An Alternative Use of Horticultural Crops: Stressed Plants as Biofactories of Bioactive Phenolic Compounds



Abstract
:1. Introduction
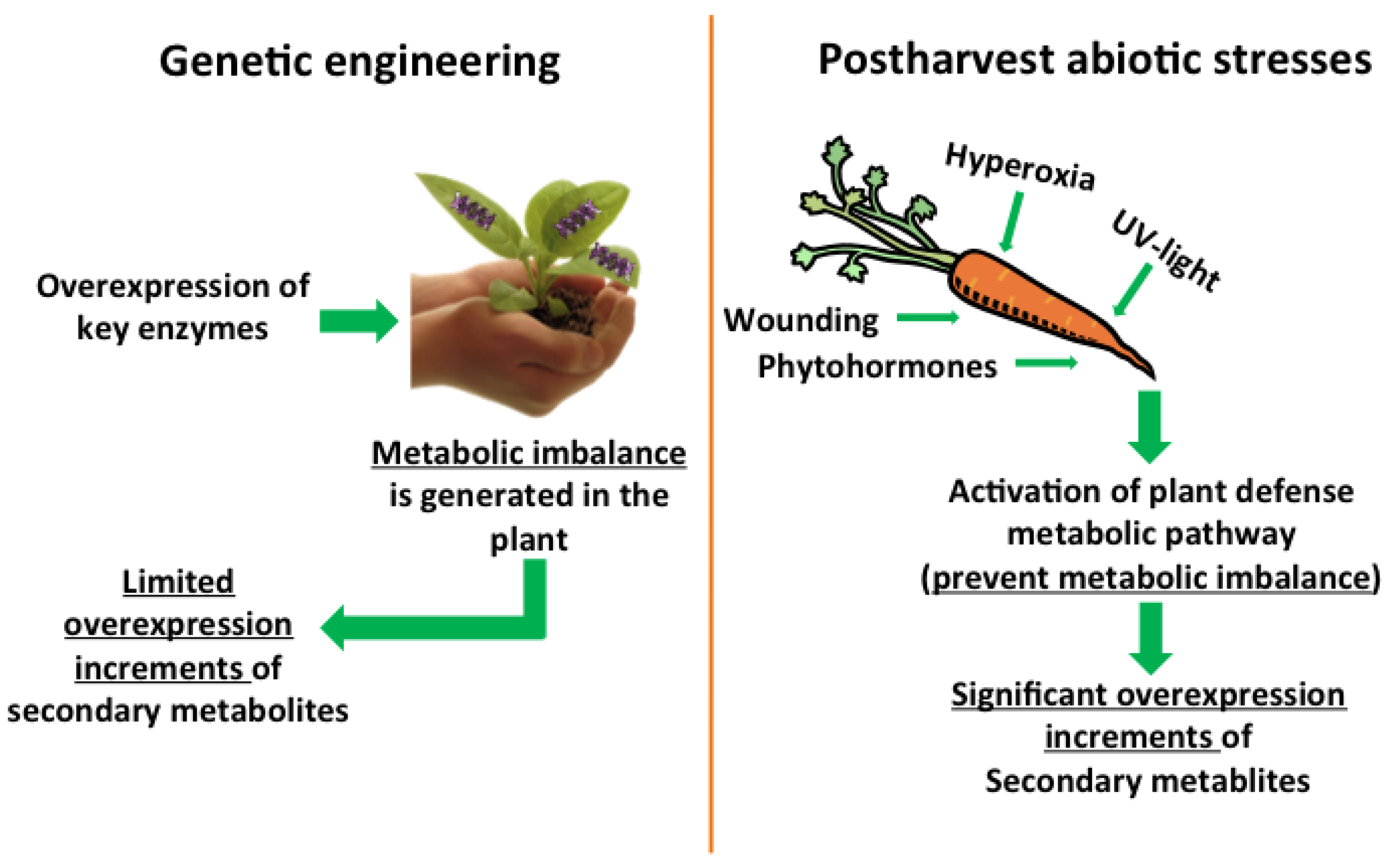
2. Plants as Biofactories of Phenolic Compounds: Use of Abiotic Stresses
2.1. Carrots as Biofactories of Hydroxycinnamic Acids
2.1.1. Chlorogenic Acid (CQA)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Abiotic stress applied | Storage conditions | Phenolic content (mg/100 g FW) | Increase (%) | Reference | |
|---|---|---|---|---|---|---|
| Before * | After | |||||
| Chlorogenic acid | W | 4 d/15 °C | 5.3 | 44.70 | 743 | [4] |
| W + UV-C | 4 d/15 °C | 5.3 | 75.52 | 1324 | [4] | |
| W + E | 6 d/15 °C | 14.8 | 92.6 | 526 | [9] | |
| W + H | 2 d/20 °C | 1.62 | 68.8 | 4148 | [11] | |
| 4,5-Dicaffeoylquinic acid | W | 4 d/15 °C | 4.59 | 13.23 | 188 | [4] |
| W + UV-C | 4 d/15 °C | 4.59 | 14.00 | 205 | [4] | |
| W + E | 6 d/15 °C | 3.2 | 28.1 | 778 | [9] | |
| W + H | 2 d/20 °C | 0.13 | 7.1 | 6191 | [11] | |
| 3,5-Dicaffeoylquinic acid | W | 4 d/15 °C | nd | 4.91 | - | [4] |
| W + UV-C | 4 d/15 °C | nd | 6.03 | - | [4] | |
| W + H | 2 d/20 °C | nd | 1.65 | - | [11] | |
2.1.2. Chlorogenic Acid Derivatives: 4,5-diCQA and 3,5-diCQA.
3. Physiological and Molecular Basis for the Accumulation of Phenolics as a Stress Response
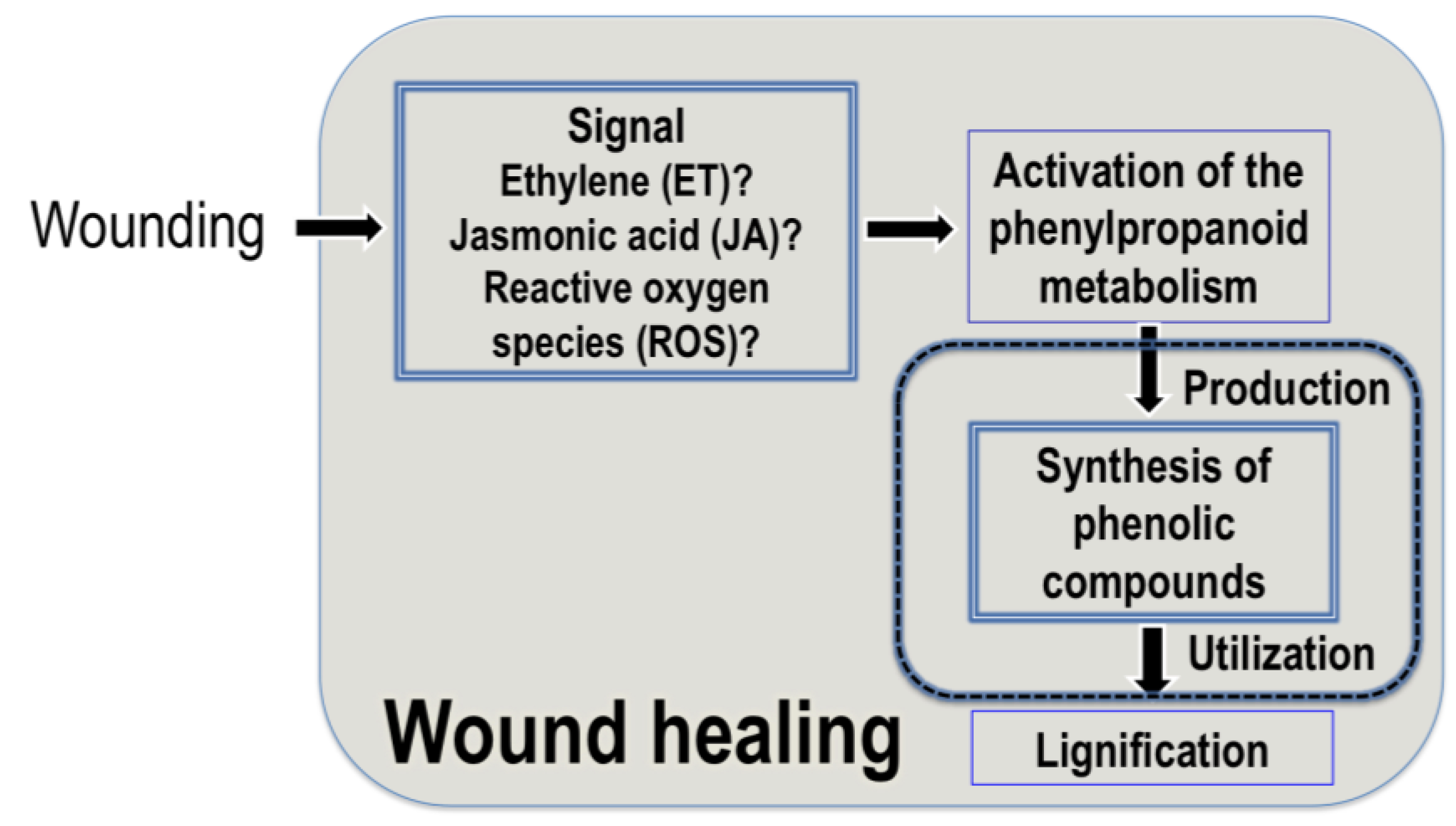
3.1. Wound Healing and Accumulation of Hydroxycinnamic Acids in Plants
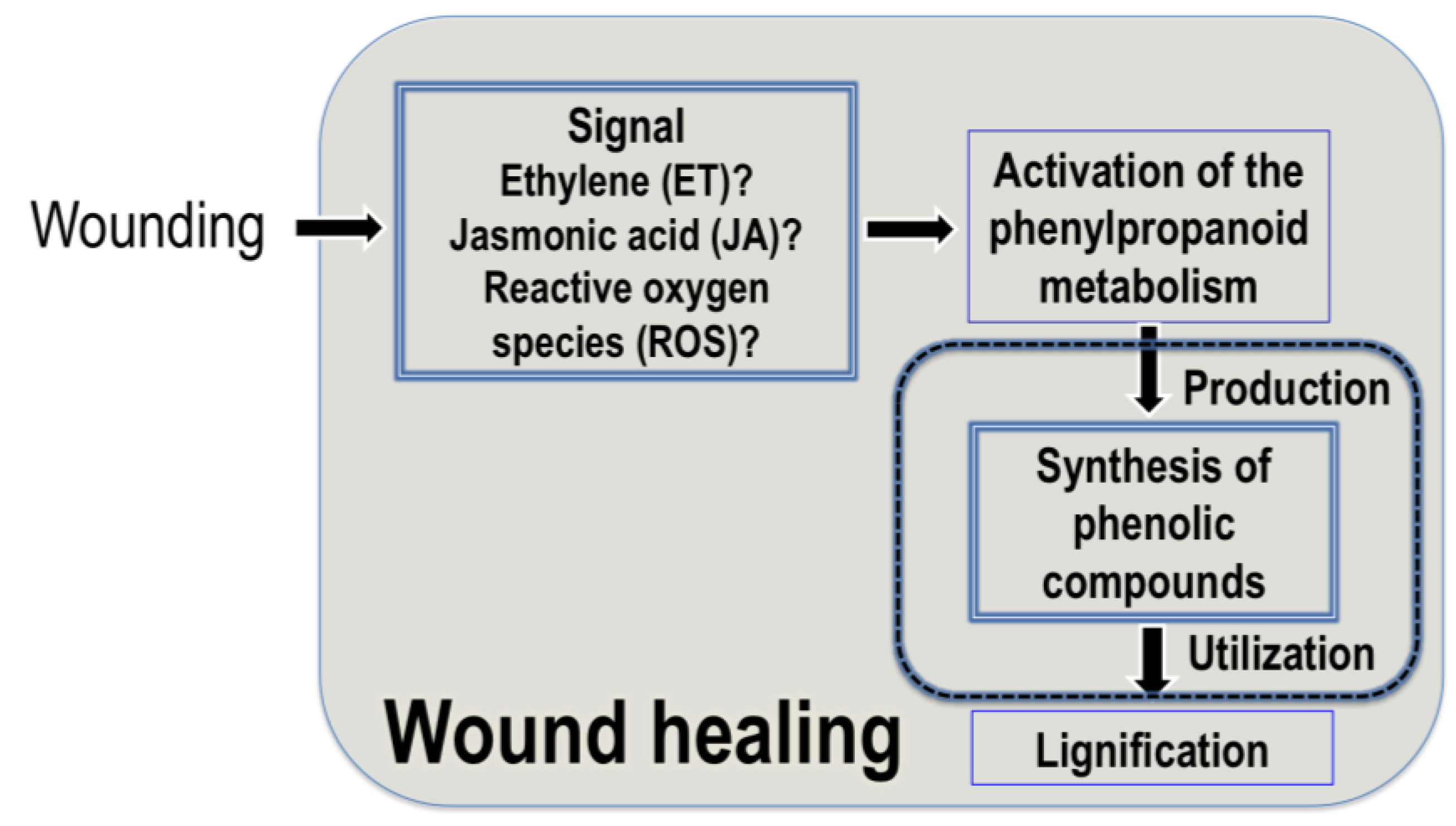
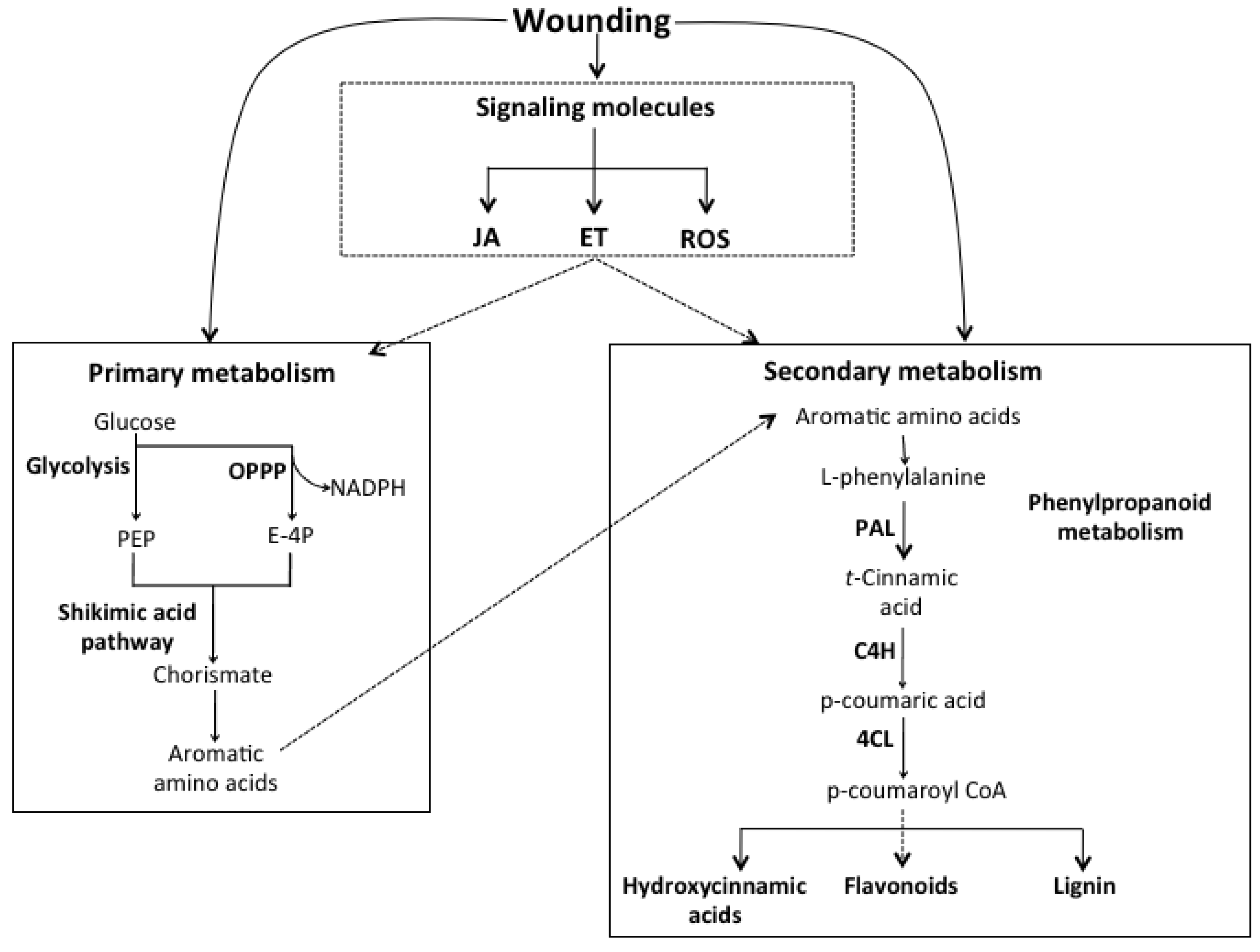
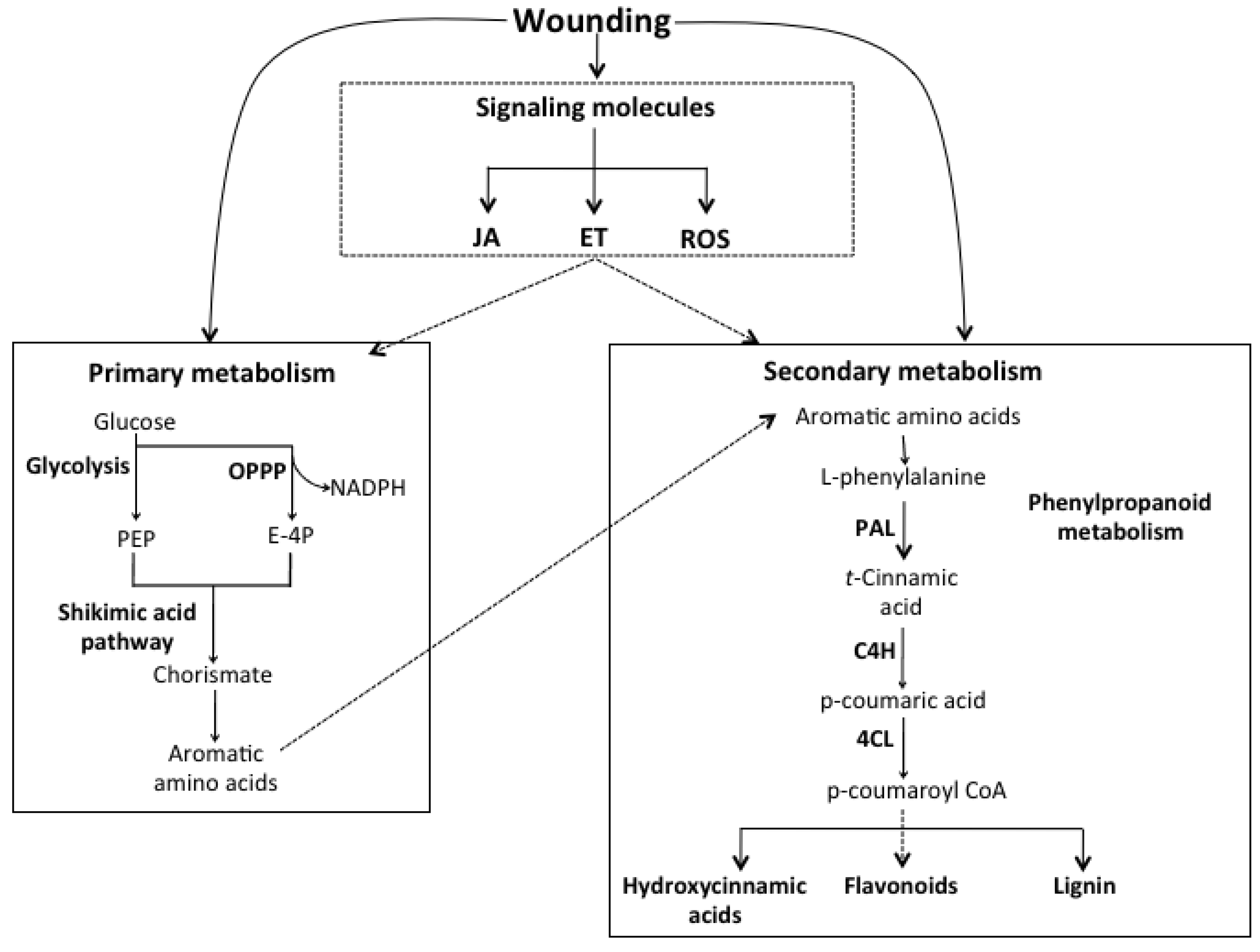
3.2. Signaling Molecules Involved in the Wound-Induced Activation of the Phenylpropanoid Metabolism
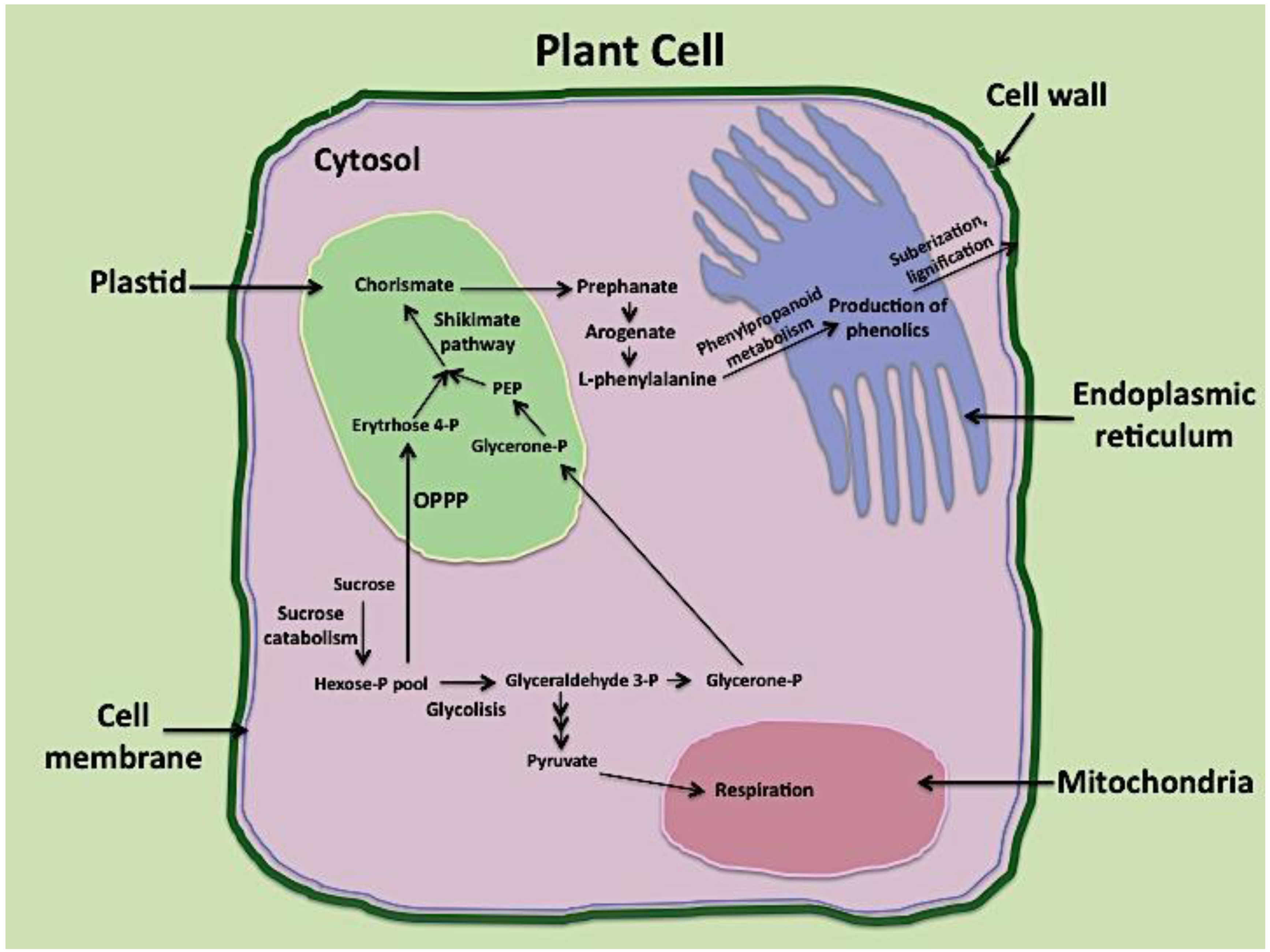
3.3. Wounding and the Synthesis of Aromatic Amino Acids (Carbon Source for Phenolics Biosynthesis)
4. Conclusions
Acknowledgments
References
- Flinn, J.E.; Zavon, J.A. Green plants as biofactories for drugs. Biopharm. Int. 2004, 17, 42–49. [Google Scholar]
- Colwell, R.K.; Norse, E.A.; Pimentel, D.; Sharples, F.E.; Simberloff, D. Genetic engineering in agriculture. Science 1985, 229, 111–112. [Google Scholar]
- Cisneros-Zevallos, L. The use of controlled postharvest abiotic stresses as a tool for enhancing the nutraceutical content and adding-value of fresh fruits and vegetables. J. Food Sci. 2003, 68, 1560–1565. [Google Scholar] [CrossRef]
- Surjadinata, B.B. Wounding and Ultraviolet Radiation Stresses Affect the Phenolic Profile and Antioxidant Capacity of Carrot Tissue. Ph.D. Thesis, Texas A&M University, College Station, TX, USA, 2006. [Google Scholar]
- Heredia, J.B. The Enhancement of Fresh Produce Antioxidant Capacity by Wounding Stress and Phytohormones. Ph.D. Thesis, Texas A&M University, College Station, TX, USA, 2006. [Google Scholar]
- Jacobo-Velázquez, D.A. Physiological and Molecular Mechanisms Governing the Postharvest Stress-Induced Accumulation of Antioxidant Phenolic Compounds in Carrots. Ph.D. Thesis, Texas A&M University, College Station, TX, USA, 2010. [Google Scholar]
- Surjadinata, B.B.; Cisneros-Zevallos, L. Biosynthesis of phenolic antioxidants in carrot tissue increases with wounding intensity. Food Chem. 2012, 134, 615–624. [Google Scholar] [CrossRef]
- Heredia, J.B.; Cisneros-Zevallos, L. The effect of exogenous ethylene and methyl jasmonate on the accumulation of phenolic antioxidants in selected whole and wounded fresh produce. Food Chem. 2009, 115, 1500–1508. [Google Scholar] [CrossRef]
- Heredia, J.B.; Cisneros-Zevallos, L. The effect of exogenous ethylene and methyl jasmonate on pal activity, phenolic profiles and antioxidant capacity of carrots (Daucus carota) under different wounding intensities. Postharvest Biol. Technol. 2009, 51, 242–249. [Google Scholar] [CrossRef]
- Jacobo-Velázquez, D.A.; Cisneros-Zevallos, L. Correlations of antioxidant activity against phenolic content revisited: A new approach in data analysis for food and medicinal plants. J. Food Sci. 2009, 74, R107–R113. [Google Scholar] [CrossRef]
- Jacobo-Velázquez, D.A.; Martínez-Hernández, G.B.; Rodríguez, S.; Cao, C.-M.; Cisneros-Zevallos, L. Plants as biofactories: Physiological role of reactive oxygen species on the accumulation of phenolic antioxidants in carrot tissue under wounding and hyperoxia stress. J. Agric. Food Chem. 2011, 59, 6583–6593. [Google Scholar]
- Ye, X.; Al-Babili, S.; Klöti, A.; Zhang, J.; Lucca, P.; Beyer, P.; Potrykus, I. Engineering the provitamin A (beta411 carotene) biosynthetic pathway into (carotenoid-free) rice endosperm. Science 2000, 287, 303–305. [Google Scholar]
- Niggeweg, R.; Michael, A.J.; Martin, C. Engineering plants with increased levels of the antioxidant chlorogenic acid. Nat. Biotechnol. 2004, 22, 746–754. [Google Scholar] [CrossRef]
- Diretto, G.; Al-Babili, S.; Tavazza, R.; Papacchioli, V.; Beyer, P.; Giuliano, G. Metabolic engineering of potato carotenoid content through tuber-specific over expression of a bacterial mini-pathway. PLoS ONE 2007, 2, e350. [Google Scholar] [CrossRef]
- Vinson, J.A.; Su, X.; Zubik, L.; Bose, P. Phenol antioxidant quantity and quality in foods: Fruits. J. Agric. Food Chem. 2001, 49, 5315–5321. [Google Scholar] [CrossRef]
- Heo, H.J.; Kim, Y.J.; Chung, D.; Kim, D.-O. Antioxidant capacities of individual and combined phenolics in a model system. Food Chem. 2007, 104, 87–92. [Google Scholar] [CrossRef]
- Reyes, L.F.; Cisneros-Zevallos, L. Wounding stress increases the phenolic content and antioxidant capacity of purple-flesh potatoes (Solanum tuberosum L.). J. Agric. Food Chem. 2003, 51, 5296–5300. [Google Scholar]
- Reyes, L.F.; Villarreal, J.E.; Cisneros-Zevallos, L. The increase in antioxidant capacity after wounding depends on the type of fruit and vegetable tissue. Food Chem. 2007, 101, 1254–1262. [Google Scholar] [CrossRef]
- Olthof, M.R.; Hollman, P.C.H.; Katan, M.B. Chlorogenic acid and caffeic acid are absorbed in humans. J. Nutr. 2001, 131, 66–71. [Google Scholar]
- Olthof, M.R.; Hollman, P.C.H.; Buijsman, M.N.C.P.; van Amelsvoort, J.M.M.; Katan, M.B. Chlorogenic acid, quercetin-3-rutinoside and black tea phenols are extensively metabolized in humans. J. Nutr. 2003, 133, 1806–1814. [Google Scholar]
- Farah, A.; Monteiro, M.; Donangelo, C.M.; Lafay, S. Chlorogenic acids from green coffee extract are highly bioavailabe in humans. J. Nutr. 2008, 138, 2309–2315. [Google Scholar] [CrossRef]
- Thom, E. The effect of chlorogenic acid enriched coffee on glucose absorption in healthy volunteers and its effect on body mass when used long-term in overweight and obese people. J. Int. Med. Res. 2007, 35, 900–908. [Google Scholar]
- Wang, G.-F.; Shi, L.-P.; Ren, Y.-D.; Liu, Q.-F.; Liu, H.-.F.; Zhang, R.-J.; Li, Z.; Zhu, F.-H.; He, P.-L.; Tang, W.; et al. Anti-hepatitis B virus activity of chlorogenic acid, quinic acid and caffeic acid in vitro and in vivo. Antivir. Res. 2009, 83, 186–190. [Google Scholar] [CrossRef]
- Belkaid, A.; Currie, J.-C.; Desgagnés, J.; Annabi, B. The chemopreventive properties of chlorogenic acid reveal a potencial new role for the microsomal glucose-6-phosphate translocase in brain tumor progression. Cancer Cell Int. 2006, 6, 7:1–7:12. [Google Scholar]
- Robinson, W.E., Jr.; Cordeiro, M.; Abdel-Malek, S.; Jia, Q.; Chow, S.A.; Reinecke, M.G.; Mitchell, W.M. Dicaffeoylquinic acid inhibitors of human immunodeficiency virus integrase: Inhibition of the core catalytic domain of human immunodeficiency virus integrase. Mol. Pharmacol. 1996, 50, 845–855. [Google Scholar]
- Zhu, K.; Cordeiro, M.L.; Atienza, J.; Robinson, W.E., Jr.; Chow, S.A. Irreversible inhibition of human immunodeficiency virus type 1 integrase by dicaffeoylquinic acids. J. Virol. 1999, 73, 3309–3316. [Google Scholar]
- Choi, J.; Park, J.K.; Lee, K.T.; Park, K.K.; Kim, W.B.; Lee, J.H.; Jung, H.J.; Park, H.J. In vivo antihepatotoxic effects of Ligularia fischeri var. spiciformis and the identification of the active component, 3,4-dicaffeoylquinic acid. J. Med. Food 2005, 8, 348–352. [Google Scholar]
- Kim, S.-S.; Park, R.-Y.; Jeon, H.-J.; Kwon, Y.-S.; Chun, W. Neuroprotective effect of 3,5-dicaffeoylquinic acid on hydrogen peroxide-induced cell death in SH-SY5Y cells. Phytother. Res. 2005, 19, 243–245. [Google Scholar] [CrossRef]
- Rittinger, P.A.; Biggs, A.R.; Peirson, D.R. Histochemistry of lignin and suberin deposition in boundary layers formed after wounding in various plant species and organs. Can. J. Bot. 1987, 65, 1886–1892. [Google Scholar] [CrossRef]
- Boerjan, W.; Ralph, J.; Baucher, M. Lignin biosynthesis. Annu. Rev. Plant Biol. 2003, 54, 519–546. [Google Scholar] [CrossRef]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar]
- Franke, R.; Schreiber, L. Suberin—A biopolyester forming apoplastic plant interfaces. Curr. Opin. Plant Biol. 2007, 10, 252–259. [Google Scholar] [CrossRef]
- Cottle, W.; Kolattukudy, P.E. Biosynthesis, deposition, and partial characterization of potato suberin phenolics. Plant Physiol. 1982, 69, 393–399. [Google Scholar] [CrossRef]
- Tomás-Barerán, F.A.; Loaiza-Velararde, J.; Bonfanti, A.; Saltveit, M.E. Early wound- and ethylene-induced changes in phenylpropanoid metabolism in harvested lettuce. J. Am. Soc. Hort. Sci. 1997, 122, 399–404. [Google Scholar]
- Davey, M.W.; Franck, C.; Keulemans, J. Distribution, developmental and stress responses of antioxidant metabolism in Malus. Plant Cell Environ. 2004, 27, 1309–1320. [Google Scholar] [CrossRef]
- Saltveit, M.E. Effect of 1-methylcyclopropene on phenylpropanoid metabolism, the accumulation of phenolic compounds, and browning of whole and fresh-cut ‘iceberg’ lettuce. Postharvest Biol. Technol. 2004, 34, 75–80. [Google Scholar] [CrossRef]
- Kim, Y.J.; Kim, D.G.; Lee, S.H.; Lee, I. Wound-induced expression of the ferulate 5-hydroxylase gene in Camptotheca acuminata. Biochim. Biophys. Acta 2006, 1760, 182–190. [Google Scholar] [CrossRef]
- Choi, Y.-J.; Tomás-Barberán, F.A.; Saltveit, M.E. Wound-induced phenolic accumulation and browning in lettuce (Lactuca sativa L.) leaf tissue is reduced by exposure to n-alcohols. Postharvest Biol. Technol. 2005, 37, 47–55. [Google Scholar]
- Bernards, M.A.; Razem, F.A. The poly(phenolic) domain of potato suberin: A non-lignin cell wall bio-polymer. Phytochemistry 2001, 57, 1115–1122. [Google Scholar] [CrossRef]
- Razem, F.A.; Bernards, M.A. Reactive oxygen species production in association with suberization: Evidence for an NADPH-dependent oxidase. J. Exp. Bot. 2003, 54, 935–941. [Google Scholar] [CrossRef]
- Herrmann, K.M.; Weaver, L.M. The shikimate pathway. Annu. Rev. Plant Phys. 1999, 50, 473–503. [Google Scholar] [CrossRef]
- Amthor, J.S. Efficiency of lignin biosynthesis: A quantitative analysis. Ann. Bot. 2003, 91, 673–695. [Google Scholar] [CrossRef]
- Surjadinata, B.B.; Cisneros-Zevallos, L. Modeling wound-induced respiration of fresh-cut carrots (Daucus carota L.). J. Food Sci. 2003, 68, 2735–2740. [Google Scholar]
- Klotz, K.L.; Finger, F.L.; Anderson, M.D. Wounding increases glycolytic but not soluble sucrolytic activities in stored sugarbeet root. Postharvest Biol. Technol. 2006, 41, 48–55. [Google Scholar] [CrossRef]
- Pryke, J.A.; Rees, T. The pentose phosphate pathway as a source of NADPH for lignin synthesis. Phytochemistry 1977, 16, 557–560. [Google Scholar] [CrossRef]
- Dyer, W.E.; Henstrand, J.M.; Handa, A.K.; Herrmann, K.M. Wounding induces the first enzyme of the shikimate pathway in Solanaceae. Proc. Natl. Acad. Sci. USA 1989, 86, 7370–7373. [Google Scholar]
- Sharma, R.; Jain, M.; Bhatnagar, R.K.; Bhalla-Sarin, N. Differential expression of DAHP synthase and chorismate mutase in various organs of Brassica juncea and the effect of external factors on enzyme activity. Physiol. Plant. 1999, 105, 739–745. [Google Scholar] [CrossRef]
- Keith, B.; Dong, X.N.; Ausubel, F.M.; Fink, G.R. Differential induction of 3-deoxy-D-arabino-heptulosonate 7-phosphate synthase genes in Arabidopsis thaliana by wounding and pathogenic attack. Proc. Natl. Acad. Sci. USA 1991, 88, 8821–8825. [Google Scholar] [CrossRef]
- Corpas, F.J.; Barroso, J.B.; Sandalio, L.M.; Distefano, S.; Palma, J.M.; Lupiáñez, J.A.; del Río, L.A. A dehydrogenase-mediated recycling system of NADPH in plant peroxisomes. Biochem. J. 1998, 330, 777–784. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jacobo-Velázquez, D.A.; Cisneros-Zevallos, L. An Alternative Use of Horticultural Crops: Stressed Plants as Biofactories of Bioactive Phenolic Compounds. Agriculture 2012, 2, 259-271. https://doi.org/10.3390/agriculture2030259
Jacobo-Velázquez DA, Cisneros-Zevallos L. An Alternative Use of Horticultural Crops: Stressed Plants as Biofactories of Bioactive Phenolic Compounds. Agriculture. 2012; 2(3):259-271. https://doi.org/10.3390/agriculture2030259
Chicago/Turabian StyleJacobo-Velázquez, Daniel A., and Luis Cisneros-Zevallos. 2012. "An Alternative Use of Horticultural Crops: Stressed Plants as Biofactories of Bioactive Phenolic Compounds" Agriculture 2, no. 3: 259-271. https://doi.org/10.3390/agriculture2030259
APA StyleJacobo-Velázquez, D. A., & Cisneros-Zevallos, L. (2012). An Alternative Use of Horticultural Crops: Stressed Plants as Biofactories of Bioactive Phenolic Compounds. Agriculture, 2(3), 259-271. https://doi.org/10.3390/agriculture2030259