
Effect of Ozone Treatment on Inactivation of Escherichia coli and Listeria sp. on Spinach

Abstract
:1. Introduction
2. Results and Discussion
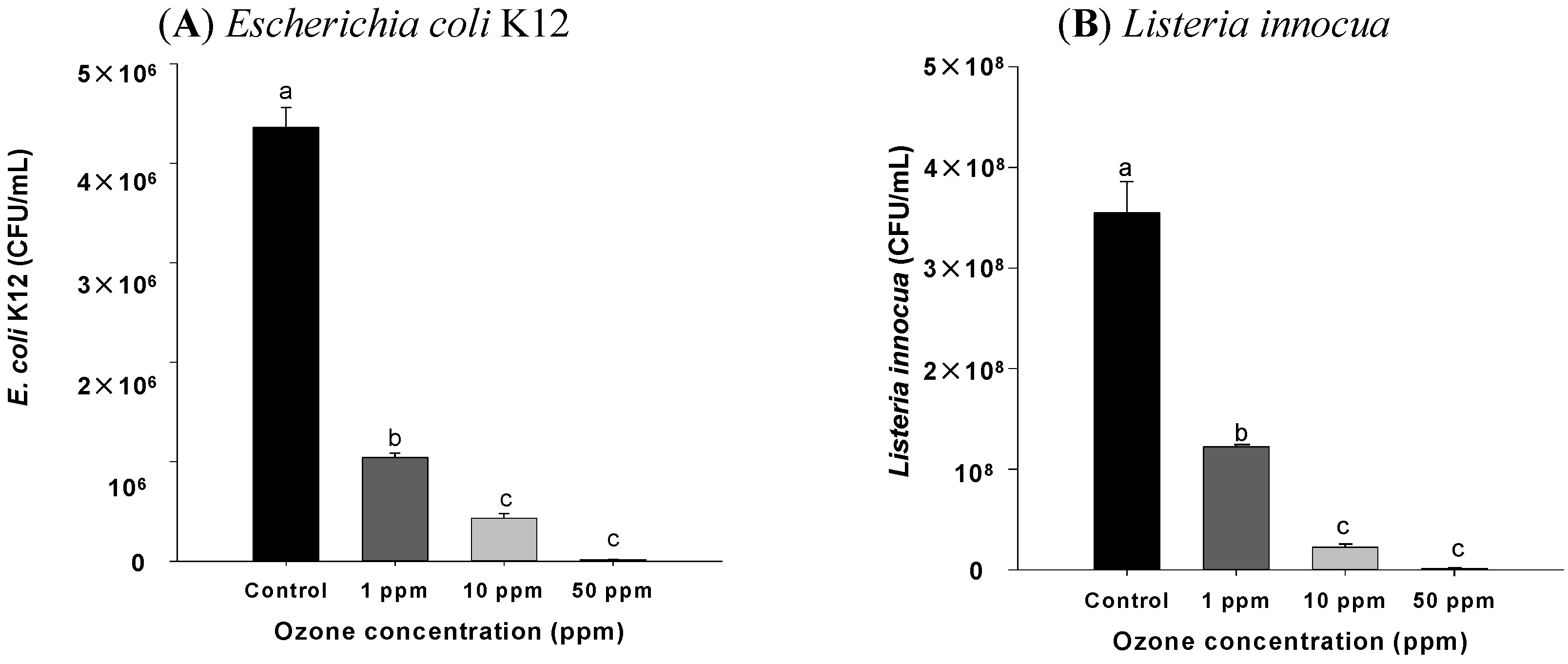
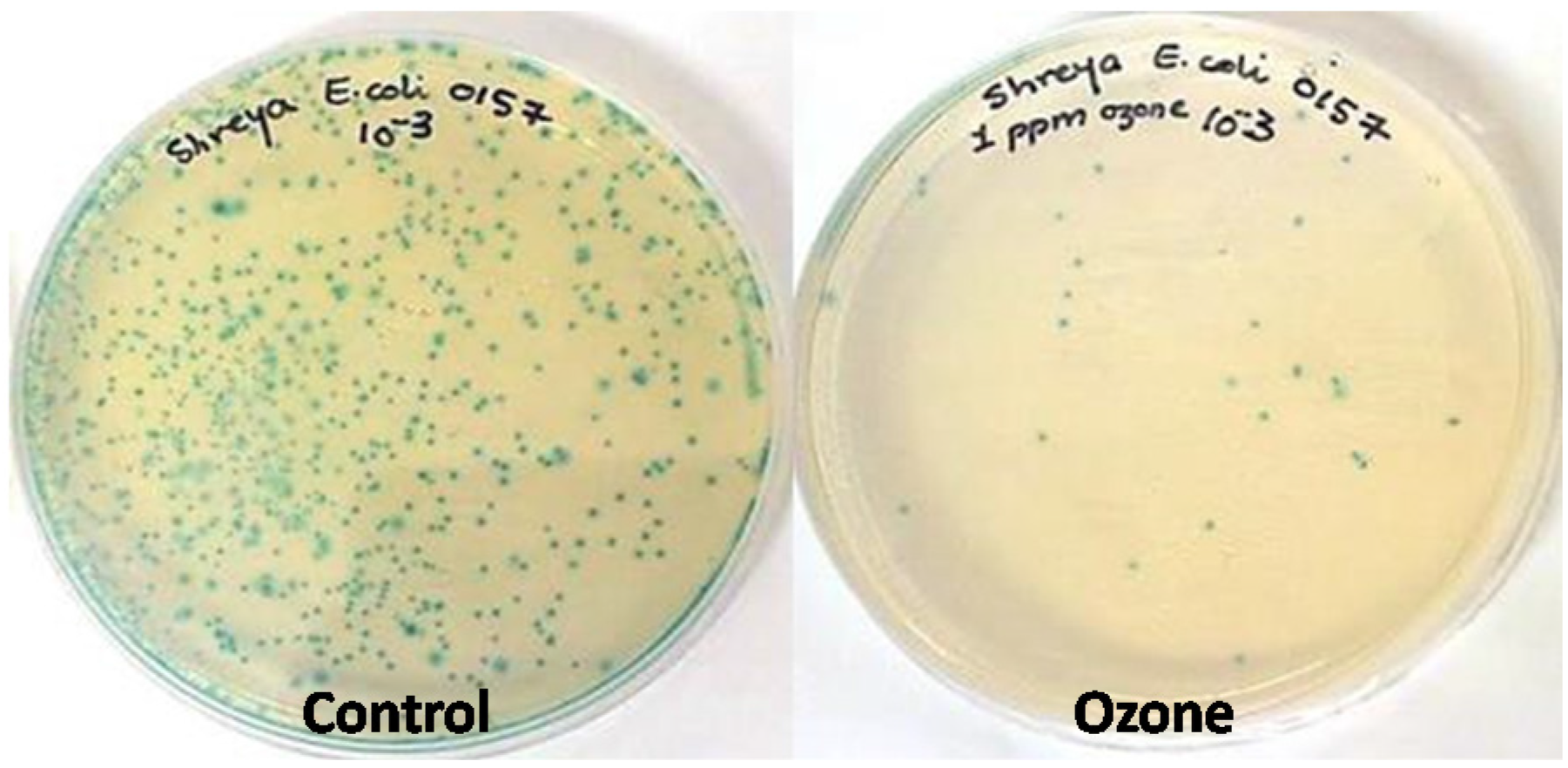
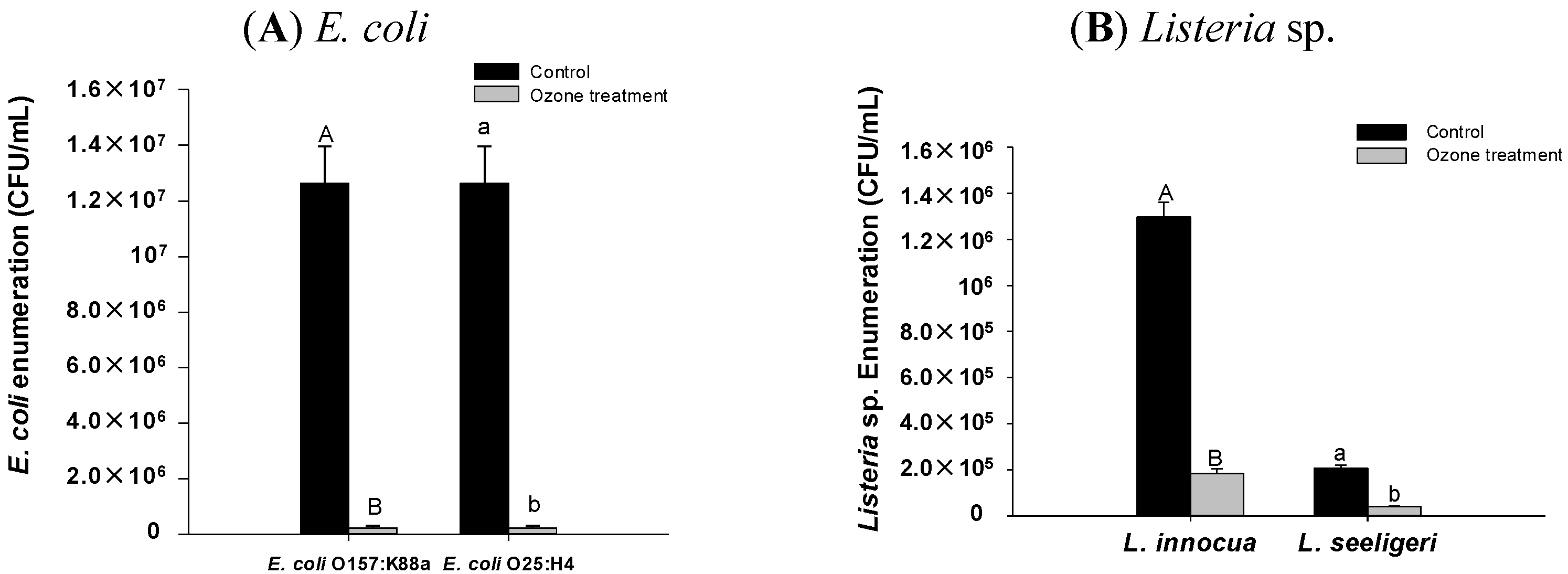
2.1. Effect of Ozone Exposure on E. coli and Listeria sp. in Vitro
2.2. Optimization of the Concentration and Duration of Ozone Exposure Levels to Treat Spinach without Causing Visual Damage

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Duration of the Exposure of Spinach | ||||
|---|---|---|---|---|
| Ozone concentration | 10 ppm | 15 ppm | 20 ppm | 25 ppm |
| Time | 2 min | 45 s | 30 s | Damaged at 30 s, 45 s and 2 min |

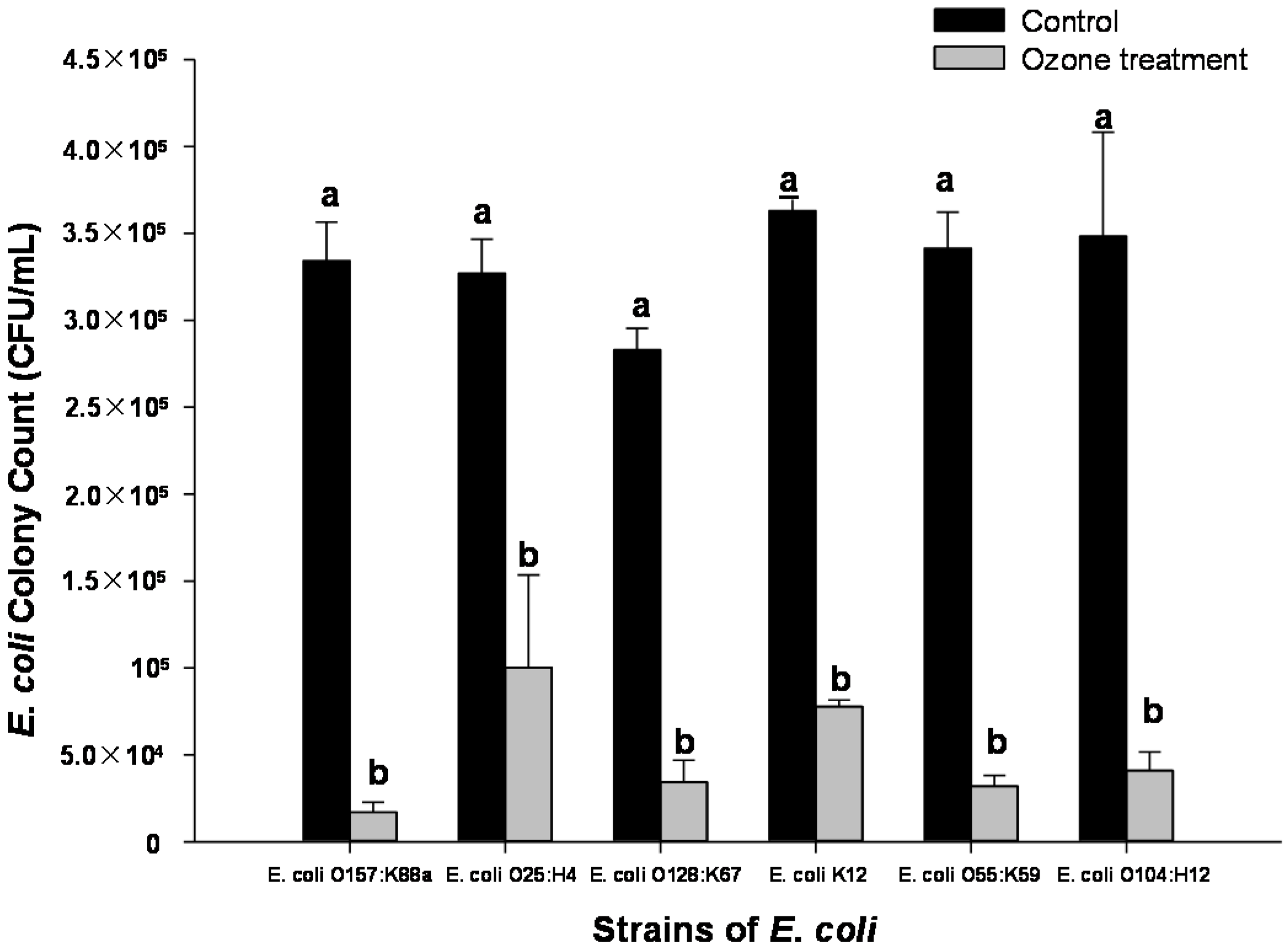
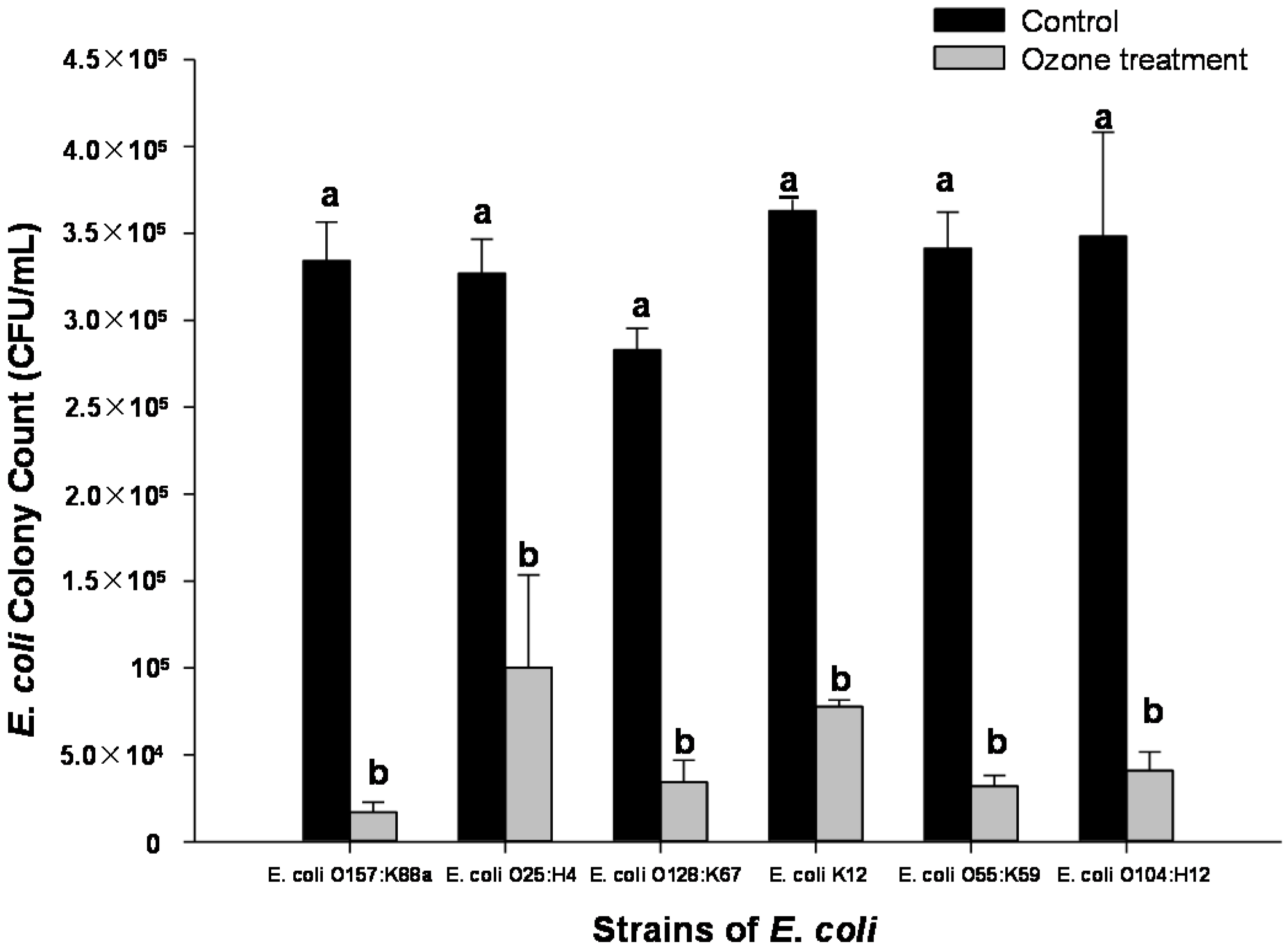
2.3. Effect of Ozone Exposure (1 ppm for 10 min) on Different Strains of E. coli Inoculated onto Spinach Leaf Surfaces
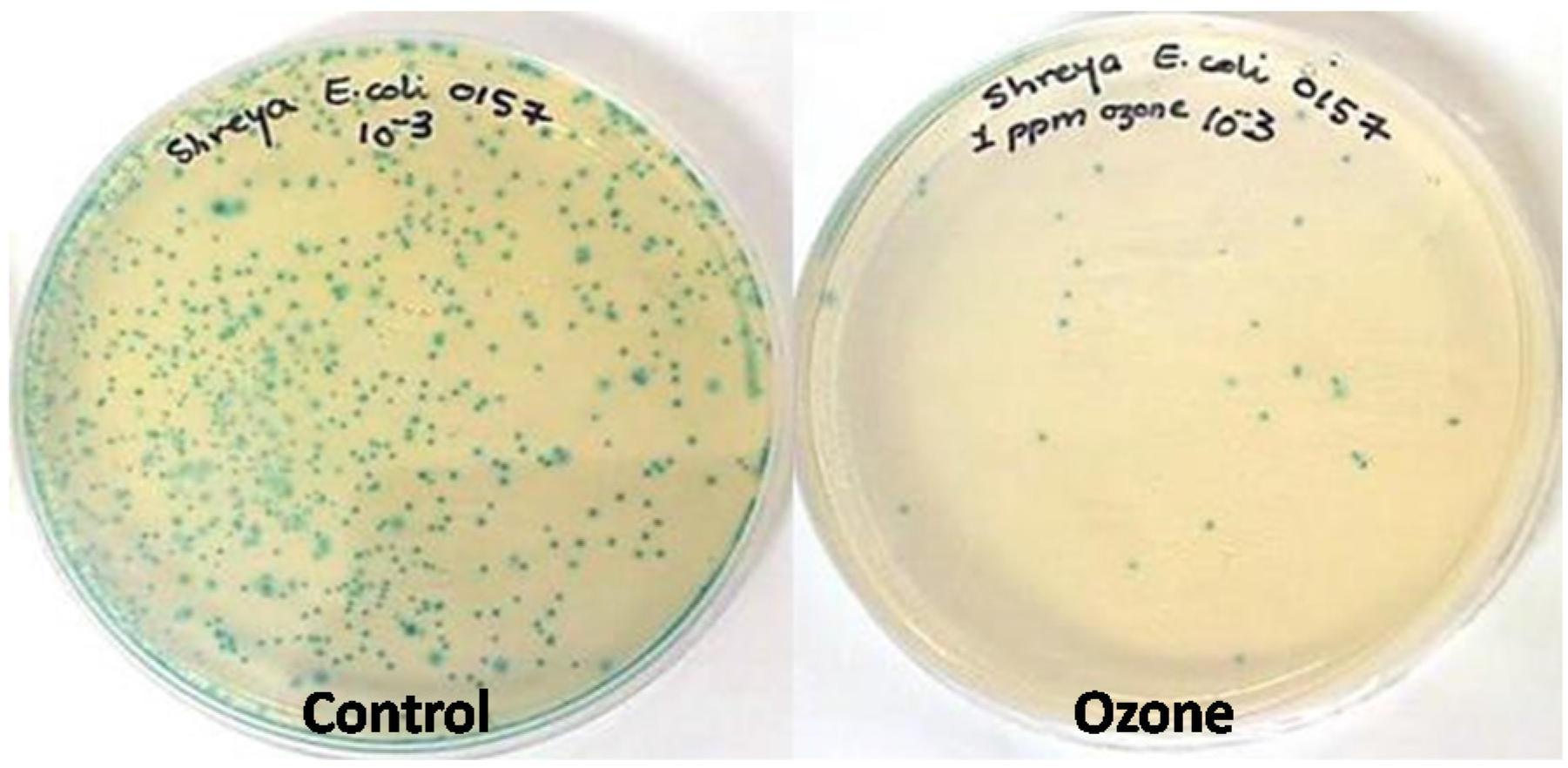
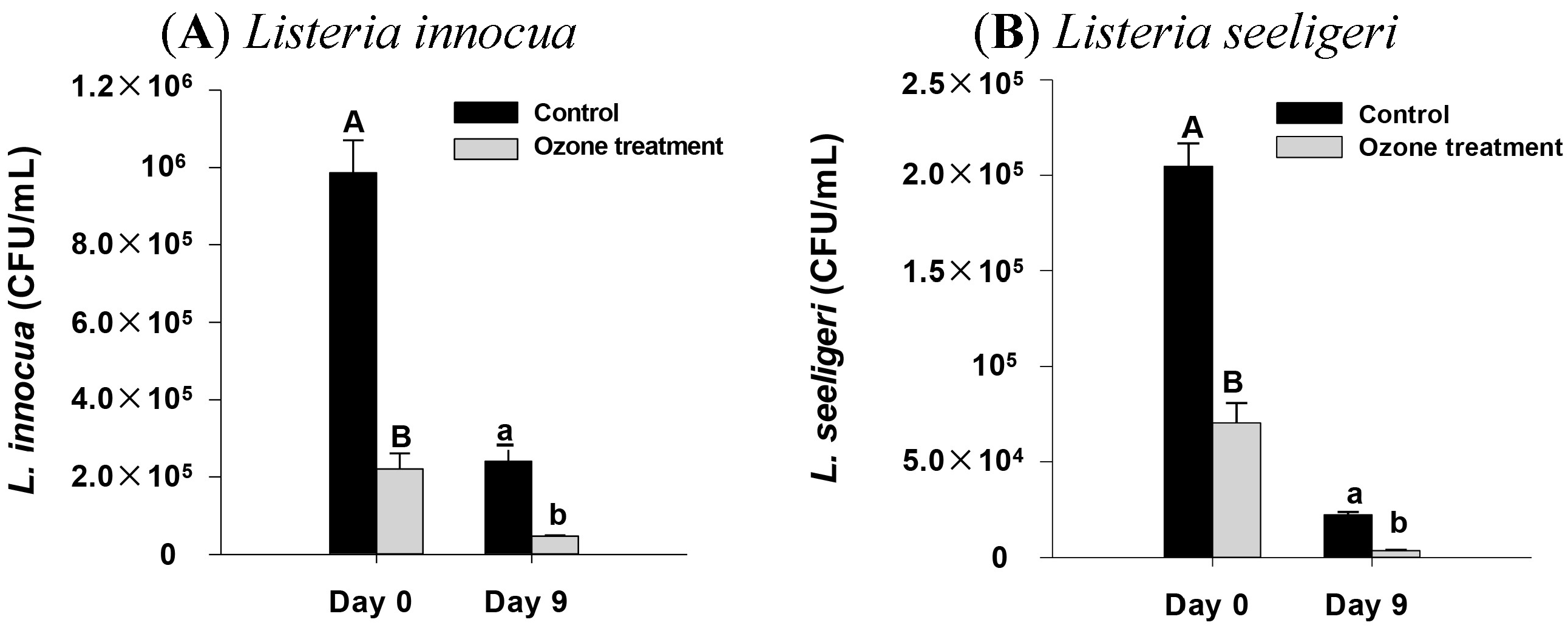
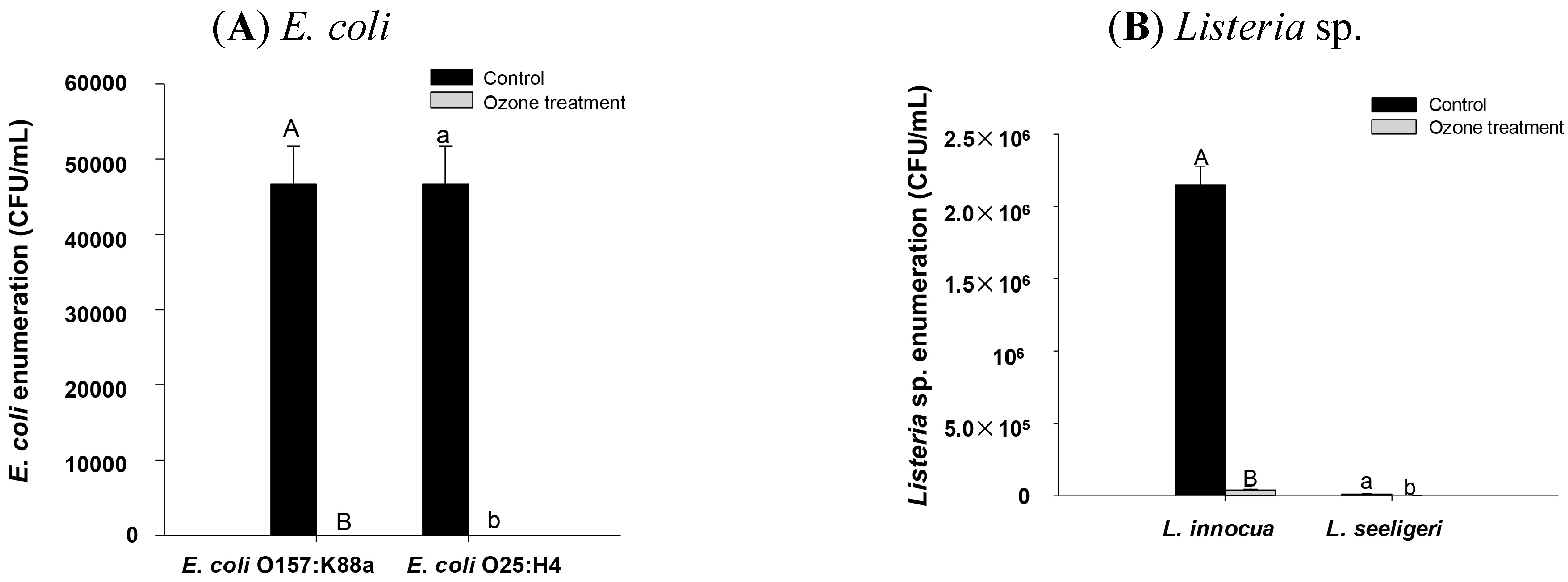
2.4. Impact of Ozone Treatment on Listeria innocua and L. seeligeri Inoculated onto Spinach Leaves
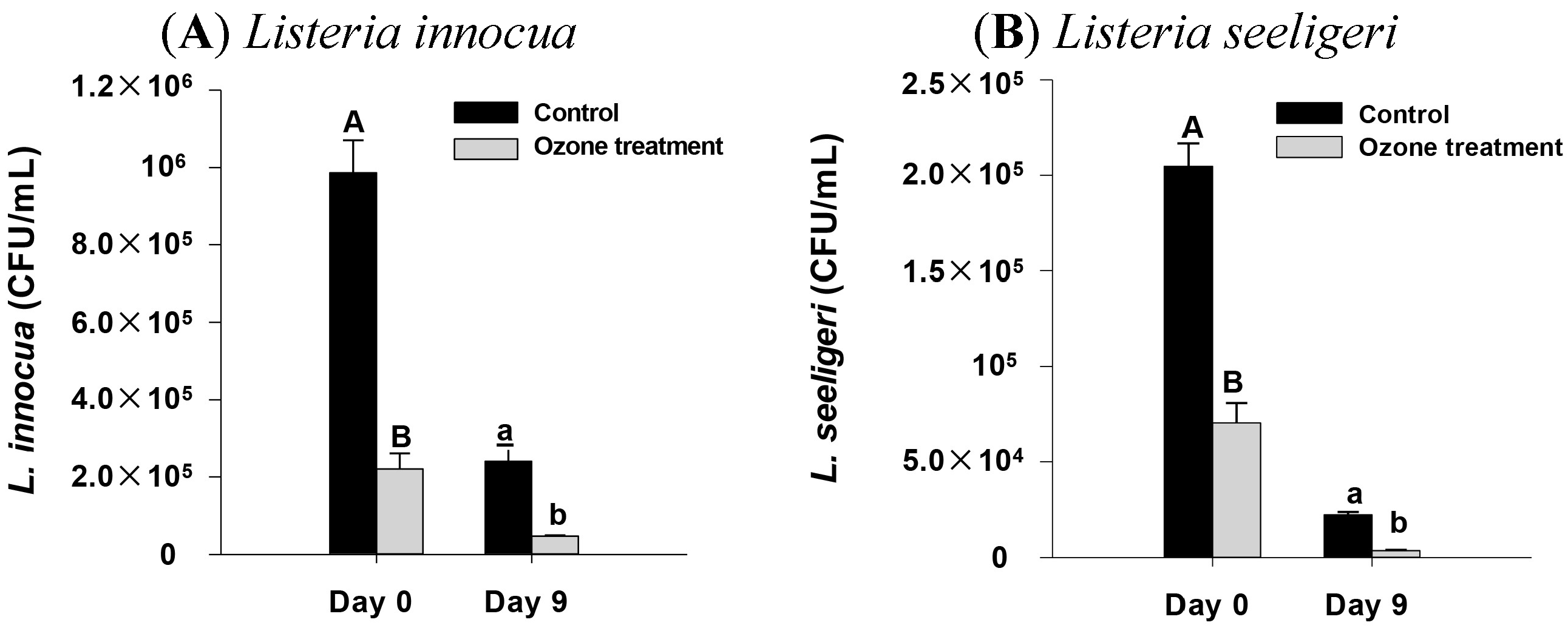
2.5. Effect of Higher Ozone Treatment on E. coli and Listeria sp. Inoculated onto Spinach Leaf Surface

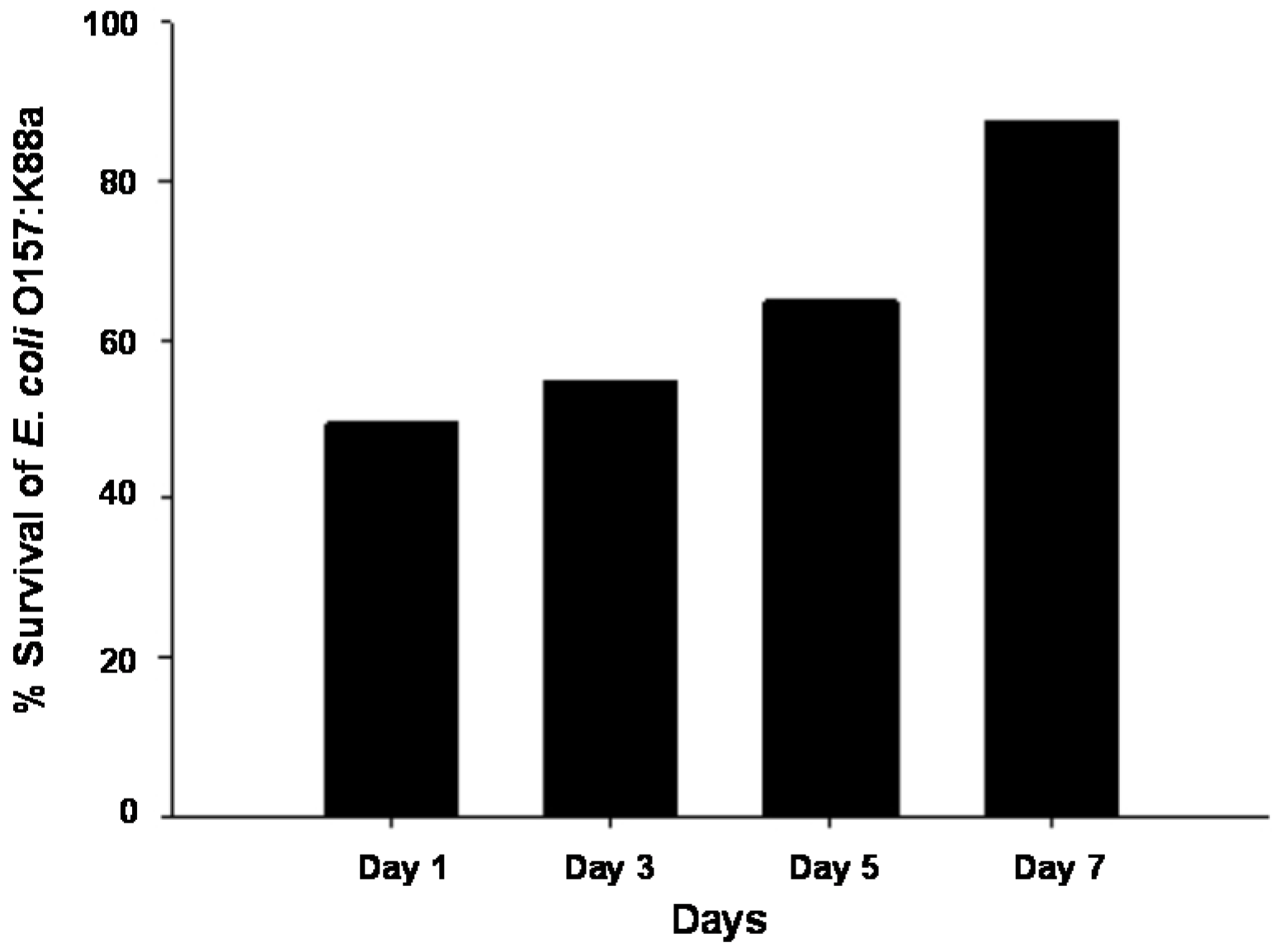
2.6. Effect of Age on Ozone Resistance of E. coli O157:K88a in Vitro
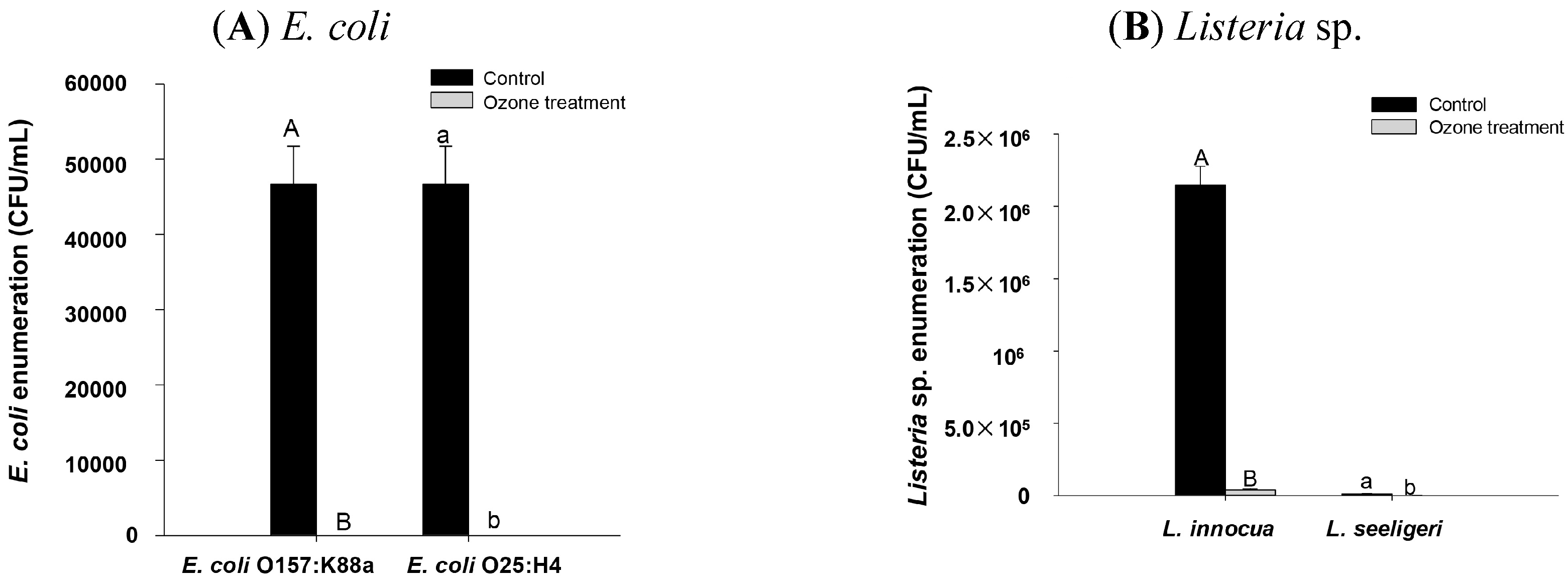
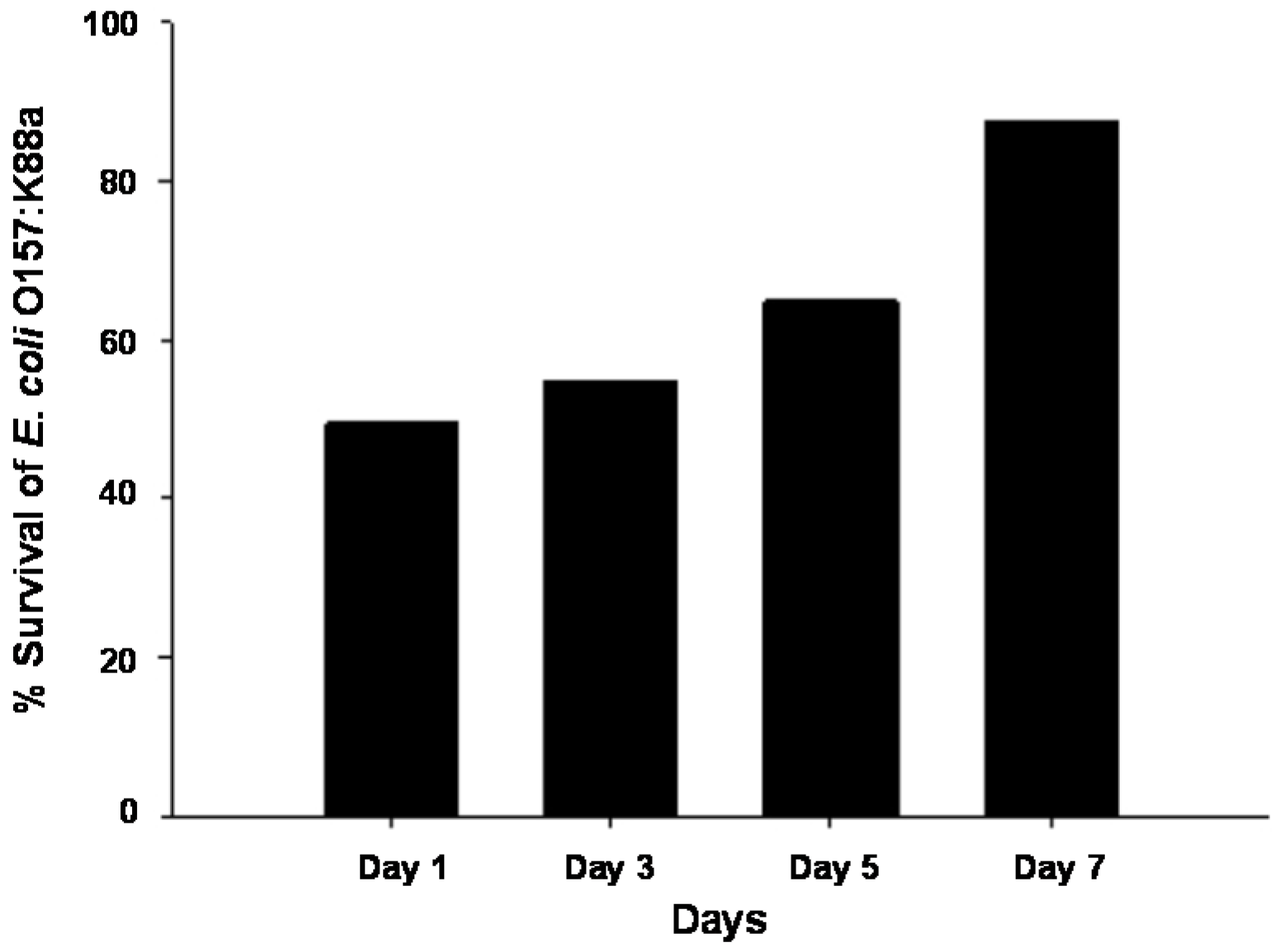
3. Experimental Section
3.1. Ozone Fumigation System

3.2. Assessing the Impact of Ozone Treatment on Food Pathogens E. coli and L. innocua in Vitro
3.3. Optimization of Ozone Exposure Levels (Concentration and Duration) to Treat Leafy Salads without Causing Visual Damage to Produce
3.4. Ozone Resistance of Different Strains of E. coli: Inoculation of E. coli onto Spinach Leaves and Ozone Exposure Conditions
3.5. Impact of Ozone Treatment on L. innocua and L. seeligeri Inoculated onto Spinach Leaves
3.6. Modified Ozone Fumigation System—Delivery of High Ozone Concentrations for Short Time Durations (Seconds)

3.7. Exploration of Higher Ozone Exposure Levels to Treat Spinach without Causing Visual Damage
3.8. Impact of Higher/Increased Ozone Concentrations on Two Strains of E. coli and Listeria Inoculated onto Spinach Leaves
3.9. Age Effects on Ozone Resistance of E. coli in Vitro
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Burnett, S.L.; Beuchat, L.R. Human pathogens associated with raw produce and unpasteurized juices, and difficulties in decontamination. J. Ind. Microbiol. Biotechnol 2000, 25, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Abadias, M.; Usall, J.; Anguera, M.; Solsona, C.; Vinas, I. Microbiological quality of fresh, minimally-processed fruit and vegetables, and sprouts from retail establishments. Int. J. Food Microbiol. 2008, 123, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Velusamy, V.; Arshak, K.; Korostynska, O.; Oliwa, K.; Adley, C. An overview of foodborne pathogen detection: In the perspective of biosensors. Biotechnol. Adv. 2010, 28, 232–254. [Google Scholar] [CrossRef] [PubMed]
- Olaimat, A.N.; Holley, R.A. Factors influencing the microbial safety of fresh produce: A review. Food Microbiol. 2012, 32, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Engels, C.; Weiss, A.; Carle, R.; Schmidt, H.; Schieber, A.; Ganzle, M.G. Effect of gallotannin treatment on attachment, growth and survival of Escherichia coli O157:H7 and Listeria monocytogenes on spinach and lettuce. Eur. Food Res. Technol. 2012, 234, 1081–1090. [Google Scholar] [CrossRef]
- Griffin, P.M.; Tauxe, R.V. The epidemiology of infections caused by Escherichia coli O157:H7, other enterohemorrhagic E.coli, and the associated Hemolytic Uremic Syndrome. Epidemiol. Rev. 1991, 13, 60–99. [Google Scholar] [PubMed]
- Tomas-Callejas, A.; Lopez-Velasco, G.; Camacho, A.B.; Artes, F.; Artes-Hernandez, F.; Suslow, T.V. Survival and distribution of Escherichia coli on diverse fresh-cut baby leafy greens under preharvest through postharvest conditions. Int. J. Food Microbiol. 2011, 151, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Coia, J.E. Clinical, microbiological and epidemiological aspects of Escherichia coli O157 infection. FEMS Immunol. Med. Microbiol. 1998, 20, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Farber, J.M.; Peterkin, P.I. Listeria monocytogenes, a Food-Borne pathogen. Microbiol. Rev. 1991, 55, 476–511. [Google Scholar] [PubMed]
- Notermans, S.; Todd, E.C.D. Surveillance of listeriosis and its causative pathogen, Listeria monocytogenes. Food Control 2011, 22, 1484–1490. [Google Scholar] [CrossRef]
- Launders, N.; Byrne, L.; Adams, N.; Glen, K.; Jenkins, C.; Tubin-Delic, D.; Locking, M.; Williams, C.; Morgan, D.; Outbreak Control Team. Outbreak of Shiga toxin-producing E.coli O157 associated with consumption of watercress, United Kingdom, August to September 2013. Euro Surveill. 2013, 18. [Google Scholar] [CrossRef]
- Bower, C.K.; Daeschel, M.A. Resistance responses of microorganisms in food environments. Int. J. Food Microbiol. 1999, 50, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Mahapatra, A.K.; Muthukumarappan, K.; Julson, J.L. Applications of ozone, bacteriocins and irradiation in food processing: A review. Crit. Rev. Food Sci. Nutr. 2005, 45, 447–461. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Singh, R.K.; Bhunia, A.K.; Stroshine, R.L. Efficacy of chlorine dioxide, ozone, and thyme essential oil or a sequential washing in killing Escherichia coli O157:H7 on lettuce and baby carrots. LWT Food Sci. Technol. 2002, 35, 720–729. [Google Scholar] [CrossRef]
- Goncalves, A.A. Ozone—An emerging technology for the seafood industry. Braz. Arch. Biol. Technol. 2009, 52, 1527–1539. [Google Scholar] [CrossRef]
- Tzortzakis, N.; Borland, A.; Singleton, I.; Barnes, J. Impact of atmospheric ozone-enrichment on quality-related attributes of tomato fruit. Postharvest Biol. Technol. 2007, 45, 317–325. [Google Scholar] [CrossRef]
- Karaca, H.; Velioglu, Y.S. Effects of ozone treatments on microbial quality and some chemical properties of lettuce, spinach, and parsley. Postharvest Biol. Technol. 2014, 88, 46–53. [Google Scholar] [CrossRef]
- Alwi, N.A.; Ali, A. Reduction of Escherichia coli O157, Listeria monocytogenes and Salmonella enterica sv. Typhimurium populations on fresh-cut bell pepper using gaseous ozone. Food Control 2014, 46, 304–311. [Google Scholar] [CrossRef]
- Yuk, H.G.; Yoo, M.Y.; Yoon, J.W.; Marshall, D.L.; Oh, D.H. Effect of combined ozone and organic acid treatment for control of Escherichia coli O157:H7 and Listeria monocytogenes on enoki mushroom. Food Control 2007, 18, 548–553. [Google Scholar] [CrossRef]
- Fan, L.; Song, J.; McRae, K.B.; Walker, B.A.; Sharpe, D. Gaseous ozone treatment inactivates Listeria innocua in vitro. J. Appl. Microbiol. 2007, 103, 2657–2663. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, E.; O’Beirne, D. Effects of process severity on survival & growth of Escherichia coli & Listeria innocua on minimally processed vegetables. Food Control 2005, 16, 677–685. [Google Scholar] [CrossRef]
- O’Beirne, D.; Francis, A.G. Effects of the indigenous microflora of minimally processed lettuce on the survival and growth of Listeria innocua. Int. J. Food Sci. Technol. 1998, 33, 477–488. [Google Scholar] [CrossRef]
- Rico, D.; Martín-Diana, A.B.; Barat, J.M.; Barry-Ryan, C. Extending and measuring the quality of fresh-cut fruit and vegetables: A review. Trends Food Sci. Technol. 2007, 18, 373–386. [Google Scholar] [CrossRef]
- Alexopoulos, A.; Plessas, S.; Ceciu, S.; Lazar, V.; Mantzourani, I.; Voidarou, C.; Stavropouloua, E.; Bezirtzogloua, E. Evaluation of ozone efficacy on the reduction of microbial population of fresh cut lettuce (Lactuca sativa) and green bell pepper (Capsicum annuum). Food Control 2013, 30, 491–496. [Google Scholar] [CrossRef]
- Klockow, P.A.; Keener, K.M. Safety and quality assessment of packaged spinach treated with a novel ozone-generation system. LWT Food Sci. Technol. 2009, 42, 1047–1053. [Google Scholar] [CrossRef]
- Bialka, K.L.; Demirci, A. Decontamination of Escherichia coli O157:H7 and Salmonella enterica on blueberries using ozone and pulsed UV-light. J. Food Sci. 2007, 72, 391–396. [Google Scholar] [CrossRef]
- Akbas, M.Y.; Ozdemir, M. Application of gaseous ozone to control populations of Escherichia coli, Bacillus cereus and Bacillus cereus spores in dried figs. Food Microbiol. 2008, 25, 386–391. [Google Scholar] [CrossRef]
- Scifò, G.O.; Randazzo, C.L.; Restuccia, C.; Fava, G.; Caggia, C. Listeria innocua growth in fresh cut mixed leafy salads packaged in modified atmosphere. Food Control 2009, 20, 611–617. [Google Scholar] [CrossRef]
- Bermúdez-Aguirre, D.; Barbosa-Cánovas, G.V. Disinfection of selected vegetables under nonthermal treatments: Chlorine, acid citric, ultraviolet light and ozone. Food Control 2013, 29, 82–90. [Google Scholar] [CrossRef]
- Zuma, F.; Lin, J.; Jonnalagadda, S.B. Ozone-initiated disinfection kinetics of Escherichia coli in water. J. Environ. Sci. Health A Tox. Hazard. Subst. Environ. Eng. 2009, 44, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Vaz-Velho, M.; Silva, M.; Pessoa, J.; Gibbs, P. Inactivation by ozone of Listeria innocua on salmon-trout during cold-smoke processing. Food Control 2006, 17, 609–619. [Google Scholar] [CrossRef]
- Navarro Llorens, J.M.; Tormo, A.; Martinez-Garcia, E. Stationary phase in gram-negative bacteria. FEMS Microbiol. Rev 2010, 34, 476–495. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wani, S.; Maker, J.K.; Thompson, J.R.; Barnes, J.; Singleton, I. Effect of Ozone Treatment on Inactivation of Escherichia coli and Listeria sp. on Spinach. Agriculture 2015, 5, 155-169. https://doi.org/10.3390/agriculture5020155
Wani S, Maker JK, Thompson JR, Barnes J, Singleton I. Effect of Ozone Treatment on Inactivation of Escherichia coli and Listeria sp. on Spinach. Agriculture. 2015; 5(2):155-169. https://doi.org/10.3390/agriculture5020155
Chicago/Turabian StyleWani, Shreya, Jagpreet K. Maker, Joseph R. Thompson, Jeremy Barnes, and Ian Singleton. 2015. "Effect of Ozone Treatment on Inactivation of Escherichia coli and Listeria sp. on Spinach" Agriculture 5, no. 2: 155-169. https://doi.org/10.3390/agriculture5020155