) | 4.90 (4.70; 5.10) | 4.95 (4.80; 5.10) | 0.446 |
| HBA1C (%) | 5.00 (4.90; 5.10) | 5.00 (4.90; 5.10) | 0.876 |
| LDH (U/L) | 159.00 (153.00; 170.00) | 168.50 (160.00; 173.00) | 0.177 |
| Bilirubin (µmol/L) | 8.40 (6.20; 10.80) | 9.60 (6.90; 11.40) | 0.364 |
| ALAT (U/L) | 16.00 (14.00; 19.00) | 20.00 (15.00; 25.00) | 0.347 |
| ASPAT (U/L) | 18.00 (17.00; 19.00) | 19.00 (17.00; 20.00) | 0.550 |
| GGT (U/L) | 16.00 (13.00; 19.00) | 24.50 (15.00; 31.00) | 0.035 |
| Fibrinogen (mg/dl) | 249.00 (221.00; 294.00) | 261.00 (242.00; 282.00) | 0.496 |
| ALP (U/L) | 57.00 (55.00; 64.00) | 59.00 (55.00; 66.00) | 0.675 |
| 25(OH)D (ng/mL) | 45.00 (39.00; 47.00) 43.40 ± 8.63 | 7.00 (7.00; 8.00) 7.13 ± 2.16 | <0,001 |
| Total calcium (mmol/L) | 2.42 (2.38; 2.46) | 2.40 (2.35; 2.42) | 0.169 |
| Phosphorus (mmol/L) | 1.08 (1.00; 1.16) | 1.06 (1.00; 1.10) | 0.297 |
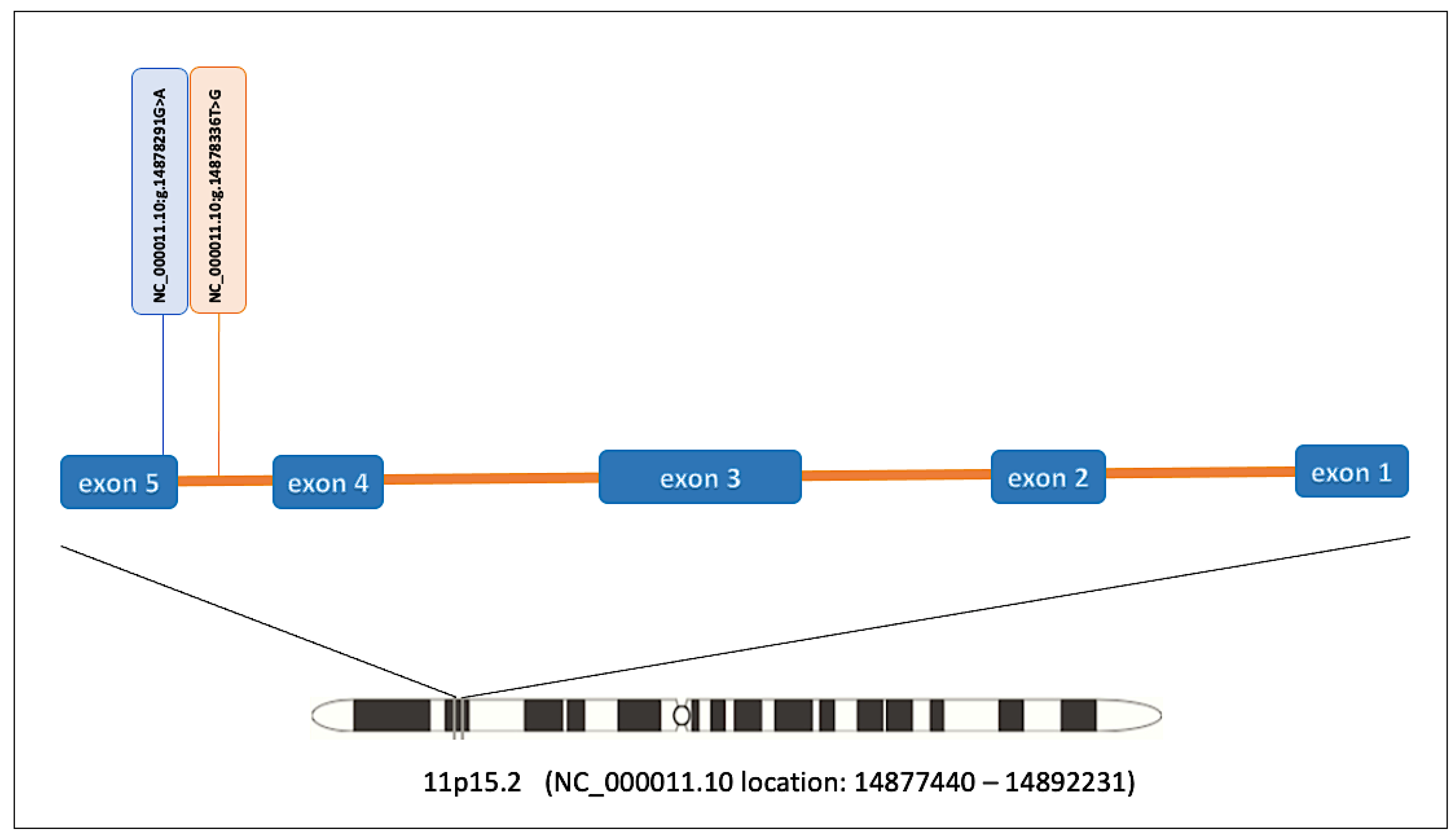
| Variant Description | ||
|---|---|---|
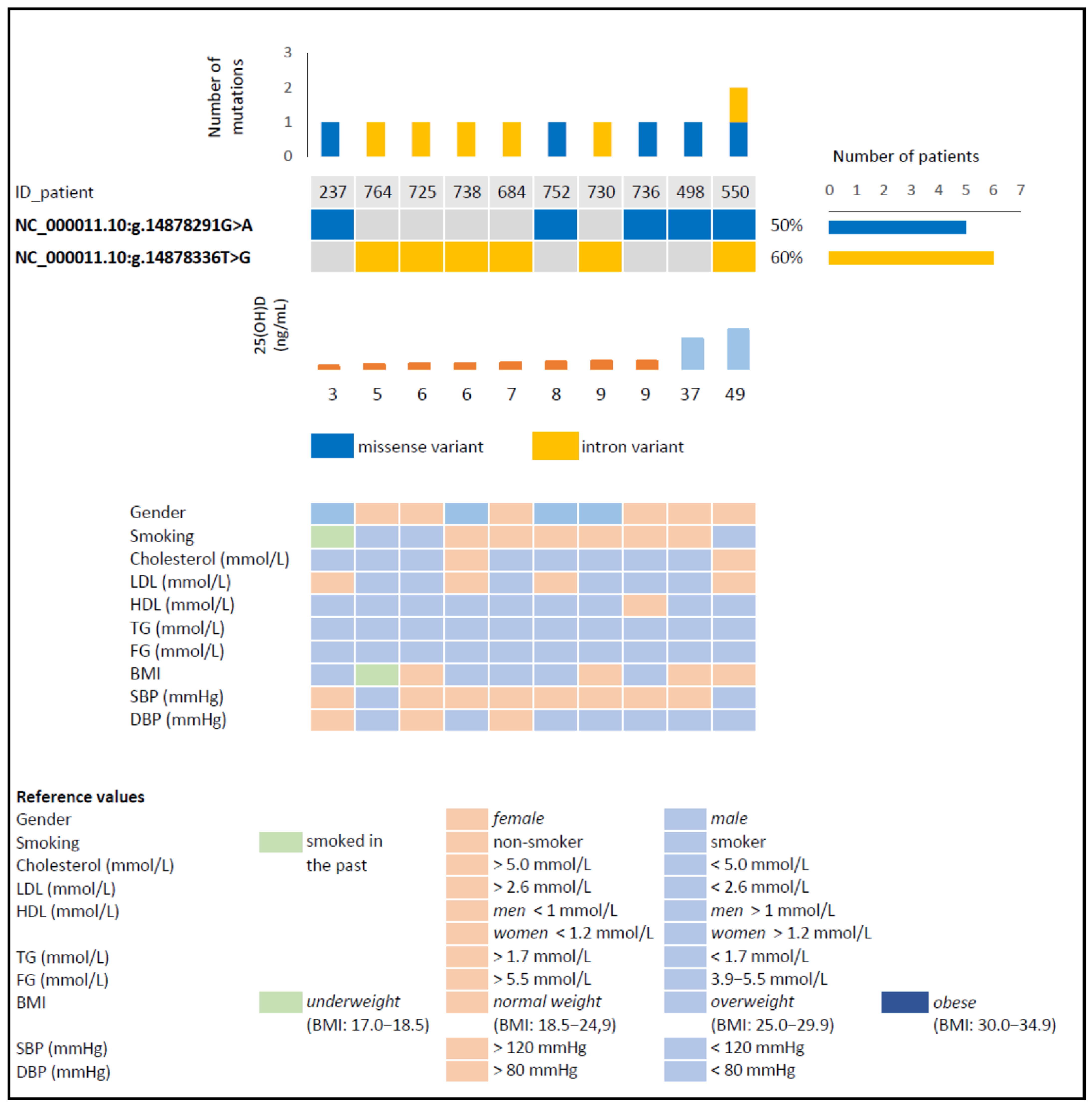
| Variant (GRCh38) | NC_000011.10:g.14878291G>A | NC_000011.10:g.14878336T>G |
| NM_024514.5 | c.1337G>A (p.Arg446Lys) | c.1331-39T>G |
| Variant type | Missense variant | Intron variant |
| Bioinformatic Analysis | ||
| SIFT score | 0.0 (deleterious) | - |
| PolyPhen score | 1.0 (probably damaging) | - |
| SpliceAI DS (bp) | ||
| acceptor loss | 0.25 (6 bp) | 0.04 (−39 bp) |
| donor loss | 0.00 (−36 bp) | 0.00 (2 bp) |
| acceptor gain | 0.00 (−31 bp) | 0.00 (4 bp) |
| donor gain | 0.00 (47 bp) | 0.00 (−2 bp) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fronczek, M.; Strzelczyk, J.K.; Biernacki, K.; Salatino, S.; Osadnik, T.; Ostrowska, Z. New Variants of the Cytochrome P450 2R1 (CYP2R1) Gene in Individuals with Severe Vitamin D-Activating Enzyme 25(OH)D Deficiency. Biomolecules 2021, 11, 1867. https://doi.org/10.3390/biom11121867
Fronczek M, Strzelczyk JK, Biernacki K, Salatino S, Osadnik T, Ostrowska Z. New Variants of the Cytochrome P450 2R1 (CYP2R1) Gene in Individuals with Severe Vitamin D-Activating Enzyme 25(OH)D Deficiency. Biomolecules. 2021; 11(12):1867. https://doi.org/10.3390/biom11121867
Chicago/Turabian StyleFronczek, Martyna, Joanna Katarzyna Strzelczyk, Krzysztof Biernacki, Silvia Salatino, Tadeusz Osadnik, and Zofia Ostrowska. 2021. "New Variants of the Cytochrome P450 2R1 (CYP2R1) Gene in Individuals with Severe Vitamin D-Activating Enzyme 25(OH)D Deficiency" Biomolecules 11, no. 12: 1867. https://doi.org/10.3390/biom11121867