Predicting the Response of Molluscs to the Impact of Ocean Acidification

 ,
,
Abstract
:1. Introduction
2. Life Cycle
3. Hypotheses on Responses of Molluscs
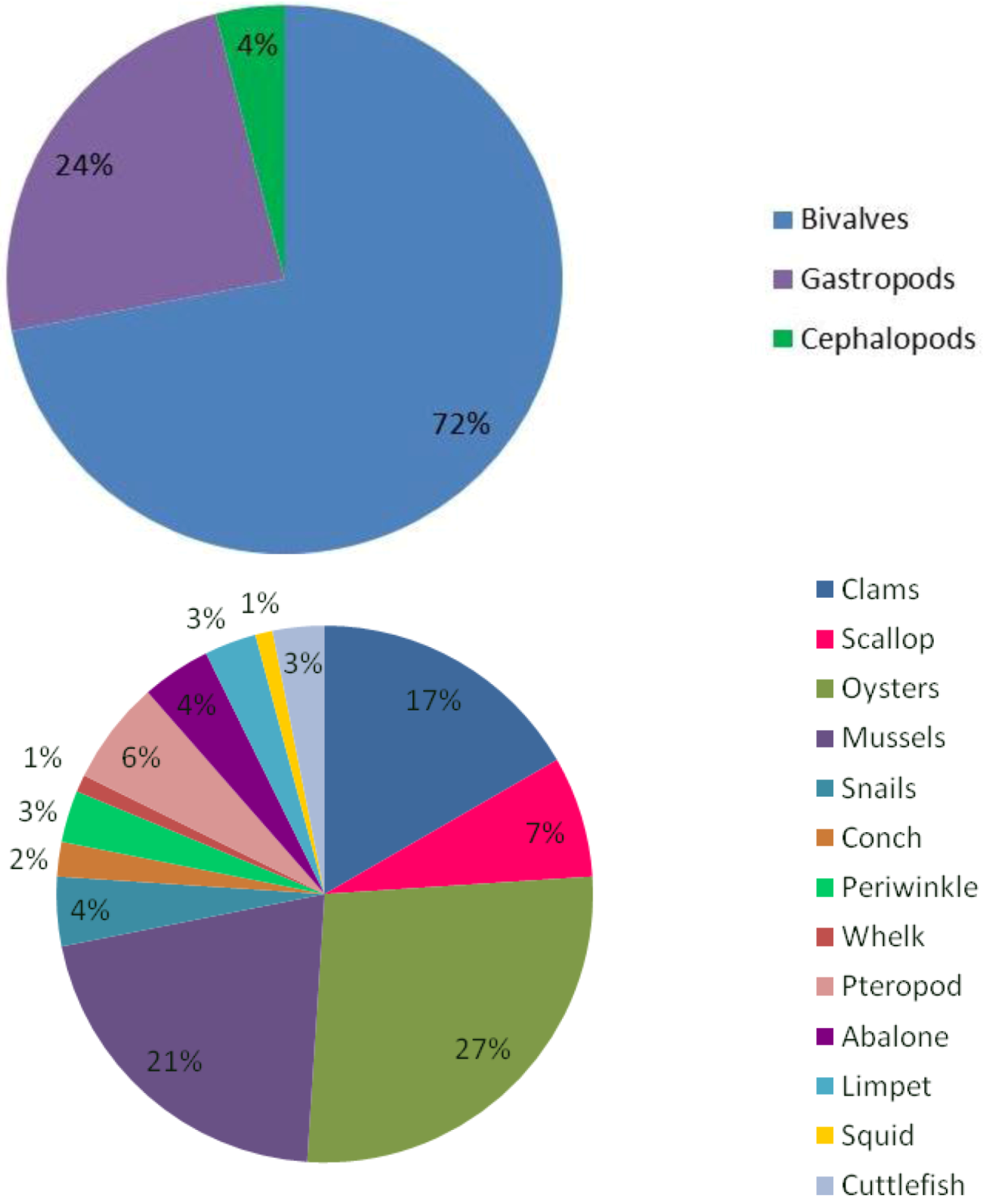
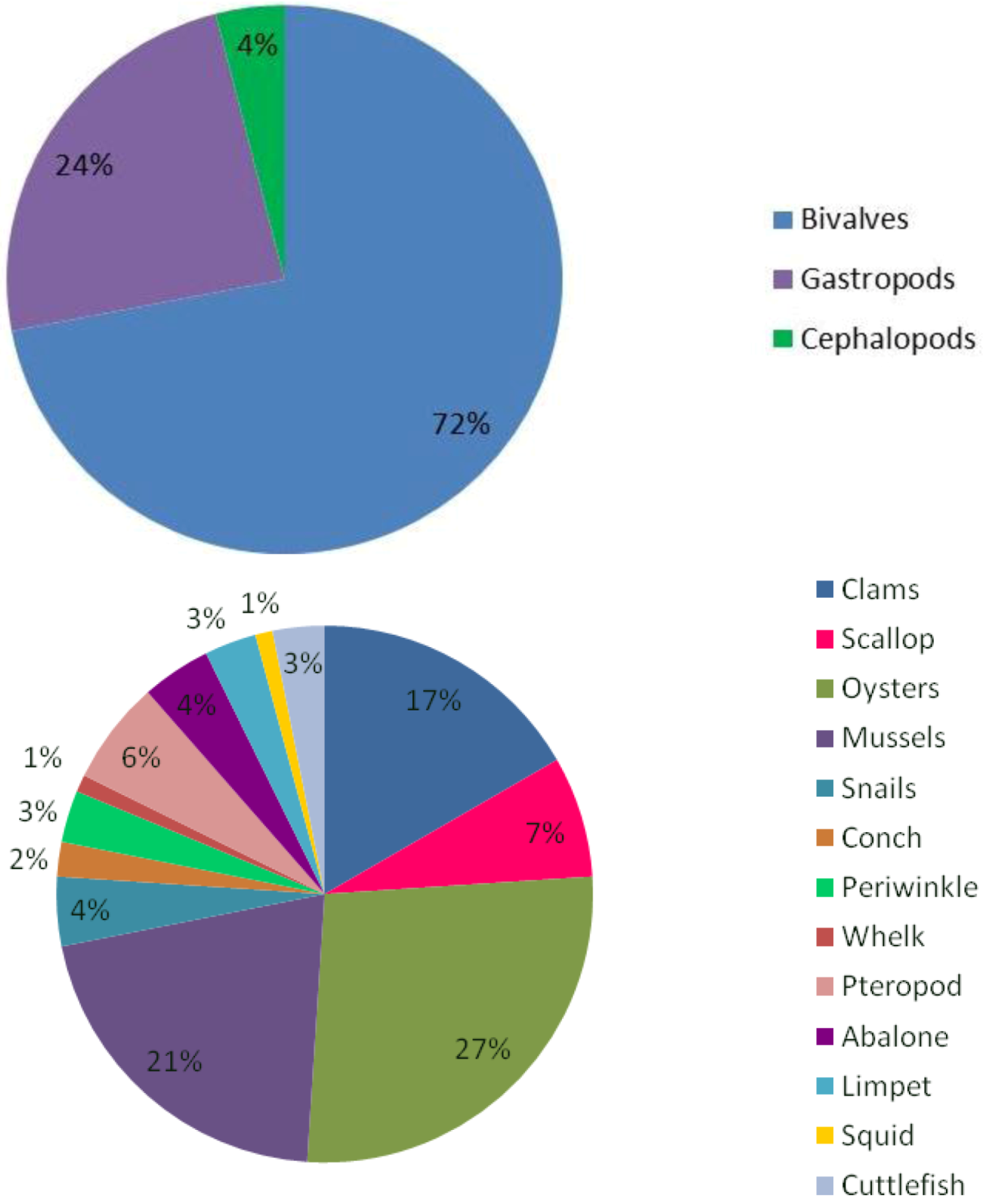
- (a)
- Molluscs will experience a reduction in calcification with the severity of this reduction differing depending on the CaCO3 polymorph which they secrete, their capacity to regulate acid-base status and the physicochemical environment of the population:

{kind=link}
{kind=link}
| Species | Experiment Duration | CO2/pH range | Measured | Impact | CO2/pH level that first caused an impact | Author |
|---|---|---|---|---|---|---|
| BIVALVES | ||||||
| Acesta excavata (clam) | 4 d | 530, 33,000 µatm 8.12, 6.35 | Respiration | ↓/= | −1.72 (33,000 µatm) −1.72 (33,000 µatm) −1.72 (33,000 µatm) | [161] |
| Excretion | = | |||||
| pHe | ↓ | |||||
| pHi | ↓ | |||||
| Argopecten irradians (scallop) | 60 d | 370, 547, 892, 2,551 ppm 8.15, 8.02, 7.83, 7.45 | Calcification | ↓ | −0.13 (547 ppm) | [48] |
| Chamelea nobilis (clam) | 7 d | n/a | Immune response | Changes | −0.4 | [135] |
| 8.1, 7.7, 7.4 | ||||||
| +Temp. 22, 28 °C | ||||||
| +Salinity 28, 34, 40 | ||||||
| Chlamys nobilis (scallop) | 2 h | n/a 8.1, 7.7, 7.4 | Respiration rate | ↓ | −0.4 | [158] |
| Clearance rate | ↓ | −0.4 | ||||
| Excretion rate | ↓ | −0.4 | ||||
| Crassostrea gigas (oyster) | 14 d | 380, 740 ppmv n/a | Calcification | ↓ | −0.2 (740 ppmv) | [45] |
| 30 d | n/a | Shell density | ↓ | −1.2 | [55] | |
| 8.2–5.4 | Shell weight | ↓ | −1.2 | |||
| 55 d | 582, 1480 ppm | pHe | ↓ | −0.39 (1,480 ppm) | [137] | |
| 8.07, 7.68 | PeCO2 | ↑ | −0.39 (1,480 ppm) | |||
| +Temp. 15, 25 °C | PeO2 | = | ||||
| CeCO2 | ↑ | −0.39 (1,480 ppm) | ||||
| HCO3− | ↑ | −0.39 (1,480 ppm) | ||||
| Ca2+ | = | |||||
| Na2+ | ↓ | −0.39 (1,480 ppm) | ||||
| K+ | ↑ | −0.39 (1,480 ppm) | ||||
| Tissue metabolite levels | ↓ | −0.39 (1,480 ppm) | ||||
| Metabolic rate (+temp.) | ↑ | −0.39 (1,480 ppm) | ||||
| Crassostrea virginica (oyster) | 15 h | n/a | Calcification (+salinity) | ↓ | −0.63 | [30] |
| 8.29, 7.66, 7.56 | ||||||
| +Temp 20, 30 °C | ||||||
| +Salinity 16, 32 | ||||||
| 14–140 d | 380, 3500 µatm 8.2, 7.5 | Survival | ↓ | −0.7 (3,500 µatm) | [40] | |
| Shell growth | ↓ | −0.7 (3,500 µatm) | ||||
| Somatic growth | ↓ | −0.7 (3,500 µatm) | ||||
| Shell ultrastructure | ↓ | −0.7 (3,500 µatm) | ||||
| Respiration rate | ↑ | −0.7 (3,500 µatm) | ||||
| 60 d | 370, 547, 892, 2551 ppm | Calcification | ↓ | −0.13 (547 ppm) | [48] | |
| 8.15, 8.02, 7.83, 7.45 | ||||||
| 77 d | 400, 700–800 µatm 8.11, 7.97 (Sal 15) +Salinity 15, 30 | Survival | ↓ | −0.14 (700–800 µatm) | [140] | |
| Tissue energy stores | ↓ | −0.14 (700–800 µatm) | ||||
| Somatic growth | ↓ | −0.14 (700–800 µatm) | ||||
| Tissue ATP | = | |||||
| Carbonic anhydrase | = | |||||
| 14 d | n/a | Shell dissolution | ↑ | −0.23 | [62] | |
| 7.9, 7.67, 7.38, 7.17 | ||||||
| 14 d | 395, 3,617 ppm | Oxidative stress | ↑ | −0.8 (3,617 ppm) | [66] | |
| 8.3, 7.5 | Cytoskeletal proteins | ↑ | ||||
| Laternula elliptica (clam) | 28 d | 390, 2,639 µatm | Shell dissolution | ↑ | −0.6 (2,639 µatm) | [54] |
| 8.2, 7.4 | ||||||
| 120 d | 187 *, 438, 735 µatm 8.32 *, 7.99, 7.78 | Metabolic rate | ↑ | −0.21 (735 µatm) | [152] | |
| HSP70 expression | ↑ | −0.21 (735 µatm) | ||||
| Chitin synthase expression | ↑ | −0.21 (735 µatm) | ||||
| Mercenaria mercenaria (clam) | 14 d | Ωarag 1.2, 0.6, 0.4 | Survival | ↓ | −0.6 (Ωarag 0.6) | [153] |
| 7.9, 7.3, 7.0 | ||||||
| 60 d | 370, 547, 892, 2551 ppm | Calcification | ↓ | −0.7 (2,551 ppm) | [48] | |
| 8.15, 8.02, 7.83, 7.45 | ||||||
| 8 h | 424, 1,120, 1950 µatm | Calcification | ↓ | −0.38 (1,120 µatm) | [61] | |
| 8.02, 7.64, 7.41 | ||||||
| Mya arebaria (clam) | 60 d | 370, 547, 892, 2551 ppm | Calcification | ↓ | −0.13 (547 ppm) | [48] |
| 8.15, 8.02, 7.83, 7.45 | ||||||
| Mytilus chilensis (mussel) | 70 d | 380, 750, 1200 ppm7.91, 7.71, 7.57 | Respiration rate | ↓ | −0.34 (1,200 ppm) | [160] |
| Excretion rate | = | |||||
| Ingestion rate | = | |||||
| Clearance rate | ↓ | −0.34 (1,200 ppm) | ||||
| Adsorption rate | ↓ | −0.34 (1,200 ppm) | ||||
| Mytilus edulis (mussel) | 2 h | 380, 740 ppmv | Calcification | ↓ | −0.2 (740 ppmv) | [45] |
| n/a | ||||||
| 30 d | n/a | Shell density | ↓ | −1.2 | [55] | |
| 8.2–5.4 | Shell weight | ↓ | −1.2 | |||
| 32 d | 665, 1,161, 1435, 3316 µatm | Immune response | ↓ | −0.1 (1,161 µatm) | [144] | |
| 7.8, 7.7, 7.5, 6.7 | Immune surveillance parameters | = | ||||
| 44 d | n/a | Survival | ↓ | −0.5 | [60] | |
| 8.1, 7.6, 7.4, 7.1, 6.7 | Shell growth | ↓ | −0.5 | |||
| 14–56 d | 385, 560, 840, 1120, 1400, 4000 µatm 8.05, 7.89, 7.81, 7.7, 7.56, 7.08 | Shell growth | ↓ | −0.49 (4,000 µatm) | [79] | |
| Somatic growth | = | |||||
| pHe | ↓ | −0.49 (4,000 µatm) | ||||
| PeCO32− | ↓ | −0.49 (4,000 µatm) | ||||
| PeCO2 | ↑ | −0.49 (4,000 µatm) | ||||
| HCO3− | = | |||||
| Ca2+ | = | |||||
| Na2+ | ↑ | −0.49 (4,000 µatm) | ||||
| K+ | = | |||||
| Mg2+ | = | |||||
| 2 mo | 385, 1120, 2400, 4000 µatm 8.03, 7.7, 7.38, 7.14 | Shell growth | ↓ | −0.33 (1,120 µatm) | [51] | |
| Somatic growth | = | |||||
| Metabolic rate |  | −0.33 (1,120 µatm) | ||||
| NH4+ excretion rate | ↑ | −0.33 (1,120 µatm) | ||||
| 2 mo | 385, 1120, 2400, 4000 µatm 8.03, 7.7, 7.38, 7.16 | Shell growth | ↑ | −0.65 (2,400 µatm) | [59] | |
| Gene expression (tyrosinases) | ↓ | −0.65 (2,400 µatm) | ||||
| Gene expression | ↓ | −0.65 (2,400 µatm) | ||||
| (chitinases) | ||||||
| 60 d | 370, 547, 892, 2551 ppm | Calcification | = | [48] | ||
| 8.15, 8.02, 7.83, 7.45 | ||||||
| 35 d | 472, 1021, 2114, 3350 µatm | Shell length | = | [65] | ||
| 8.01, 7.7, 7.4, 7.19 | Inorganic shell growth | ↓ | −0.31 (1,021 µatm) | |||
| +Food high, low | Organic shell growth | = | ||||
| Mytilus galloprovincialis (mussel) | 7 d | n/a 8.1, 7.7, 7.4 +Temp. 22, 28 °C +Salinity 28, 34, 40 | Immune response | Changes | −0.4 | [135] |
| 78 d | 963, 1989, 3790 µatm 8.03, 7.74, 7.48 | Somatic growth | ↑ | −0.3 | [159] | |
| Absorption efficiency | ↑ | −0.3 | ||||
| Ammonium excretion | ↑ | −0.3 | ||||
| Excretion rate | ↑ | −0.3 | ||||
| Clearance rate | = | |||||
| Ingestion rate | = | |||||
| Respiration rate | = | |||||
| 3 mo | 0.82, 3.82 mmHg CO2 8.05, 7.3 | Shell growth | ↓ | −0.75 (3.82 mmHg) | [43] | |
| Metabolic rate | ↓ | −0.75 (3.82 mmHg) | ||||
| pHe level | ↓ | −0.75 (3.82 mmHg) | ||||
| Protein degradation | ↑ | −0.75 (3.82 mmHg) | ||||
| HCO3− | ↓ | −0.75 (3.82 mmHg) | ||||
| Ca2+ | ↓ | −0.75 (3.82 mmHg) | ||||
| 84 d | 805, 1,698, 4,345 µatm 8.13, 7.84, 7.46 | Survival | = | −0.67 (4,345 µatm) | [63] | |
| Shell growth | ↓ | |||||
| Tissue growth | = | |||||
| Ostrea edulis (oyster) | 30 d | n/a | Shell density | ↓ | −1.2 | [55] |
| 8.2–5.4 | Shell weight | ↓ | −1.2 | |||
| Pecten maximus (scallop) | 30 d | 395, 1,086 µatm | Clapping performance | ↓ | −0.43 (1,086 µatm) | [165] |
| 4 °C: 8.19, 7.76 | Aerobic scope | ↓ | −0.43 (1,086 µatm) | |||
| 10 °: 8.25, 7.81 | ||||||
| (+temp. 4, 10 °C) | ||||||
| Perna viridis (green-lipped mussel) | 2 h | n/a 8.1, 7.7, 7.4 | Respiration rate | ↓ | −0.4 | [158] |
| Clearance rate | ↓ | −0.4 | ||||
| Excretion rate | ↓ | −0.4 | ||||
| Pinctada fucata (pearl oyster) | 2 h | n/a 8.1, 7.7, 7.4 | Respiration rate | ↓ | −0.4 | [158] |
| Clearance rate | ↓ | −0.4 | ||||
| Excretion rate | ↓ | −0.4 | ||||
| 28 d | n/a 8.1, 7.8, 7.6 | Shell strength | ↓ | −0.3 | [52] | |
| Ruditapes decussatus (clam) | 75 d | 8.05, 1698, 4345 µatm 8.13, 7.84, 7.46 | Survival | ↑ | −0.39 (1,698 µatm) | [72] |
| Calcification | = | |||||
| Shell growth | = | |||||
| Weight | = | |||||
| 87 d | 730, 1813, 3702 µatm 8.16, 7.82, 7.53 | Respiration rate | ↓ | −0.4 | [157] | |
| Clearance rate | ↓ | −0.4 | ||||
| Ingestion rate | ↓ | −0.4 | ||||
| Excretion rate | ↑ | −0.4 | ||||
| Ruditapes philippinarum (clam) | 28 d | n/a | Metal uptake | ↑ | −1.0 | [113] |
| 8.5–6.5 | ||||||
| Saccostrea glomerata (oyster) | 4 d | 375, 1000 ppm | Shell growth | ↓ | −0.36 (1,000 ppm) | [53] |
| 8.2, 7.84 | ||||||
| Yoldia eightsi (clam) | 35 d | 390, 2639 µatm | Shell dissolution | ↑ | −0.8 (2,639 µatm) | [54] |
| 8.2, 7.4 | ||||||
| Community | 30 d | 729–3010 µatm | Structure | ↓ | −0.3 | [138] |
| 8.0, 7.7, 7.3, 6.7 | ||||||
| (+Temp. 12, 16 °C) | ||||||
| GASTROPODS | ||||||
| Creithium vulgatum (snail) | ecosystem | 304–20812 µatm | Shell strength | ↓ | −0.7 | [46] |
| 8.17–6.57 | ||||||
| Clio pyramidata (pteropod) | 2 d | Ωarag > 1, < 1 | Shell dissolution | ↑ | −0.35 (Ωarag >1) | [5] |
| Crepidula fornicata (limpet) | 60 d | 370, 547, 892, 2551 ppm | Calcification | ↑ | −0.13 (547 ppm) | [48] |
| 8.15, 8.02, 7.83, 7.45 | ||||||
| Hexaplex trunculus (snail) | ecosystem | 304–20812 µatm | Shell strength | ↓ | −0.7 | [46] |
| 8.17–6.57 | ||||||
| Limacina helicina (pteropod) | 5 d | 350, 760 µatm | Calcification | ↓ | −0.3 (760 µatm) | [47] |
| 8.1, 7.8 | ||||||
| 4 - 14 d | 4 d: 318, 940 µatm 4 d: 8.13, 7.7 14 d: 318, 883 µatm 14 d: 8.13, 7.73 | Dissolution | ↑ | −0.43 (940 µatm) | [56] | |
| Dissolution | ↑ | −0.4 (883 µatm) | ||||
| 29 d | 180 *, 380, 750, 1150 µatm | Survival | ↓ | −0.43 (1,150 µatm) | [49] | |
| 8.27 *, 8.12, 7.81, 7.69 | Shell growth | ↓ | −0.31 (750 µatm) | |||
| (+temp.: 3, 5.5, 8 °C) | Shell degradation | ↑ | −0.31 (750 µatm) | |||
| Limacina retroversa (pteropod) | 8 d | 280 *, 350, 750, 1000 ppm | Shell growth | ↓ | −0.2 (750 ppm) | [57] |
| 8.2 *, 8.0, 7.8, 7.6 | Shell dissolution | ↑ | −0.2 (750 ppm) | |||
| Littorina littorea (periwinkle) | 15 d | n/a 7.97, 6.63 | Predator avoidance | = | [58] | |
| Predator avoidance (+predator cue) | ↑ | −1.34 | ||||
| Respiration rate | = | |||||
| Respiration rate (+ predator cue) | ↓ | −1.34 | ||||
| Shell thickness | = | |||||
| Shell thickness (+predator cue) | ↓ | −1.34 | ||||
| 60 d | 409, 606, 903, 2856 ppm | Calcification | ↓ | −0.09 (606 ppm) | [48] | |
| 8.09, 8.00, 7.86, 7.42 | ||||||
| 30 d | 428, 998 µatm 8.03, 7.67 | Shell length | ↓ | −0.36 | [64] | |
| Shell weight | ↑ | |||||
| Shell thickness | ↓ | |||||
| Aspect ratio | ↓ | |||||
| 30 d | 428, 998 µatm | Metabolic rate | ↓ | −0.36 | [162] | |
| 8.03, 7.67 | Energy metabolites (ATP) | ↓ | ||||
| Nacella concinna (limpet) | 14 d | Ωarag 2.66, 0.47 | Shell dissolution | ↑ | −0.8 (Ωarag 0.47) | [54] |
| 8.2, 7.4 | ||||||
| Nucella lamellosa (snail) | 6 d | 385, 785, 1200, 1585 ppm | Shell weight | ↓ | −0.18 (785 ppm) | [50] |
| 7.98, 7.80, 7.54 | Shell dissolution | ↑ | −0.18 (785 ppm) | |||
| Osilinus turbinata (snail) | ecosystem | 304–20812 µatm | Abundance | ↓ | −0.7 | [46] |
| 8.17–6.57 | ||||||
| Patella caerulea (snail) | ecosystem | 304–20812 µatm | Abundance | ↓ | −0.7 | [46] |
| 8.17–6.57 | ||||||
| Patella vulgata (limpet) | 5 d | 419, 2804 µatm 8.2, 7.6 | Metabolic rate | = | [155] | |
| Protein content | = | |||||
| Feeding rate | = | |||||
| Radula structure | ↓ | −0.6 (2,804 µatm) | ||||
| pHe | = | |||||
| PeCO2 | ↑ | −0.6 (2,804 µatm) | ||||
| HCO3− | ↑ | −0.6 (2,804 µatm) | ||||
| Ca2+ | ↑ | −0.6 (2,804 µatm) | ||||
| Mg2+ | = | |||||
| Strombus alatus (conch) | 60 d | 409, 606, 903, 2856 ppm | Calcification | ↓ | −0.67 (2,856 ppm) | [48] |
| 8.09, 8.00, 7.86, 7.42 | ||||||
| Strombus lubuanus (conch) | 6 mo | 360, 560 ppm 7.94, 7.90 | Survival | ↓ | −0.04 (560 ppm) | [44] |
| Shell growth | ↓ | −0.04 (560 ppm) | ||||
| Shell weight | ↓ | −0.04 (560 ppm) | ||||
| Urosalpinx cinerea (whelk) | 60 d | 409, 606, 903, 2856 ppm | Calcification | ↓ | −0.09 (606 ppm) | [48] |
| 8.09, 8.00, 7.86, 7.42 | ||||||
| Range of gastropods | 336–5148 ppm | Settlement | ↓ | −0.4 | [126] | |
| 8.09–7.08 | ||||||
| CEPHALOPODS | ||||||
| Dosidicus gigas (squid) | 8–12 h | 300, 1000 ppm | [136] | |||
| 7.93, 7.62 | Metabolic rate | ↓ | −0.31 (1,000 ppm) | |||
| +Temp. 10, 20, 25 °C | Activity level | ↓ | −0.31 (1,000 ppm) | |||
| + PO2 21, 1% | ||||||
| Sepia officinalis (cuttlefish) | 48 h | 493, 5922 ppm 8.12, 7.10 | pHe | ↓ | −1.02 (5,922 ppm) | [71] |
| pHi | = | |||||
| PeO2 | = | |||||
| PeCO2 | ↑ | −1.02 (5,922 ppm) | ||||
| HCO3− | ↑ | −1.02 (5,922 ppm) | ||||
| 42 d | 628, 4271, 6068 ppm 8.01, 7.23, 7.10 | Calcification | = | [67] | ||
| Growth | = | |||||
| Metabolic rate | = | |||||
| 42 d | 632, 6070 ppm | Calcification | ↑ | −1.0 (6,070 ppm) | [68] | |
| 8.10, 7.10 |
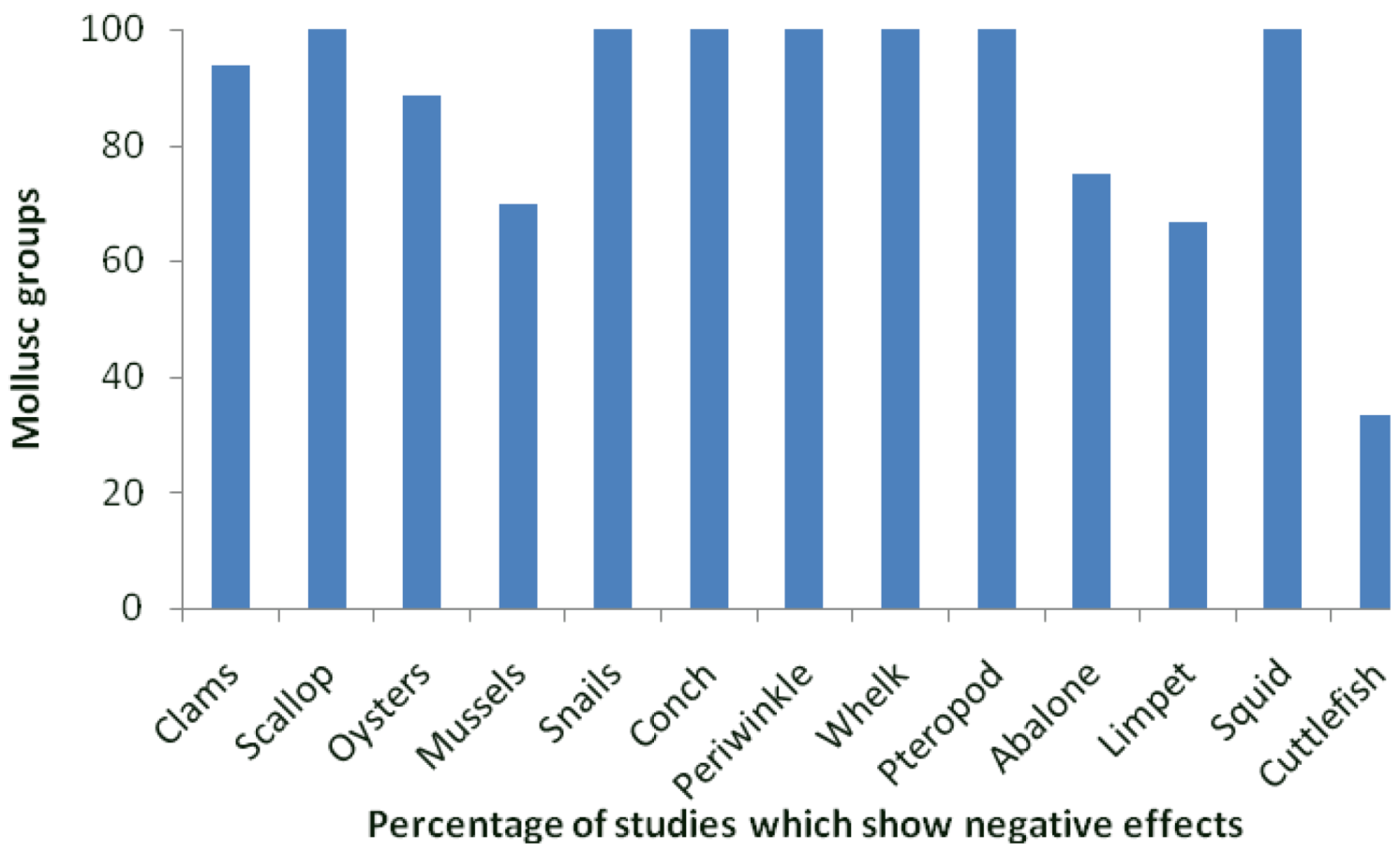
denotes parabolic response.- (b)
- Early-life history stages are more sensitive than juveniles and adults:
| Species | Experiment Duration | CO2/pH range | Measured | Impact | CO2/pH level that first caused an impact | Author |
|---|---|---|---|---|---|---|
| BIVALVES | ||||||
| Crassostrea gigas (oyster) | 1 h | 432, 1051 µatm 8.15, 7.8 | Fertilisation | = | [77] | |
| Sperm swimming speed | = | |||||
| Sperm motility | = | |||||
| 2 h | 348, 2268 µatm | Fertilisation | = | [74] | ||
| 8.21, 7.42 | ||||||
| 2 h | 375, 600, 750, 1000 µatm | Fertilisation | ↓ | −0.18 (600 µatm) | [78] | |
| 8.19, 8.01, 7.93, 7.82 | ||||||
| (+Temp. 18, 22, 26, 30 °C) | ||||||
| Macoma balthica (clam) | 24 h | n/a | Fertilisation | ↓ | −0.6 | [96] |
| 8.1, 7.8, 7.5 | ||||||
| Mytilus edulis (mussel) | 2 h | 368–381, 1291–1332 µatm | Fertilisation | = | [94] | |
| 8.13, 7.63 | ||||||
| Mytilus galloprovincialis (mussel) | 2 h | 380, 2000 ppm | Fertilisation | = | [35] | |
| 8.13, 7.42 | ||||||
| Saccostrea glomerata (oyster) | 2 h | 375, 600, 750, 1000 ppm | Fertilisation | ↓ | −0.17 (600 ppm) | [37] |
| (+Temp. 18, 22, 26, 30 °C) | ||||||
| 2 h | 375, 600, 750, 1000 µatm | Fertilisation | ↓ | −0.18 (600 µatm) | [78] | |
| 8.19, 8.01, 7.93, 7.82 | ||||||
| (+Temp. 18, 22, 26, 30 °C) | ||||||
| GASTROPODS | ||||||
| Haliotis coccoradiata (abalone) | 15 min | 327–335, 814–851, 1051–1104, 1729–1828 µatm | Fertilisation | = | [95] | |
| 8.25, 7.9, 7.8, 7.6 | ||||||
| (+Temp. +2, +4 °C) | ||||||
| Haliotis discus hannai (abalone) | 15 h | 450, 500, 1100, 1650, 2150 µatm | Fertilisation | ↓ | −0.48 (1,650 ppm) | [97] |
| 8.02, 7.94, 7.68, 7.54, 7.43 |
| Species | Experiment Duration | CO2/pH range | Measured | Impact | CO2/pH level that first caused an impact | Author |
|---|---|---|---|---|---|---|
| BIVALVES | ||||||
| Argopecten irradians (scallop) | 19 d | 355, 681, 1609 ppm 8.08, 7.83, 7.48 | Survival | ↓ | −0.25 (681 ppm) | [103] |
| Development time | ↓ | −0.25 (681 ppm) | ||||
| Size | ↓ | −0.25 (681 ppm) | ||||
| 52 d | 244 *, 387, 739, 1,529 ppm 8.17, 8.04, 7.80, 7.53 | Survival | ↓ | −0.24 | [104] | |
| Shell thickness | ↓ | −0.24 | ||||
| Shell diameter | ↓ | −0.24 | ||||
| Lipid index | ↓ | −0.24 | ||||
| 18 d | 240 *, 387, 773 ppm 8.21, 8.08, 7.81 | Survival | ↓ | −0.27 | [108] | |
| Shell diameter | ↓ | −0.27 | ||||
| Metamorphosis | ↓ | −0.27 | ||||
| Lipid index | ↓ | −0.27 | ||||
| 20 d | 240 *, 390, 850 ppm | Survival | ↓ | −0.1 | [109] | |
| Shell diameter | ↓ | −0.1 | ||||
| Metamorphosis | ↓ | −0.1 | ||||
| Lipid index | ↓ | −0.1 | ||||
| Crassostrea angulata (oyster) | 5 d | 469, 964, 1447, 2390 µatm 8.08, 7.81, 7.64, 7.43 (+Salinity 24, 34 ppt) | Shell growth | ↓ | −0.65 | [110] |
| Crassostrea ariakensis (oyster) | 30 d | 291 *, 386, 581, 823 µatm | Shell growth | = | [100] | |
| 8.17 *, 8.08, 7.92, 7.79 | Shell calcification | = | ||||
| Crassostrea gigas (oyster) | 48 h | 348, 2268 µatm 8.21, 7.42 | Survival | ↓ | −0.79 (2,268 µatm) | [76] |
| Size | ↓ | −0.79 (2,268 µatm) | ||||
| Development rate | ↓ | −0.79 (2,268 µatm) | ||||
| Shell normality | ↓ | −0.79 (2,268 µatm) | ||||
| 48 h–4 d | 375, 600, 750, 1000 µatm 8.19, 8.01, 7.93, 7.82 (+Temp. 18, 22, 26, 30 °C) | Survival | ↓ | −0.18 (600 µatm) | [78] | |
| Size | ↓ | −0.18 (600 µatm) | ||||
| Shell normality | ↓ | −0.18 (600 µatm) | ||||
| % Umbonate larvae | ↓ | −0.18 (600 µatm) | ||||
| % Eyed larvae | ↓ | −0.18 (600 µatm) | ||||
| 24–48 h | n/a 8.2–7.6 (in situ) | Early size (24 h) | = | −0.4 | [26] | |
| Mid size (48 h) | ↓ | |||||
| Shell normality | = | |||||
| 6 d | Control, 2275 µatm 8.0, 7.5 | Protein expression | ↓ | −0.5 (2,275 µatm) | [124] | |
| 3 d | 449, 1020, 2171 µatm 8.03, 7.72, 7.41 | Hatching | = | [116] | ||
| Size | = | |||||
| Calcification | = | |||||
| 1–3d | Day 1 466, 812, 1150 µatm 7.97, 7.75, 7.61 | Survival | = | [111] | ||
| Size | = | |||||
| Development rate | = | |||||
| Calcification | ↑ | −0.23 (781 µatm) | ||||
| Day 3 428, 781, 1031 µatm 8.00, 7.77, 7.66 | Survival | ↓ | −0.23 (781 µatm) | |||
| Size | ↓ | −0.34 (1,031 µatm) | ||||
| Development rate | = | |||||
| Calcification | ↓ | −0.34 (1,031 µatm) | ||||
| Crassostrea virginica (oyster) | 30 d | 291 *, 386, 581, 823 µatm | Size | ↓ | −0.16 (581 µatm) | [100] |
| 8.17 *, 8.08, 7.92, 7.79 | Calcification | ↓ | −0.16 (581 µatm) | |||
| 20 d | 355, 681, 1609 ppm 8.08, 7.83, 7.48 | Survival | ↓ | −0.6 (1,609 ppm) | [103] | |
| Size | ↓ | −0.25 (681 ppm) | ||||
| Development rate | ↓ | −0.25 (681 ppm) | ||||
| 20 d | 240 *, 390, 850 ppm | Survival | ↓ | −0.2 (850 ppm) | [109] | |
| Shell diameter | ↓ | −0.2 (850 ppm) | ||||
| Metamorphosis | ↓ | −0.2 (850 ppm) | ||||
| Lipid index | ↓ | −0.2 (850 ppm) | ||||
| Macoma balthica (clam) | 19 d | n/a 8.1, 7.8, 7.5 | Hatching | ↓ | −0.3 | [96] |
| Survival | ↓ | −0.6 | ||||
| Size | ↓ | −0.3 | ||||
| Metamorphosis | = | |||||
| Mytilus californianus (mussel) | 5–8 d | 380, 540, 970 ppm 8.07, 7.95, 7.75 | Shell area | ↓ | −0.12 (540 ppm) | [107] |
| Shell thickness | ↓ | −0.12 (540 ppm) | ||||
| Shell strength | ↓ | −0.12 (540 ppm) | ||||
| Tissue mass | ↓ | −0.12 (540 ppm) | ||||
| Mytilus edulis (mussel) | 13 d | 460, 1100, 1900 µatm | Survival | = | [102] | |
| 8.1, 7.8, 7.6 | Hatching | ↓ | −0.5 (1,900 µatm) | |||
| Size | ↓ | −0.3 (1,100 µatm) | ||||
| 64 d | 368, 1291 µatm 8.13, 7.63 | Size | ↓ | −0.5 (1,291 µatm) | [94] | |
| Development rate | = | |||||
| Shell normality | = | |||||
| Feeding rate | = | |||||
| Mytilus galloprovincialis (mussel) | 6 d | 380, 2000 ppm 8.13, 7.42 | Size | ↓ | −0.71 (2,000 ppm) | [36] |
| Development rate | ↓ | −0.71 (2,000 ppm) | ||||
| Shell normality | ↓ | −0.71 (2,000 ppm) | ||||
| Mytilus trossolus (mussel) | 60 h | 400, 1000 ppm 8.3, 7.9 | Shell length | ↓ | −0.4 (1,000 ppm) | [112] |
| Mercenaria mercenaria (clam) | 18 d | 355, 681, 1609 ppm 8.08, 7.83, 7.48 | Survival | ↓ | −0.25 (681 ppm) | [103] |
| Size | ↓ | −0.25 (681 ppm) | ||||
| Development rate | ↓ | −0.25 (681 ppm) | ||||
| 36 d | 244 *, 387, 739, 1529 ppm 8.17, 8.04, 7.80, 7.53 | Survival | ↓ | −0.24 | [104] | |
| Shell thickness | ↓ | −0.24 | ||||
| Shell diameter | ↓ | −0.24 | ||||
| Lipid index | ↓ | −0.24 | ||||
| 18 d | 240 *, 387, 773 ppm 8.21, 8.08, 7.81 | Survival | ↓ | −0.27 | [108] | |
| Shell diameter | ↓ | −0.27 | ||||
| Metamorphosis | ↓ | −0.27 | ||||
| Lipid index | ↓ | −0.27 | ||||
| Saccostrea glomerata (oyster) | 48 h | 375, 600, 750, 1000 ppm | Survival | ↓ | −0.17 | [37] |
| 8.20, 8.03, 7.95, 7.84 | Size | ↓ | −0.17 | |||
| Temp.: 18, 22, 26, 30 °C | Shell normality | ↓ | −0.17 | |||
| 48 h–4 d | 375, 600, 750, 1000 µatm 8.19, 8.01, 7.93, 7.82 Temp.: 18, 22, 26, 30 °C | Survival | ↓ | −0.18 | [78] | |
| Size | ↓ | −0.18 | ||||
| Shell normality | ↓ | −0.18 | ||||
| % Umbonate larvae | ↓ | −0.18 | ||||
| % Eyed larvae | ↓ | −0.18 | ||||
| 8 d | 220, 509, 756 µatm 8.11, 7.81, 7.64 | Survival | ↓ | −0.3 | [101] | |
| Size | ↓ | −0.3 | ||||
| Shell normality | ↓ | −0.3 | ||||
| Range of bivalves | 336–5148 µatm | Settlement | ↓ | −0.4 | [126] | |
| 8.15–7.08 | ||||||
| GASTROPODS | ||||||
| Cavolina inflexia (pteropod) | 13 d | 380, 857, 1713 µatm | Size | ↓ | −0.28 (857 µatm) | [105] |
| 8.1, 7.82, 7.58 | Shell normality | ↓ | −0.28 (857 µatm) | |||
| Haliotis cocoradiata (abalone) | 21 h | 306, 1,077, 1770 ppm | Calcification | ↓ | −0.4 (1,077 ppm) | [12] |
| 8.2, 7.8, 7.6 | Shell normality | ↓ | −0.4 (1,077 ppm) | |||
| Haliotis discus hannai (abalone) | 90 h | 450, 500, 1100, 1650, 2150 µatm 8.02, 7.94, 7.68, 7.54, 7.43 | Hatching rate | ↓ | −0.34 (1,100 ppm) | [97] |
| Survival | = | |||||
| Shell normality | ↓ | −0.34 (1,100 ppm) | ||||
| Haliotis rufescens (abalone) | 6 d | 380, 570, 990 ppm | Thermal tolerance | ↓ | −0.18 (990 ppm) | [123] |
| 8.05, 7.97, 7.87 | Ap24 expression | = | ||||
| Littoria obtusata (periwinkle) | 24 d | Control, 1093 ppm 8.1, 7.6 | Viability | ↓ | −0.5 (1,093 ppm) | [106] |
| Development rate | ↓ | −0.5 (1,093 ppm) | ||||
| Heart rate | ↓ | −0.5 (1,093 ppm) | ||||
| Altered behaviour | −0.5 (1,093 ppm) | |||||
| Altered shell | −0.5 (1,093 ppm) |
- (c)
- Multiple stressors will exacerbate the effects of ocean acidification:
- (d)
- Exposure of adults to elevated CO2 during reproductive conditioning will result in positive carry-over effects passed from adults to their offspring and will increase the resilience of molluscs to ocean acidification:
4. Physiological Responses of Molluscs
5. Food Availability and Energy “Trade-offs”
6. Conclusions and Future Research

Acknowledgments
References
- Climate Change 2001: The Scientific Basis Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change; Houghton, J.T.; Ding, Y.; Griggs, D.G.; Noguer, M.; van der Linden, P.J.; Dai, X.; Maskell, K.; Johnson, C.A. (Eds.) Cambridge University Press: Cambridge, UK, 2001; Volume 881.
- Solomon, S. Climate Change 2007: The Physical Science Basis: Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Doney, S.C.; Fabry, V.J.; Feely, R.A.; Kleypas, J.A. Ocean acidification: The other CO2 problem. Mar. Sci. 2009, 1, 169–192. [Google Scholar]
- Raven, J.; Caldeira, K.; Elderfield, H.; Hoegh-Guldberg, O.; Liss, P.; Riebesell, U.; Shepherd, J.; Turley, C.; Watson, A. Ocean Acidification due to Increasing Atmospheric Carbon Dioxide; The Royal Society: London, UK, 2005. [Google Scholar]
- Orr, J.C.; Fabry, V.J.; Aumont, O.; Bopp, L.; Doney, S.C.; Feely, R.A.; Gnanadesikan, A.; Gruber, N.; Ishida, A.; Joos, F. Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms. Nature 2005, 437, 681–686. [Google Scholar] [CrossRef] [Green Version]
- Feely, R.A.; Sabine, C.L.; Lee, K.; Berelson, W.; Kleypas, J.; Fabry, V.J.; Millero, F.J. Impact of anthropogenic CO2 on the CaCO3 system in the oceans. Science 2004, 305, 362–366. [Google Scholar] [CrossRef]
- Cooley, S.R.; Doney, S.C. Anticipating ocean acidification’s economic consequences for commercial fisheries. Environ. Res. Lett. 2009, 4, 024007. [Google Scholar] [CrossRef]
- Guinotte, J.M.; Fabry, V.J. Ocean acidification and its potential effects on marine ecosystems. Ann. N. Y. Acad. Sci. 2008, 1134, 320–342. [Google Scholar] [CrossRef]
- Hendriks, I.; Duarte, C.; Alvarez, M. Vulnerability of marine biodiversity to ocean acidification: A meta-analysis. Estuar. Coast. Shelf Sci. 2010, 86, 157–164. [Google Scholar] [CrossRef]
- Dupont, S.; Dorey, N.; Thorndyke, M. What meta-analysis can tell us about vulnerability of marine biodiversity to ocean acidification? Estuar. Coast. Shelf Sci. 2010, 89, 182–185. [Google Scholar] [CrossRef]
- Duarte, C.A.; Giarratano, E.; Amin, O.A.; Comoglio, L.I. Heavy metal concentrations and biomarkers of oxidative stress in native mussels (Mytilus edulis chilensis) from Beagle Channel coast (Tierra del Fuego, Argentina). Mar. Pollut. Bull. 2011, 62, 1895–1904. [Google Scholar] [CrossRef]
- Byrne, M.; Ho, M.; Wong, E.; Soars, N.A.; Selvakumaraswamy, P.; Shepard-Brennand, H.; Dworjanyn, S.A.; Davis, A.R. Unshelled abalone and corrupted urchins: Development of marine calcifiers in a changing ocean. Proc. Roy. Soc. B 2011, 278, 2376–2383. [Google Scholar] [CrossRef]
- Kroeker, K.J.; Kordas, R.L.; Crim, R.N.; Singh, G.G. Meta-analysis reveals negative yet variable effects of ocean acidification on marine organisms. Ecol. Lett. 2010, 13, 1419–1434. [Google Scholar] [CrossRef]
- Ross, P.M.; Parker, L.; O’Connor, W.A.; Bailey, E.A. The impact of ocean acidification on reproduction, early development and settlement of marine organisms. Water 2011, 3, 1005–1030. [Google Scholar] [CrossRef]
- Gutiérrez, J.L.; Jones, C.G.; Strayer, D.L.; Iribarne, O.O. Mollusks as ecosystem engineers: The role of shell production in aquatic habitats. Oikos 2003, 101, 79–90. [Google Scholar] [CrossRef]
- Norling, P.; Kautsky, N. Patches of the mussel Mytilus sp. Are islands of high biodiversity in subtidal sediment habitats in the Baltic Sea. Aquat. Biol. 2008, 4, 75–87. [Google Scholar]
- Buschbaum, C.; Dittmann, S.; Hong, J.S.; Hwang, I.S.; Strasser, M.; Thiel, M.; Valdivia, N.; Yoon, S.P.; Reise, K. Mytilid mussels: Global habitat engineers in coastal sediments. Helgol. Mar. Res. 2009, 63, 47–58. [Google Scholar] [CrossRef] [Green Version]
- Comeau, S.; Jeffree, R.; Teyssié, J.L.; Gattuso, J.P. Response of the arctic pteropod Limacina helicina to projected future environmental conditions. PLoS One 2010, 5, e11362. [Google Scholar]
- Gosling, E. Bivalve Molluscs; Blackwell Publishing: Oxford, UK, 2003. [Google Scholar]
- Cooley, S.R.; Lucey, N.; Kite‐Powell, H.; Doney, S.C. Nutrition and income from molluscs today imply vulnerability to ocean acidification tomorrow. Fish Fish. 2012, 13, 182–215. [Google Scholar] [CrossRef]
- Doney, S.C.; Ruckelshaus, M.; Duffy, J.E.; Barry, J.P.; Chan, F.; English, C.A.; Galindo, H.M.; Grebmeier, J.M.; Hollowed, A.B.; Knowlton, N. Climate change impacts on marine ecosystems. Mar. Sci. 2012, 4, 11–37. [Google Scholar]
- Narita, D.; Rehdanz, K.; Tol, R.S.J. Economic costs of ocean acidification: A look into the impacts on global shellfish production. Clim. Change 2012, 113, 1049–1065. [Google Scholar] [CrossRef]
- Fisheries Aquaculture Department, The State of World Fisheries and Aquaculture 2008; Food and Agriculture Organization of the United Nations: Rome, Italy, 2010.
- Smith, M.D.; Roheim, C.A.; Crowder, L.B.; Halpern, B.S.; Turnipseed, M.; Anderson, J.L.; Asche, F.; Bourillón, L.; Guttormsen, A.G.; Khan, A. Sustainability and global seafood. Science 2010, 327, 784–786. [Google Scholar] [CrossRef]
- Swartz, W.; Rashid Sumaila, U.; Watson, R.; Pauly, D. Sourcing seafood for the three major markets: The EU, Japan and the USA. Mar. Policy 2010, 34, 1366–1373. [Google Scholar] [CrossRef]
- Barton, A.; Hales, B.; Waldbusser, G.G.; Langdon, C.; Feely, R.A. The pacific oyster, Crassostrea gigas, shows negative correlation to naturally elevated carbon dioxide levels: Implications for near-term ocean acidification effects. Limnol. Oceanogr. 2012, 57, 698–710. [Google Scholar] [CrossRef] [Green Version]
- Crim, R.N.; Sunday, J.M.; Harley, C.D.G. Elevated seawater CO2 concentrations impair larval development and reduce larval survival in endangered northern abalone (Haliotis kamtschatkana). J. Exp. Mar. Biol. Ecol. 2011, 400, 272–277. [Google Scholar] [CrossRef]
- Hankewich, S.; Lessard, J. Resurvey of Northern Abalone, Haliotis Kamtschatkana, Populations along the Central Coast of British Columbia, May 2006; Fisheries and Oceans Canada: Nanaimo, Canada, 2008. [Google Scholar]
- Feely, R.A.; Alin, S.R.; Newton, J.; Sabine, C.L.; Warner, M.; Devol, A.; Krembs, C.; Maloy, C. The combined effects of ocean acidification, mixing, and respiration on pH and carbonate saturation in an urbanized estuary. Estuar. Coast. Shelf Sci. 2010, 88, 442–449. [Google Scholar] [CrossRef]
- Waldbusser, G.G.; Voigt, E.P.; Bergschneider, H.; Green, M.A.; Newell, R.I.E. Biocalcification in the Eastern oyster (Crassostrea virginica) in relation to long-term trends in Chesapeake Bay pH. Estuar. Coasts 2011, 34, 221–231. [Google Scholar] [CrossRef]
- Feely, R.A.; Sabine, C.L.; Hernandez-Ayon, J.M.; Ianson, D.; Hales, B. Evidence for upwelling of corrosive “acidified” water onto the continental shelf. Science 2008, 320, 1490–1492. [Google Scholar] [CrossRef]
- Jackson, A.; Chapman, M.; Underwood, A. Ecological interactions in the provision of habitat by urban development: Whelks and engineering by oysters on artificial seawalls. Austral Ecol. 2008, 33, 307–316. [Google Scholar] [CrossRef]
- Thorson, G. Reproductive and larval ecology of marine bottom invertebrates. Biol. Rev. 1950, 25, 1–45. [Google Scholar] [CrossRef]
- Wilt, F.H. Developmental biology meets materials science: Morphogenesis of biomineralized structures. Dev. Biol. 2005, 280, 15–25. [Google Scholar] [CrossRef]
- Kurihara, H.; Asai, T.; Kato, S.; Ishimatsu, A. Effects of elevated pCO2 on early development in the mussel Mytilus galloprovincialis. Aquat. Biol. 2009, 4, 225–233. [Google Scholar] [CrossRef]
- Kurihara, H. Effects of CO2-driven ocean acidification on the early developmental stages of invertebrates. Mar. Eco. Prog. Ser. 2008, 373, 275–284. [Google Scholar] [CrossRef]
- Parker, L.M.; Ross, P.M.; O’Connor, W.A. The effect of ocean acidification and temperature on the fertilization and embryonic development of the Sydney rock oyster Saccostrea glomerata (Gould 1850). Glob. Change Biol. 2009, 15, 2123–2136. [Google Scholar] [CrossRef]
- Parker, L.M.; Ross, P.M.; O’Connor, W.A.; Borysko, L.; Raftos, D.A.; Pörtner, H.O. Adult exposure influences offspring response to ocean acidification in oysters. Glob. Change Biol. 2012, 18, 82–92. [Google Scholar] [CrossRef]
- ESF, European Food Systems in a Changing World. Report of a ESF/cost Forward Look; European Science Foundation: Strasbourg, France, 2009.
- Beniash, E.; Ivanina, A.; Lieb, N.S.; Kurochkin, I.; Sokolova, I.M. Elevated level of carbon dioxide affects metabolism and shell formation in oysters Crassostrea virginica. Mar. Ecol. Prog. Ser. 2010, 419, 95–108. [Google Scholar] [CrossRef]
- Fabry, V.J.; Seibel, B.A.; Feely, R.A.; Orr, J.C. Impacts of ocean acidification on marine fauna and ecosystem processes. ICES J. Mar. Sci. 2008, 65, 414–432. [Google Scholar] [CrossRef]
- Wicks, L.C.; Roberts, M. Benthic invertebrates in a high-CO2 world. Oceanog. Mar. Biol. Ann. Rev. 2012, 50, 127–188. [Google Scholar]
- Michaelidis, B.; Ouzounis, C.; Paleras, A.; Pörtner, H.O. Effects of long-term moderate hypercapnia on acid-base balance and growth rate in marine mussels Mytilus galloprovincialis. Mar. Ecol. Prog. Ser. 2005, 293, 109–118. [Google Scholar] [CrossRef]
- Shirayama, Y.; Thornton, H. Effect of increased atmospheric CO2 on shallow water marine benthos. J. Geophys. Res. 2005, 110, C09S08. [Google Scholar] [CrossRef]
- Gazeau, F.; Quiblier, C.; Jansen, J.M.; Gattuso, J.P.; Middelburg, J.J.; Heip, C.H.R. Impact of elevated CO2 on shellfish calcification. Geophys. Res. Lett. 2007, 34. [Google Scholar] [CrossRef]
- Hall-Spencer, J.M.; Rodolfo-Metalpa, R.; Martin, S.; Ransome, E.; Fine, M.; Turner, S.M.; Rowley, S.J.; Tedesco, D.; Buia, M.C. Volcanic carbon dioxide vents show ecosystem effects of ocean acidification. Nature 2008, 454, 96–99. [Google Scholar]
- Comeau, S.; Gorsky, G.; Jeffree, R.; Teyssie, J.; Gattuso, J. Impact of ocean acidification on a key Arctic pelagic mollusc (Limacina helicina). Biogeosciences 2009, 6, 1877–1882. [Google Scholar] [CrossRef]
- Ries, J.B.; Cohen, A.L.; McCorkle, D.C. Marine calcifiers exhibit mixed responses to CO2-induced ocean acidification. Geology 2009, 37, 1131–1134. [Google Scholar] [CrossRef]
- Lischka, S.; Buedenbender, J.; Boxhammer, T.; Riebesell, U. Impact of ocean acidification and elevated temperatures on early juveniles of the polar shelled pteropod Limacina helicina: Mortality, shell degradation, and shell growth. Biogeosciences 2010, 7, 8177–8214. [Google Scholar] [CrossRef]
- Nienhuis, S.; Palmer, A.R.; Harley, C.D.G. Elevated CO2 affects shell dissolution rate but not calcification rate in a marine snail. Proc. Roy. Soc. B 2010, 277, 2553–2558. [Google Scholar] [CrossRef]
- Thomsen, J.; Melzner, F. Moderate seawater acidification does not elicit long-term metabolic depression in the blue mussel Mytilus edulis. Mar. Biol. 2010, 157, 2667–2676. [Google Scholar] [CrossRef] [Green Version]
- Welladsen, H.M.; Southgate, P.C.; Heimann, K. The effects of exposure to near-future levels of ocean acidification on shell characteristics of Pinctada fucata (Bivalvia: Pteriidae). Molluscan Res. 2010, 30, 125–130. [Google Scholar]
- Parker, L.M.; Ross, P.M.; O’Connor, W.A. Populations of the Sydney rock oyster, Saccostrea glomerata, vary in response to ocean acidification. Mar. Biol. 2011, 158, 689–697. [Google Scholar] [CrossRef]
- McClintock, J.B.; Angus, R.A.; Mcdonald, M.R.; Amsler, C.D.; Catledge, S.A.; Vohra, Y.K. Rapid dissolution of shells of weakly calcified antarctic benthic macroorganisms indicates high vulnerability to ocean acidification. Antarct. Sci. 2009, 21, 449–456. [Google Scholar] [CrossRef]
- Bamber, R. The effects of acidic seawater on three species of lamellibranch mollusc. J. Exp. Mar. Biol. Ecol. 1990, 143, 181–191. [Google Scholar] [CrossRef]
- Bednaršek, N.; Tarling, G.A.; Bakker, D.C.E.; Fielding, S.; Cohen, A.; Kuzirian, A.; McCorkle, D.; Lézé, B.; Montagna, R. Description and quantification of pteropod shell dissolution: A sensitive bioindicator of ocean acidification. Glob. Change Biol. 2012, 18, 2378–2388. [Google Scholar] [CrossRef] [Green Version]
- Manno, C.; Morata, N.; Primicerio, R. Limacina retroversa response to combined effects of ocean acidification and sea water freshening. Estuar. Coast. Shelf Sci. 2012, 113, 163–171. [Google Scholar] [CrossRef]
- Bibby, R.; Cleall-Harding, P.; Rundle, S.; Widdicombe, S.; Spicer, J. Ocean acidification disrupts induced defences in the intertidal gastropod Littorina littorea. Biol. Lett. 2007, 3, 699–701. [Google Scholar] [CrossRef]
- Hüning, A.K.; Melzner, F.; Thomsen, J.; Gutowska, M.A.; Krämer, L.; Frickenhaus, S.; Rosenstiel, P.; Pörtner, H.O.; Philipp, E.E.R.; Lucassen, M. Impacts of seawater acidification on mantle gene expression patterns of the Baltic Sea blue mussel: Implications for shell formation and energy metabolism. Mar. Biol. 2012. [Google Scholar] [CrossRef] [Green Version]
- Berge, J.A.; Bjerkeng, B.; Pettersen, O.; Schaanning, M.T.; Øxnevad, S. Effects of increased sea water concentrations of CO2 on growth of the bivalve Mytilus edulis L. Chemosphere 2006, 62, 681–687. [Google Scholar] [CrossRef]
- Waldbusser, G.G.; Bergschneider, H.; Green, M.A. Size-dependent pH effect on calcification in post-larval hard clam Mercenaria spp. Mar. Ecol. Prog. Ser. 2010, 417, 171–182. [Google Scholar] [CrossRef]
- Waldbusser, G.G.; Steenson, R.A.; Green, M.A. Oyster shell dissolution rates in estuarine waters: Effects of pH and shell legacy. J. Shellfish Res. 2011, 30, 659–669. [Google Scholar] [CrossRef] [Green Version]
- Range, P.; Piló, D.; Ben-Hamadou, R.; Chícharo, M.A.; Matias, D.; Joaquim, S.; Oliveira, A.P.; Chícharo, L. Seawater acidification by CO2 in a coastal lagoon environment: Effects on life history traits of juvenile mussels Mytilus galloprovincialis. J. Exp. Mar. Biol. Ecol. 2012, 424–425, 89–98. [Google Scholar]
- Melatunan, S.; Calosi, P.; Rundle, S.D.; Widdicombe, S.; Moody, A.J. Effects of ocean acidification and elevated temperature on shell plasticity and its energetic basis in an intertidal gastropod. MEPS 2013, 472, 155–168. [Google Scholar] [CrossRef]
- Thomsen, J.; Casties, I.; Pansch, C.; Körtzinger, A.; Melzner, F. Food availability outweighs ocean acidification effects in juvenile Mytilus edulis: Laboratory and field experiments. Glob. Change Biol. 2013. [Google Scholar] [CrossRef] [Green Version]
- Tomanek, L.; Zuzow, M.J.; Ivanina, A.V.; Beniash, E.; Sokolova, I.M. Proteomic response to elevated PCO2 level in eastern oysters, Crassostrea virginica: Evidence for oxidative stress. J. Exp. Biol. 2011, 214, 1836–1844. [Google Scholar] [CrossRef]
- Gutowska, M.A.; Pörtner, H.O.; Melzner, F. Growth and calcification in the cephalopod Sepia officinalis under elevated seawater pCO2. Mar. Ecol. Prog. Ser. 2008, 373, 303–309. [Google Scholar] [CrossRef]
- Gutowska, M.A.; Melzner, F.; Pörtner, H.O.; Meier, S. Cuttlebone calcification increases during exposure to elevated seawater pCO2 in the cephalopod Sepia officinalis. Mar. Biol. 2010, 157, 1653–1663. [Google Scholar] [CrossRef] [Green Version]
- Ries, J.B. A physicochemical framework for interpreting the biological calcification response to CO2-induced ocean acidification. Geochim. Cosmochim. Acta 2011, 75, 4053–4064. [Google Scholar] [CrossRef]
- Morse, J.W.; Andersson, A.J.; Mackenzie, F.T. Initial responses of carbonate-rich shelf sediments to rising atmospheric pCO2 and “ocean acidification”: Role of high Mg-Calcites. Geochim. Cosmochim. Acta 2006, 70, 5814–5830. [Google Scholar] [CrossRef]
- Gutowska, M.A.; Melzner, F.; Langenbuch, M.; Bock, C.; Claireaux, G.; Pörtner, H.O. Acid-base regulatory ability of the cephalopod (Sepia officinalis) in response to environmental hypercapnia. J. Comp. Physiol. B 2010, 180, 323–335. [Google Scholar] [CrossRef]
- Range, P.; Chícharo, M.; Ben-Hamadou, R.; Piló, D.; Matias, D.; Joaquim, S.; Oliveira, A.; Chícharo, L. Calcification, growth and mortality of juvenile clams Ruditapes decussatus under increased pCO2 and reduced pH: Variable responses to ocean acidification at local scales? J. Exp. Mar. Biol. Ecol. 2011, 396, 177–184. [Google Scholar] [CrossRef]
- Ries, J.B. Skeletal mineralogy in a high CO2 world. J. Exp. Mar. Biol. Ecol. 2011, 403, 54–64. [Google Scholar] [CrossRef]
- Pörtner, H.O.; Langenbuch, M.; Reipschläger, A. Biological impact of elevated ocean CO2 concentrations: Lessons from animal physiology and earth history. J. Oceanogr. 2004, 60, 705–718. [Google Scholar] [CrossRef]
- Pörtner, H.O.; Farrell, A.P. Physiology and climate change. Science 2008, 322, 690–692. [Google Scholar] [CrossRef]
- Kurihara, H.; Kato, S.; Ishimatsu, A. Effects of increased seawater pCO2 on early development of the oyster Crassostrea gigas. Aquat. Biol. 2007, 1, 91–98. [Google Scholar] [CrossRef]
- Havenhand, J.; Schlegel, P. Near-future levels of ocean acidification do not affect sperm motility and fertilization kinetics in the oyster Crassostrea gigas. Biogeosciences 2009, 6, 3009–3015. [Google Scholar] [CrossRef]
- Parker, L.M.; Ross, P.M.; O’Connor, W.A. Comparing the effect of elevated pCO2 and temperature on the fertilization and early development of two species of oysters. Mar. Biol. 2010, 157, 2435–2452. [Google Scholar] [CrossRef]
- Thomsen, J.; Gutowska, M.; Saphörster, J.; Heinemann, A.; Trübenbach, K.; Fietzke, J.; Hiebenthal, C.; Eisenhauer, A.; Körtzinger, A.; Wahl, M. Calcifying invertebrates succeed in a naturally CO2 enriched coastal habitat but are threatened by high levels of future acidification. Biogeosciences 2010, 7, 5119–5156. [Google Scholar] [CrossRef]
- Przeslawski, R. A review of the effects of environmental stress on embryonic development within intertidal gastropod egg masses. Molluscan Res. 2004, 24, 43–63. [Google Scholar] [CrossRef]
- Byrne, M.; Ho, M.; Selvakumaraswamy, P.; Nguyen, H.D.; Dworjanyn, S.A.; Davis, A.R. Temperature, but not pH, compromises sea urchin fertilization and early development under near-future climate change scenarios. Proc. Roy. Soc. B 2009, 276, 1883–1888. [Google Scholar] [CrossRef]
- Gosselin, L.A.; Qian, P.Y. Juvenile mortality in benthic marine invertebrates. Mar. Ecol. Prog. Ser. 1997, 146, 265–282. [Google Scholar] [CrossRef]
- Melzner, F.; Gutowska, M.; Langenbuch, M.; Dupont, S.; Lucassen, M.; Thorndyke, M.C.; Bleich, M.; Pörtner, H.O. Physiological basis for high CO2 tolerance in marine ectothermic animals: Pre-adaptation through lifestyle and ontogeny? Biogeosciences 2009, 6, 2313–2331. [Google Scholar]
- Weiss, I.M.; Tuross, N.; Addadi, L.; Weiner, S. Mollusc larval shell formation: Amorphous calcium carbonate is a precursor phase for aragonite. J. Exp. Zool. 2002, 293, 478–491. [Google Scholar] [CrossRef]
- Dupont, S.; Thorndyke, M. Impact of CO2-driven ocean acidification on invertebrates early life-history—What we know, what we need to know and what we can do. Biogeosciences 2009, 6, 3109–3131. [Google Scholar]
- Gibson, R.; Atkinson, R.; Gordon, J.; Smith, I.; Hughes, D. Impact of ocean warming and ocean acidification on marine invertebrate life history stages: Vulnerabilities and potential for persistence in a changing ocean. Oceanogr. Mar. Biol. Ann. Rev. 2011, 49, 1–42. [Google Scholar]
- Calabrese, A.; Davis, H.C. The pH tolerance of embryos and larvae of Mercenaria mercenaria and Crassostrea virginica. Biol. Bull. 1966, 131, 427–436. [Google Scholar] [CrossRef]
- Desrosiers, R.R.; Desilets, J.; Dube, F. Early developmental events following fertilization in the giant scallop Placopecten magellanicus. Can. J. Fish. Aquat. Sci. 1996, 53, 1382–1392. [Google Scholar] [CrossRef]
- Wilson, S.P.; Hyne, R.V. Toxicity of acid-sulfate soil leachate and aluminum to embryos of the Sydney rock oyster. Ecotoxicol. Environ. Saf. 1997, 37, 30–36. [Google Scholar] [CrossRef]
- Pagano, G.; Cipollaro, M.; Corsale, G.; Esposito, A.; Ragucci, E.; Giordano, G.G. pH-induced changes in mitotic and developmental patterns in sea urchin embryogenesis. I. Exposure of embryos. Teratog. Carcinog. Mutagen. 1985, 5, 101–112. [Google Scholar] [CrossRef]
- Pagano, G.; Cipollaro, M.; Corsale, G.; Esposito, A.; Ragucci, E.; Giordano, G.G. pH-induced changes in mitotic and developmental patterns in sea urchin embryogenesis. II. Exposure of sperm. Teratog. Carcinog. Mutagen. 1985, 5, 113–121. [Google Scholar] [CrossRef]
- Cipollaro, M.; Corsale, G.; Esposito, A.; Ragucci, E.; Staiano, N.; Giordano, G.G.; Pagano, G. Sublethal pH decrease may cause genetic damage to eukaryotic cell: A study on sea urchins and Salmonella typhimurium. Teratog. Carcinog. Mutagen. 1986, 6, 275–287. [Google Scholar] [CrossRef]
- Boyd, P.W. Beyond ocean acidification. Nat. Geosci. 2011, 4, 273–274. [Google Scholar] [CrossRef]
- Bechmann, R.K.; Taban, I.C.; Westerlund, S.; Godal, B.F.; Arnberg, M.; Vingen, S.; Ingvarsdottir, A.; Baussant, T. Effects of ocean acidification on early life stages of shrimp (Pandalus borealis) and mussel (Mytilus edulis). J. Toxicol. Environ. Health A 2011, 74, 424–438. [Google Scholar] [CrossRef]
- Byrne, M.; Soars, N.A.; Ho, M.A.; Wong, E.; McElroy, D.; Selvakumaraswamy, P.; Dworjanyn, S.A.; Davis, A.R. Fertilization in a suite of coastal marine invertebrates from SE Australia is robust to near-future ocean warming and acidification. Mar. Biol. 2010, 157, 2061–2069. [Google Scholar] [CrossRef]
- Van Colen, C.; Debusschere, E.; Braeckman, U.; van Gansbeke, D.; Vincx, M. The early life history of the clam Macoma balthica in a high CO2 world. PLoS One 2012, 7. [Google Scholar] [CrossRef] [Green Version]
- Kimura, R.Y.O.; Takami, H.; Ono, T.; Onitsuka, T.; Nojiri, Y. Effects of elevated pCO2 on the early development of the commercially important gastropod, Ezo abalone Haliotis discus hannai. Fish Oceanogr. 2011, 20, 357–366. [Google Scholar] [CrossRef]
- Byrne, M. Global change ecotoxicology: Identification of early life history bottlenecks in marine invertebrates, variable species responses and variable experimental approaches. Mar. Environ. Res. 2012, 76, 3–15. [Google Scholar] [CrossRef]
- Reuter, K.I.M.E.; Lotterhos, K.E.; Crim, R.N.; Thompson, C.A.; Harley, C.D.G. Elevated pCO2 increases sperm limitation and risk of polyspermy in the red sea urchin Strongylocentrotus franciscanus. Glob. Change Biol. 2010, 17, 163–171. [Google Scholar]
- Miller, A.W.; Reynolds, A.C.; Sobrino, C.; Riedel, G.F. Shellfish face uncertain future in high CO2 world: Influence of acidification on oyster larvae calcification and growth in estuaries. PLoS One 2009, 4, e5661. [Google Scholar] [CrossRef]
- Watson, S.A.; Southgate, P.C.; Tyler, P.A.; Peck, L.S. Early larval development of the Sydney rock oyster Saccostrea glomerata under near-future predictions of CO2-driven ocean acidification. J. Shellfish Res. 2009, 28, 431–437. [Google Scholar]
- Gazeau, F.; Gattuso, J.; Dawber, C.; Pronker, A.; Peene, F.; Peene, J.; Heip, C.; Middelburg, J. Effect of ocean acidification on the early life stages of the blue mussel Mytilus edulis. Biogeosciences 2010, 7, 2051–2060. [Google Scholar]
- Talmage, S.C.; Gobler, C.J. The effects of elevated carbon dioxide concentrations on the metamorphosis, size, and survival of larval hard clams (Mercenaria mercenaria), bay scallops (Argopecten irradians), and eastern oysters (Crassostrea virginica). Limnol. Oceanog. 2009, 54, 2072–2080. [Google Scholar]
- Talmage, S.C.; Gobler, C.J. Effects of past, present, and future ocean carbon dioxide concentrations on the growth and survival of larval shellfis. Proc. Natl. Acad. Sci. USA 2010, 107, 17246–17251. [Google Scholar]
- Comeau, S.; Gorsky, G.; Alliouane, S.; Gattuso, J.P. Larvae of the pteropod Cavolinia inflexa exposed to aragonite undersaturation are viable but shell-less. Mar. Biol. 2010, 157, 2341–2345. [Google Scholar]
- Ellis, R.P.; Bersey, J.; Rundle, S.D.; Hall-Spencer, J.M.; Spicer, J.I. Subtle but significant effects of CO2 acidified seawater on embryos of the intertidal snail, Littorina obtusata. Aquat. Biol. 2009, 5, 41–48. [Google Scholar]
- Gaylord, B.; Hill, T.M.; Sanford, E.; Lenz, E.A.; Jacobs, L.A.; Sato, K.N.; Russell, A.D.; Hettinger, A. Functional impacts of ocean acidification in an ecologically critical foundation species. J. Exp. Biol. 2011, 214, 2586–2594. [Google Scholar]
- Talmage, S.C.; Gobler, C.J. Effects of elevated temperature and carbon dioxide on the growth and survival of larvae and juveniles of three species of Northwest Atlantic bivalves. PLoS One 2011, 6, e26941. [Google Scholar]
- Talmage, S.C.; Gobler, C.J. Effects of CO2 and the harmful alga Aureococcus anophagefferens on growth and survival of oyster and scallop larvae. Mar. Ecol. Prog. Ser. 2012, 464, 121–147. [Google Scholar]
- Thiyagarajan, V.; Ko, G.W.K. Larval growth response of the Portuguese oyster (Crassostrea angulata) to multiple climate change stressors. Aquaculture 2012, 370–371, 90–95. [Google Scholar]
- Timmins-Schiffman, E.; O’Donnell, M.J.; Friedman, C.S.; Roberts, S.B. Elevated pCO2 causes developmental delay in early larval Pacific oysters, Crassostrea gigas. Mar. Biol. 2012. [Google Scholar] [CrossRef]
- Sunday, J.M.; Crim, R.N.; Harley, C.D.G.; Hart, M.W. Quantifying rates of evolutionary adaptation in response to ocean acidification. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- López, I.R.; Kalman, J.; Vale, C.; Blasco, J. Influence of sediment acidification on the bioaccumulation of metals in Ruditapes philippinarum. Environ. Sci. Pollut. R. 2010, 17, 1519–1528. [Google Scholar]
- Riba, I.; García-Luque, E.; Kalman, J.; Blasco, J.; Vale, C. Effects of Sediment Acidification on the Bioaccumulation of Zn in R. Philippinarum. In Oceans and the Atmospheric Carbon Content; Duarte, P., Santana-Casiano, J.M., Eds.; Springer: Berlin, Germany, 2010; pp. 115–132. [Google Scholar]
- Lacoue-Labarthe, T.; Martin, S.; Oberhänsli, F.; Teyssié, J.L.; Markich, S.; Jeffree, R.; Bustamante, P. Effects of increased pCO2 and temperature on trace element (Ag, Cd and Zn) bioaccumulation in the eggs of the common cuttlefish, Sepia officinalis. Biogeosciences 2009, 6, 2561–2573. [Google Scholar]
- Gazeau, F.; Gattuso, J.P.; Greaves, M.; Elderfield, H.; Peene, J.; Heip, C.H.R.; Middelburg, J.J. Effect of carbonate chemistry alteration on the early embryonic development of the Pacific oyster (Crassostrea gigas). PLoS One 2011, 6, e23010. [Google Scholar]
- Goldberg, J.I.; Doran, S.A; Shartau, R.B.; Pon, J.R.; Ali, D.W.; Tam, R; Kuang, S. Integratingbiology of an embryonic respiratory behaviour in pond snails: The embryo stir-bar hypothesis. J. Exp. Biol. 2008, 211, 1729–1736. [Google Scholar]
- Diefenbach, T.J.; Koehncke, N.K.; Goldberg, J.I. Characterization and development of rotational behavior in Helisoma embryos: Role of endogenous serotonin. J. Neurobiol. 1991, 22, 922–934. [Google Scholar]
- Waller, T.R. Functional Morphology and Development of Veliger Larvae of the European Oyster, Ostrea Edulis Linne; Smithsonian Institution Press: Washington, DC, USA, 1981; pp. 1–70. [Google Scholar]
- Hayakaze, E.; Tanabe, K. Early larval shell development in mytilid bivalve Mytilus galloprovincialis. Venus Jpn. J. Malacol. 1999, 58, 119–127. [Google Scholar]
- O’Donnell, M.J.; Hammond, L.T.M.; Hofmann, G.E. Predicted impact of ocean acidification on a marine invertebrate: Elevated CO2 alters response to thermal stress in sea urchin larvae. Mar. Biol. 2009, 156, 439–446. [Google Scholar]
- Todgham, A.E.; Hofmann, G.E. Transcriptomic response of sea urchin larvae Strongylocentrotus purpuratus to CO2-driven seawater acidification. J. Exp. Biol. 2009, 212, 2579–2594. [Google Scholar]
- Zippay, M.K.L.; Hofmann, G.E. Effect of pH on gene expression and thermal tolerance of early life history stages of red abalone (Haliotis rufescens). J. Shellfish Res. 2010, 29, 429–439. [Google Scholar]
- Dineshram, R.; Wong, K.K.W.; Xiao, S.; Yu, Z.; Qian, P.Y.; Thiyagarajan, V. Analysis of Pacific oyster larval proteome and its response to high-CO2. Mar. Pollut. Bull. 2012, 64, 2160–2167. [Google Scholar]
- Rodriguez, S.R.; Ojeda, F.P.; Inestrosa, N.C. Settlement of benthic marine invertebrates. Mar. Ecol. Prog. Ser. 1993, 97, 193–207. [Google Scholar]
- Cigliano, M.; Gambi, M.; Rodolfo-Metalpa, R.; Patti, F.; Hall-Spencer, J. Effects of ocean acidification on invertebrate settlement at volcanic CO2 vents. Mar. Biol. 2010, 157, 2489–2502. [Google Scholar]
- Byrne, M.; Selvakumaraswamy, P.; Ho, M.; Woolsey, E.; Nguyen, H. Sea urchin development in a global change hotspot, potential for southerly migration of thermotolerant propagules. Deep Sea Res. Part II 2011, 58, 712–719. [Google Scholar]
- Caldeira, K.; Wickett, M.E. Anthropogenic carbon and ocean pH. Nature 2003, 425, 365–365. [Google Scholar]
- Howarth, R.; Chan, F.; Conley, D.J.; Garnier, J.; Doney, S.C.; Marino, R.; Billen, G. Coupled biogeochemical cycles: Eutrophication and hypoxia in temperate estuaries and coastal marine ecosystems. Front. Ecol. Environ. 2011, 9, 18–26. [Google Scholar]
- Reynaud, S.; Leclercq, N.; Romaine‐Lioud, S.; Ferrier‐Pagés, C.; Jaubert, J.; Gattuso, J.P. Interacting effects of CO2 partial pressure and temperature on photosynthesis and calcification in a scleractinian coral. Glob. Change Biol. 2003, 9, 1660–1668. [Google Scholar]
- Metzger, R.; Sartoris, F.J.; Langenbuch, M.; Pörtner, H.O. Influence of elevated CO2 concentrations on thermal tolerance of the edible crab Cancer pagurus. J. Therm. Biol. 2007, 32, 144–151. [Google Scholar]
- Anthony, K.; Kline, D.; Diaz-Pulido, G.; Dove, S.; Hoegh-Guldberg, O. Ocean acidification causes bleaching and productivity loss in coral reef builders. Proc. Natl. Acad. Sci. USA 2008, 105, 17442. [Google Scholar]
- Gooding, R.A.; Harley, C.D.G.; Tang, E. Elevated water temperature and carbon dioxide concentration increase the growth of a keystone echinoderm. Proc. Natl. Acad. Sci. USA 2009, 106, 9316–9321. [Google Scholar]
- Martin, S.; Gattuso, J.P. Response of mediterranean coralline algae to ocean acidification and elevated temperature. Glob. Change Biol. 2009, 15, 2089–2100. [Google Scholar]
- Matozzo, V.; Chinellato, A.; Munari, M.; Finos, L.; Bressan, M.; Marin, M.G. First evidence of immunomodulation in bivalves under seawater acidification and increased temperature. PLoS One 2012, 7, e33820. [Google Scholar]
- Rosa, R.; Seibel, B.A. Synergistic effects of climate-related variables suggest future physiological impairment in a top oceanic predator. Proc. Natl. Acad. Sci. USA 2008, 105, 20776–20780. [Google Scholar]
- Lannig, G.; Eilers, S.; Pörtner, H.O.; Sokolova, I.M.; Bock, C. Impact of ocean acidification on energy metabolism of oyster, Crassostrea gigas—Changes in metabolic pathways and thermal response. Mar. Drugs 2010, 8, 2318–2339. [Google Scholar]
- Hale, R.; Calosi, P.; McNeill, L.; Mieszkowska, N.; Widdicombe, S. Predicted levels of future ocean acidification and temperature rise could alter community structure and biodiversity in marine benthic communities. Oikos 2011, 120, 661–674. [Google Scholar]
- Byrne, M.; Soars, N.; Selvakumaraswamy, P.; Dworjanyn, S.A.; Davis, A.R. Sea urchin fertilization in a warm, acidified and high pCO2 ocean across a range of sperm densities. Mar. Environ. Res. 2010, 69, 234–239. [Google Scholar]
- Dickinson, G.H.; Ivanina, A.V.; Matoo, O.B.; Pörtner, H.O.; Lannig, G.; Bock, C.; Beniash, E.; Sokolova, I.M. Interactive effects of salinity and elevated CO2 levels on juvenile eastern oysters, Crassostrea virginica. J. Exp. Biol. 2012, 215, 29–43. [Google Scholar]
- Pörtner, H.O. Oxygen-and capacity-limitation of thermal tolerance: A matrix for integrating climate-related stressor effects in marine ecosystems. J. Exp. Biol. 2010, 213, 881–893. [Google Scholar]
- Przeslawski, R.; Davis, A.; Benkendorff, K. Synergistic effects associated with climate change and the development of rocky shore molluscs. Glob. Change Biol. 2005, 11, 515–522. [Google Scholar]
- McLusky, D.S.; Bryant, V.; Campbell, R. The effects of temperature and salinity on the toxicity of heavy metals to marine and estuarine invertebrates. Oceanogr. Mar. Biol. Ann. Rev. 1986, 24, 481–520. [Google Scholar]
- Bibby, R.; Widdicombe, S.; Parry, H.; Spicer, J.; Pipe, R. Effect of ocean acidification on the immune response of the blue mussel, Mytilus edulis. Aquat. Biol. 2008, 2, 67–74. [Google Scholar]
- Beesley, A.; Lowe, D.; Pascoe, C.; Widdicombe, S. Effects of CO2-induced seawater acidification on the health of Mytilus edulis. Clim. Res. 2008, 37, 215–225. [Google Scholar]
- Davis, H.C. Survival and growth of clam and oyster larvae at different salinities. Biol. Bull. 1958, 114, 296–307. [Google Scholar]
- Podolsky, R.D.; Moran, A.L. Integrating function across marine life cycles. ICB 2006, 46, 577–586. [Google Scholar]
- Allen, R.M.; Buckley, Y.M.; Marshall, D.J. Offspring size plasticity in response to intraspecific competition: An adaptive maternal effect across life-history stages. Am. Nat. 2008, 171, 225–237. [Google Scholar]
- Moran, A.L.; McAlister, J.S. Egg size as a life history character of marine invertebrates: Is it all it's cracked up to be? Biol. Bull. 2009, 216, 226–242. [Google Scholar]
- Dupont, S.; Dorey, N.; Stumpp, M.; Melzner, F.; Thorndyke, M. Long-term and trans-life-cycle effects of exposure to ocean acidification in the green sea urchin Strongylocentrotus droebachiensis. Mar. Biol. 2012. [Google Scholar] [CrossRef] [Green Version]
- Reipschläger, A.; Pörtner, H. Metabolic depression during environmental stress: The role of extracellular versus intracellular pH in Sipunculus nudus. J. Exp. Biol. 1996, 199, 1801–1807. [Google Scholar]
- Cummings, V.; Hewitt, J.; van Rooyen, A.; Currie, K.; Beard, S.; Thrush, S.; Norkko, J.; Barr, N.; Heath, P.; Halliday, N.J. Ocean acidification at high latitudes: Potential effects on functioning of the antarctic bivalve Laternula elliptica. PLoS One 2011, 6, e16069. [Google Scholar] [CrossRef]
- Green, M.A.; Waldbusser, G.G.; Reilly, S.L.; Emerson, K. Death by dissolution: Sediment saturation state as a mortality factor for juvenile bivalves. Limnol. Oceanogr. 2009, 54, 1037–1047. [Google Scholar] [CrossRef]
- Seibel, B.A.; Walsh, P.J. Biological impacts of deep-sea carbon dioxide injection inferred from indices of physiological performance. J. Exp. Biol. 2003, 206, 641–650. [Google Scholar] [CrossRef]
- Marchant, H.K.; Calosi, P.; Spicer, J.I. Short-term exposure to hypercapnia does not compromise feeding, acid-base balance or respiration of Patella vulgata but surprisingly is accompanied by radula damage. J. Mar. Biol. Assoc. UK 2010, 90, 1379–1384. [Google Scholar] [CrossRef]
- Pörtner, H.O. Ecosystem effects of ocean acidification in times of ocean warming: A physiologist’s view. Mar. Ecol. Prog. Ser. 2008, 373, 203–217. [Google Scholar] [CrossRef]
- Fernández-Reiriz, J.; Range, P.; Álvarez-Salgado, X.A.; Labarta, U. Physiological energetics of juvenile clams (Ruditapes decussatus) in a high CO2 coastal ocean. Mar. Ecol. Prog. Ser. 2011, 433, 97–105. [Google Scholar]
- Liu, W.; He, M. Effects of ocean acidification on the metabolic rates of three species of bivalve from southern coast of China. Chin. J. Oceanol. Limn. 2012, 30, 206–211. [Google Scholar] [CrossRef]
- Fernández-Reiriz, M.J.; Range, P.; Alvarez-Salgado, X.A.; Espinosa, J.; Labarta, U. Tolerance of juvenile Mytilus galloprovincialis to experimental seawater acidification. Mar. Ecol. Prog. Ser. 2012, 454, 65–74. [Google Scholar] [CrossRef]
- Navarro, J.M.; Torres, R.; Acuña, K.; Duarte, C.; Manriquez, P.H.; Lardies, M.; Lagos, N.A.; Vargas, C.; Aguilera, V. Impact of medium-term exposure to elevated pCO2 levels on the physiological energetics of the mussel Mytilus chilensis. Chemosphere 2013. [Google Scholar] [CrossRef]
- Hammer, K.M.; Kristiansen, E.; Zachariassen, K.E. Physiological effects of hypercapnia in the deep-sea bivalve Acesta excavata (Fabricius, 1779) (Bivalvia; Limidae). Mar. Environ. Res. 2011, 72, 135–142. [Google Scholar] [CrossRef]
- Melatunan, S.; Calosi, P.; Rundle, S.D.; Moody, A.J.; Widdicombe, S. Exposure to elevated temperature and PCO2 reduces respiration rate and energy status in the periwinkle Littorina littorea. Physiol. Biochem. Zool. 2011, 84, 583–594. [Google Scholar] [CrossRef]
- Sokolova, I.; Pörtner, H. Temperature effects on key metabolic enzymes in Littorina saxatilis and L. obtusata from different latitudes and shore levels. Mar. Biol. 2001, 139, 113–126. [Google Scholar] [CrossRef] [Green Version]
- Lindinger, M.; Lauren, D.; McDonald, D. Acid-base balance in the sea mussel, Mytilus edulis. III: Effects of environmental hypercapnia on intra-and extracellular acid-base balance. Mar. Biol. Lett. 1984, 5, 371–381. [Google Scholar]
- Schalkhausser, B.; Bock, C.; Stemmer, K.; Brey, T.; Pörtner, H.-O.; Lannig, G. Impact of ocean acidification on escape performance of the king scallop, Pecten maximus, from Norway. Mar. Biol. 2012. [Google Scholar] [CrossRef] [Green Version]
- Melzner, F.; Stange, P.; Trübenbach, K.; Thomsen, J.; Casties, I.; Panknin, U.; Gorb, S.N.; Gutowska, M.A. Food supply and seawater pCO2 impact calcification and internal shell dissolution in the blue mussel Mytilus edulis. PLoS One 2011, 6, e24223. [Google Scholar]
- Chassot, E.; Bonhommeau, S.; Dulvy, N.K.; Mélin, F.; Watson, R.; Gascuel, D.; Le Pape, O. Global marine primary production constrains fisheries catches. Ecol. Lett. 2010, 13, 495–505. [Google Scholar] [CrossRef]
- Anger, K. The D0 threshold: A critical point in the larval development of decapod crustaceans. J. Exp. Mar. Biol. Ecol. 1987, 108, 15–30. [Google Scholar] [CrossRef]
- Strathmann, R. Larval Feeding. In Reproduction of Marine Invertebrates. General Aspects, Seeking Unity in Diversity; Giese, A.C., Pearse, A.C., Pearse, J.S., Eds.; Blackwell Scientific Publishing: San Francisco, CA, USA, 1987; Volume 9, pp. 465–550. [Google Scholar]
- Hart, M.W.; Strathmann, R.R. Functional consequences of phenotypic plasticity in echinoid larvae. Biol. Bull. 1994, 186, 291–299. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Parker, L.M.; Ross, P.M.; O'Connor, W.A.; Pörtner, H.O.; Scanes, E.; Wright, J.M. Predicting the Response of Molluscs to the Impact of Ocean Acidification. Biology 2013, 2, 651-692. https://doi.org/10.3390/biology2020651
Parker LM, Ross PM, O'Connor WA, Pörtner HO, Scanes E, Wright JM. Predicting the Response of Molluscs to the Impact of Ocean Acidification. Biology. 2013; 2(2):651-692. https://doi.org/10.3390/biology2020651
Chicago/Turabian StyleParker, Laura M., Pauline M. Ross, Wayne A. O'Connor, Hans O. Pörtner, Elliot Scanes, and John M. Wright. 2013. "Predicting the Response of Molluscs to the Impact of Ocean Acidification" Biology 2, no. 2: 651-692. https://doi.org/10.3390/biology2020651