Biotechnological Screening of Microalgal and Cyanobacterial Strains for Biogas Production and Antibacterial and Antifungal Effects

Abstract
:1. Introduction
1.1. Microalgae Biomass as Substrate in Biogas Plants
1.2. Distribution of Compounds with Bioactive Activities
1.3. Aims of the Study
2. Materials and Methods
Organisms and Culture Conditions
3. Analytical Methods
3.1. Growth Analysis
3.2. Determination of Dry Mass (DM) and Organic Dry Mass (ODM)
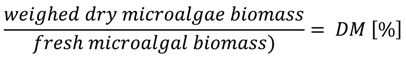
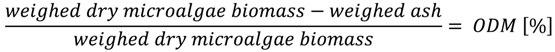
3.3. Measuring Biogas Production
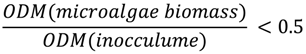
3.4. Productivity of Biogas
3.5. Extraction of Bioactive Compounds
3.6. Bacterial and Fungal Organisms Used and Culture Conditions
3.7. Determination of Optimal Antibiotic Concentration
4. Results and Discussion
4.1. Biomass Production of Microalgal and Cyanobacterial Strains
4.2. Screening for Microalgal and Cyanobacterial Biogas Production and Productivity

| Division/Class | Genus/Species | SAG Strain No | Culture Medium | Biogas yield (BY) | Biomass dry mass (DW) | Biogas productivity (P) |
|---|---|---|---|---|---|---|
| Chlorophyta | ||||||
| Chlorophyceae | Desmodesmus. sp. | 2389 | 3NBBM | 456 | 3.15±0.21 | 1436.40 |
| D. armatus | 276–4e | 3NBBM | 440 | 3.00±0.43 | 1318.53 | |
| D. armatus | 276–4d | 3NBBM | 518 | 2.55±0.75 | 1319.17 | |
| Haematococcus. pluvialis | 44.96 | 3NBBM | 300 | 2.03±0.27 | 610.00 | |
| H. pluvialis | 192.80 | 3NBBM | 413 | 3.05±0.77 | 1261.03 | |
| Scenedesmus. sp. | Kiel* | 3NBBM | 373 | 2.00±0.07 | 746.00 | |
| S. obliquus | 276–1 | 3NBBM | 433 | 1.27±0.22 | 548.47 | |
| Chlorodendrophyceae | Tetraselmis striata | 41.85 | 3NBBM | 385 | 2.91±0.16 | 1121.63 |
| Trebouxiophyceae | Chlorella sorokiniana | 211–8k | 3NBBM | 320 | 1.43±0.11 | 456.53 |
| C. vulgaris | 211–11b | 3NBBM | 410 | 4.85±0.15 | 1987.13 | |
| C. vulgaris | 211–1e | 3NBBM | 510 | 3.81±0.07 | 1944.80 | |
| C. vulgaris | 211–8l | 3NBBM | 436 | 3.00±0.05 | 1306.55 | |
| C. vulgaris | 211–11f | 3NBBM | 428 | 3.88±0.13 | 1660.64 | |
| C. vulgaris | 211–8m | 3NBBM | 463 | 3.64±0.06 | 1683.78 | |
| C. vulgaris | 211–11s | 3NBBM | 364 | 3.86±0.07 | 1406.25 | |
| C. vulgaris | 2.80 | 3NBBM | 397 | 3.66±0.10 | 1454.34 | |
| C. vulgaris | 9.88 | 3NBBM | 431 | 3.70±0.23 | 1594.70 | |
| Chloroidium. saccharophilum | 2149 | 3NBBM | 304 | 1.63±0.08 | 495.52 | |
| C. saccharophilum | 56.87 | 3NBBM | 303 | 2.93±0.11 | 887.79 | |
| Geminella. sp. | 57.90 | 3NBBM | 448 | 2.19±0.39 | 981.12 | |
| Geminella sp. | 49.80 | 3NBBM | 440 | 1.92±0.15 | 843.33 | |
| G. minor | 22.88 | 3NBBM | 376 | 2.75±0.14 | 1032.74 | |
| G. terricola | 20.91 | 3NBBM | 316 | 1.42±0.35 | 449.77 | |
| Nannochloris. sp. | 251–2 | 3NBBM | 494 | 2.41±0.18 | 1192.19 | |
| Stichococcus. sp. | 2118 | 3NBBM | 576 | 2.40±0.06 | 1384.32 | |
| Trebouxia. showmanii | 2009 | 3NBBM | 461 | 1.18±0.18 | 542.44 | |
| unclear taxonomy | Chlorella stigmatophora | 9.86 | 3NBBM | 380 | 4.63±0.42 | 1761.93 |
| Heterokontophyta | ||||||
| Eustigmatophyceae | Chloridella. neglecta | 48.84 | 3NBBM | 380 | 2.82±0.18 | 1071.60 |
| Eustigmatos. magnus | 36.89 | 3NBBM | 405 | 2.66±0.13 | 1077.30 | |
| Monodus. unipapilla | 8.83 | 3NBBM | 217 | 1.75±0.07 | 379.03 | |
| Nannochloropsis. salina | 40.85 | ASM30 | 202 | 3.35±0.05 | 677.37 | |
| Vischeria. punctata | 887–1 | 3NBBM | 464 | 2.26±0.06 | 1047.09 | |
| Xanthophyceae | Botrydiopsis intercedens | 806–3 | 3NBBM | 235 | 2.18±0.05 | 512.30 |
| Bumilleriopsis. filiformis | 809–2 | 3NBBM | 353 | 2.24±0.04 | 789.54 | |
| Heterococcus. viridis | 835–7 | 3NBBM | 411 | 1.72±0.08 | 706.92 | |
| Sphaerosorus. composita | 53.91 | 3NBBM | 399 | 1.00±0.04 | 399.00 | |
| Xanthonema. debile | 2289 | 3NBBM | 421 | 1.73±0.14 | 726.93 | |
| X. sessile | 2193 | 3NBBM | 238 | 2.28±0.05 | 541.85 | |
| Streptophyta | ||||||
| Klebsormidiophyceae | Klebsormidium. sp. | Namibia 5* | 3NBBM | 308 | 2.11±0.03 | 649.88 |
| Rhodophyta | ||||||
| Porphyridiophyceae | Porphyridium. purpureum | 1380–1d | ASM15 | 264 | 3.98±0.03 | 1050.72 |
| P. sordidum | 44.94 | ASM15 | 236 | 4.12±0.03 | 973.11 | |
| Cyanobacteria | ||||||
| Cyanophyceae | Arthrospira. platensis | 86.79 | Spirul | 376 | 2.93±0.09 | 1102.93 |
| A. platensis | 21.99 | Spirul | 395 | 3.95±0.06 | 1560.25 | |
| A. maxima | 84.79 | Spirul | 260 | 5.72±0.18 | 1488.07 | |
| Oscillatoria sp. | 76.79 | Spirul | 356 | 3.31±0.17 | 1178.36 |
4.3. Screening for Microalgal and Cyanobacterial Bioactivities Against Bacteria and Fungi
| Division/Class | Genus/Species | SAG Strain No | Culture Medium | Axenic culture | Methanol extract | Pellet | supernatant | Degree of inhibition in % |
|---|---|---|---|---|---|---|---|---|
| Chlorophyta | ||||||||
| Chlorophyceae | Bracteacoccus sp. | 2137 | 3NBBM | yes | P | - | - | 40 |
| B. bullatus | 2032 | 3NBBM | yes | - | - | - | ||
| Chromochloris cinnabarinus | 221–2 | 3NBBM | yes | - | - | - | ||
| Coleochlamys oleifera | 6.90 | 3NBBM | yes | - | - | B | 5 | |
| Desmodesmus. sp.b | 2389 | 3NBBM | yes | - | - | - | ||
| D. armatusb | 276–4e | 3NBBM | yes | E | - | - | 10 | |
| Haematococcus pluvialisb | 44.96 | 3NBBM | yes | - | B | - | 20 | |
| H. pluvialis | 34–1a | 3NBBM | yes | - | B | - | 20 | |
| H. pluvialis | 34–1b | 3NBBM | yes | - | - | - | ||
| Neospongiococcum gelatinosum | 64.80 | 3NBBM | yes | B | - | - | 40 | |
| Pectinodesmus. pectinatus | 2003 | 3NBBM | yes | - | - | - | ||
| Scenedesmus sp. | 2125 | 3NBBM | no | - | - | - | ||
| Scenedesmus sp.b | Kiel* | 3NBBM | yes | - | - | - | ||
| S. acuminatus | 38.81 | 3NBBM | yes | B | B | B | 40/20/20 | |
| S. costatus | 46.88 | 3NBBM | yes | B | B | B | 20/20/20 | |
| S. falcatus | 2.81 | 3NBBM | yes | B | B | B | 40/20/20 | |
| S. ovalternus | 52.80 | 3NBBM | yes | B | B | B | 20/20/20 | |
| S. obliquusb | 276–1 | 3NBBM | yes | - | - | - | ||
| S. pectinatus | 2003 | 3NBBM | yes | B | - | B | 40/20 | |
| S. rubescens | 5.95 | 3NBBM | yes | - | - | - | ||
| S. wisconsinensis | 22.81 | 3NBBM | yes | B | B | B | 40/40/40 | |
| Scotiellopsis. oocystiformis | 277–1 | 3NBBM | yes | - | - | - | ||
| Tetraedron caudatum | 2092 | 3NBBM | yes | B | - | - | 20 | |
| T. minimum | 44.81 | 3NBBM | yes | B | - | - | 20 | |
| Chlorodendrophyceae | Tetraselmis sp. | 3.98 | 3NBBM | yes | B | B | B | 40/40/20 |
| T. striatab | 41.85 | 3NBBM | yes | B | B | B | 20/20/20 | |
| T. tetrathele | 161–2b | 3NBBM | yes | - | - | - | ||
| Trebouxiophyceae | Chlorella mirabilis | 38.88 | 3NBBM | yes | - | - | - | |
| C. sorokiniana | 211–31 | 3NBBM | yes | - | - | - | ||
| C. sorokiniana | 211–32 | 3NBBM | yes | - | - | E | 40 | |
| C. sorokinianab | 211–8k | 3NBBM | yes | - | - | - | ||
| C. vulgarisb | 211–11b | 3NBBM | yes | - | - | - | ||
| Chloroidium. ellipsoideum | 2140 | 3NBBM | yes | - | - | - | ||
| C. angustoellipsoideum | 2041 | 3NBBM | yes | - | - | - | ||
| C. saccharophilumb | 2149 | 3NBBM | yes | - | - | B | 20 | |
| C. saccharophilumb | 56.87 | 3NBBM | yes | - | - | - | ||
| Coccomyxa. sp. | 2391 | 3NBBM | yes | - | - | - | ||
| C. elongata | 216–3a | 3NBBM | yes | P | - | - | 5 | |
| Coenocystis. sp. | LS5-R4* | 3NBBM | yes | - | - | - | ||
| Diplosphaera. mucosa | 48.86 | 3NBBM | yes | - | - | - | ||
| Geminella sp.b | 57.90 | 3NBBM | yes | E/B | - | - | 5/10 | |
| G. minor | 10.91 | 3NBBM | yes | P | - | - | 20 | |
| G. terricolab | 20.91 | 3NBBM | yes | P | - | - | 10 | |
| Heterochlorella luteoviridis | 211–2a | 3NBBM | yes | - | - | - | ||
| H. luteoviridis | 211–3 | 3NBBM | yes | - | - | C | + | |
| Muriella. terrestris | 2390 | 3NBBM | yes | - | - | - | ||
| Nannochloris eucaryotum | 55.87 | 3NBBM | no | - | - | - | ||
| N. normandinae | 9.82 | 3NBBM | no | - | - | - | ||
| Neocystis. mucosa | 40.88 | 3NBBM | yes | - | - | - | ||
| Pabia signiensis | 7.90 | 3NBBM | yes | - | - | B | 5 | |
| Pseudochlorella. pringsheimii | 211–1a | 3NBBM | yes | - | - | - | ||
| Raphidonema sp. | LS11-R7A* | 3NBBM | yes | P | - | - | 5 | |
| Stichococcus sp. | 249.80 | 3NBBM | yes | - | - | - | ||
| Stichococcus sp. | 2059 | 3NBBM | yes | - | - | - | ||
| S. bacillaris | 379–1b | 3NBBM | yes | - | B | - | 40 | |
| S. bacillaris | 379–2 | 3NBBM | yes | - | - | - | ||
| S. deasonii | 2139 | 3NBBM | yes | - | - | |||
| Chlorophyta | ||||||||
| Trebouxiophyceae | Trebouxia. aggregata | 219–1d | 3NBBM | yes | - | - | - | |
| T. asymmetrica | 48.88 | 3NBBM | yes | - | - | - | ||
| T. showmaniib | 2009 | 3NBBM | yes | - | - | - | ||
| unclear taxonomy | Chlorella stigmatophorab | 9.86 | 3NBBM | no | - | - | - | |
| Chlorella sp. | 15.93 | 3NBBM | yes | - | - | - | ||
| Heterokontophyta | ||||||||
| Eustigmatophyceae | Chloridella. neglecta | 7.88 | 3NBBM | yes | P | - | - | 25 |
| C. simplex | 51.91 | 3NBBM | yes | - | - | - | ||
| Eustigmatos magnusb | 36.89 | 3NBBM | yes | - | - | - | ||
| Monodus. unipapillab | 8.83 | 3NBBM | yes | - | - | - | ||
| Nannochloropsis. gaditana | 2.99 | ASM15 | no | - | - | - | ||
| N. salinab | 40.85 | ASM30 | yes | - | - | - | ||
| Vischeria. helvetica | 876–1 | 3NBBM | yes | - | B | - | 40 | |
| V. stellata | 33.83 | 3NBBM | yes | - | - | - | ||
| V. stellata | 887–2 | 3NBBM | yes | - | B | - | 20 | |
| Xanthophyceae | Botrydiopsis alpina | CCALA 217* | 3NBBM | yes | - | - | - | |
| B. intercedensb | 806–3 | 3NBBM | yes | - | - | - | ||
| Bumilleriopsis. filiformisb | 809–2 | 3NBBM | yes | P | - | - | 20 | |
| Sphaerosorus compositab | 53.91 | 3NBBM | yes | - | - | - | ||
| Xanthonema. exile | 2286 | 3NBBM | no | P | - | 5 | ||
| X. sessileb | 2193 | 3NBBM | no | - | - | - | ||
| Streptophyta | - | - | - | |||||
| Klebsormidiophyceae | Klebsormidium sp.b | Namibia 5* | 3NBBM | yes | B | B | 40/70 | |
| Zygnematophyceae | Fottea pyrenoidosa | 1.88 | 3NBBM | yes | - | - | - | |
| Rhodophyta | - | - | - | |||||
| Porphyridiophyceae | Porphyridium. purpureum | 113.79 | ASM15 | no | E | - | - | 40 |
| P. purpureum | 1380–1a | ASM15 | yes | - | - | - | ||
| P. purpureum | 1380–1b | ASM15 | yes | - | - | - | ||
| P. purpureum | 1380–1c | ASM15 | yes | E/B | - | - | 60/30 | |
| P. purpureumb | 1380–1d | ASM15 | yes | - | - | C | + | |
| P. purpureum | 1380–1f | ASM15 | yes | E/B | - | - | 40/10 | |
| P. sordidumb | 44.94 | ASM15 | no | - | E/B/P | - | 40/20/40 | |
| Rhodellophyceae | Rhodella. maculata | 45.85 | ASM15 | no | B | B | B | 20/40/40 |
| R. violaceae | 115.79 | ASM15 | yes | B | B | - | 40/40 | |
| Cyanobacteria | - | - | - | |||||
| Cyanophyceae | Arthrospira platensis | 85.79 | Spirul | yes | S | - | - | ++ |
| A. platensisb | 86.79 | Spirul | no | - | - | - | ||
| A. platensis | 257.80 | Spirul | yes | E/B | - | - | 40/5 | |
| A. platensisb | 21.99 | Spirul | yes | - | - | - | ||
| A. maximab | 84.79 | Spirul | no | S | - | - | + | |
| A. maxima | 49.88 | Spirul | no | - | - | - | ||
| Oscillatoria sp. | 50.96 | ASM30 | yes | - | - | - | ||
| Oscillatoria. sp.b | 76.79 | Spirul | no | B | B | B | 30/50/40 | |
| unrevised taxon | „Spirulina“ laxissima | 256.80 | Spirul | no | E | - | - | 40 |
Conclusions
Outlook
Acknowledgements
Conflicts of Interests
Author Conrtibutions
References
- Montzka, S.A.; Dlugokencky, E.J.; Butler, J.H. Non-CO2 greenhouse gases and climate change. Nature 2011, 476, 43–50. [Google Scholar] [CrossRef]
- Chynoweth, D.P.; Turick, C.E.; Owens, J.M.; Jerger, D.E.; Peck, M.W. Biochemical methane potential of biomass and waste feedstocks. Biomass Bioenergy 1993, 5, 95–111. [Google Scholar] [CrossRef]
- Amon, T.; Amon, B.; Kryvoruchko, V.; Zollitsch, W.; Mayer, K.; Gruber, L. Biogas production from maize and dairy cattle manure Influence of biomass composition on the methane yield. Agric. Ecosyst. Environ. 2007, 118, 173–182. [Google Scholar] [CrossRef]
- Johansson, T.; Kelly, H.; Reddy, A.K.N.; Williams, R.H. Renewable fuels and electricity for a growing world economy. In Renewable Energy Sources for Fuels and Electricity; Burnham, L., Ed.; Island Press: Washington, USA, 1993; p. 5. [Google Scholar]
- Lastella, G.; Testa, C.; Cornacchia, G.; Notornicola, M.; Voltasio, F.; Sharma, V.K. Anaerobic digestion of semi-solid organic waste: Biogas production and its purification. Energ. Convers. Manage. 2002, 43, 63–75. [Google Scholar] [CrossRef]
- Muyiiya, N.D.; Kasisira, L.L. Assessment of the Effect of Mixing Pig and Cow Dung on Biogas Yield. Agricultural Engineering International: The CIGR Journal 2009, 11. Manuscript PM 1329. [Google Scholar]
- Hanagata, N.; Takeuchi, T.; Fukuju, Y.; Barnes, D.J.; Karube, I. Tolerance of microalgae to high CO2 and high temperature. Phytochemistry 1992, 31, 3345–3348. [Google Scholar] [CrossRef]
- Yoo, C.; Choi, G.G.; Kim, S.C.; Oh, H.M. Ettlia sp. YC001 showing high growth rate and lipid content under high CO2. Bioresour. Technol. 2013, 127, 482–488. [Google Scholar] [CrossRef]
- Mudimu, O.; Rybalka, N.; Bauersachs, T.; Friedl, T.; Schulz, R. Influence of different CO2 concentrations on microalgae growth rate, α-tocopherol content and fatty acid composition. Geomicrobiol. J. 2014, unpublished work. [Google Scholar]
- Wang, B.; Li, Y.; Wu, N.; Lan, C.Q. CO2 biomitigation using microalgae. Appl. Microbiol. Biotechnol. 2008, 79, 707–718. [Google Scholar] [CrossRef]
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef]
- Chisti, Y. Biodiesel from microalgae beats bioethanol. Trends Biotechnol. 2008, 26, 126–131. [Google Scholar] [CrossRef]
- Golueke, C.; Oswald, W.J.; Gotaas, H.B. Anaerobic digestion of algae. Appl. Microbiol. 1957, 5, 47–55. [Google Scholar]
- Mussgnug, J.H.; Klassen, V.; Schlüter, A.; Kruse, O. Microalgae as substrates for fermentative biogas production in a combined biorefinery concept. J. Biotechnol. 2010, 150, 51–56. [Google Scholar]
- Ras, M.; Lardon, L.; Bruno, S.; Bernet, N.; Steyer, J.P. Experimental study on a coupled process of production and anaerobic digestion of Chlorella vulgaris. Bioresour. Technol. 2011, 102, 200–206. [Google Scholar] [CrossRef]
- Collet, P.; Hélias, A.; Lardon, L.; Ras, M.; Goy, R.A.; Steyer, J.P. Life-cycle assessment of microalgae culture coupled to biogas production. Bioresour. Technol. 2011, 102, 207–214. [Google Scholar] [CrossRef]
- Sialve, B.; Bernet, N.; Bernard, O. Anaerobic digestion of microalgae as a necessary step to make microalgal biodiesel sustainable. Biotechnol. Adv. 2009, 27, 409–416. [Google Scholar] [CrossRef]
- Courvalin, P. Transfer of antibiotic resistance genes between gram-positive and gram-negative bacteria. Antimicrob. Agents Chemother. 1994, 38, 1447–1451. [Google Scholar] [CrossRef]
- Pulz, O.; Gross, W. Valuable products from biotechnology of microalgae. Appl. Microbiol. Biotechnol. 2004, 65, 635–648. [Google Scholar] [CrossRef]
- Wehr, J.D.; Sheath, R.G. Freshwater habitats of algae: Freshwater algae of North America. In Ecology and Classification; Academic Press: New York, NY, USA, 2003; pp. 11–58. [Google Scholar]
- Hallmann, C.; Rüdrich, J.; Enseleit, M.; Friedl, T.; Hoppert, M. Microbial diversity on a marble monument: a case study. Environmental Earth Sciences 2011, 63, 1701–1711. [Google Scholar] [CrossRef]
- Harris, E.H. Introduction to Chlamydomonas and its laboratory use. In The Chlamydomonas Sourcebook, 2nd ed.; Harris, E.H., Stern, D.B., Witman, G.B., Eds.; Academic Press: San Diego, CA, USA, 2009; Volume 1, p. 444. [Google Scholar]
- Arp, G.; Bissett, A.; Brinkmann, N.; Cousin, S.; Beer, D.D.; Friedl, T.; Mohr, K.I.; Neu, T.R.; Reimer, A.; Shiraishi, F.; Stackebrandt, E.; Zippel, B. Tufa-forming biofilms of German karstwater streams: microorganisms, exopolymers, hydrochemistry and calcification. Geol. Soc. Spec. Publ. 2010, 336, 83–118. [Google Scholar] [CrossRef]
- Patterson, G.M.L.; Baker, K.K.; Baldwin, C.L.; Bolis, C.M.; Caplan, F.R.; Larsen, L.K.; Levine, I.A.; Moore, R.E.; Nelson, C.S.; Tschappat, K.D.; Tuang, G.D.; Boyd, M.R.; Cardellina, J.H.; Collins, R.P.; Gustafson, K.R.; Snader, K.M.; Weislow, O.S.; Lewin, R.A. Antiviral activity of cultured blue-green algae (Cyanophyta). J. Phycol. 1993, 29, 125–130. [Google Scholar] [CrossRef]
- Patterson, G.M.L.; Larsen, L.K.; Moore, R.E. Bioactive natural products from blue-green algae. J. Appl. Phycol. 1994, 6, 151–157. [Google Scholar] [CrossRef]
- Borowitzka, A.M. Microalgae as source of pharmaceuticals and biologically active compounds. J. Appl. Phycol. 1995, 7, 3–15. [Google Scholar] [CrossRef]
- Falch, B.S.; König, G.M.; Wright, A.D.; Sticher, O.; Angerhofer, C.K.; Pezzuto, J.M.; Bachmann, H. Biological activities of cyanobacteria: Evaluation of extracts and pure compounds. Planta. Med. 1995, 61, 321–328. [Google Scholar]
- Kulik, M.M. The potential for using cyanobacteria (blue-greenalgae) and algae in the biological control of plant pathogenic bacteria and fungi. European Journal of Plant Pathology 1995, 101, 585–599. [Google Scholar] [CrossRef]
- Schlegel, I.; Doan, N.T.; de Chazal, N.; Smith, G.D. Antibiotic activity of new cyanobacterial isolates from Australia and Asia against green algae and cyanobacteria. J. Appl. Phycol. 1999, 10, 471–479. [Google Scholar]
- Del Val, A.G.; Platas, G.; Basilio, A.; Cabella, A.; Gorrochategui, J.; Suay, I.; Vicente, F.; Portillo, E.; del Rio, M.J.; Reina, G.G.; Peláez, F. Screening of antimicrobial activities in red, green and brown macroalgae from Gran Canaria (Canary Islands, Spain). Int. Microbiol. 2001, 4, 35–40. [Google Scholar]
- Culture Collection of Algae at the Goettingen University (SAG), Germany. Available online: http://www.epsag.uni-goettingen.de (accessed on 10 October 2012).
- Culture Collection of Algal Laboratory (CCALA), Czech Republic. Available online: http://www.butbn.cas.cz/ccala/index.php (accessed on 05 July 2012).
- Starr, R.C.; Zeikus, J.A. UTEX – The Culture Collection of Algae at the University of Texas at Austin. J. Phycol. 1993, 29, 1–160. [Google Scholar]
- Aiba, S.; Ogawa, T. Assessment of Growth Yield of a Blue-green Alga, Spirulina platensis, in Axenic and Continuous Culture. J. Gen. Microbiol. 1977, 102, 179–182. [Google Scholar] [CrossRef]
- VDI 4630. Fermentation of Organic Materials—Characterisation of the Substrate, Sampling, Collection of Material Data, Fermentation Tests. Verlag des Vereins Deutscher Ingenieure: Berlin, Germany, 2008.
- German Collection of Microorganisms and Cell Cultures (DMSZ), Germany. Available online: Available online: https://www.dsmz.de (accessed on 11 May 2012).
- American Type Culture Collection (ATCC). Available online: http://www.lgcstandards-atcc.org (accessed on 10 May 2012).
- Cannell, R.J.P.; Owsianka, A.M.; Walker, J.M. Results of a large-scale screening programme to detect antibacterial activity from fresh water algae. British Phycological Journal 1988, 23, 41–44. [Google Scholar] [CrossRef]
- Degen, J.; Uebele, A.; Retze, A.; Schmid-Staiger, U.; Trösch, W. A novel airlift photobioreactor with baffles for improved light utilization through the flashing light effect. J. Biotechnol. 2000, 92, 89–94. [Google Scholar]
- Campo, J.A.; Moreno, J.; Rodrıguez, H.; Vargas, M.A.; Rivas, J.; Guerrero, M.G. Carotenoid content of chlorophycean microalgae: factors determining lutein accumulation in Muriellopsis. sp. (Chlorophyta). J. Biotechnol. 2000, 76, 51–59. [Google Scholar]
- Oswald, W.J.; Goluecke, C.G. Biological Transformation of Solar Energy. Adv. Appl. Microbiol. 1960, 2, 223–262. [Google Scholar] [CrossRef]
- Samson, R.; LeDuy, A. Influence of mechanical and thermochemical pretreatments on anaerobic digestion of Spirulina maxima algal biomass. Biotechnol. Lett. 1983, 5, 671–676. [Google Scholar] [CrossRef]
- Müller, J.; Friedl, T.; Hepperle, D.; Lorenz, M. Distinction between multiple isolates of Chlorella vulgaris (Chlorophyta, Trebouxiophyceae) and testing for conspecificity using amplified fragment length polymorphism and IST rDNA sequences. Journal of Phycology 2005, 41, 1236–1247. [Google Scholar] [CrossRef]
- Schwede, S.; Rehman, Z.U.; Gerber, M.; Theiss, C.; Span, R. Effects of thermal pretreatment on anaerobic digestion of Nannochloropsis salina biomass. Bioresour. Technol. 2013, 143, 505–511. [Google Scholar] [CrossRef]
- Van den Hoek, C.; Mann, D.G.; Jahns, H.M. Algae: An introduction to phycology, 2nd ed.Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Viola, R.; Nyvall, P.; Pedersen, M. The unique features of starch metabolism in red algae. Proc. R. Soc. Lond. B 2001, 268, 1417–1422. [Google Scholar] [CrossRef]
- Kromkamp, J. Formation and functional significance of storage products in cyanobacteria. New Zeal. J. Mar Fresh. 1987, 21, 457–465. [Google Scholar] [CrossRef]
- Chen, P.H.; Oswald, W.J. Thermochemical treatment for algal fermentation. Environ. Int. 1998, 24, 889–897. [Google Scholar] [CrossRef]
- Flores, E.; Wolk, C.P. Production, by filamentous, nitrogenfixing cyanobacteria, of a bacteriocin and of other antibiotics that kill related strains. Arch. Microbiol. 1986, 145, 215–219. [Google Scholar] [CrossRef]
- Debro, L.L.; Ward, H.B. Antibacterial activity of freshwater green algae. Planta. Med. 1979, 36, 375–378. [Google Scholar] [CrossRef]
- Katircioglu, H.; Beyatli, Y.; Aslim, B.; Yüksekdag, Z.; Atici, T. Screening for Antimicrobial Agent Production of Some Microalgae in Freshwater. Int. J. Microbiol. 2005, 2, 2. [Google Scholar]
- Han, S.M.; Ahn, B.J.; Lee, C.G.; Park, J.K. Expression and characterization of a chitinolyt ic enzyme in Chlorella vulgaris. J. Chitin Chitosan 2008, 13, 205–209. [Google Scholar]
- Park, J.K.; Kim, J.D.; Patil, S.S.; Kim, S.J.; Lee, C.G. Expression and characterization of chitinolytic enzymes in Chlamydomonas reinhardtii. J. Chitin Chitosan 2008, 13, 95–100. [Google Scholar]
- Chetsumon, A.; Maeda, I.; Umeda, F.; Yagi, K.; Miura, Y.; Mizoguchi, T. Antibiotic pro-duction by the immobilized cyanobacterium, Scytonema sp.TISTR 8208, in a seaweed-type photobioreactor. J. Appl. Phycol. 1994, 6, 539–543. [Google Scholar] [CrossRef]
- Bloor, S.; England, R.R. Elucidation and optimization of the medium constituents controlling antibiotic production by the cyanobacterium Nostoc muscorum. Enzyme Microb. Technol. 1991, 13, 76–81. [Google Scholar] [CrossRef]
- Noaman, N.H.; Fattah, A.; Khaleafa, M.; Zaky, S.H. Factors affecting antimicrobial activity of Synechococcus leopoliensis. Microbiol. Res. 2004, 159, 395–402. [Google Scholar] [CrossRef]
- Gacheva, G.; Gigova, L.; Ivanova, N.; Iliev, I.; Toshkova, R.; Gardeva, E.; Kussovski, V.; Najdenski, H. Suboptimal growth temperatures enhance the biological activity of cultured cyanobacterium Gloeocapsa. sp. J. Appl. Phycol. 2013, 25, 183–194. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mudimu, O.; Rybalka, N.; Bauersachs, T.; Born, J.; Friedl, T.; Schulz, R. Biotechnological Screening of Microalgal and Cyanobacterial Strains for Biogas Production and Antibacterial and Antifungal Effects. Metabolites 2014, 4, 373-393. https://doi.org/10.3390/metabo4020373
Mudimu O, Rybalka N, Bauersachs T, Born J, Friedl T, Schulz R. Biotechnological Screening of Microalgal and Cyanobacterial Strains for Biogas Production and Antibacterial and Antifungal Effects. Metabolites. 2014; 4(2):373-393. https://doi.org/10.3390/metabo4020373
Chicago/Turabian StyleMudimu, Opayi, Nataliya Rybalka, Thorsten Bauersachs, Jens Born, Thomas Friedl, and Rüdiger Schulz. 2014. "Biotechnological Screening of Microalgal and Cyanobacterial Strains for Biogas Production and Antibacterial and Antifungal Effects" Metabolites 4, no. 2: 373-393. https://doi.org/10.3390/metabo4020373
APA StyleMudimu, O., Rybalka, N., Bauersachs, T., Born, J., Friedl, T., & Schulz, R. (2014). Biotechnological Screening of Microalgal and Cyanobacterial Strains for Biogas Production and Antibacterial and Antifungal Effects. Metabolites, 4(2), 373-393. https://doi.org/10.3390/metabo4020373