Time-Dependent Effects of Pre-Aging 3D Polymer Scaffolds in Cell Culture Medium on Cell Proliferation

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Experimental Section
2.1. Fabrication of Macroporous Scaffolds
2.2. Pre-Ageing and Cell Seeding
2.3. Protein Assay
2.4. Measuring Cell Response
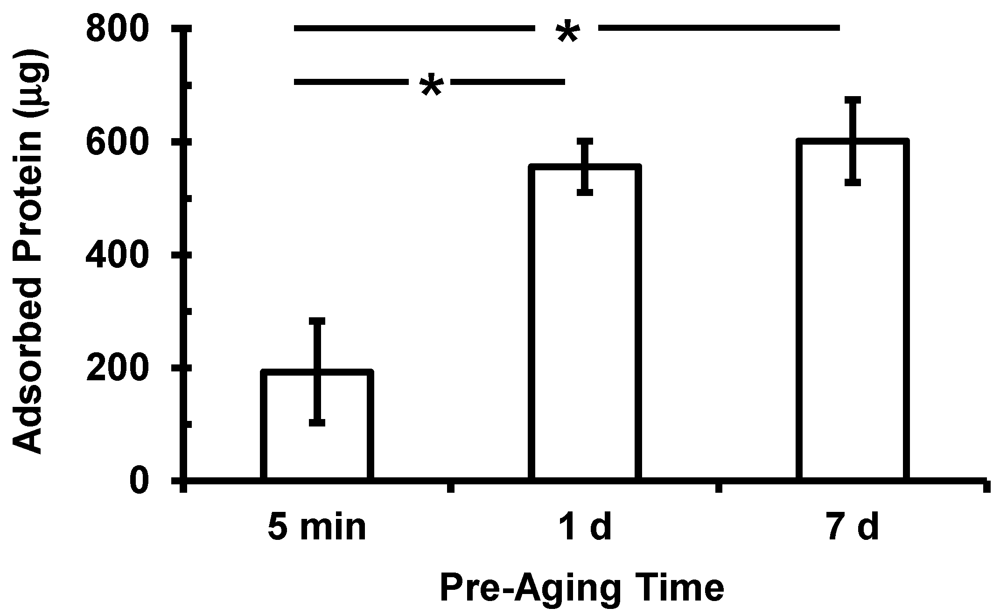
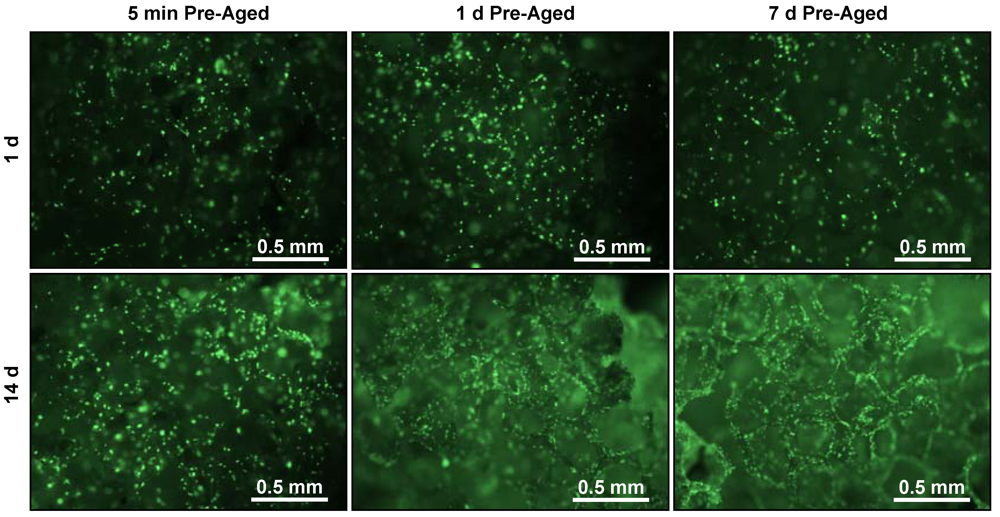
3. Results

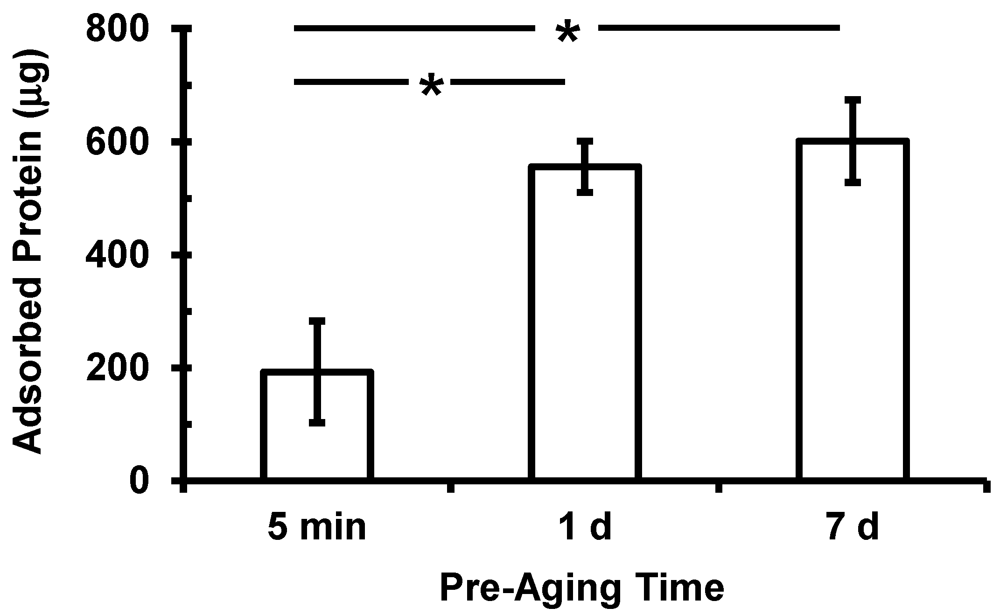

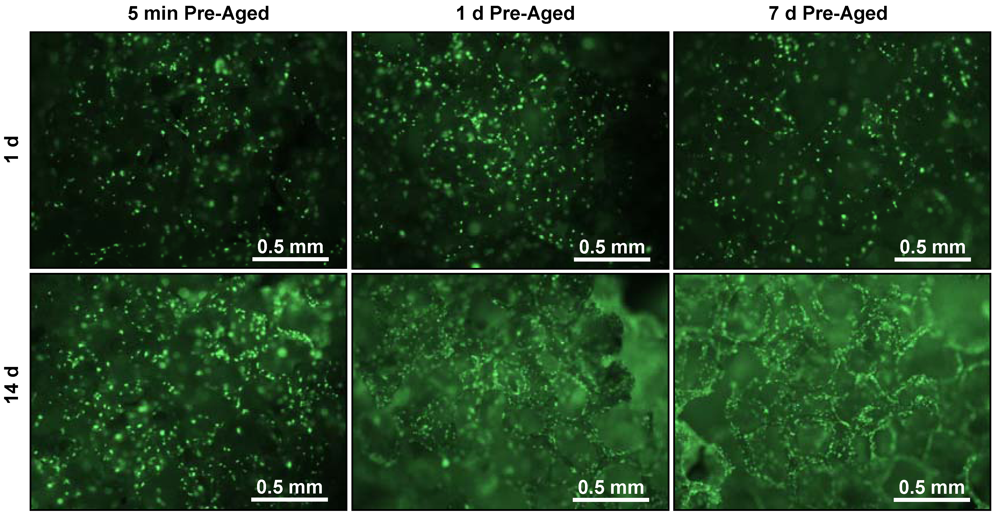
4. Discussion
5. Conclusions
Acknowledgments
References
- Horbett, T.A. The role of adsorbed proteins in tissue response to biomaterials. In Biomaterials Science: An Introduction to Materials in Medicine; Ratner, B.D., Hoffman, A.S., Schoen, F.J., Lemons, J.E., Eds.; Elsevier Academic Press: San Diego, CA, USA, 2004; pp. 237–245. [Google Scholar]
- Soderquist, M.E.; Walton, A.G. Structural changes in proteins adsorbed on polymer surfaces. J. Colloid Interf. Sci. 1980, 75, 386–397. [Google Scholar] [CrossRef]
- Keselowsky, B.G.; Collard, D.M.; García, A.J. Surface chemistry modulates fibronectin conformation and directs integrin binding and specificity to control cell adhesion. J. Biomed. Mater. Res. A 2003, 66A, 247–259. [Google Scholar] [CrossRef]
- Anderson, N.L.; Polanski, M.; Pieper, R.; Gatlin, T.; Tirumalai, R.S.; Conrads, T.P.; Veenstra, T.D.; Adkins, J.N.; Pounds, J.G.; Fagan, R.; Lobley, A. The human plasma proteome. Mol. Cell. Proteomics 2004, 3, 311–326. [Google Scholar] [CrossRef]
- Jung, S.Y.; Lim, S.M.; Albertorio, F.; Kim, G.; Gurau, M.C.; Yang, R.D.; Holden, M.A.; Cremer, P.S. The Vroman effect: A molecular level description of fibrinogen displacement. J. Am. Chem. Soc. 2003, 125, 12782–12786. [Google Scholar]
- Noh, H.; Vogler, E.A. Volumetric interpretation of protein adsorption: Competition from mixtures and the Vroman effect. Biomaterials 2007, 28, 405–422. [Google Scholar] [CrossRef]
- Scotchford, C.A.; Gilmore, C.P.; Cooper, E.; Leggett, G.J.; Downes, S. Protein adsorption and human osteoblast-like cell attachment and growth on alkylthiol on gold self-assembled monolayers. J. Biomed. Mater. Res. 2002, 59, 84–99. [Google Scholar] [CrossRef]
- Arima, Y.; Iwata, H. Effects of surface functional groups on protein adsorption and subsequent cell adhesion using self-assembled monolayers. J. Mater. Chem. 2007, 17, 4079–4087. [Google Scholar] [CrossRef]
- Yildirim, E.D.; Besunder, R.; Pappas, D.; Allen, F.; Guceri, S.; Sun, W. Accelerated differentiation of osteoblast cells on polycaprolactone scaffolds driven by a combined effect of protein coating and plasma modification. Biofabrication 2010, 2, 014109–1. [Google Scholar]
- Woo, K.M.; Seo, J.; Zhang, R.; Ma, P.X. Suppression of apoptosis by enhanced protein adsorption on polymer/hydroxyapatite composite scaffolds. Biomaterials 2007, 28, 2622–2630. [Google Scholar] [CrossRef]
- Chen, R.; Gallant, N.D.; Smith, J.; Kipper, M.; Simon, C.G. Time-dependent effects of pre-aging polymer films in cell culture medium on cell adhesion and spreading. J. Mater. Sci. Mater. Med. 2008, 19, 1759–1766. [Google Scholar] [CrossRef]
- Abbott, A. Cell culture: Biology’s new dimension. Nature 2003, 424, 870–872. [Google Scholar] [CrossRef]
- Yang, Y.; Bolikal, D.; Becker, M.L.; Kohn, J.; Zeiger, D.N.; Simon, C.G. Combinatorial polymer scaffold libraries for screening cell-biomaterial interactions in 3D. Adv. Mater. 2008, 20, 2037–2043. [Google Scholar]
- Mikos, A.G.; Lu, L.; Temenoff, J.S.; Tessmar, J.K. Synthetic bioresorable polymer scaffolds. In Biomaterials Science: An Introduction to Materials in Medicine; Ratner, B.D., Hoffman, A.S., Schoen, F.J., Lemons, J.E., Eds.; Elsevier Academic Press: San Diego, CA, USA, 2004; pp. 237–245. [Google Scholar]
- Sachlos, E.; Czernuszka, J.T. Making tissue engineering scaffolds work. Review: The application of solid freeform fabrication technology to the production of tissue engineering scaffolds. Eur. Cells Mater. 2003, 5, 29–39. [Google Scholar]
- Ahmad, Z.; Rasekh, M.; Edirisinghe, M. Electrohydrodynamic direct rwiting of biomedical polymers and composites. Macromol. Mater. Eng. 2010, 295, 315–319. [Google Scholar] [CrossRef]
- Rasekh, M.; Ahmad, Z.; Day, R.; Edirisinghe, M. Direct writing of polycaprolactone polymer for potential biomedical engineering applications. Adv. Eng. Mater. 2011, 13, B296–B305. [Google Scholar]
- Shea, L.D.; Wang, D.; Franceschi, R.T.; Mooney, D.J. Engineered bone development from a pre-osteoblast cell line on three-dimensional scaffolds. Tissue Eng. 2000, 6, 605–617. [Google Scholar] [CrossRef]
- Kumar, G.; Tison, C.K.; Chatterjee, K.; Pine, P.S.; McDaniel, J.H.; Salit, M.L.; Young, M.F.; Simon, C.G. The determination of stem cell fate by 3D scaffold structures through the control of cell shape. Biomaterials 2011, 32, 9188–9196. [Google Scholar]
- Sudo, H.; Kodama, H.A.; Amagai, Y.; Yamamoto, S.; Kasai, S. In vitro differentiation and calcification in a new clonal osteogenic cell line derived from newborn mouse calvaria. J. Cell Biol. 1983, 96, 191–198. [Google Scholar] [CrossRef]
- Powell, K. Dishing up bone formation. J. Cell Biol. 2005, 171, 409. [Google Scholar] [CrossRef]
- Chatterjee, K.; Lin-Gibson, S.; Wallace, W.E.; Parekh, S.H.; Lee, Y.J.; Cicerone, M.T.; Young, M.F.; Simon, C.G. The effect of 3D hydrogel scaffold modulus on osteoblast differentiation and mineralization revealed by combinatorial screening. Biomaterials 2010, 31, 5051–5062. [Google Scholar]
- Chatterjee, K.; Sun, L; Chow, L.C.; Young, M.F.; Simon, C.G. Combinatorial screening of osteoblast response to 3D calcium phosphate/poly(ε-caprolactone) scaffolds using gradients and arrays. Biomaterials 2011, 32, 1361–1369. [Google Scholar] [CrossRef]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar]
- Chadwick, R.B.; Conrad, M.P.; McGinnis, M.D.; Johnston-Dow, L.; Spurgeon, S.L.; Kronick, M.N. Heterozygote and mutation detection by direct automated fluorescent DNA sequencing using a mutant Taq DNA polymerase. Biotechniques 1996, 20, 676–83. [Google Scholar]
- Simon, C.G.; Yang, Y.; Dorsey, S.M.; Ramalingam, M.; Chatterjee, K. 3D polymer scaffold arrays. Methods Mol. Biol. 2011, 671, 161–174. [Google Scholar] [CrossRef]
- Hayman, E.G.; Ruoslahti, E. Distribution of fetal bovine serum fibronectin and endogenous rat-cell fibronectin in extracellular matrix. J. Cell Biol. 1979, 83, 255–259. [Google Scholar] [CrossRef]
- Pierschbacher, M.D.; Ruoslahti, E. Cell attachment activity of fibronectin can be duplicated by small synthetic fragments of the molecule. Nature 1984, 309, 30–33. [Google Scholar] [CrossRef]
- Petrie, T.A.; Raynor, J.E.; Dumbauld, D.W.; Lee, T.T.; Jagtap, S.; Templeman, K.L.; Collard, D.M.; Garcia, A.J. Multivalent integrin-specific ligands enhance tissue healing and biomaterial integration. Sci. Transl. Med. 2010, 2, 45–60. [Google Scholar]
- Savarino, L.; Baldini, N.; Greco, M.; Capitani, O.; Pinna, S.; Valentini, S.; Lombardo, B.; Esposito, M.T.; Pastore, L.; Ambrosio, L.; et al. The performance of poly-ε-caprolactone scaffolds in a rabbit femur model with and without autologous stromal cells and BMP4. Biomaterials 2007, 28, 3101–3109. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chatterjee, K.; Hung, S.; Kumar, G.; Simon, C.G., Jr. Time-Dependent Effects of Pre-Aging 3D Polymer Scaffolds in Cell Culture Medium on Cell Proliferation. J. Funct. Biomater. 2012, 3, 372-381. https://doi.org/10.3390/jfb3020372
Chatterjee K, Hung S, Kumar G, Simon CG Jr. Time-Dependent Effects of Pre-Aging 3D Polymer Scaffolds in Cell Culture Medium on Cell Proliferation. Journal of Functional Biomaterials. 2012; 3(2):372-381. https://doi.org/10.3390/jfb3020372
Chicago/Turabian StyleChatterjee, Kaushik, Stevephen Hung, Girish Kumar, and Carl G. Simon, Jr. 2012. "Time-Dependent Effects of Pre-Aging 3D Polymer Scaffolds in Cell Culture Medium on Cell Proliferation" Journal of Functional Biomaterials 3, no. 2: 372-381. https://doi.org/10.3390/jfb3020372