How Management Factors Influence Weed Communities of Cereals, Their Diversity and Endangered Weed Species in Central Europe

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Areas and Field Selection
2.2. Vegetation Recordings and Farmer Surveys
2.3. Data Analysis
3. Results
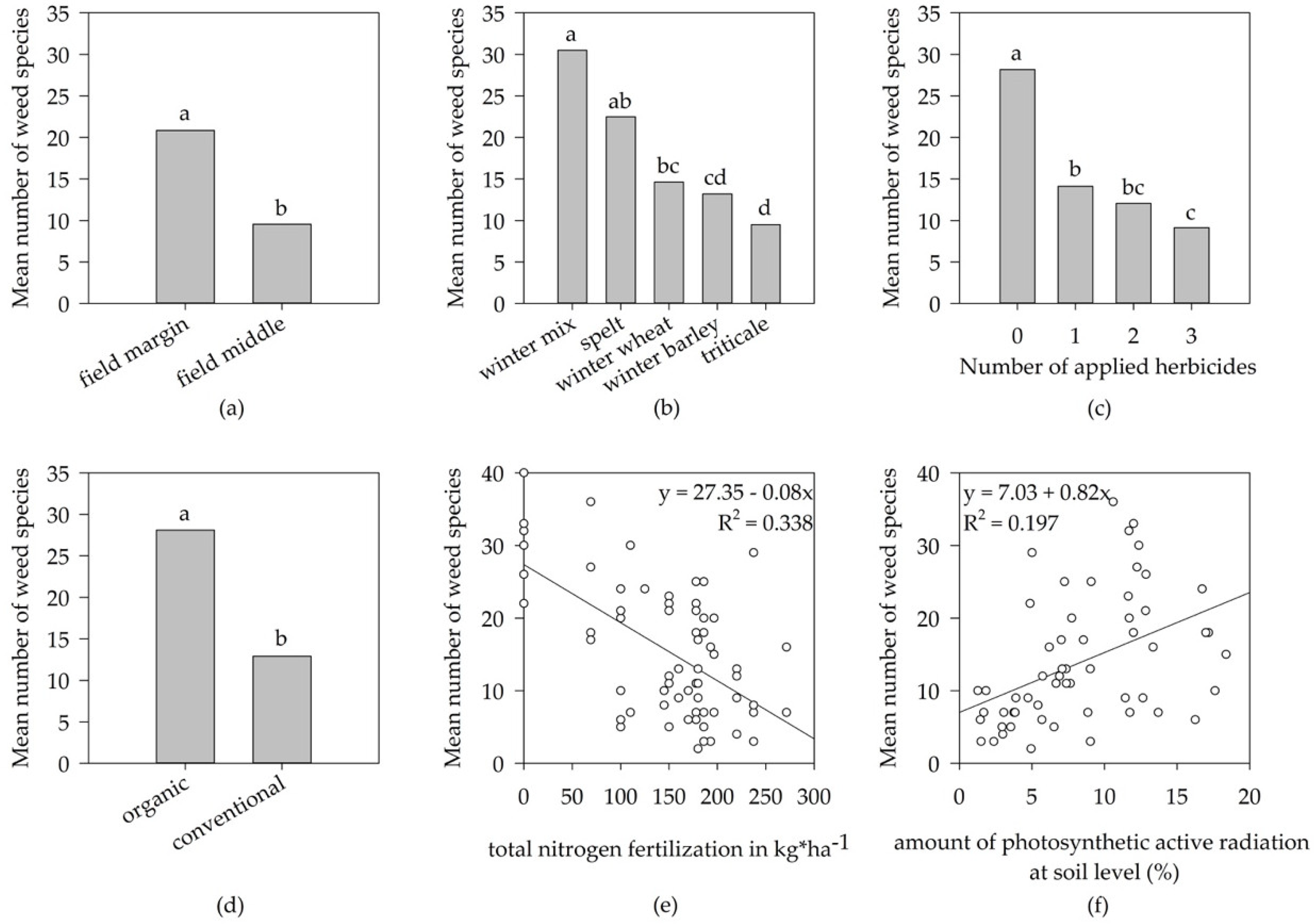
3.1. Weed Species Diversity
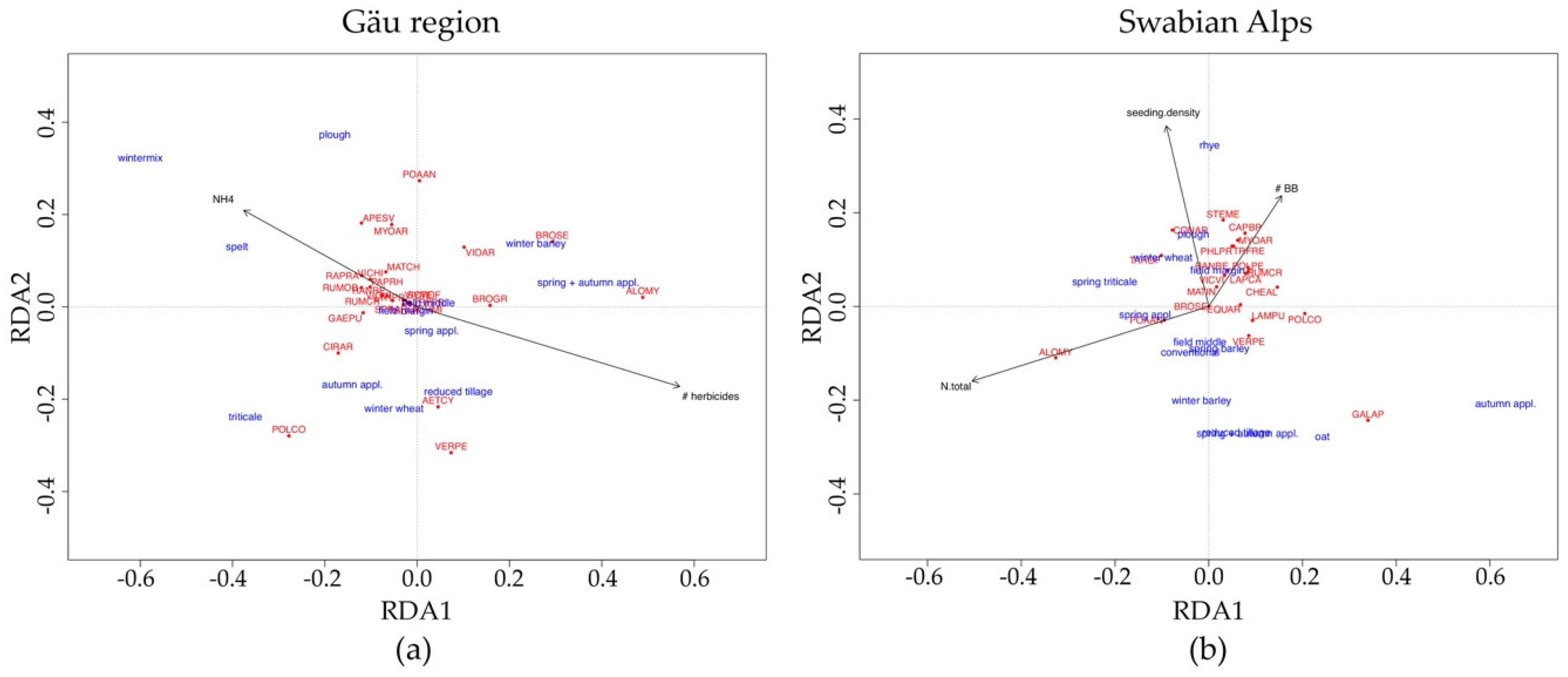
3.2. Weed Species Community
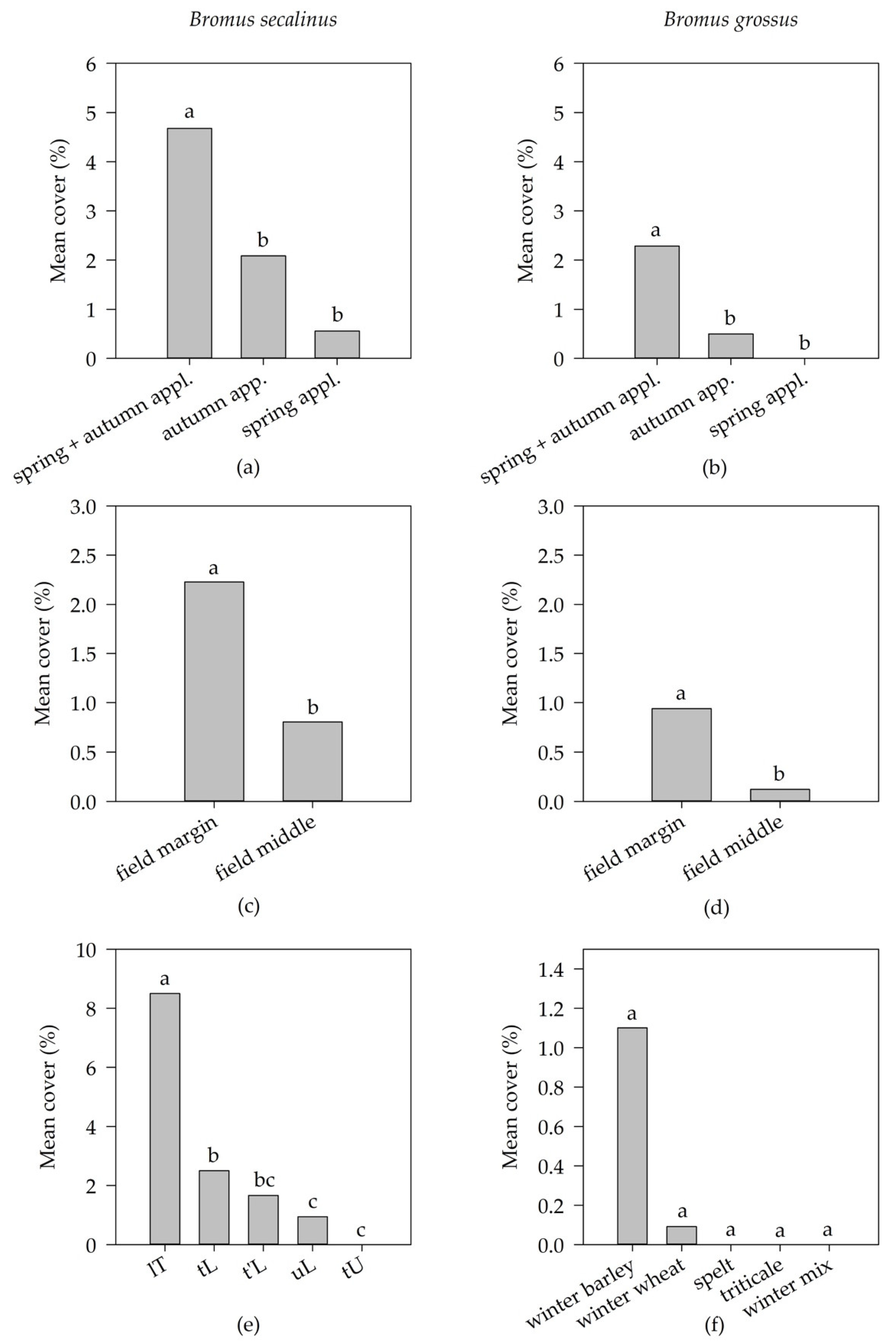
3.3. Endangered Weed Species (EWS)
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Convention on Biological Diversity—Safeguarding Life on Earth. Available online: https://www.cbd.int/information/parties.shtml (accessed on 14 October 2018).
- Power, M.E. Top-Down and Bottom-Up Forces in Food Webs: Do Plants Have Primacy. Ecology 1992, 73, 733–746. [Google Scholar] [CrossRef]
- Scherber, C.; Eisenhauer, N.; Weisser, W.W.; Schmid, B.; Voigt, W.; Fischer, M.; Schulze, E.-D.; Roscher, C.; Weigelt, A.; Allan, E.; et al. Bottom-up effects of plant diversity on multitrophic interactions in a biodiversity experiment. Nature 2010, 468, 553–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holt, A.R.; Alix, A.; Thompson, A.; Maltby, L. Food production, ecosystem services and biodiversity: We can’t have it all everywhere. Sci. Total Environ. 2016, 573, 1422–1429. [Google Scholar] [CrossRef] [PubMed]
- Flynn, D.F.B.; Gogol-Prokurat, M.; Nogeire, T.; Molinari, N.; Richers, B.T.; Lin, B.B.; Simpson, N.; Mayfield, M.M.; DeClerck, F. Loss of functional diversity under land use intensification across multiple taxa. Ecol. Lett. 2009, 12, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Meyer, S.; Wesche, K.; Krause, B.; Leuschner, C. Dramatic losses of specialist arable plants in Central Germany since the 1950s/60s—A cross-regional analysis. Divers. Distrib. 2013, 19, 1175–1187. [Google Scholar] [CrossRef]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef] [PubMed]
- Donald, P.F.; Green, R.E.; Heath, M.F. Agricultural intensification and the collapse of Europe’s farmland bird populations. Proc. R. Soc. B Biol. Sci. 2001, 268, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Convention on Biological Diversity—Safeguarding Life on Earth. Available online: https://www.cbd.int/agro/whatstheproblem.shtml (accessed on 14 October 2018).
- Oerke, E.-C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Blaix, C.; Moonen, A.C.; Dostatny, D.F.; Izquierdo, J.; Le Corff, J.; Morrison, J.; Von Redwitz, C.; Schumacher, M.; Westerman, P.R. Quantification of regulating ecosystem services provided by weeds in annual cropping systems using a systematic map approach. Weed Res. 2018, 58, 151–164. [Google Scholar] [CrossRef] [Green Version]
- Marshall, E.J.P.; Brown, V.K.; Boatman, N.D.; Lutman, P.J.W.; Squire, G.R.; Ward, L.K. The role of weeds in supporting biological diversity within crop fields. Weed Res. 2003, 43, 77–89. [Google Scholar] [CrossRef]
- Gerhards, R.; Dieterich, M.; Schumacher, M. Rückgang von Ackerunkräutern in Baden-Württemberg—Ein Vergleich von vegetationskundlichen Erhebungen in den Jahren 1948/49, 1975–1978 und 2011 im Raum Mehrstetten—Empfehlungen für Landwirtschaft und Naturschutz. Gesunde Pflanzen 2013, 65, 151–160. [Google Scholar] [CrossRef]
- Atakan, E. Influence of weedy field margins on abundance patterns of the predatory bugs Orius spp. and their prey, the western flower thrips (Frankliniella occidentalis), on faba bean. Phytoparasitica 2010, 38, 313–325. [Google Scholar] [CrossRef]
- Brütting, C.; Wesche, K.; Meyer, S.; Hensen, I. Genetic diversity of six arable plants in relation to their Red List status. Biodivers. Conserv. 2012, 21, 745–761. [Google Scholar] [CrossRef]
- Gerowitt, B.; Bertke, E.; Hespelt, S.-K.; Tute, C. Towards multifunctional agriculture–weeds as ecological goods? Weed Res. 2003, 43, 227–235. [Google Scholar] [CrossRef]
- Meyer, S.; Van Elsen, T.; Blümlein, B.; Kaerlein, M.; Metzger, J.; Gottwald, F.; Wehke, S.; Dieterich, M.; Wahmhoff, W.; Stock, R.; et al. Conserving agrobiodiversity through arable field sanctuaries. Natur und Landschaft 2014, 9, 434–441. [Google Scholar]
- Firbank, L.G. Agrostemma githago L. (Lychnis githago (L.) Scop.). J. Ecol. 1988, 76, 1232–1246. [Google Scholar] [CrossRef]
- Van Elsen, T.; Hotze, C.; Meyer, S.; Gottwald, F.; Wehke, S. Empfehlungen für die Bewirtschaftung von Schutzäckern; University of Kassel: Kassel, Germany, 2009. [Google Scholar]
- Meyer, S.; Van Elsen, T.; Gottwald, F.; Hotze, C.; Wehke, S. Monitoring-Konzept für die Entwicklung der Vegetation von Schutzäckern; University of Kassel: Kassel, Germany, 2010. [Google Scholar]
- Waldhardt, R.; Simmering, D.; Albrecht, H. Floristic diversity at the habitat scale in agricultural landscapes of Central Europe—Summary, conclusions and perspectives. Agric. Ecosyst. Environ. 2003, 98, 79–85. [Google Scholar] [CrossRef]
- Fried, G.; Petit, S.; Dessaint, F.; Reboud, X. Arable weed decline in Northern France: Crop edges as refugia for weed conservation? Biol. Conserv. 2009, 142, 238–243. [Google Scholar] [CrossRef]
- Heap, I. Global perspective of herbicide-resistant weeds. Pest Manag. Sci. 2014, 70, 1306–1315. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, H. Suitability of arable weeds as indicator organisms to evaluate species conservation effects of management in agricultural ecosystems. Agric. Ecosyst. Environ. 2003, 98, 201–211. [Google Scholar] [CrossRef]
- Schumacher, M.; Hahn, A.-K.; Gerhards, R. The Influence of Farming on Weed Flora in the Gäu Region of Southwestern Germany with an Emphasis on Rare Arable Weed Species; Julius-Kühn-Archiv: Braunschweig, Germany, 2018; Volume 458, pp. 30–34. [Google Scholar]
- Torra, J.; Recasens, J.; Royo-Esnal, A. Seedling emergence response of rare arable plants to soil tillage varies by species. PLoS ONE 2018, 13, e0199425. [Google Scholar] [CrossRef] [PubMed]
- Van Elsen, T. Ackerwildkraut-Gesellschaften herbizidfreier Ackerränder und des herbzidbehandelten Bestandesinneren im Vergleich. Tuexenia 1989, 9, 75–105. [Google Scholar]
- Wilmanns, O. Ökologische Pflanzensoziologie: Eine Einführung in die Vegetation Mitteleuropas; 6. Auflage; Quelle & Meyer: Wiesbaden, Germany, 1998; ISBN 978-3-8252-0269-9. [Google Scholar]
- Van der Maarel, E. Transformation of cover-abundance values for appropriate numerical treatment – Alternatives to the proposals by Podani. J. Veg. Sci. 2007, 18, 767–770. [Google Scholar] [CrossRef]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, J.; Monette, G. Generalized collinearity diagnostics. J. Am. Stat. Assoc. 1992, 87, 178–183. [Google Scholar] [CrossRef]
- Lososová, Z.; Chytrý, M.; Cimalová, Š.; Kropáč, Z.; Otýpková, Z.; Pyšek, P.; Tichý, L. Weed vegetation of arable land in Central Europe: Gradients of diversity and species composition. J. Veg. Sci. 2004, 15, 415–422. [Google Scholar] [CrossRef]
- Koch, M.A.; Meyer, N.; Engelhardt, M.; Thiv, M.; Bernhardt, K.-G.; Michling, F. Morphological and genetic variation of highly endangered Bromus species and the status of these Neolithic weeds in Central Europe. Plant Syst. Evol. 2016, 302, 515–525. [Google Scholar] [CrossRef]
- Van Elsen, T. Species diversity as a task for organic agriculture in Europe. Agric. Ecosyst. Environ. 2000, 77, 101–109. [Google Scholar] [CrossRef]
- Piqueray, J.; Gilliaux, V.; Gaillard, T.; Mahy, G.; Delescaille, L.-M. Uncleaned crop seed sowing as a tool to conserve Bromus grossus and restore species-rich arable-dependent plant communities. Conserv. Evid. 2018, 15, 26–31. [Google Scholar]
- Peigné, J.; Ball, B.C.; Roger-Estrade, J.; David, C. Is conservation tillage suitable for organic farming? A review. Soil Use Manag. 2007, 23, 129–144. [Google Scholar] [CrossRef]
- Lang, M.; Prestele, J.; Wiesinger, K.; Kollmann, J.; Albrecht, H. Reintroduction of rare arable plants: Seed production, soil seed banks, and dispersal 3 years after sowing: Reintroduction of rare arable plants. Restor. Ecol. 2018, 26, 170–178. [Google Scholar] [CrossRef]
- Keller, M.; Böhringer, N.; Möhring, J.; Rueda-Ayala, V.; Gutjahr, C.; Gerhards, R. Long-term changes in weed occurrence, yield and use of herbicides in maize in south-western Germany, with implications for the determination of economic thresholds. Weed Res. 2014, 54, 457–466. [Google Scholar] [CrossRef]
- Dessaint, F.; Chadoeuf, R. Barralis Diversity of weed communities of annual crops in côte-d’Or, France. Biotechnol. Agron. Soc. Environ. 2001, 5, 91–98. [Google Scholar]
- Rühl, A.T.; Eckstein, R.L.; Otte, A.; Donath, T.W. Future challenge for endangered arable weed species facing global warming: Low temperature optima and narrow moisture requirements. Biol. Conserv. 2015, 182, 262–269. [Google Scholar] [CrossRef]
- McDonald, A.; Riha, S.; DiTommaso, A.; DeGaetano, A. Climate change and the geography of weed damage: Analysis of U.S. maize systems suggests the potential for significant range transformations. Agric. Ecosyst. Environ. 2009, 130, 131–140. [Google Scholar] [CrossRef]
- Nowak, A.; Nowak, S.; Nobis, M.; Nobis, A. Crop type and altitude are the main drivers of species composition of arable weed vegetation in Tajikistan. Weed Res. 2015, 55, 525–536. [Google Scholar] [CrossRef]
- Nagy, K.; Lengyel, A.; Kovács, A.; Türei, D.; Csergő, A.M.; Pinke, G. Weed species composition of small-scale farmlands bears a strong crop-related and environmental signature. Weed Res. 2018, 58, 46–56. [Google Scholar] [CrossRef]
- Derksen, D.A. Impact of post-emergence herbicides on weed community diversity within conservation-tillage systems. Weed Res. 1995, 35, 311–320. [Google Scholar] [CrossRef]
- Chadwigk, M.J. The response to variations in nitrogen level of some weed species of the Sudan. Weed Res. 1963, 3, 230–241. [Google Scholar] [CrossRef]
- De Mol, F.; von Redwitz, C.; Gerowitt, B. Weed species composition of maize fields in Germany is influenced by site and crop sequence. Weed Res. 2015, 55, 574–585. [Google Scholar] [CrossRef]
- Pinke, G.; Blazsek, K.; Magyar, L.; Nagy, K.; Karácsony, P.; Czúcz, B.; Botta-Dukát, Z. Weed species composition of conventional soyabean crops in Hungary is determined by environmental, cultural, weed management and site variables. Weed Res. 2016, 56, 470–481. [Google Scholar] [CrossRef]
- Zarina, L.; Gerowitt, B.; Melander, B.; Salonen, J.; Krawczuk, R.; Verwijst, T. Crop diversification for weed management in organic arable cropping systems. Environ. Technol. Resour. Proc. Int. Sci. Pract. Conf. 2015, 2, 333. [Google Scholar] [CrossRef] [Green Version]
- Kleijn, D.; van der Voort, L.A.C. Conservation headlands for rare arable weeds: The effects of fertilizer application and light penetration on plant growth. Biol. Conserv. 1997, 81, 57–67. [Google Scholar] [CrossRef]
- Schumacher, W. Schutz und Erhaltung gefährdeter Ackerwildkräuter durch Integration von landwirtschaftlicher Nutzung und Naturschutz. Natur und Landschaft 1980, 55, 447–453. [Google Scholar]
- Marshall, E.J.P.; Moonen, A.C. Field margins in northern Europe: Their functions and interactions with agriculture. Agric. Ecosyst. Environ. 2002, 89, 5–21. [Google Scholar] [CrossRef]
- Kunz, C.; Weber, J.; Gerhards, R. Benefits of Precision Farming Technologies for Mechanical Weed Control in Soybean and Sugar Beet—Comparison of Precision Hoeing with Conventional Mechanical Weed Control. Agronomy 2015, 5, 130–142. [Google Scholar] [CrossRef] [Green Version]
- Kristensen, L.; Olsen, J.; Weiner, J. Crop Density, Sowing Pattern, and Nitrogen Fertilization Effects on Weed Suppression and Yield in Spring Wheat. Weed Sci. 2008, 56, 97–102. [Google Scholar] [CrossRef]
- Blackmer, A.M.; White, S.E. Using precision farming technologies to improve management of soil and fertilizer nitrogen. Aust. J. Agric. Res. 1998, 49, 555–564. [Google Scholar] [CrossRef]
- Storkey, J.; Moss, S.R.; Cussans, J.W. Using Assembly Theory to Explain Changes in a Weed Flora in Response to Agricultural Intensification. Weed Sci. 2010, 58, 39–46. [Google Scholar] [CrossRef]
- Denys, C.; Tscharntke, T. Plant-insect communities and predator-prey ratios in field margin strips, adjacent crop fields, and fallows. Oecologia 2002, 130, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Meek, B.; Loxton, D.; Sparks, T.; Pywell, R.; Pickett, H.; Nowakowski, M. The effect of arable field margin composition on invertebrate biodiversity. Biol. Conserv. 2002, 106, 259–271. [Google Scholar] [CrossRef]
- Krompa, B.; Steinberger, K.-H. Grassy field margins and arthropod diversity: A case study on ground beetles and spiders in eastern Austria (Coleoptera: Carabidae; Arachnida: Aranei, Opiliones). Biot. Divers. Agroecosyst. 2012, 71–93. [Google Scholar]
- Meyer, S.; Leuschner, C.; Van Elsen, T. Sanctuaries for the segetal vegetation in Germany—Inventory and new impulses by the project “Biodiversity in agricultural landscapes”. J. Plant Diseases Protect. 2008, 21, 363–368. [Google Scholar]
- Landis, D.A. Designing agricultural landscapes for biodiversity-based ecosystem services. Basic Appl. Ecol. 2017, 18, 1–12. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Gäu Region | Swabian Alps |
|---|---|---|
| Levels | Levels | |
| Location in the field | Field margin; field middle | Field margin; field middle |
| Farming system | Organic; conventional | Organic; conventional |
| Crop species | winter mix 1; spelt; winter barley; winter wheat; triticale | Oat; rye; spring barley; triticale; winter barley; winter wheat |
| Photosynthetic active radiation (PAR) at soil level (%) | 1.3–18.4 | 2.8–70.0 |
| Crop cover (%) | 55.0–95.0 | 30.0–97.0 |
| Nitrate in the soil in June (NO3) (kg ha−1) | 1.7–115.9 | - |
| NH4 in the soil in June (kg ha−1) | 0.0–17.5 | - |
| Soil type | lT, tL, t’L, uL, tU 2 | - |
| Timing of herbicide application | Spring appl.; autumn appl.; spring + autumn appl. 3 | Spring appl.; autumn appl.; spring + autumn appl. |
| Total nitrogen fertilization (kg ha−1) | 0.0–271.5 | 44.0–110.0 |
| Crop row distance (cm) | 12.0–17.0 | 12.2–15.0 |
| Seeding density (kg ha−1) | 140.0–260.0 | 100.0–250.0 |
| Number of applied herbicides | 0–3 | - |
| Number of tillage operations | 2–4 | 2–4 |
| Tillage | Plough; reduced tillage | Plough; reduced tillage |
| Crop height (cm) | 51–153 | 45–150 |
| Field size (ha) | - | 0.15–5.00 |
| Nitrogen fertilization | - | Mineral; organic; mineral + organic |
| Variables | Gäu Region | Swabian Alps | ||||
|---|---|---|---|---|---|---|
| Gross Effect | Net Effect | F-Value | Gross Effect | Net Effect | F-Value | |
| Full model 1 | 42.54 | 3.634 *** | 46.63 | 2.185 *** | ||
| Crop species | 21.92 | 12.61 | 2.963 *** | 19.05 | 20.83 | 2.276 *** |
| Tillage | 7.79 | 5.37 | 5.049 *** | 3.39 | 3.31 | 2.174 ** |
| Number of herbicide applications | 7.13 | 2.60 | 2.441 ** | - | - | - |
| Location in the field | 2.77 | 2.63 | 2.471 *** | 3.20 | 3.20 | 2.101 ** |
| Herbicide timing | 16.51 | 5.93 | 1.857 *** | 11.44 | 7.51 | 2.463 *** |
| NH4 | 4.56 | 2.37 | 2.235 ** | |||
| Total N | 4.21 | 4.53 | 2.968 *** | |||
| Farming system | 5.59 | NA | NA | |||
| Number of tillage operations | 3.11 | 2.76 | 1.812 ** | |||
| Seeding density | 2.87 | 2.53 | 1.659 * | |||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schumacher, M.; Ohnmacht, S.; Rosenstein, R.; Gerhards, R. How Management Factors Influence Weed Communities of Cereals, Their Diversity and Endangered Weed Species in Central Europe. Agriculture 2018, 8, 172. https://doi.org/10.3390/agriculture8110172
Schumacher M, Ohnmacht S, Rosenstein R, Gerhards R. How Management Factors Influence Weed Communities of Cereals, Their Diversity and Endangered Weed Species in Central Europe. Agriculture. 2018; 8(11):172. https://doi.org/10.3390/agriculture8110172
Chicago/Turabian StyleSchumacher, Matthias, Simon Ohnmacht, Ronja Rosenstein, and Roland Gerhards. 2018. "How Management Factors Influence Weed Communities of Cereals, Their Diversity and Endangered Weed Species in Central Europe" Agriculture 8, no. 11: 172. https://doi.org/10.3390/agriculture8110172