Smooth Muscle Cell Functionality on Collagen Immobilized Polycaprolactone Nanowire Surfaces



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
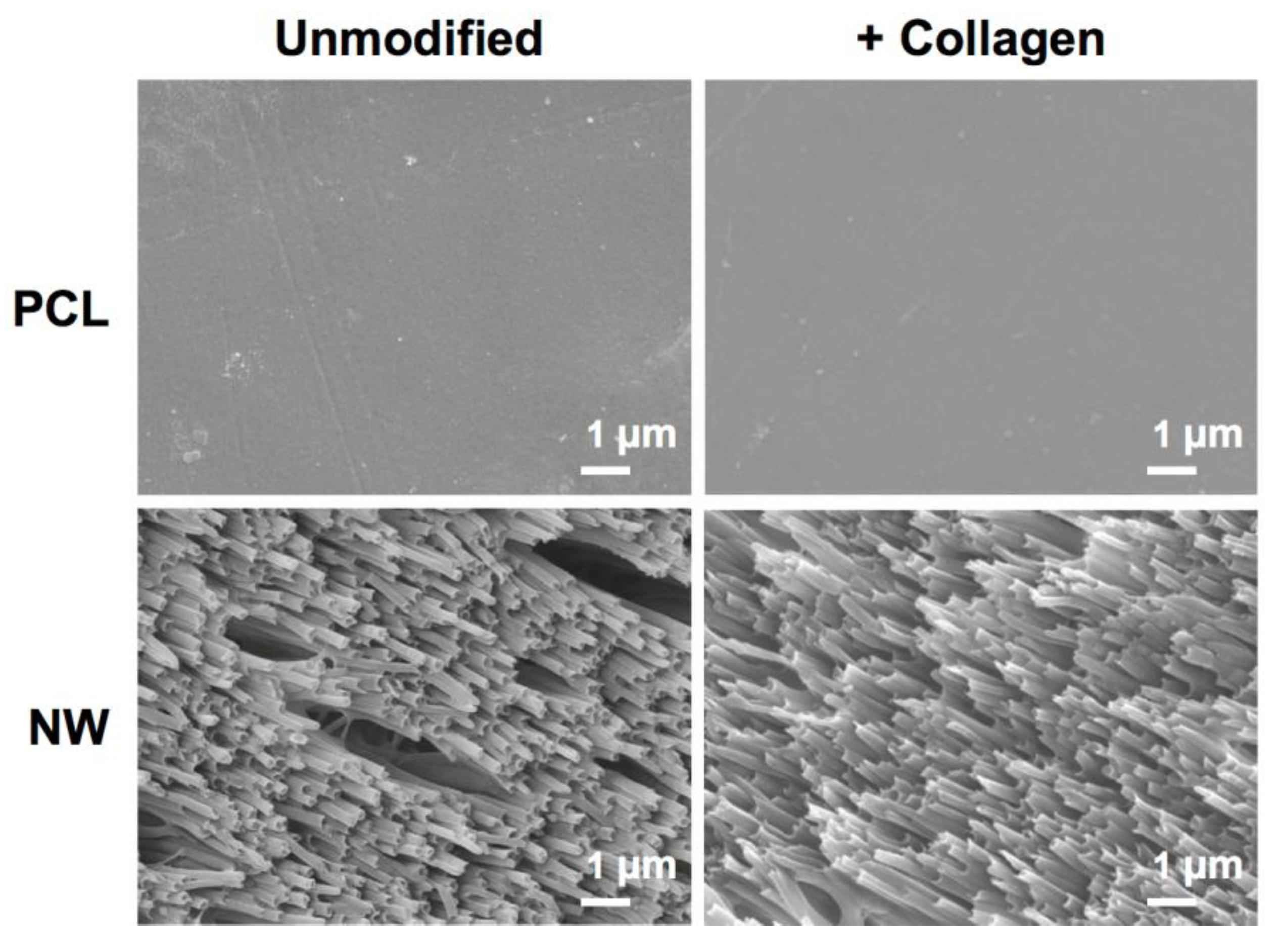
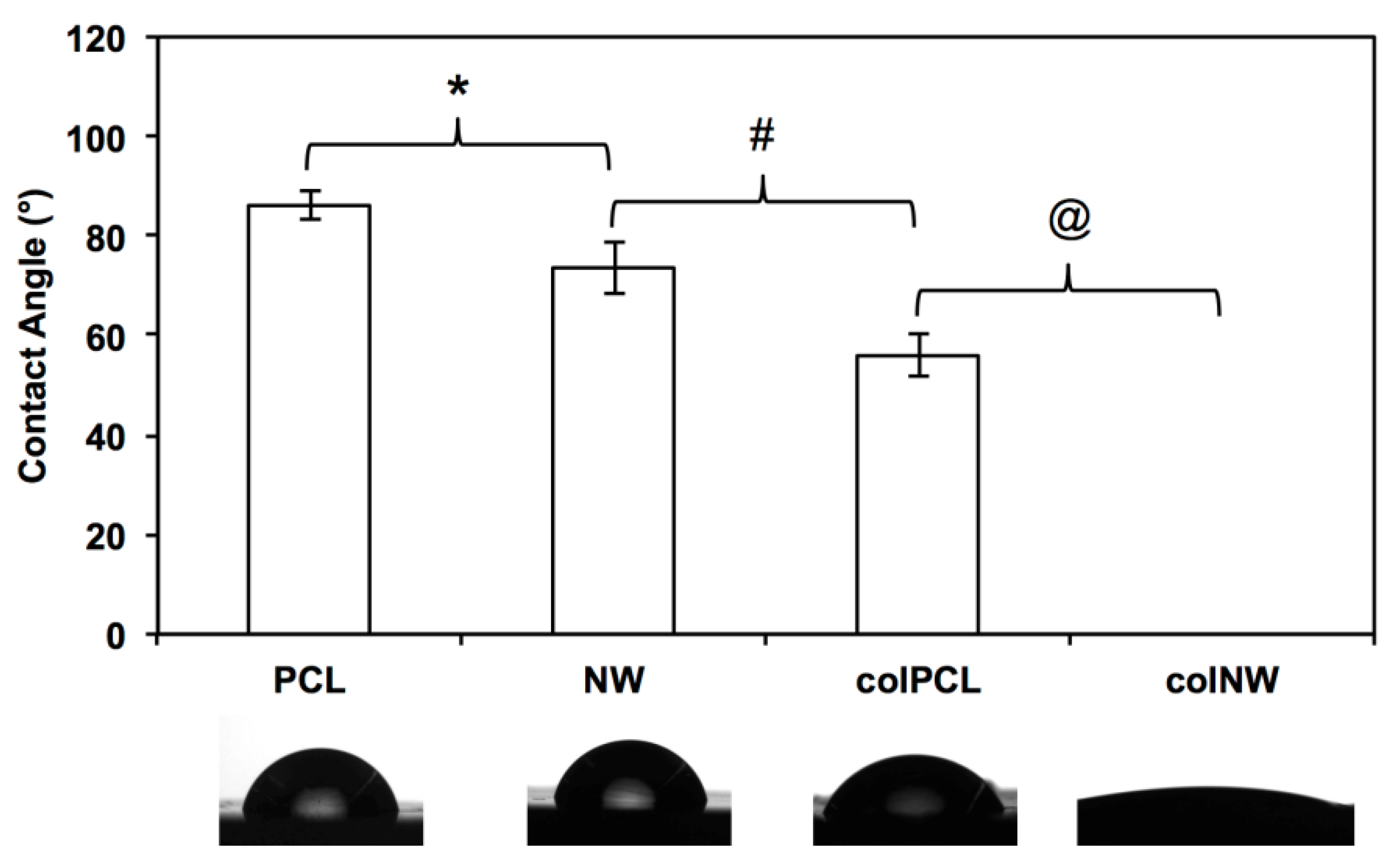
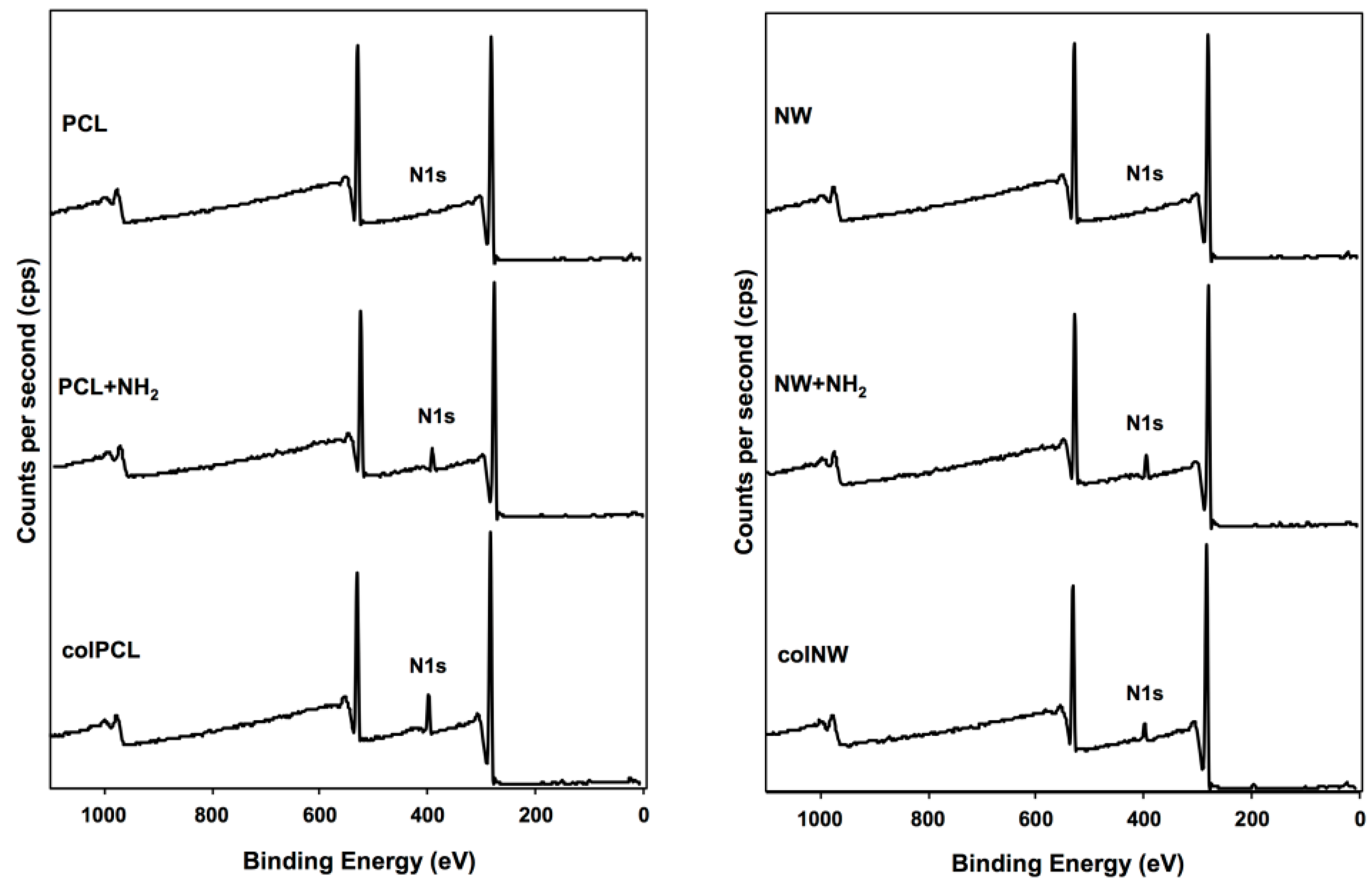
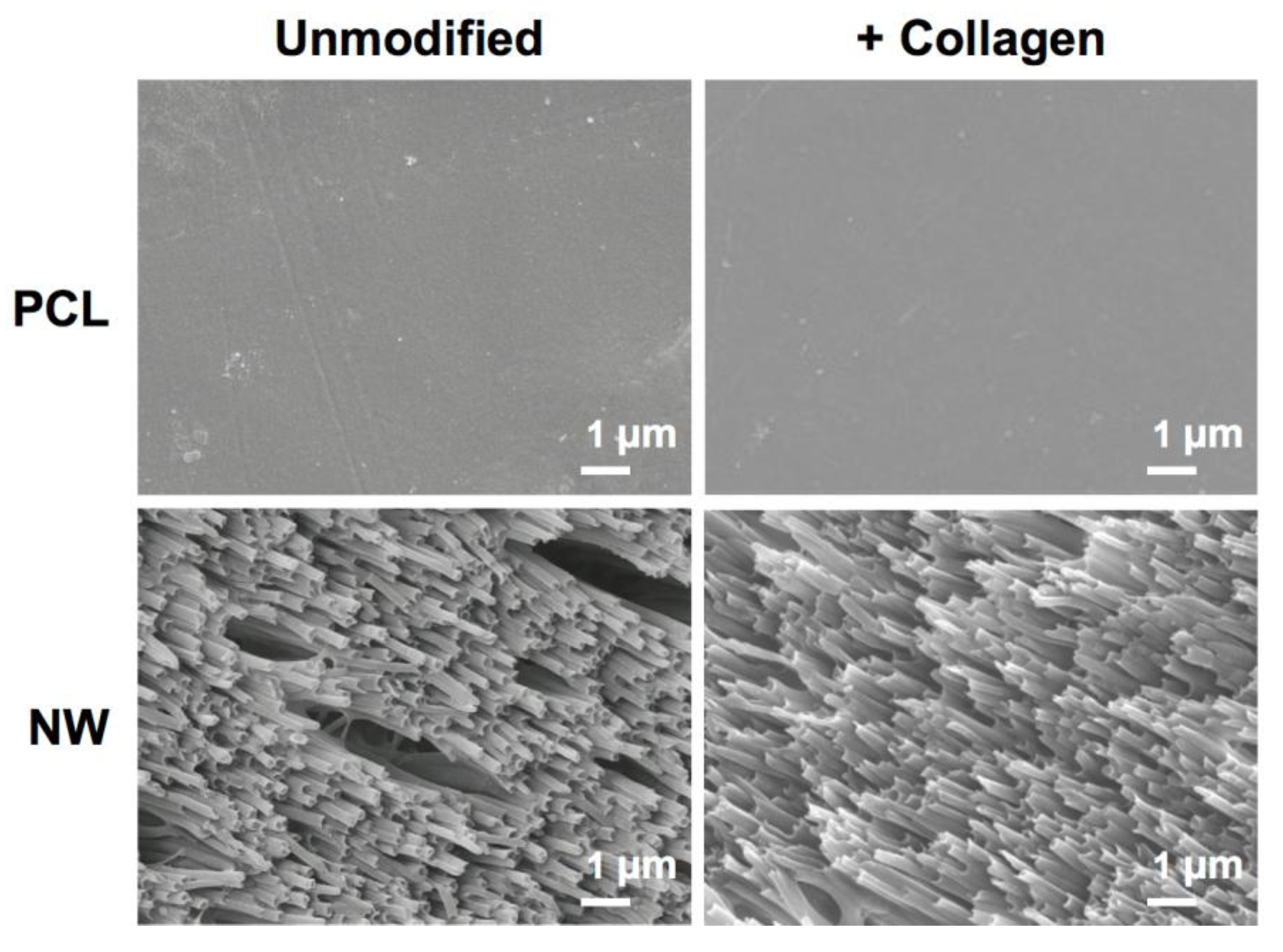
2.1. Characterization of Surfaces


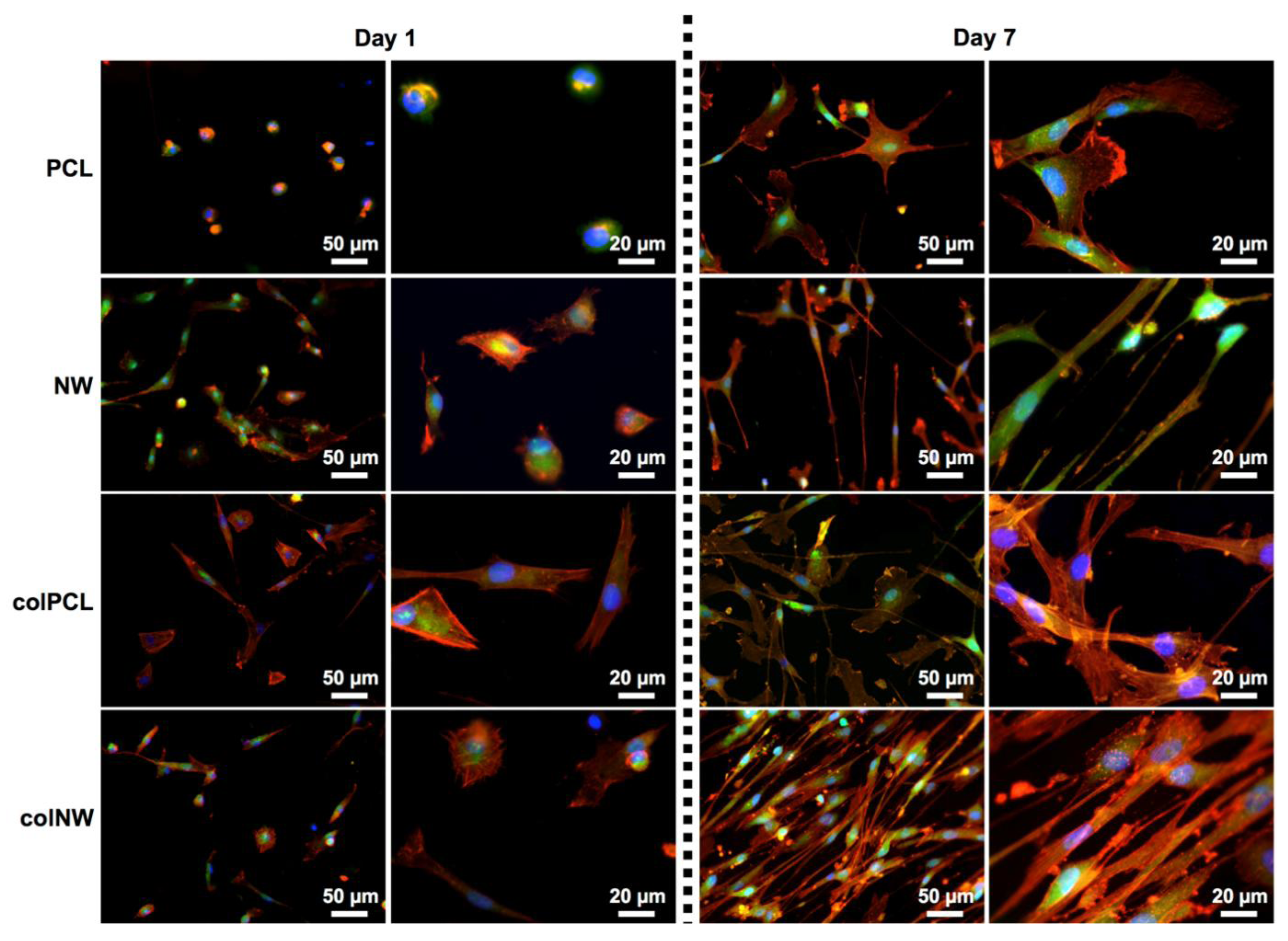
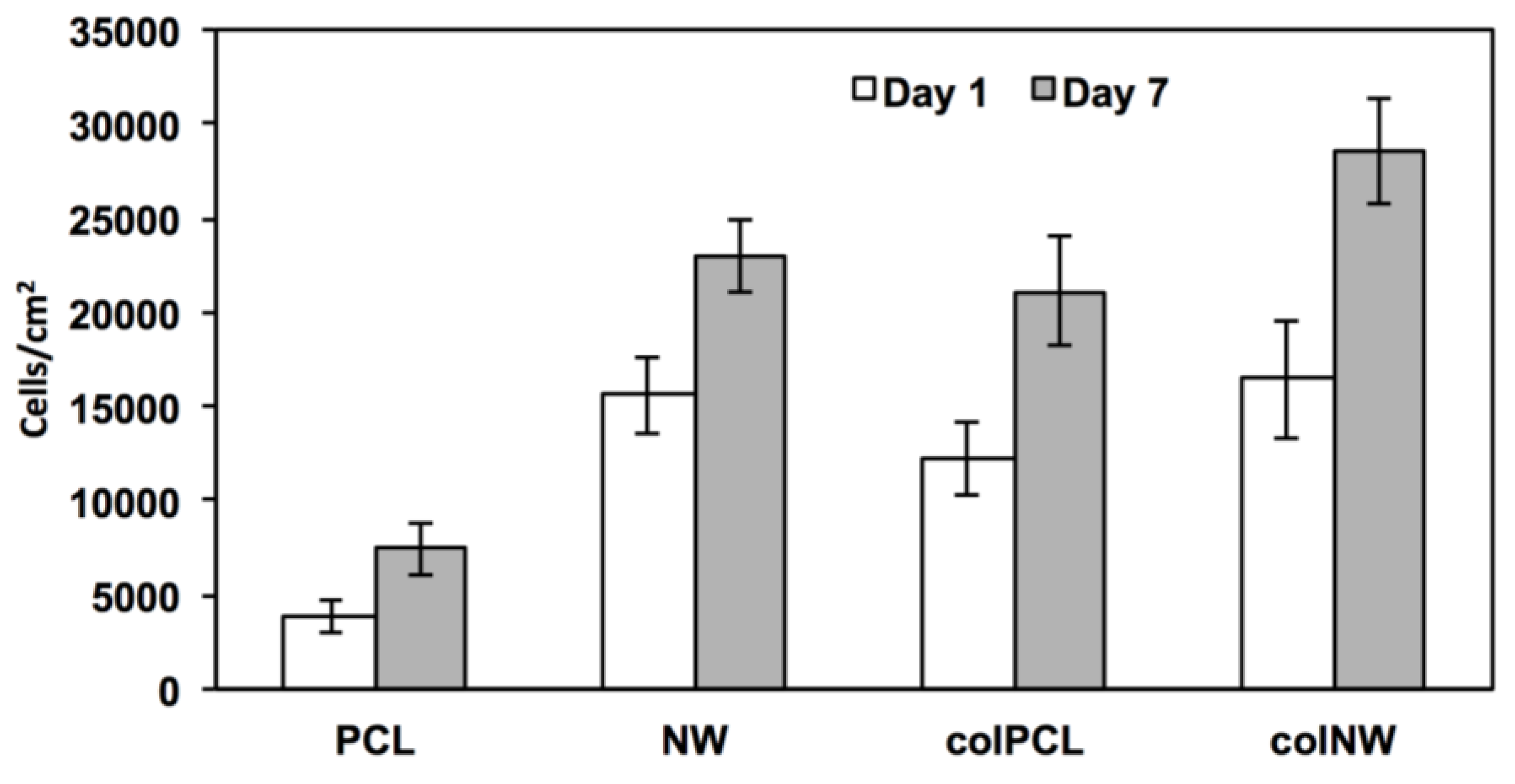
2.2. Adhesion and Proliferation of SMCs


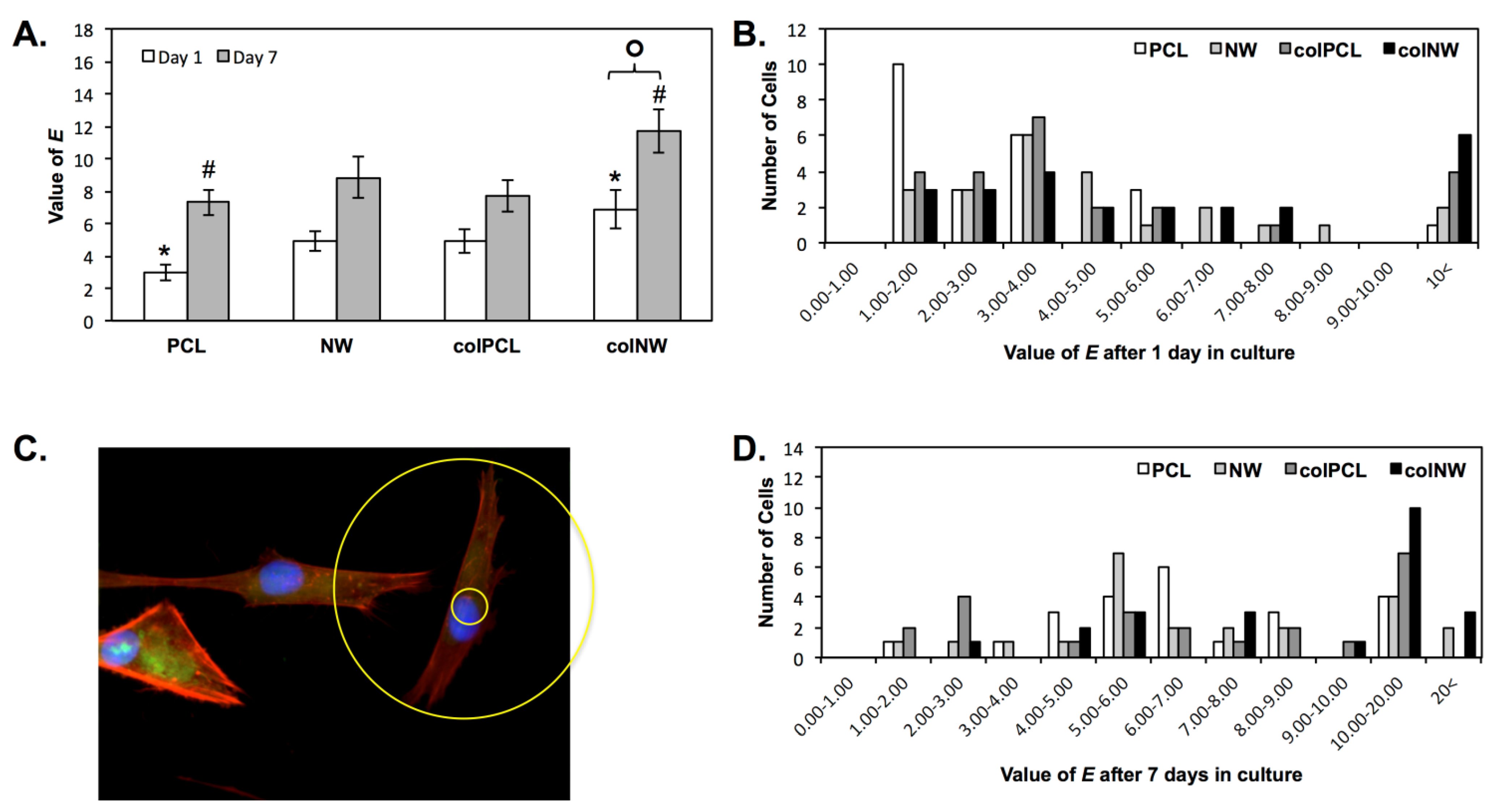
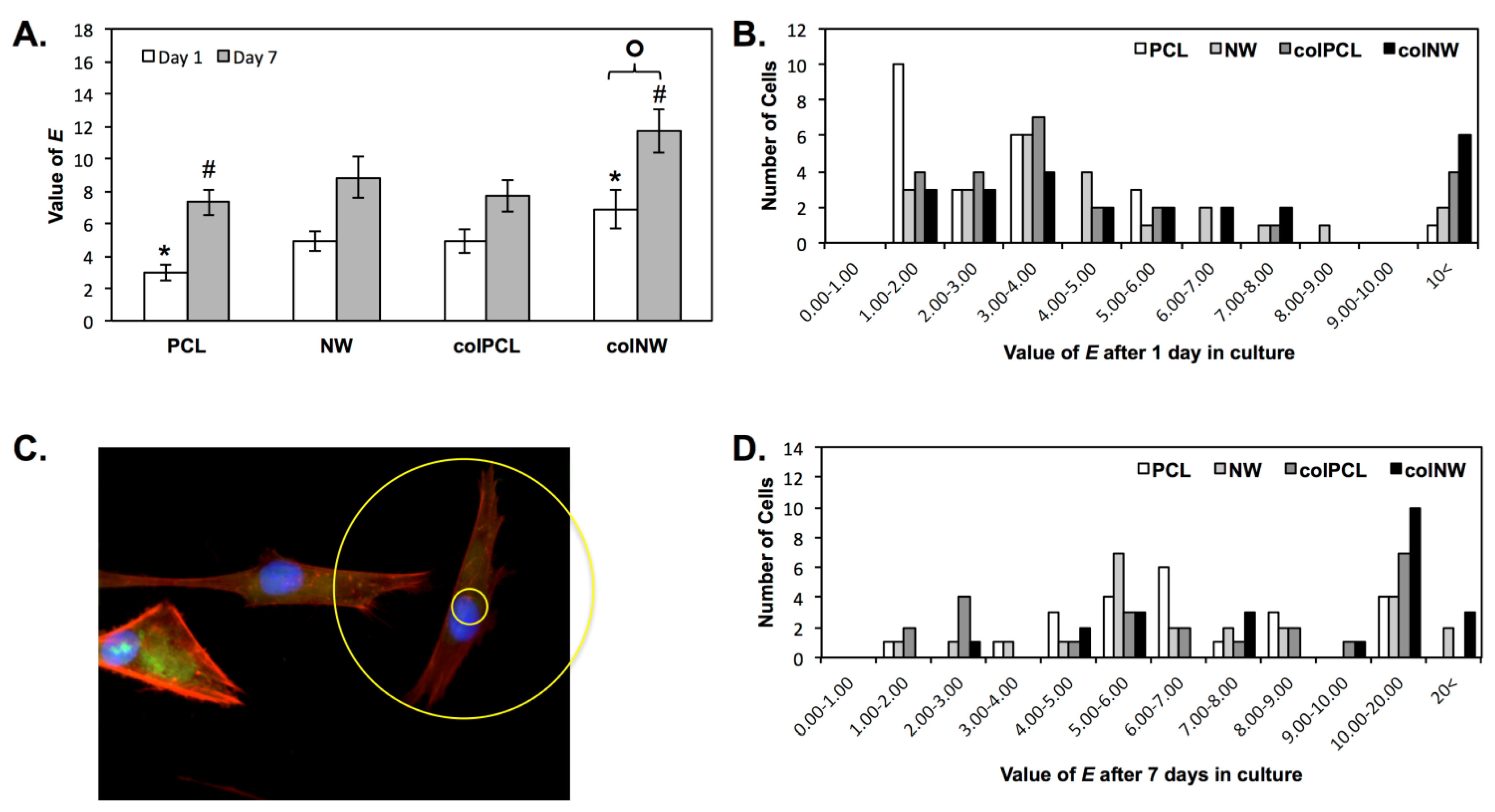
2.3. SMC Elongation on Different Surfaces
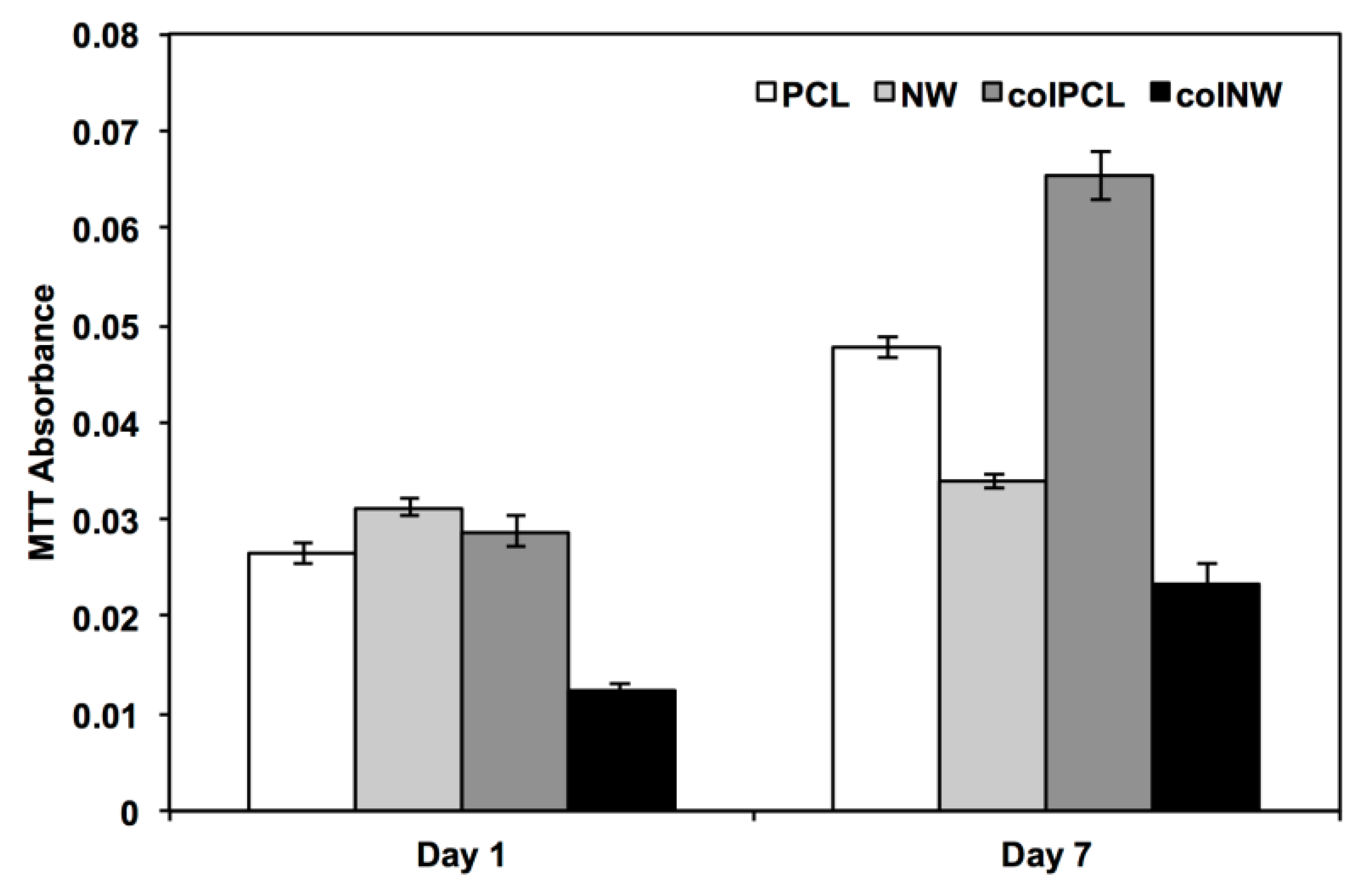
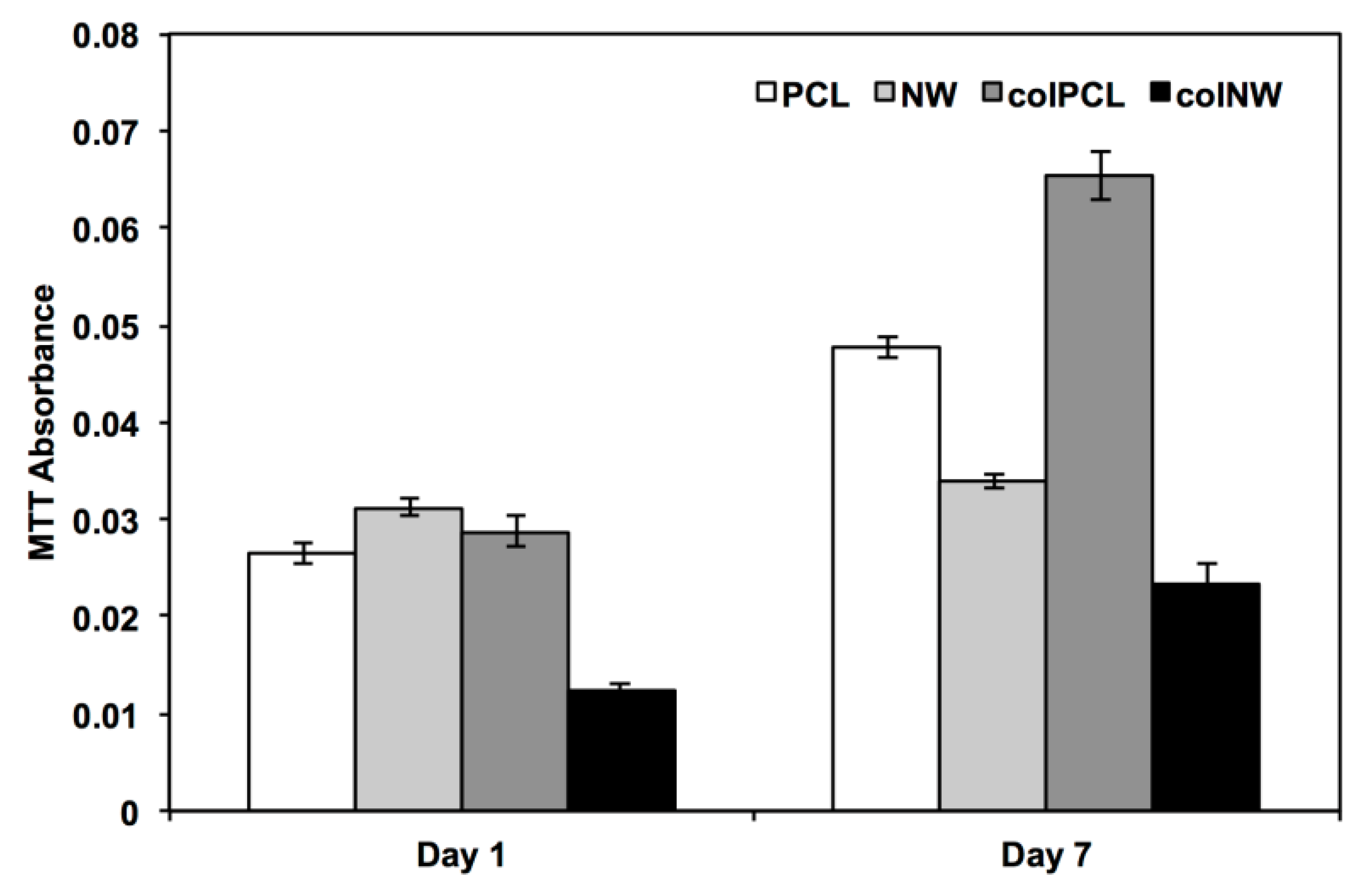
2.4. Viability of SMCs on Surfaces
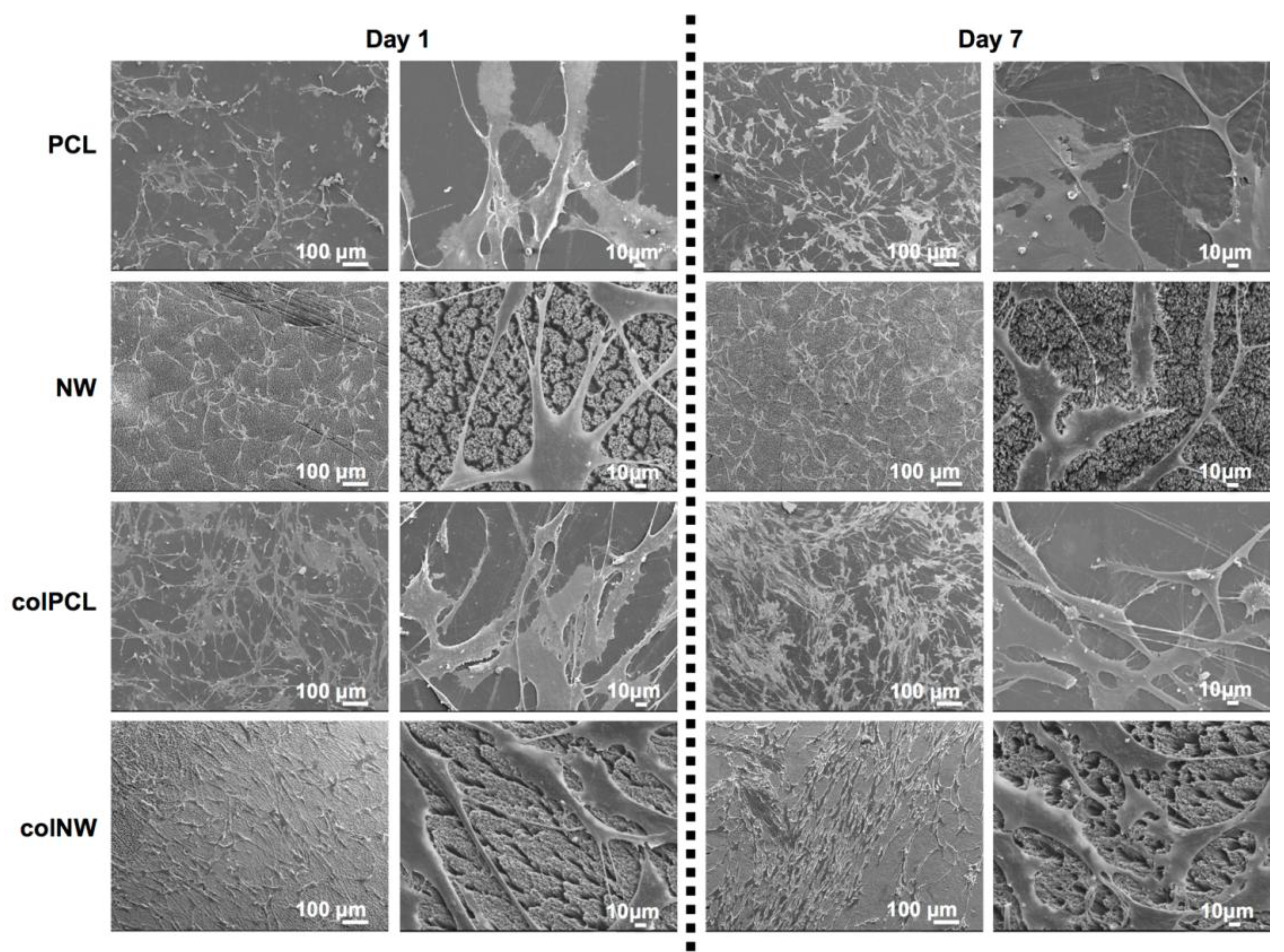
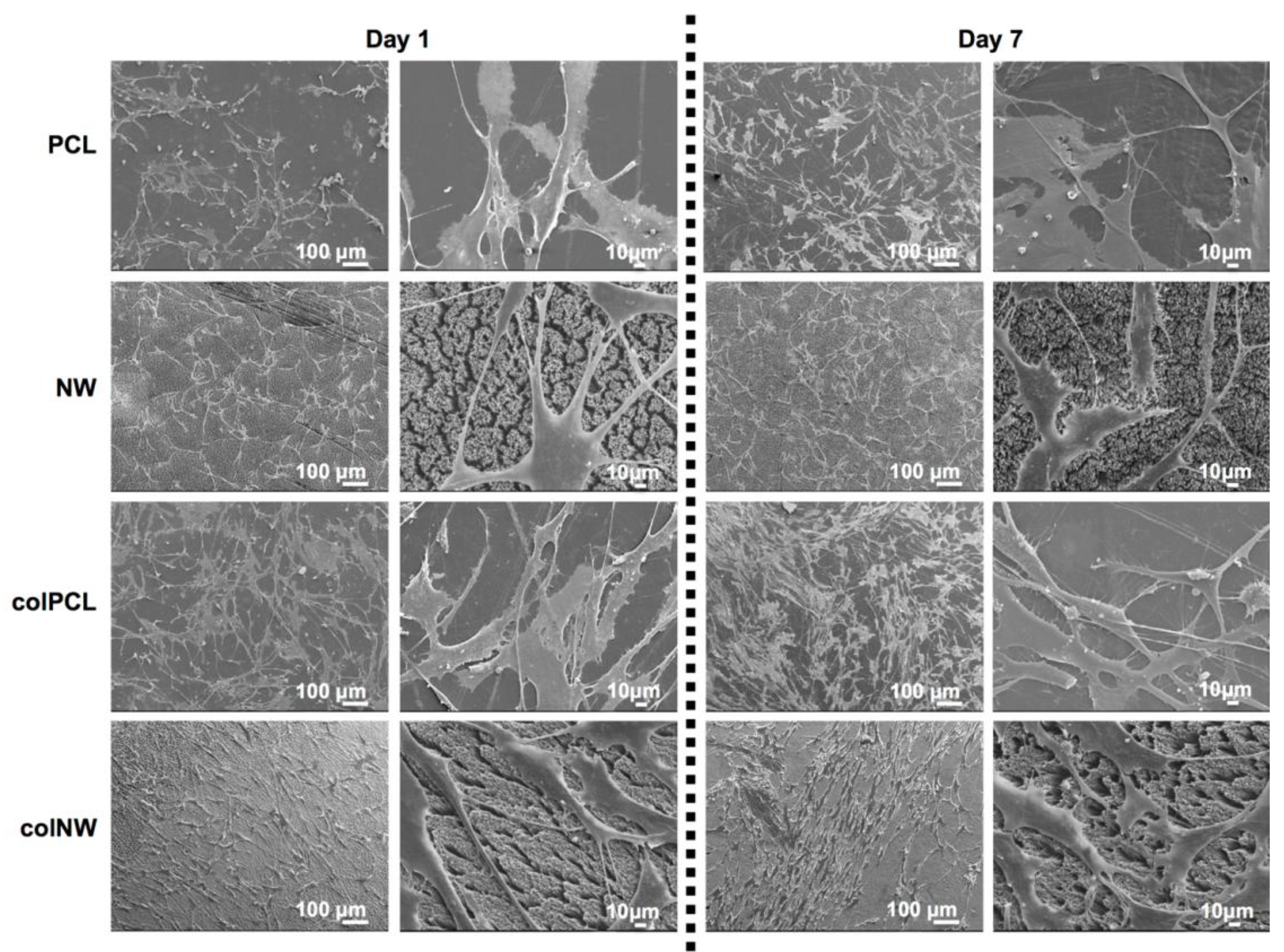
2.5. SMC Morphology on Different Surfaces
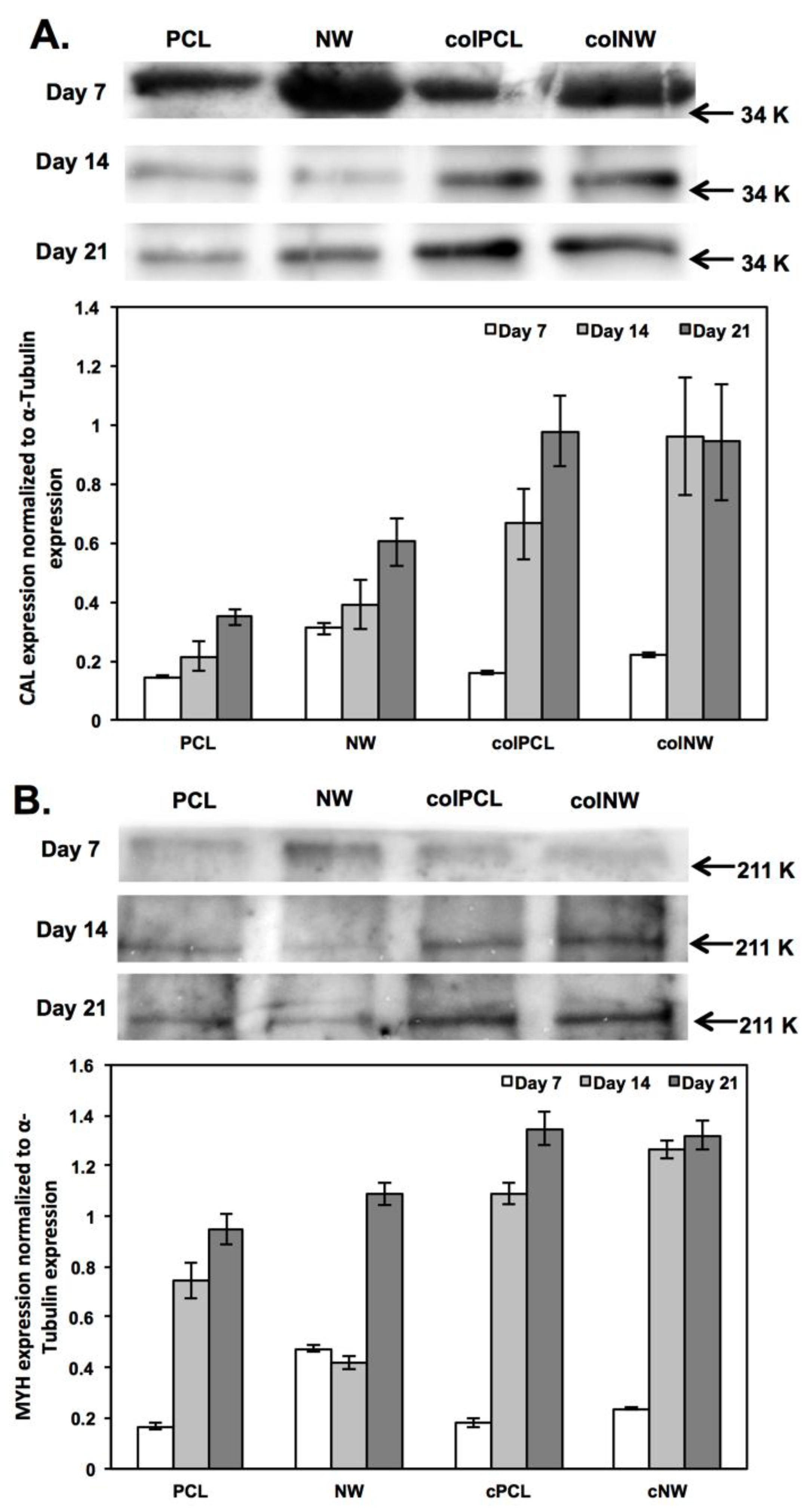
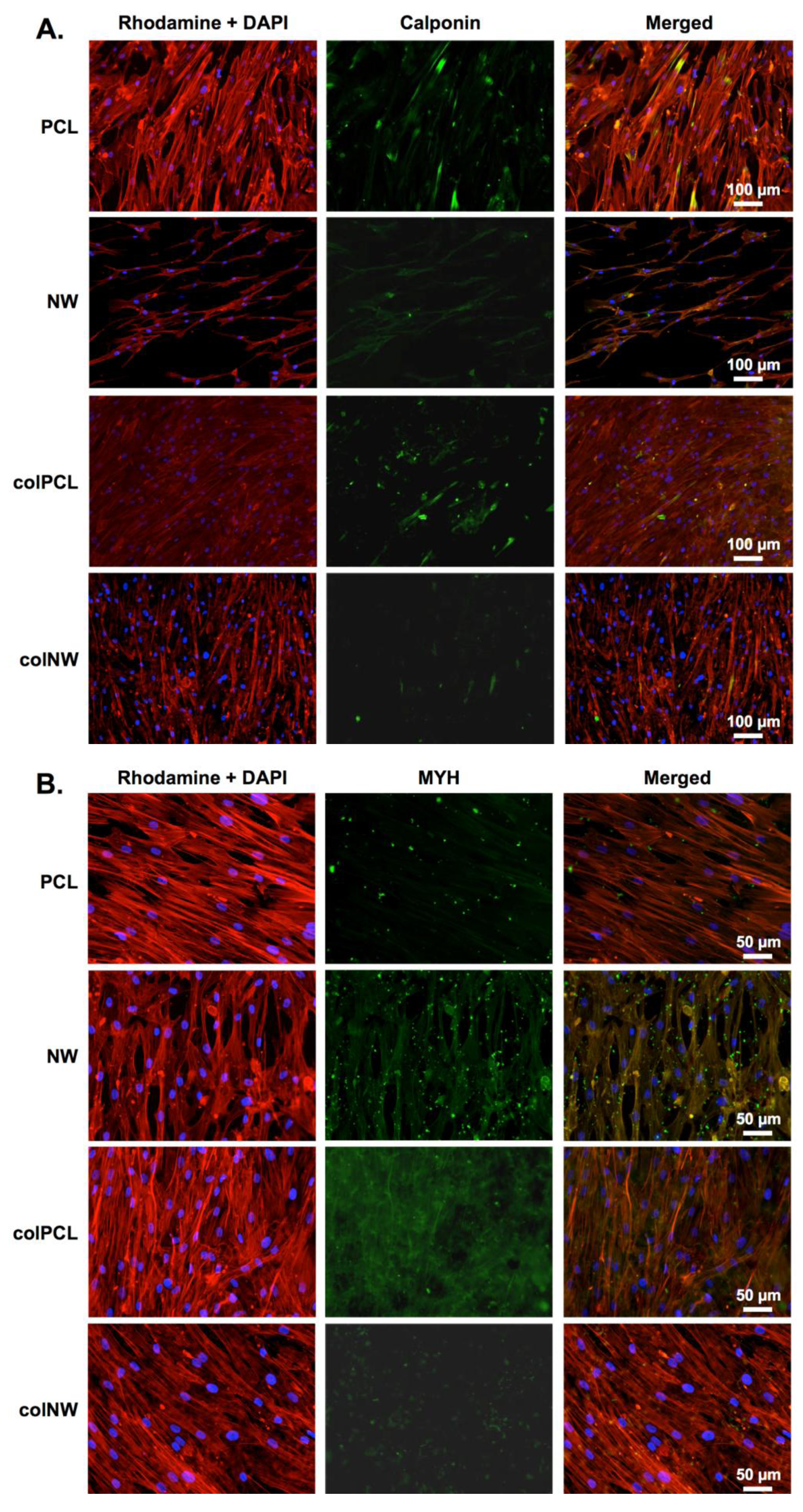
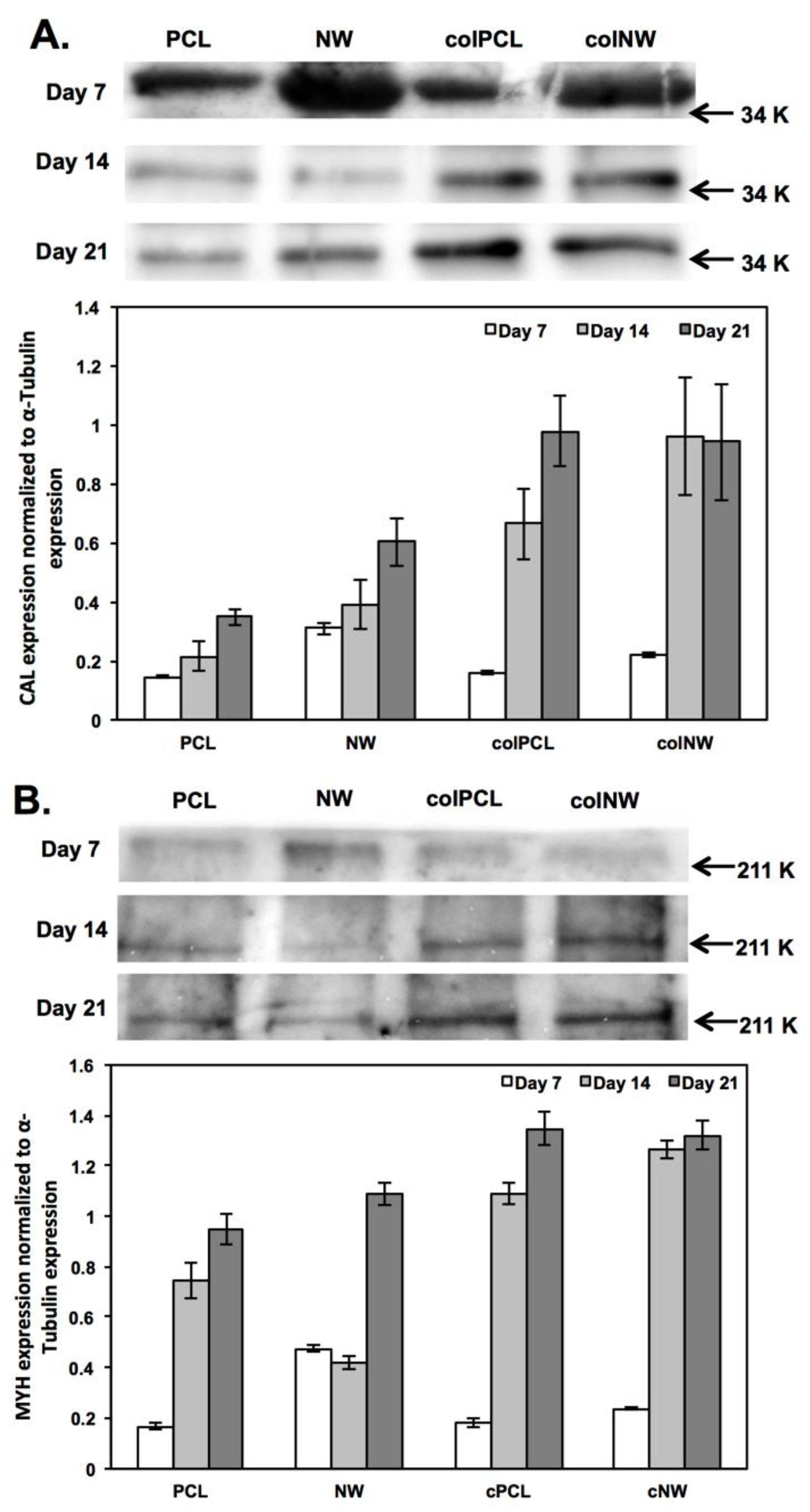
2.6. Differentiation of SMCs on Different Surfaces

3. Experimental Section
3.1. Fabrication of PCL Nanowire Surfaces
3.2. Immobilization of Collagen on PCL and NW Surfaces
3.3. Characterization of colPCL and colNW Surfaces
3.4. Smooth Muscle Cell Culture
3.5. Adhesion and Proliferation of SMCs on Different Surfaces
3.6. SMC Elongation on Different Surfaces
3.7. Viability of SMCs on Different Surfaces
3.8. Morphology of SMCs on Different Surfaces
3.9. Differentiation of SMCs on Different Surfaces
3.10. Statistics
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Owens, G.K.; Kumar, M.S.; Wamhoff, B.R. Molecular regulation of vascular smooth muscle cell differentiation in development and disease. Physiol. Rev. 2004, 84, 767–801. [Google Scholar] [CrossRef]
- Alexander, M.R.; Owens, G.K. Epigenetic control of smooth muscle cell differentiation and phenotypic switching in vascular development and disease. Ann. Rev. Physiol. 2012, 74, 13–40. [Google Scholar] [CrossRef]
- Halayko, A.J.; Solway, J. Molecular mechanisms of phenotypic plasticity in smooth muscle cells. J. Appl. Physiol. 2001, 90, 358–368. [Google Scholar]
- Owens, G.K. Molecular control of vascular smooth muscle cell differentiation and phenotypic plasticity. Novartis Foundation Symp. 2007, 283, 174–191. [Google Scholar] [CrossRef]
- Johnson, J.L.; van Eys, G.J.; Angelini, G.D.; George, S.J. Injury induces dedifferentiation of smooth muscle cells and increased matrix-degrading metalloproteinase activity in human saphenous vein. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 1146–1151. [Google Scholar] [CrossRef]
- Doevendans, P.A.; van Eys, G. Smooth muscle cells on the move: The battle for actin. Cardiovasc. Res. 2002, 54, 499–502. [Google Scholar] [CrossRef]
- Hehrlein, C.; Gollan, C.; Dönges, K.; Metz, J.; Riessen, R.; Fehsenfeld, P.; von Hodenberg, E.; Kübler, W. Low-dose radioactive endovascular stents prevent smooth muscle cell proliferation and neointimal hyperplasia in rabbits. Circulation 1995, 92, 1570–1575. [Google Scholar] [CrossRef]
- Rensen, S.S.; Doevendans, P.A.; van Eys, G.J. Regulation and characteristics of vascular smooth muscle cell phenotypic diversity. Neth. Heart J. 2007, 15, 100–108. [Google Scholar]
- Owens, G.K. Regulation of differentiation of vascular smooth muscle cells. Physiol. Rev. 1995, 75, 487–517. [Google Scholar]
- Chamley-Campbell, J.H.; Campbell, G.R.; Ross, R. Phenotype-dependent response of cultured aortic smooth muscle to serum mitogens. J. Cell Biol. 1981, 89, 379–383. [Google Scholar] [CrossRef]
- Bentzon, J.F.; Weile, C.; Sondergaard, C.S.; Hindkjaer, J.; Kassem, M.; Falk, E. Smooth muscle cells in atherosclerosis originate from the local vessel wall and not circulating progenitor cells in apoe knockout mice. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 2696–2702. [Google Scholar] [CrossRef]
- Costa, M.A.; Simon, D.I. Molecular basis of restenosis and drug-eluting stents. Circulation 2005, 111, 2257–2273. [Google Scholar] [CrossRef]
- Mao, X.; Peng, H.; Ling, J.; Friis, T.; Whittaker, A.K.; Crawford, R.; Xiao, Y. Enhanced human bone marrow stromal cell affinity for modified poly(l-lactide) surfaces by the upregulation of adhesion molecular genes. Biomaterials 2009, 30, 6903–6911. [Google Scholar] [CrossRef] [Green Version]
- Marcovich, R.; Seifman, B.; Beduschi, R.; Wolf, J.S. Surface modification to improve in vitro attachment and proliferation of human urinary tract cells. BJU Int. 2003, 92, 636–640. [Google Scholar] [CrossRef]
- Bisson, I.; Kosinski, M.; Ruault, S.; Gupta, B.; Hilborn, J.; Wurm, F.; Frey, P. Acrylic acid grafting and collagen immobilization on poly(ethylene terephthalate) surfaces for adherence and growth of human bladder smooth muscle cells. Biomaterials 2002, 23, 3149–3158. [Google Scholar] [CrossRef]
- Glukhova, M.; Koteliansky, V.; Fondacci, C.; Marotte, F.; Rappaport, L. Laminin variants and integrin laminin receptors in developing and adult human smooth muscle. Dev. Biol. 1993, 157, 437–447. [Google Scholar] [CrossRef]
- Yim, E.K.; Reano, R.M.; Pang, S.W.; Yee, A.F.; Chen, C.S.; Leong, K.W. Nanopattern-induced changes in morphology and motility of smooth muscle cells. Biomaterials 2005, 26, 5405–5413. [Google Scholar] [CrossRef]
- Nivison-Smith, L.; Weiss, A.S. Alignment of human vascular smooth muscle cells on parallel electrospun synthetic elastin fibers. J. Biomed. Mater. Res. Part A 2012, 100A, 155–161. [Google Scholar] [CrossRef]
- Choudhary, S.; Haberstroh, K.M.; Webster, T.J. Enhanced functions of vascular cells on nanostructured ti for improved stent applications. Tissue Eng. 2007, 13, 1421–1430. [Google Scholar] [CrossRef]
- Choudhary, S.; Berhe, M.; Haberstroh, K.M.; Webster, T.J. Increased endothelial and vascular smooth muscle cell adhesion on nanostructured titanium and cocrmo. Int. J. Nanomed. 2006, 1, 41–49. [Google Scholar]
- Wulf, K.; Teske, M.; Lobler, M.; Luderer, F.; Schmitz, K.P.; Sternberg, K. Surface functionalization of poly(epsilon-caprolactone) improves its biocompatibility as scaffold material for bioartificial vessel prostheses. J. Biomed. Mater. Res. B Appl. Biomater. 2011, 98, 89–100. [Google Scholar]
- Williamson, M.R.; Woollard, K.J.; Griffiths, H.R.; Coombes, A.G. Gravity spun polycaprolactone fibers for applications in vascular tissue engineering: Proliferation and function of human vascular endothelial cells. Tissue Eng. 2006, 12, 45–51. [Google Scholar]
- Sarkar, S.; Lee, G.Y.; Wong, J.Y.; Desai, T.A. Development and characterization of a porous micro-patterned scaffold for vascular tissue engineering applications. Biomaterials 2006, 27, 4775–4782. [Google Scholar] [CrossRef]
- Leszczak, V.; Smith, B.S.; Popat, K.C. Hemocompatibility of polymeric nanostructured surfaces. J. Biomater. Sci. Polym. Ed. 2013, 24, 1529–1548. [Google Scholar] [CrossRef]
- Bechara, S.L.; Judson, A.; Popat, K.C. Template synthesized poly(ε-caprolactone) nanowire surfaces for neural tissue engineering. Biomaterials 2010, 31, 3492–3501. [Google Scholar] [CrossRef]
- Porter, J.R.; Henson, A.; Ryan, S.; Popat, K.C. Biocompatibility and mesenchymal stem cell response to poly(epsilon-caprolactone) nanowire surfaces for orthopedic tissue engineering. Tissue Eng. Part A 2009, 15, 2547–2559. [Google Scholar] [CrossRef]
- Solouk, A.; Mirzadeh, H.; Shokrgozar, M.A.; Solati-Hashjin, M.; Najarian, S.; Seifalian, A.M. The study of collagen immobilization on a novel nanocomposite to enhance cell adhesion and growth. Iran. Biomed. J. 2011, 15, 6–14. [Google Scholar]
- Clyman, R.I.; McDonald, K.A.; Kramer, R.H. Integrin receptors on aortic smooth muscle cells mediate adhesion to fibronectin, laminin, and collagen. Circ. Res. 1990, 67, 175–186. [Google Scholar] [CrossRef]
- He, W.; Ma, Z.; Yong, T.; Teo, W.E.; Ramakrishna, S. Fabrication of collagen-coated biodegradable polymer nanofiber mesh and its potential for endothelial cells growth. Biomaterials 2005, 26, 7606–7615. [Google Scholar] [CrossRef]
- Berridge, M.V.; Herst, P.M.; Tan, A.S. Tetrazolium dyes as tools in cell biology: New insights into their cellular reduction. Biotechnol. Ann. Rev. 2005, 11, 127–152. [Google Scholar] [CrossRef]
- Berridge, M.V.; Tan, A.S. Characterization of the cellular reduction of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (mtt): Subcellular localization, substrate dependence, and involvement of mitochondrial electron transport in mtt reduction. Arch. Biochem. Biophys. 1993, 303, 474–482. [Google Scholar] [CrossRef]
- Lord, M.S.; Foss, M.; Besenbacher, F. Influence of nanoscale surface topography on protein adsorption and cellular response. Nano Today 2010, 5, 66–78. [Google Scholar] [CrossRef]
- Sobue, K.; Hayashi, K.i.; Nishida, W. Expressional regulation of smooth muscle cell-specific genes in association with phenotypic modulation. Mol. Cell. Biochem. 1999, 190, 105–118. [Google Scholar] [CrossRef]
- Steitz, S.A.; Speer, M.Y.; Curinga, G.; Yang, H.Y.; Haynes, P.; Aebersold, R.; Schinke, T.; Karsenty, G.; Giachelli, C.M. Smooth muscle cell phenotypic transition associated with calcification: Upregulation of cbfa1 and downregulation of smooth muscle lineage markers. Circ. Res. 2001, 89, 1147–1154. [Google Scholar] [CrossRef]
- Winder, S.J.; Walsh, M.P. Calponin: Thin filament-linked regulation of smooth muscle contraction. Cell. Signal. 1993, 5, 677–686. [Google Scholar] [CrossRef]
- Miano, J.M.; Cserjesi, P.; Ligon, K.L.; Periasamy, M.; Olson, E.N. Smooth muscle myosin heavy chain exclusively marks the smooth muscle lineage during mouse embryogenesis. Circ. Res. 1994, 75, 803–812. [Google Scholar] [CrossRef]
- Popat, K.C.; Porter, J.R.; Henson, A. Biodegradable poly(epsilon-caprolactone) nanowires for bone tissue engineering applications. Biomaterials 2009, 30, 780–788. [Google Scholar] [CrossRef]
- Andersson, A.-S.; Bäckhed, F.; von Euler, A.; Richter-Dahlfors, A.; Sutherland, D.; Kasemo, B. Nanoscale features influence epithelial cell morphology and cytokine production. Biomaterials 2003, 24, 3427–3436. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Leszczak, V.; Baskett, D.A.; Popat, K.C. Smooth Muscle Cell Functionality on Collagen Immobilized Polycaprolactone Nanowire Surfaces. J. Funct. Biomater. 2014, 5, 58-77. https://doi.org/10.3390/jfb5020058
Leszczak V, Baskett DA, Popat KC. Smooth Muscle Cell Functionality on Collagen Immobilized Polycaprolactone Nanowire Surfaces. Journal of Functional Biomaterials. 2014; 5(2):58-77. https://doi.org/10.3390/jfb5020058
Chicago/Turabian StyleLeszczak, Victoria, Dominique A. Baskett, and Ketul C. Popat. 2014. "Smooth Muscle Cell Functionality on Collagen Immobilized Polycaprolactone Nanowire Surfaces" Journal of Functional Biomaterials 5, no. 2: 58-77. https://doi.org/10.3390/jfb5020058