
Effects of Knotweed-Enriched Feed on the Blood Characteristics and Fitness of Horses


1
Department of Forest Botany, Dendrology and Geobiocoenology, Mendel University in Brno, Zemědělská 1665/1, 613 00 Brno, Czech Republic
2
Department of Mycorrhizal Symbioses, Institute of Botany of the CAS, 25243 Průhonice, 100 00 Prague, Czech Republic
3
Dibaq, Helvíkovice 90, 564 01 Žamberk, Czech Republic
4
Veterinary Laboratory, Jirchářská 217, 517 41 Kostelec nad Orlicí, Czech Republic
*
Author to whom correspondence should be addressed.
Agriculture 2022, 12(1), 109; https://doi.org/10.3390/agriculture12010109
Submission received: 28 November 2021
/
Revised: 5 January 2022
/
Accepted: 10 January 2022
/
Published: 13 January 2022
(This article belongs to the Special Issue Safety and Efficacy of Feed Additives in Animal Production)
Abstract
:The aboveground biomass of dry knotweed was administered daily to large groups of young (1- to 3-year-old) stallions of the Czech Warmblood, Czech-Moravian Coldblood and Silesian Norik breeds, fed individually for 4 and 6 months in two successive winter experiments. Their fitness was compared with control groups consisting of equally numerous subgroups comparable in age, breed, body mass and initial blood parameters. The effects of knotweed on the horses’ fitness were evaluated based on changes in blood characteristics. Even if administered in small amounts, 150 g per day, knotweed could (1) increase the thrombocyte numbers, (2) increase the globulin content (thus improving the horses’ immunity, which is desired in large groups of animals), (3) stimulate lipid metabolism in cold-blooded horses and (4) decrease the concentration of cholesterol. The long-lasting effect of knotweed on both the urea and triglyceride–cholesterol ratio presumably reflected, between the two experiments, the temporary protein starvation of horses on pastures with poor quality of grass in a dry summer.
1. Introduction
Many countries have been trying to eradicate knotweed due to its invasiveness, although it has many practical uses, including improving the health of animals through feed supplementation. This report suggests that it could be used as a feed supplement for horses, thereby improving their fitness. This study was part of a larger project in which knotweed had been administered to several animal species, the health of which improved.
Japanese knotweed (Reynoutria japonica, syn. Polygonum cuspidatum), including its hybrid (R. × bohemica), has been thoroughly studied from a range of perspectives, both as an “enemy” of native flora due to its invasiveness and as a plant with many beneficial uses, including positive health effects [1,2,3,4]. As a source of resveratrol and other substances with anti-ageing and other positive effects on both animals and humans, the entire knotweed plant can also be expected to have positive effects on the health of horses. Resveratrol has already been used in horses with positive results. Resveratrol is, however, extracted mainly from roots and rhizomes; the aboveground portions of knotweed, which contain equally powerful antioxidants, deserve more thorough investigation [5]. Neochlorogenic acid was identified as an efficient antioxidant in the aboveground knotweed biomass [6]. High amounts of carotenoids in knotweed leaves are comparable to those in spinach [7]. In addition, knotweed plants produce a significant number of bioactive constituents, namely, phenolic substances derived from resveratrol, such as piceid, piceatannol, astringin and emodin, which are also found in aboveground biomass albeit in smaller quantities than in rhizomes and roots.
Among the bioactive components of knotweed, resveratrol and, more recently, piceid (also called polydatin) have been thoroughly studied. Resveratrol was found to modulate many different pathways, as it binds to numerous cell signalling molecules, modulates cell regulatory genes, activates transcription factors, suppresses pro-inflammatory genes’ expression and inflammatory biomarkers, induces antioxidant enzymes and inhibits protein kinases and the expression of angiogenic and metastatic gene products. It thus has strong potential as a treatment for inflammatory, cardiovascular, pulmonary and age-related diseases such as cancer, diabetes and even Alzheimer’s disease. These chronic illnesses and neurological and autoimmune diseases are accompanied by the dysregulation of multiple cell regulating pathways and are connected with inflammation. Resveratrol targets sirtuin, adenosine monophosphate kinase, nuclear factor-κB, inflammatory cytokines, anti-oxidant enzymes along with cellular processes such as gluconeogenesis, lipid metabolism, mitochondrial biogenesis, angiogenesis and apoptosis. Immunity is regulated by its interfering with immune cell regulation, pro-inflammatory cytokines’ synthesis and gene expression [8]. Similarly, piceid regulates lipid metabolism, helps treat cardiovascular diseases [9] and diabetic cardiomyopathy via its anti-inflammatory and anti-oxidative effects [10], preserves mitochondrial function in the central nervous system, offers a therapeutic option for Spinal cord ischemia/reperfusion injury [11], decreases the levels of reactive oxygen species in neurons from the ischemic cortex, ameliorates oxidative stress and mitochondria-dependent apoptosis [12], fights neurodegenerative diseases including Alzheimer’s, Parkinson’s, cognition/memory dysfunction, brain/spinal cord injuries, ischemic stroke and miscellaneous neuronal dysfunctionalities [13], promotes a radiosensitising effect on osteosarcoma cancer cells, reduces clonogenic survival of tumor cells and induces osteogenic differentiation, alone and in the presence of ionising therapy [14]. Emodin has recently garnered increased interest, with refs. [15,16] reporting its antiviral and anti-inflammatory activities against SARS-CoV-2, for the reason that it is one of the components of the Chinese medicine LQF, the Lianhua-Qingwen formula based on 11 herbs with 61 compounds that is used for the prevention and treatment of viral diseases, including coronavirus disease 2019 (COVID-19).
Three resveratrol analogue glucosides, namely, piceid, piceatannol glucoside and resveratroloside, were found to have antibacterial effects [11], and their bioactivity was comparable to that of resveratrol, which is released in the gut due to glucoside hydrolysis. Piceid exhibited more powerful effects than resveratrol against hepatitis B virus [17,18]. Piceatannol had even more efficient anti-inflammatory, immunomodulatory, anti-proliferative, anti-leishmanial, and anti-leukaemic activities than resveratrol. Resveratroloside competitively inhibited α-glucosidase, thus alleviating postprandial hyperglycemia in diabetic mice [19]. Crude extract from knotweed roots and rhizomes inhibited such troublesome bacteria as Staphylococus aureus, Pseudomonas aeruginosa, Acinetobacter baumannii, Bacillus cereus, Escherichia coli, Listeria monocytogenes and Salmonella anatum [20,21].
It might therefore be expected that there are even greater health effects of knotweed plants than there are of their individual components.
In horses, resveratrol has already been used with positive results. Inflammation is accompanied by increased enzymatic activity of the granulocytic enzyme myeloperoxidase (MPO), resulting in the production of the highly oxidative acid HOCl [22,23]. Resveratrol was shown [24] to substantially decrease HOCl production and mitigate inflammation in horses. Inhibitory effects of resveratrol on equine neutrophil myeloperoxidase were described [25,26]. It was found that four weeks of supplementation with 1 g/day resveratrol in old horses decreased the inflammation-induced production of cytokinin both in vitro and in vivo, indicating that resveratrol has substantial potential for the treatment of acute and chronic inflammatory horse diseases [27]. Resveratrol was also reported to reduce the gene expression of inflammatory mediators, thus allowing even old horses to move freely in training and competitions [28,29]. Resveratrol has been administered to performance horses in a number of food supplements, such as Equithrive Joint® and Resverasyn®, with the aims of reducing the effects of laminitis and slowing ageing.
Metabolic efficiency in animals decreases with age, leading to increased levels of creatine kinase and glucose. A decrease in both creatine kinase and glucose levels in horses receiving resveratrol and hyaluronic acid (EquithriveJoint®) was found [30]. EquithriveJoint® was administered to old, lame horses and a decreased serum malondialdehyde (MDA) concentration and modulated serum levels of glutathionperoxidase (GPx), catalase and superoxiddismutase (SOD) were found, indicating protective effects of EquithriveJoint® against oxidative stress and ageing [31]. A beneficial effect of EquithriveJoint® on the performance of horses with hindlimb lameness treated with triamcinolone was also demonstrated [32]. Supplementation with these plant-derived phenolics in old horses was suggested [33] to lower the doses of nonsteroidal anti-inflammatory drugs and thus reduce their side effects. Since laminitis and other diseases impair older horses, the effects of resveratrol on the health of young horses are unclear.
Based on the above evidence, there is good reason to assume that knotweed would help treat horse health issues in a similar way to resveratrol. Although the diseases occur mainly in older horses, the aim of this project was to find out whether knotweed can also improve the fitness of young horses. Reports on the health effects of supplementation with knotweed herbs in animals are scarce; a drug containing knotweed (Praziver®) helped cure equine helminthiasis [34].
Knotweed is not only one of the best sources of resveratrol, its derivatives, carotenoids [7] and neochlorogenic acid [6], but it also contains other substances, the effects of which are still not well known. A conservative approach to the use of knotweed as a dietary supplement in animals thus persists, although there have been many reports of various animal species grazing on it without problems. It is also on the list of safe plants for fodder, as it was introduced to Europe for use as feed for domesticated animals. Aboveground parts of knotweed are consumed by humans in some areas, e.g., in Japan and North America [6].
This study thus aimed to fill the knowledge gap regarding the effects of knotweed on the health status of young horses. We performed two consecutive experiments, during which the dry aboveground knotweed biomass was administered as a dietary supplement to a large number of young stallions.
2. Materials and Methods
2.1. Horses and Husbandry
Most of the horses involved in the two experiments belonged to the warmblood breed known as the Czech Warmblood, while the minority of coldblood horses belonged to two breeds, the Czech-Moravian Belgian Horse, a breed constituted from the Czech Coldblood and Moravian Coldblood, and the Silesian Norik. These horses were kept at the Regional Stud Farm Tlumačov, Czech Republic, which provides breeding services, rears foals and young stallions and performs early testing and training of stallions. Most of the stallions are sold at the age of 2–3 years, and only the selected ones are further kept for breeding and training. In October 2021, i.e., 2 years after the experiments, those warmblood horses that had been involved in the experiments and later sold to the horse keepers in the Czech Republic who got them involved in sport activities, were tracked in the records of the Czech Equestrian Federation: https://www.jezdectvi.org/kone (accessed on 25 October 2021).
In winter, the horses were kept in individual boxes on a deep straw bed and fed individually with hay, with a feed supplement and with an experimental mixture, which were always consumed without leftovers. From May to October/November they stayed on pastures at grass only. In boxes, only the 3-year-old stallions were under training, receiving 30 min of daily exercise. All of the horses were healthy and under veterinary control.
2.2. Experimental Design
Two feeding experiments in two successive winters were run with 74 and 62 young stallions. Experiment 1 lasted from 10 January 2018 to 10 May 2018, Experiment 2 from 12 November 2018 to 14 May 2019. In Experiment 1, there were 15 horses born in 2015, 24 horses born in 2016 and 35 horses born in 2017. From those 24 horses born in 2016 and 35 horses born in 2017, 11 and 27 horses, respectively, were also included in Experiment 2, according to the rule that the knotweed-fed horses from Experiment 1 were also knotweed-fed in Experiment 2 to avoid affecting the control horses with recent treatment. In addition, 24 new horses born in 2018 were included in Experiment 2. Each group of 1–3 years old stallions (foals) was divided in both seasons into two subgroups based on age, body weight and blood test values. The resulting experimental groups were not significantly different before the experiments in any of the baseline parameters, including weight, blood test results, age, breed and/or parentage. The health status of the stallions was monitored by testing blood samples.
2.3. Feed and Feeding Regimen
All stallions were fed individually in boxes, receiving hay ad libitum, a regular feed supplement and the experimental mixture, which were always consumed without leftovers. As they were kept on a deep straw bed, straw also became an occasional component of their diet.
Each horse was fed daily with:
- (1)
- Local grass hay. The real consumption of hay per day, per horse by the 1-year-old, 2-year-old and 3-year-old warmbloods was 4, 5.5 and 8 kg hay, respectively; by the 1-year-old and 2-year-old coldbloods, it was 6 and 10 kg hay, respectively. For the nutrient content, see Table 1;
- (2)
- A regular feed supplement for foals and stallions in amounts covering their nutrient requirements with respect to their age and breed, i.e., 1.5 and 2.0 kg for 1-year old, 2.0 and 2.5 kg for 2-year old and 2.5 and 3.0 kg for 3-year old warm- and cold-blooded horses, supplied by a regional feed producer RenoFarmy (Troubky, Czech Republic), specially prepared for the horse breeding farm Tlumačov to meet the feeding requirements of the young, 1–3-year-old stallions kept there. The feed supplement was fed to them, together with grain oats. For the composition and amounts of the individual components of the feed supplement and grain oats, see Table 1;
- (3)
- 0.5 kg of the experimental mixture prepared and supplied by Dibaq (Helvíkovice, Czech Republic). For the composition and nutrient contents, see Table 1.
The amounts of the feed supplement and composition of the experimental mixture for individual horses respecting their breed, age, weight and condition were determined according to the nutrient requirements recommended by the NRC (National Research Council, 2007), based on experience with the development of similar feeds for horses enriched with bioactive compounds of plant origin, such as Fitmin herbs Regeneration, Fitmin herbs Calmer, or Fitmin Bronchial, see https://www.fitmin.cz (accessed on 30 October 2021).
The control group of horses (not fed knotweed) received 500 g/day/individual barley pellets with soya oil and beet sugar; horses from the treatment group (fed knotweed) received a mixture of dry aboveground knotweed biomass and barley with soya oil and beet sugar (500 g/day/individual). Each horse from the treatment group received daily 150 g of dry knotweed biomass containing 0.087 g of emodin, 0.028 g of resveratrol, 0.554 g of oxyresveratrol and 0.139 g of piceid. Oxyresveratrol, which is resveratrol with an additional -OH group, and piceid, which is resveratrol glucoside, split into glucose and resveratrol in the gut, thus contributing to the overall amount of available resveratrol administered daily to each individual horse, which ultimately amounted to 0.63 g. As some of these horses had been involved in both experiments, the stallions that had already received knotweed in Experiment 1 continued receiving it in Experiment 2.
There was a warm and dry season between the two experiments, from May to November 2018, during which the horses only fed on grass from pastures.
2.4. Measured Characteristics
2.4.1. Analysis of Resveratrol, Piceid, Astringin and Piceatannol in Knotweed Biomass
Finely ground (10 mm sieve) samples (500 mg) of dry aboveground biomass were extracted with 10 mL 60% ethanol, as it was the most efficient extractant for both resveratrol and its glucosides. Prior to analysis, the samples were filtered with syringes (NY, 0.2 µm). The extracts were analysed by a validated HPLC-UV method. Instrument: Shimadzu NEXERA with PDA detection 306 nm; column: Phenomenex synergi Hydro-RP 80A, 250 mm × 4.6 mm, 4µm (30 °C); flow rate: 1.5 mL.min−1; mobile phases: A–10 mM ammonium acetate at pH 4.15 using acetic acid, B-acetonitrile, concentration gradient from 7 to 90%. Standards: Sigma-Aldrich, Piceid: 98%, Resveratrol: 99%, Emodin: 98%, Astringin: 99%, Piceatannol: 99%. Mixed calibration was used for quantification. Validation report called Determination of selected biologically active substances present in knotweed by HPLC/UV method for AKP, has been deposited in the Library of VUOS in Pardubice Rybitví as SOP 568, VU 4661, 2007.
2.4.2. Blood Testing: Haematology and Biochemistry
At the beginning and end of each experiment, before the horses were released into pastures, they were weighed, and blood samples were drawn on the stud farm in the early morning for biochemical and haematological tests. Blood was drawn from the jugular vein and collected in a vacuum tube. One millilitre of blood was collected in a tube with EDTAK3 and was used for haematological tests, and 10 mL was collected in a tube with pro-coagulation gel and used for biochemical tests. The samples were kept cool during their rapid transport to the laboratory. Blood samples from Experiment 1 were analysed at the veterinary clinic in Brno, where they were analysed with a BC-2800 Vet haematological analyser (Mindray, Shenzhen, China) in horse mode. White blood cells were counted under a microscope in blood smears stained according to Pappenheim (May-Grünwald, Giemsa-Romanowski) in 200-cell subsamples. Blood samples from Experiment 2 were analysed in the Medila laboratory for biochemical parameters, and the haematological samples were analysed in the veterinary laboratory of the veterinary clinic Štrossovka s.r.o., Pardubice, using an automated haematological counter. The blood smears were simultaneously fixed and stained with Hemacolor®. The cellular elements (neutrophils, eosinophils, basophils, monocytes, lymphocytes and activated lymphocytes) were counted with the LeukoCounter system, and the individual populations of leukocytes were expressed as percentages of 200-cell subsamples. The following parameters were measured: total protein, albumin, urea, creatinine, cholesterol, glucose and triglyceride levels; AST, ALT and ALP activity; haemoglobin content (as the mean corpuscular haemoglobin (MCH), which is calculated by dividing the total weight of the haemoglobin by the total number of red blood cells in a particular volume of the blood sample and gives an average concentration of haemoglobin in each red blood cell throughout the sample); plateletcrit (PCT); and leucocyte, erythrocyte, thrombocyte, lymphocyte, neutrophil and monocyte counts. The albumin results could not be compared between the two experiments because different methods were used to determine the levels in Experiment 1 (photometry) and Experiment 2 (electrophoresis). As all biochemical laboratories in the Czech Republic use the same methods, maximum divergence should not exceed 20% and low bias due to lab change is to be expected.
2.5. Statistical Data Evaluation
The data were statistically evaluated with ANOVA (STATISTICA v.12, Dell Inc., Tulsa, OK, USA). One-way ANOVA (factor: experimental treatment—control or knotweed-fed) or two-way ANOVA (factors: experimental treatment, horse age or horse type) was applied. The mean differences between the control and knotweed-fed horses were measured. If homoscedasticity was not met, the Kruskal-Wallis test was used. Multivariate redundancy analysis (RDA) was performed using Canoco 5.0 (Biometris, Plant Research International, Wageningen, The Netherlands) [35]. Biochemical and haematological characteristics and their differences between the beginning and the end of the experiments were used as response data. Experimental treatment (control or knotweed-fed) was used as the explanatory variable.
3. Results
3.1. Blood Characteristics—Overview
Since body weight was not affected by knotweed in either experiment, only blood characteristics are dealt with here. Biochemical and haematological characteristics measured at the end of the two experiments are presented in Table 2. Besides the blood characteristics of significance described below, there were also a higher number of monocytes in knotweed-fed horses but only in Experiment 1.
The parameters shown in Figure 1 are the biochemical (Figure 1a) and haematological (Figure 1b) characteristics measured in the blood samples drawn at the end of Experiment 1, explaining the variability due to treatment—the closer a vector of a certain characteristic towards “Fed knotweed”, the more significantly would “feeding with knotweed” influence this characteristic. The similar direction of vectors indicates correlation among characteristics.
Additional variability is explained by the horse type and age, revealed in more detailed analyses of the individual characteristics described below. Only significantly different blood parameters are reported in the following section.
3.2. Blood Characteristics of Significance
3.2.1. Total Protein
The stallions fed knotweed had a significantly larger decrease in the amount of total protein in the oldest age group (born in 2016) than the control stallions (Experiment 2, Figure 2).
3.2.2. Globulin and Albumin/Globulin Ratio
Blood tests also revealed a statistically significant effect of knotweed on the globulin level in Experiment 1. Stallions fed knotweed had a higher level of globulin than horses in the control group (p = 0.05), and the difference was more distinct among warm-blooded horses (Figure 3).
Additionally, the albumin/globulin ratio was significantly affected by knotweed supplementation (p = 0.04); the effect of knotweed was significant in warm-blooded horses (Figure 4).
3.2.3. Thrombocytes, Plateletcrit (PCT) and Mean Cell Hemoglobin (MCH)
In Experiment 2, the stallions fed knotweed also produced more thrombocytes (p = 0.006) and PCT (p = 0.002) than the control stallions. The MCH increase in the stallions in the control group during Experiment 2 was more pronounced than that in the stallions fed knotweed (p = 0.02).
3.2.4. Activity of ALP
ALP activity decreased more in the blood of knotweed-fed horses than in the blood of the control horses (p = 0.05). If the warm-blooded and cold-blooded breeds were compared, the decrease was significant only in cold-blooded stallions (Figure 5).
This effect was significant in Experiment 1 but not in Experiment 2, in which a number of ALP values were missing.
3.2.5. Triglycerides
The level of triglycerides was higher in the cold-blooded horses fed knotweed than in the control ones (Figure 6).
3.2.6. Cholesterol
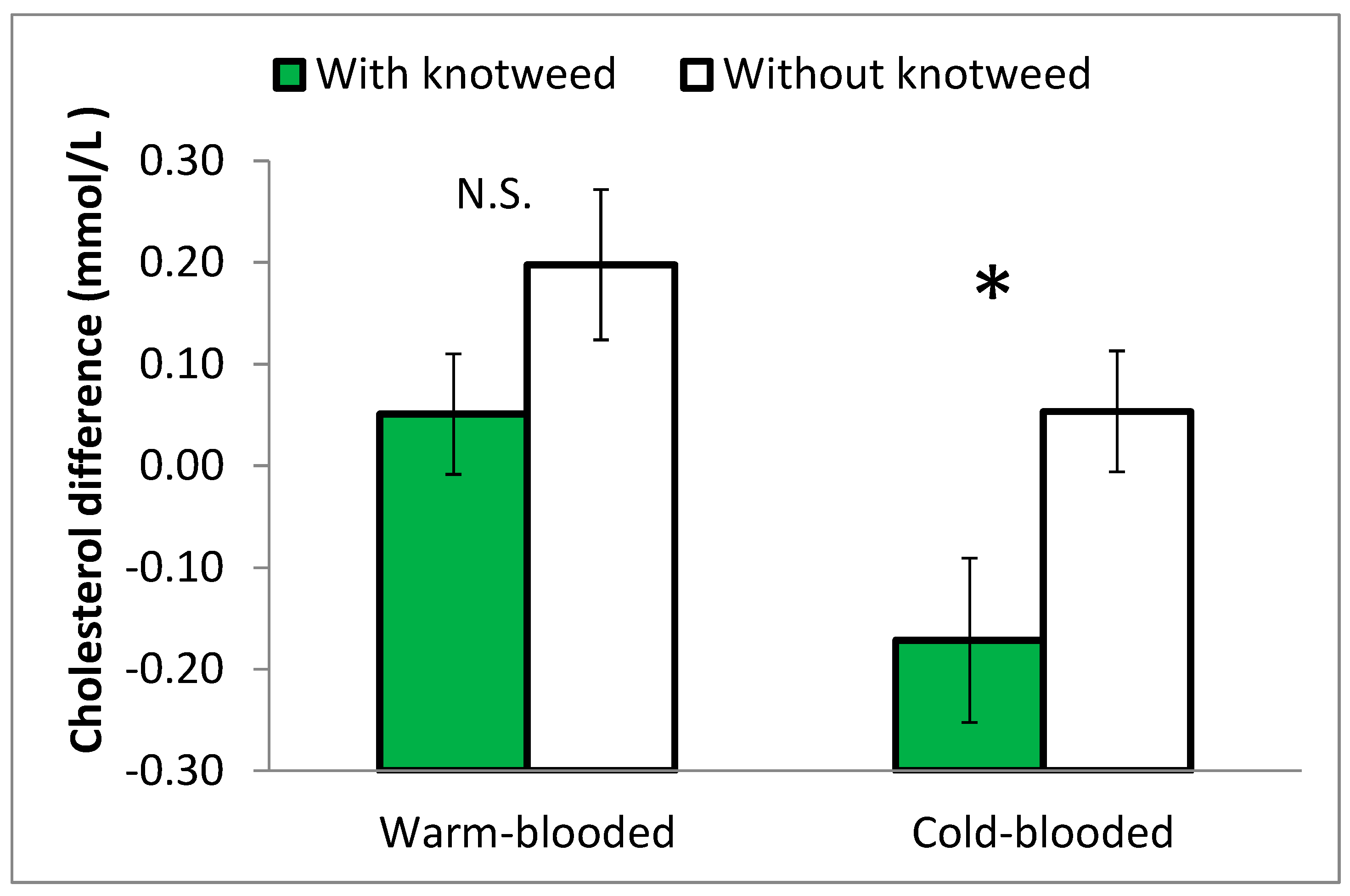
Both 2- and 3-year-old stallions fed knotweed showed a decrease in blood cholesterol levels during Experiment 2 while those not fed knotweed experienced an increase. The statistical difference between these two groups was significant at p = 0.022. However, when the warm-blooded and cold-blooded were compared separately, the effect of knotweed on cholesterol was only significant in the cold-blooded horses (Figure 7).
3.2.7. Urea
There was no difference in the urea blood levels between the knotweed-fed and control horses at the end of either of the two experiments. However, urea measured before the start of Experiment 2 was significantly higher (5.96 mmol L−1) in the 2- and 3-year-old horses, i.e., those who had been receiving knotweed in the previous winter and spring in Experiment 1, compared with the respective control horses (5.51 mmol L−1).
3.3. Sports Carrier of the Experimental Horses
Owing to a delay between the experiments and this report, it was possible to find out which warm-blooded horses were already in use in sports and what their first sport results were. By 20 October 2021, roughly equal numbers of control (14) and knotweed-fed (13) horses out of 26 in each category had already been registered for horse jumping and dressage competitions, and the control and knotweed-fed horses jumped over 104 and 109 cm high obstacles (in average; p = 0.3) and had been involved in 7 and 13 competitions (p = 0.15), respectively. These data are only of limited informative value as many other factors are involved, including socioeconomic ones. However, it is obvious that knotweed intake in feed restricted neither the fitness nor the performance of horses but rather supported it.
4. Discussion
4.1. Biochemistry and Haematology
Among the effects of knotweed on horse health, the following aspects are worth noting.
4.1.1. Total Protein, Globulins and Albumin/Globulin Ratio
The concentration of total protein represents the sum of the protein fractions in plasma: of albumin and all the fractions of globulins. Compensations occur between proteins in plasma because decreases or increases in their concentrations are common [36,37], which means that the concentration of total protein is totally dependent on the concentrations of these two groups of proteins.
Globulins are mainly involved in the immune response, transport of substances and coagulation [38]. In equine plasma, globulins account for a large proportion of the total protein. Five different groups of globulins are recognized: alpha 1 and alpha 2 globulins, beta 1 and beta 2 globulins, and gamma globulins. Altogether they account for 40–50% of the serum protein content, the remaining 50–60% of the plasma protein content is albumin. The albumin/globulin ratio is a convenient means of comparing values among horses.
In this study, concentrations of globulins in the blood of knotweed-fed horses were higher and the A/G ratio was lower than in the blood of control horses. All the values were within physiological limits and no pathology was observed in any of these experimental horses. Increased levels of globulins are often accompanied by decreased levels of albumins. Low levels of albumin may result from the decreased production of albumin or the increased loss of albumin via the kidneys, gastrointestinal tract, skin or extravascular space, or from increased catabolism of albumin [31]. Here, non-significantly lower level of albumin could indicate decreased absorption of proteins in the gut in the presence of knotweed, resulting in globulin increase, as globulins take over some of the functions of albumin. The increase of globulins suggests that the immunity of the horses was under non-specific pressure; the immunity of control horses was reduced, and their antibody levels decreased. On the contrary, the antibody levels in the stallions fed knotweed did not decline.
Increasing the plasma globulin concentration is desirable but difficult, as there are only a few means of doing so. Knotweed supplementation thus represents a suitable method.
4.1.2. Thrombocytes
Thrombocytes are non-nuclear blood elements that are crucial in the onset of blood clotting. Their number in the blood depends on their production in the bone marrow, consumption and losses. They account for the smallest proportion of blood cells. As their average lifespan in horse blood is 4–6 days [39], their population size sensitively reflects both their production and their destruction. They contain many substances involved in inflammatory reactions, blood coagulation and other specific reactions. It is impossible to assess immune function based on the thrombocyte count, as their absolute numbers do not correlate with thrombocyte function. Therefore, although stallions fed knotweed produced more thrombocytes than the control stallions in Experiment 2, the informative value of such a finding is limited with regard to the effect of knotweed supplementation on immunity. On the contrary, the number of thrombocytes decreased in the blood of knotweed-fed pigs in our previous study [40]. The different types of knotweed biomass in the feed supplements, i.e., stems and leaves in the supplement for horses, and rhizomes and roots in the supplement for the pigs (varying in their contents of bioactive substances, namely carotenoids, resveratrol, piceid and emodin), might help explain this difference and serve as a hint for further research. The decrease in thrombocyte numbers in pigs could be explained by high resveratrol content in knotweed roots [41,42] while the increase in thrombocyte numbers in horses could be due to other bioactive compounds in knotweed leaves and stems, such as carotenoids. It was reported that tomato puree rich in carotenoids improved mice health and increased their thrombocyte numbers in mice exposed to toxic fluorides [43]. The cellular membranes of thrombocytes in the carotene-deficient blood of people infected with the dengue virus suffered from dryness, which led to the death of these cells [44]. Carotenoids were not targeted in this study, however; their considerable content in knotweed leaves has been reported [7] and they deserve further study.
PCT, plateletcrit, is a number describing ratio between the volume of thrombocytes and plasma. A higher thrombocytes number means a higher PCT [42]. In this study, PCT was higher in the blood of knotweed-fed horses than in the blood of control ones.
4.1.3. Activity of Alkaline Phosphatase (ALP)
ALP is a non-specific metallo-enzyme (Mg, Zn), a hydrolase removing phosphate group from various proteins and nucleotides and hydrolyzing inorganic phosphate, produced in liver, bones, intestines, kidneys and placenta. Only activities of ALP isoenzymes from liver and bones, both coded by the same gene, could be detected here because the other ALP isoenzymes survive in the blood only for a short amount of time. ALP from bones is typical for young individuals whose bones are growing. When the growth slows down the activity of ALP physiologically decreases, being associated with mineralisation of bones [45]. The ALP, which was lower in the blood of knotweed-fed horses than in the blood of control horses in this study, was probably not produced in the liver but in the bones as there are no statistically significant changes in any other parameters connected with the liver (AST, GMT, bilirubin). It means that the decrease in ALP activity was associated with the physiological process of adolescence. In this study, the ALP decrease was enhanced by knotweed and correlated positively with urea (r = 0.411, p = 0.01) in knotweed-fed horses in Experiment 1, indicating a positive effect of knotweed on nitrogen use from the feed, as discussed below in Section 4.1.6. The ALP values from all the horses were within physiological limits [46].
4.1.4. Triglycerides
Triglycerides are another group of lipids involved in lipid metabolism that accounts for the majority of the lipids found in adipose tissue. They are synthesised primarily in adipose tissue, the liver, small intestine, and mammary glands. Circulating concentrations of triglycerides in normal animals reflect the balance in triglyceride absorption by the small intestine, synthesis/secretion by the hepatocytes and uptake by the adipose tissue. This balance is affected by the concentration of fat in the diet and by the production of hormones such as insulin and glucagon [47]. The triglyceride levels were significantly elevated in cold-blooded stallions to such an extent that the levels in cold-blooded and warm-blooded stallions were equal (Figure 6), which indicates that knotweed stimulated lipid metabolism in cold-blooded stallions but not in warm-blooded horses whose metabolic activities were already high.
Many studies ascribed the lowered concentration of triglycerides in the blood of different species to different active compounds from knotweed [48,49,50,51,52]. This study found the opposite: an increase of triglyceride concentration in the blood of knotweed-fed cold-blooded horses. The mechanism behind the increased triglycerides is not clear. This increase could be due to the different physiologies of horses or to specific active compounds from knotweed, or a combination of both. The anatomy and physiology of digestion is partly different in horses than in all of the others species, including humans, that have been tested in previous studies. A horse has no gallbladder. It means that bile is constantly passing from the liver directly to the intestine through bile duct. Pancreatic juice also constantly flows directly into the intestine. The production or release of gall and pancreatic juice in horses is not dependent on the food in the intestine or on the amount of lipids in feed like in other mammals [53]. If some active compounds from knotweed, such as piceatannol, could change the amount of bile acids in the gall, different amounts of lipids from food could be absorbed. Epicatechin-3-O-gallate from knotweed could increase the concentration of triglycerides in the blood by blocking lipase [48,54]. The horses in this study were fed a common diet without higher amounts of lipids and statistically significant results were within physiological ranges. We cannot say whether the triglyceride levels were increased due to the absorption from food or the release from adipose tissues. Answers to these questions are beyond the scope of this study. However, it shows an interesting potential of knotweed supplementation to support the breeding and performance of cold-blooded horses.
4.1.5. Cholesterol
Cholesterol is a fatty substance either obtained from food in the gut or produced in the liver when needed. It is a necessary component of all cell membranes and a fundamental component of many molecules, such as steroid hormones; it is also involved in the transport of lipids between different tissues in the body and the blood [47]. This study revealed more considerable reductions in the concentrations of blood cholesterol in the cold-blooded knotweed-fed horses. There are many studies describing different mechanisms by which resveratrol decreased the cholesterol concentration in blood, e.g., by reducing the basal and insulin-induced glucose conversion to total lipids in white adipose tissue [55]. There are also studies which found no effect of resveratrol on cholesterol concentration in horse blood [30]. Again, it seems that different active compounds are behind these diverse effects of knotweed. Another study [50] reported that a piceatannol-enriched diet decreased the cholesterol concentration in blood due to lowering its absorption in the intestine resulting from higher excretion of bile acids to the intestine. Piceid is also known for decreasing cholesterol concentrations [49].
All of the animals involved in the experiments reported here were healthy; no lipid metabolism disorders were noted. Nevertheless, their metabolisms were improved due to the reduction in cholesterol concentration. Knotweed, owing to its content of resveratrol possessing hepatoprotective effects, can help prevent steatosis, which is an abnormal retention of fat in an organ, usually the liver [56,57].
4.1.6. Urea
The urea measured in the blood of horses that had received knotweed in the previous winter and spring was higher than the urea in the blood of the control horses. All of these horses spent the extremely hot and dry summer between the two successive experiments on pastures and only fed on grass. It was reported [58] that changes in grass quality in the growth season and its protein (nitrogen) content correlate with blood urea level but do not affect the blood protein in horses on grass. The decreased grass quality resulted in lower blood urea content by up to half of its normal level [59]. In this study, the higher urea level in earlier knotweed-fed horses could indicate their better protein availability as all of the horses were on the same grass.
4.1.7. Urea, ALP, Triglycerides and Cholesterol Interactions
Urea was also positively correlated with ALP decreasing with horse weight, the decrease being enhanced by knotweed in Experiment 1. In Experiment 2, ALP was negatively correlated with cholesterol (r = −0.435, p = 0.03) in the control horses but not in the knotweed-fed horses, revealing the role of ALP in the metabolism of phosphorus associated with cholesterol presumably as phospholipids, i.e., amphiphatic molecules enabling the water-solubility of cholesterol. Similarly, the cholesterol and triglycerides showed a non-significant correlation in the horses fed knotweed compared to the horses not fed knotweed, where a highly significant correlation was found (p = 0.001).
When all of the age categories were evaluated together, the same knotweed effect was noted at the end of both experiments: the control horses not fed knotweed had a highly significant correlation between triglycerides and cholesterol (p = 0.01 in Experiment 1 and p = 0.001 in Experiment 2), but in the blood of all of the knotweed-fed horses, there was a non-significant correlation between triglycerides and cholesterol in either experiment. When only 1- and 2-year-old horses from Experiment 1 were taken into account, reappearing as 2- and 3-year-old horses in Experiment 2, a significant correlation was found only in the horses not fed knotweed before the start of Experiment 2. This reveals a strong influence on this ratio of 3-year-old horses, i.e., the heaviest ones.
These urea, ALP, cholesterol and triglyceride interrelations indicate the effect of knotweed on the protein and lipidic (lipoprotein, phospholipid) metabolism of horses. Several authors [59,60] have reported largely increased triglyceride content due to the seasonal food deprivation of only grass-fed equines. Others [61,62] have discerned three major classes of lipoproteins in equine plasma: very low density lipoproteins (VLDL, 24 per cent of the total plasma lipoprotein mass) rich in triglycerides and cholesterol and poor in protein (apolipoprotein B-100 and apoB-48), cholesterol-rich low density lipoproteins (LDL, 15 per cent) with three discrete subfractions, and protein-rich HDL with the dominant protein apoA-I (HDL, 61 per cent), all composed of lipids and apoliproteins in different proportions. Mass spectrometry revealed that the apolipoproteins contain etw. 400–500 amino acids. These might serve for protein synthesis as one of the means of lipoprotein utilisation.
Several papers [59,63] report that in horse serum (plasma), the cleavage of VLDL mainly releases triglycerides, while LDL largely releases cholesterol. In this study, the loss of correlation between triglycerides and cholesterol in knotweed-fed horses as compared with control ones could be explained if different types of lipoproteins were cleaved in knotweed-fed horses than in control horses not fed knotweed.
A similar loss of (negative) correlation in knotweed-fed horses, which was recorded between cholesterol and ALP, would suggest that, besides apolipoproteins, phospholipids as solubilising components and ALP targets are associated with the cholesterol particles.
The low quality of pasture grass later in the summer might be perceived as a period of mild fasting, producing higher blood triglyceride levels [59,60]. However, all of the changes measured in the blood plasma were within normal limits. Ref. [64] states that equine energy metabolism evolved in sparse grassland environments and has been adapted to the intake of a low energy diet with large quantities of roughage rich in cellulose and lignin, and that equids shift between the glucose-oriented metabolic pattern of non-ruminants and a metabolism similar to that of the ruminants when fed with a roughage-based diet. Since the blood glucose level is maintained within a narrow limit, preservation of its supply depends on gluconeogenesis, and protein catabolism increases to provide amin acids as precursors for glucose synthesis. However, more detailed studies are needed to reveal which metabolic processes are affected by knotweed in horse diets.
Meanwhile, we assume that even in earlier knotweed-fed horses, the higher urea levels indicating higher protein availability, together with a change in the ratio of lipidic substances, might have resulted from the long-lasting effects of knotweed on the metabolism of lipoproteins. Different types of lipoproteins would be metabolised with varying intensities, and the apolipoproteins released, together with other internal sources, might help horses overcome the periods of protein starvation on dry pastures lacking high quality grass.
5. Conclusions
The results obtained in this study indicate that knotweed consumption helps (1) improve the immunity of horses by increasing their blood globulin levels, (2) stimulate lipid metabolism in cold-blooded horses and (3) lower cholesterol levels. These effects have been found in young and healthy horses and are expected to be more pronounced in older horses.
Because knotweed has the potential to stimulate the release of fat from hepatic cells and thus speed up liver regeneration processes, it could reduce the risk of adverse health consequences in horses with impaired lipid metabolism.
The sporting success of the horses involved in the experiments was monitored. Half of the foals from both the control and knotweed-fed groups have been in use in sports in the Czech Republic since 2018, i.e., 2 years after the second experiment. The knotweed-fed horses achieved slightly better results than control horses in jumping competitions.
6. Patents
This study is a part of a more extensive project resulting in the registration of the Utility Model “Compound feed containing knotweed for game, domestic and farm animals”.
Author Contributions
Conceptualisation, P.M., M.K. and J.N.; methodology, M.K., P.M., T.F., Š.V. and J.N.; formal analysis, T.F.; investigation, M.K. and Š.V.; data curation, T.F.; writing—original draft preparation, M.K.; writing—review and editing, all authors; visualisation, T.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Technology Agency of the Czech Republic, grant number TH02010325 within project “Innovation of feed supplements for fitness improvement of domestic and wild animals”.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Institutional Ethics Committee of Mendel University in Brno, 21 October 2021. Feeding experiments are not applicable according to the law of the Czech Republic.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Data are available from the third author.
Acknowledgments
Our thanks are due to the managers of the Regional Stud Farm Tlumačov, Czech Republic, for excellent co-operation in both experiments. VUOS provided analyses of resveratrol in knotweed biomass. Graphic Abstract was prepared by BcA. Vít Kovář.
Conflicts of Interest
There are no conflict of interest.
References
- Bishayee, A. Cancer prevention and treatment with resveratrol: From rodent studies to clinical trials. Cancer Prev. Res. 2009, 2, 409–418. [Google Scholar] [CrossRef] [Green Version]
- Liu, A.; Chen, H.; Wei, W.; Ye, S.; Liao, W.; Gong, J.; Jiang, Z.; Wang, L.; Lin, S. Antiproliferative and antimetastatic effects of emodin on human pancreatic cancer. Oncol. Rep. 2011, 1257, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Kovářová, M.; Bartůňková, K.; Frantík, T.; Koblihová, H.; Prchalová, K.; Vosátka, M. Factors influencing the production of stilbenes by the knotweed, Reynoutria × bohemica. BMC Plant Biol. 2010, 10, 19. [Google Scholar] [CrossRef] [Green Version]
- Kovářová, M.; Bartůňková, K.; Frantík, T.; Koblihová, H.; Prchalová, K.; Vosátka, M. Effect of clone selection, nitrogen supply, leaf damage and mycorrhizal fungi on stilbene and emodin production in knotweed. BMC Plant Biol. 2011, 11, 98. [Google Scholar] [CrossRef] [Green Version]
- Kurita, S.; Kashiwagi, T.; Ebisu, T.; Shimamura, T.; Ukeda, H. Content of resveratrol and glycoside and its contribution to the antioxidative capacity of Polygonum cuspidatum (Itadori) harvested in Kochi. Biosci. Biotechnol. Biochem. 2014, 78, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Kurita, S.; Kashiwagi, T.; Ebisu, T.; Shimamura, T.; Ukeda, H. Identification of neochlorogenic acid as the predominant antioxidant in Polygonum cuspidatum leaves. Ital. J. Food Sci. 2016, 28, 25–31. [Google Scholar]
- Metličar, V.; Vovk, I.; Albreht, A. Japanese and Bohemian Knotweeds as Sustainable Sources of Carotenoids. Plants 2019, 8, 384. [Google Scholar] [CrossRef] [Green Version]
- Malaguarnera, L. Influence of Resveratrol on the Immune Response. Nutrients 2019, 11, 946. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Yu, C.-H.; Jiang, Y.-P.; Peng, C.; He, K.; Tang, J.-Y.; Xin, H.-L. Protective Effects of Polydatin from Polygonum cuspidatum against Carbon Tetrachloride-Induced Liver Injury in Mice. PLoS ONE 2012, 7, e46574. [Google Scholar] [CrossRef] [Green Version]
- Tan, Y.Y.; Chen, L.X.; Fang, L.; Zhang, Q. Cardioprotective effects of polydatin against myocardial injury in diabetic rats via inhibition of NADPH oxidase and NF-κB activities. BMC Complement. Med. Ther. 2020, 20, 378. [Google Scholar] [CrossRef] [PubMed]
- Zhan, J.; Li, X.; Luo, D.; Yan, W.; Hou, Y.; Hou, Y.; Chen, S.; Luan, J.; Zhang, Q.; Lin, D. Polydatin Attenuates OGD/R-Induced Neuronal Injury and Spinal Cord Ischemia/Reperfusion Injury by Protecting Mitochondrial Function via Nrf2/ARE Signaling Pathway. Oxid. Med. Cell. Longev. 2021, 2021, 6687212. [Google Scholar] [CrossRef]
- Gao, Y.; Chen, T.; Lei, X.; Li, Y.; Dai, X.; Cao, Y.; Ding, Q.; Lei, X.; Li, T.; Lin, X. Neuroprotective effects of polydatin against mitochondrial-dependent apoptosis in the rat cerebral cortex following ischemia/reperfusion injury. Mol. Med. Rep. 2016, 14, 5481–5488. [Google Scholar] [CrossRef] [Green Version]
- Fakhri, S.; Gravandi, M.M.; Abdian, S.; Akkol, E.K.; Farzaei, M.H.; Sobarzo-Sánchez, E. The Neuroprotective Role of Polydatin: Neuropharmacological Mechanisms, Molecular Targets, Therapeutic Potentials, and Clinical Perspective. Molecules 2021, 26, 5985. [Google Scholar] [CrossRef]
- Luce, A.; Lama, S.; Millan, P.C.; Itro, A.; Sangiovanni, A.; Caputo, C.; Ferranti, P.; Cappabianca, S.; Caraglia, M.; Stiuso, P. Polydatin Induces Differentiation and Radiation Sensitivity in Human Osteosarcoma Cells and Parallel Secretion through Lipid Metabolite Secretion. Oxid. Med. Cell. Longev. 2021, 2021, 3337013. [Google Scholar] [CrossRef]
- Zhou, Y.; Hou, Y.; Shen, J.; Huang, Y.; Martin, W.; Cheng, F. Network-based drug repurposing for novel coronavirus 2019-nCoV/SARS-CoV-2. Cell Discov. 2020, 6, 14. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Hou, Y.; Huang, J.; Pan, W.; Ma, Q.; Shi, Y.; Li, C.; Zhao, J.; Jia, Z.; Jiang, H.; et al. Lianhuaqingwen exerts anti-viral and anti-inflammatory activity against novel coronavirus (SARS-CoV-2). Pharmacol. Res. 2020, 156, 104761. [Google Scholar] [CrossRef]
- Jacob, C.; Kirsch, G.; Slusarenko, A.; Winyard, P.G.; Burkholz, T. Recent Advances in Redox Active Plant and Microbial Products: From Basic Chemistry to Widespread Applications in Medicine and Agriculture; Springer: New York, NY, USA, 2020. [Google Scholar]
- Park, S.; Lim, J.; Kim, J.R.; Cho, S. Inhibitory effects of resveratrol on hepatitis B virus X protein-induced hepatocellular carcinoma. J. Vet. Sci. 2017, 18, 419–429. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Tao, J.; Zhang, T.; Jiang, S.; Wei, W.; Han, H.; Shao, Y.; Zhou, G.; Yue, H. Resveratroloside alleviates postprandial hyperglycemia in diabetic mice by competitively inhibiting α-glucosidase. J. Agric. Food Chem. 2019, 67, 2886–2893. [Google Scholar] [CrossRef]
- Shan, B.; Cai, Y.Z.; Brooks, J.D.; Corke, H. Antibacterial properties of Polygonum cuspidatum roots and their major bioactive constituents. Food Chem. 2008, 109, 530–537. [Google Scholar] [CrossRef]
- Su, P.W.; Yang, C.H.; Yang, J.F.; Su, P.Y.; Chuang, L.Y. Antibacterial Activities and Antibacterial Mechanism of Polygonum cuspidatum Extracts against Nosocomial Drug-Resistant Pathogens. Molecules 2015, 20, 11119–11130. [Google Scholar] [CrossRef] [Green Version]
- Franck, T.; Kohnen, G.; Deby-Dupont, S.; Grulke, C.; Serteyn, D.D. A specific method for measurement of equine active myeloperoxidase inbiological samples and in in vitro tests. J. Vet. Diagn. Investig. 2006, 18, 326–334. [Google Scholar] [CrossRef] [Green Version]
- Klebanoff, S.J. Myeloperoxidase: Friend and foe. J. Leukoc. Biol. 2005, 77, 598–625. [Google Scholar] [CrossRef]
- Cavallaro, A.; Ainis, T.; Bottari, C.; Fimiani, V. Effect of resveratrol on some activities of isolated and in whole blood human neutrophils. Physiol. Res. 2003, 52, 555–562. [Google Scholar]
- Kohnen, S.; Franck, T.; Van Antwerpen, P.; Boudjeltia Zouaoui, K.; Mouithys-Mickalad, A.; Deby, C.; Moguilevsky, N.; Deby, G.; Lamy, N.; Serteyn, D. Resveratrol inhibits the activity of equine neutrophil myeloperoxidase by a direct interaction with the enzyme. J. Agric. Food Chem. 2007, 55, 8080–8087. [Google Scholar] [CrossRef] [PubMed]
- Zambito, J.L. Effects of Resveratrol Supplementation on Glycemic Response and Oxidant Status in Moderately Exercised Mature Quarter Horse Geldings. Master’s Thesis, West Virginia University, Morgantown, WV, USA, 2011; 96p. Available online: https://researchrepository.wvu.edu/cgi/viewcontent.cgi?article=4337&context=etd (accessed on 11 September 2021).
- Lawless, P. Two New Research Projects Study on Resveratrol’s Effect on EMS. Biological Prospects/Equithrive. 2010. Available online: https://horsenetwork.com/2017/02/resverasyn-can-help-horse/ (accessed on 13 October 2021).
- Lawless, P. What is Resverasyn and How Can It Help My Horse. Available online: https://equithrive.com/blogs/news/what-is-resverasyn-and-how-can-it-help-my-horse?_pos=2&_sid=22ad88df5&_ss=r (accessed on 13 October 2021).
- Lawless, P.; Equithrive. Resverasyn, the Key Ingredient. Available online: https://equithrive.com/pages/resverasyn (accessed on 13 October 2021).
- Ememe, M.U.; Abdullahi, U.S.; Sackey, A.K.; Ayo, J.O.; Mshelia, W.P.; Edeh, R.E. Effects of a joint supplement whose main components are resveratrol and hyaluronic acid on some biochemical parameters in aged lame horses. J. Equine Sci. 2016, 27, 19–22. [Google Scholar] [CrossRef] [PubMed]
- Ememe, M.U.; Msheliaa, W.P.; Ayo, J.O. Ameliorative Effects of Resveratrol on Oxidative Stress Biomarkers in Horses. J. Equine Vet. Sci. 2015, 35, 518–523. [Google Scholar] [CrossRef]
- Watts, A.E.; Dabareiner, R.; Marsh, C.; Carter, K.; Cummings, K.J. A randomized, controlled trial of the effects of resveratrol administration in performance horses with lameness localized to the distal tarsal joints. J. Am. Vet. Med. Assoc. 2016, 249, 650–659. [Google Scholar] [CrossRef]
- Siard, M.H.; McMurry, K.E.; Adams, A.A. Effects of polyphenols including curcuminoids, resveratrol, quercetin, pterostilbene, and hydroxypterostilbene on lymphocyte pro-inflammatory cytokine production of senior horses in vitro. Vet. Immunol. Immunopathol. 2016, 173, 50–59. [Google Scholar] [CrossRef]
- Kalughina, E.G.; Stolbova, O.A. Praziver® and Ivermek® effectiveness for horse helminthiase prevention. Eurasian J. Biosci. 2020, 14, 317–322. [Google Scholar]
- Ter Braak, C.J.F.; Šmilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination, Version 5.0; Microcomputer Power: Ithaca, NY, USA, 2012; p. 362. [Google Scholar]
- Werner, L.L.; Turnwald, G.H.; Willard, M.D. Immunologic and Plasma Protein Disorders. Small Anim. Clin. Diagn. Lab. Methods 2004, 290–305. [Google Scholar] [CrossRef]
- Gounden, V.; Vashisht, R.; Jialal, I. Hypoalbuminemia; StatPearls Publishing LLC: Treasure Island, FL, USA, 2021. [Google Scholar]
- Tothova, C.; Nagy, O.; Kovac, G. Serum proteins and their diagnostic utility in veterinary medicine: A review. Vet. Med. 2016, 61, 475–496. [Google Scholar] [CrossRef] [Green Version]
- Harvey, J.W. Veterinary Hematology II Evaluation of Hemostasis; Elsevier: St. Louis, MO, USA, 2012; pp. 191–233. [Google Scholar] [CrossRef]
- Maděra, P.; Kovářová, M.; Frantík, T.; Filipčík, R.; Novák, J.; Vencl, Š.; Maděrová, L.; Rozkot, M.; Kuchařová, S.; Václavková, E.; et al. Effect of knotweed in diet on physiological changes in pig. Agriculture 2021, 11, 169. [Google Scholar] [CrossRef]
- Ferry-Dumazet, H.; Garnier, O.; Mamani-Matsuda, M.; Vercauteren, J.; Belloc, F.; Billiard, C.; Dupouy, M.; Thiolat, D.; Kolb, J.P.; Marit, G. Resveratrol inhibits the growth and induces the apoptosis of both normal and leukemic hematopoietic cells. Carconigenesis 2002, 23, 1323–1333. [Google Scholar] [CrossRef]
- Baker, D. Diagnosis of disorders of hemostasis. In Veterinary Hematology and Clinical Chemistry; Thrall, M., Baker, D., Campbell, T., Eds.; Blackwell Publishing: Ames, IA, USA, 2006; pp. 179–196. [Google Scholar]
- Sharma, S.; Parashar, P.; Sharma, S.; Sharma, K.P. Ameliorating role of lycopene, tomato puree, and Spirulina + tomato puree on the hematology of fluoride-exposed Swiss Albino mice. J. Diet. Suppl. 2018, 15, 827–841. [Google Scholar] [CrossRef]
- Khan, S.; Ullah, R.; Saleem, M.; Bilal, M.; Rashid, R.; Khan, I.; Mahmood, A.; Nawaz, M. Raman spectroscopic analysis of dengue virus infection in human blood sera. Optik 2016, 127, 2086–2088. [Google Scholar] [CrossRef]
- Szulc, P.; Bauer, D.C. Biochemical markers of bone turnover in osteoporosis. In Osteoporosis, 4th ed.; Marcus, R., Feldman, D., Dempster, D.W., Luckey, M., Cauley, J.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 1573–1610. [Google Scholar] [CrossRef]
- Kraft, W.; Dürr, U.M. Referenzwerte in Klinische Labordiagnostik der Tiermedizin. In Klinische Labordiagnostik in der Tiermedizin, 6th ed.; Kraft, W., Ed.; Schattauer: Stuttgart, Germany, 2005; pp. 339–357. [Google Scholar]
- Lassen, E.; Fettman, M. Laboratory evaluation of lipids. In Veterinary Hematology and Clinical Chemistry; Thrall, M.A., Weiser, G., Allison, R.W., Campbell, T.W., Eds.; Willey-Blackwell: Hoboken, NJ, USA, 2006; pp. 421–429. [Google Scholar]
- Nawrot-Hadzik, I.; Hadzik, J.; Fleischer, M.; Choromańska, A.; Sterczała, B.; Kubasiewicz-Ross, P.; Saczko, J.; Gałczyńska-Rusin, M.; Gedrange, T.; Matkowski, A. Chemical Composition of East Asian Invasive Knotweeds, their Cytotoxicity and Antimicrobial Efficacy Against Cariogenic Pathogens: An In-Vitro Study. Med. Sci. Monit. 2019, 25, 3279–3287. [Google Scholar] [CrossRef] [PubMed]
- Arichi, H.; Kimura, Y.; Okuda, H.; Baba, K.; Kozawa, M.; Arichi, S. Effects of Stilbene Components of the roots of Polygonum cuspidatum Sieb. et Zucc. on Lipid Metabolism. Chem. Pharm. Bull. 1982, 30, 1766–1770. [Google Scholar] [CrossRef] [Green Version]
- Tung-Ting, S.; Meng-Heng, L.; Chi-On, C.; Huan, Z.; Shun-Wan, C.; Kam-Wah Mok, D. Cholesterol-lowering effects of piceatannol, a stilbene from wine, using untargeted metabolomics. J. Funct. Foods 2017, 28, 127–137. [Google Scholar] [CrossRef]
- Du, J.; Sun, L.N.; Xing, W.W.; Huang, B.K.; Jia, M.; Wu, J.Z.; Zhang, H.; Qin, L.P. Lipid-lowering effects of polydatin from Polygonum cuspidatum in hyperlipidemic hamsters. Phytomedicine 2009, 16, 652–658. [Google Scholar] [CrossRef]
- Patocka, J.; Navratilova, Z.; Ovando, M. Biologically Active Compounds of Knotweed (Reynoutria spp.). Mil. Med. Sci. Lett. 2017, 86, 17–31. [Google Scholar] [CrossRef] [Green Version]
- Reece, W.O. Fyziologie A Funkční Anatomie Domácích Zvířat, 2nd ed.; Grada Publishing: Prague, Czech Republic, 2011; pp. 313–360. [Google Scholar]
- Kusano, R.; Andou, H.; Fujieda, M.; Tanaka, T.; Matsuo, Y.; Kouno, I. Polymer-like Polyphenols of Black Tea and Their Lipase and Amylase Inhibitory Activities. Chem. Pharm. Bull. 2008, 56, 266–272. [Google Scholar] [CrossRef] [Green Version]
- Szkudelska, K.; Szkudelski, T. Resveratrol, obesity and diabetes. Eur. J. Pharmacol. 2010, 635, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chiang-Wen, L.; Feng-Lin, Y.; Haw-Wei, H.; Tzu-Hui, W.; Horng-Huey, K.; Wen-Sheng, T.; Chun-Ching, L. Resveratrol Nanoparticle System Improves Dissolution Properties and Enhances the Hepatoprotective Effect of Resveratrol through Antioxidant and Anti-Inflammatory Pathways. J. Agric. Food Chem. 2012, 60, 4662–4671. [Google Scholar] [CrossRef]
- Xu, L.; Huang, G.; Guo, X.; Zhou, Q.; He, S. Total flavonoids extracted from Polygonum knotweed L, exerts beneficial hepatoprotection against liver injury. J. Cell. Biochem. 2019, 120, 12677–12683. [Google Scholar] [CrossRef] [PubMed]
- Fonnesbeck, P.V.; Linda, D.; Symons, L.U. Effect of Diet on Concentration of Protein, Urea Nitrogen, Sugar and Cholesterol of Blood Plasma of Horses. J. Anim. Sci. 1969, 28, 216–219. [Google Scholar] [CrossRef]
- Owen, J.M.; McCullagh, K.G.; Crook, D.H.; Hinton, M. Seasonal Variations in the Nutrition of Horses at Grass. Equine Vet. J. 1978, 10, 260–266. [Google Scholar] [CrossRef]
- Morris, D.M.; Zilversmit, B.D.; Hintz, F.H. Hyperlipoproteinaemia in fasting ponies. J. Lipid Res. 1972, 13, 383–389. [Google Scholar] [CrossRef]
- Watson, T.D.; Burns, L.; Love, S.; Packard, C.J.; Shepherd, J. The isolation, characterisation and quantification of the equine plasma lipoproteins. Equine Vet. J. 1991, 23, 353–359. [Google Scholar] [CrossRef]
- Watson, T.D.; Packard, C.J.; Shepherd, J. Plasma lipid transport in the horse (Equus caballus). Comp. Biochem. Physiol. B 1993, 106, 27–34. [Google Scholar] [CrossRef]
- Wensing, T.H.; Van Gent, C.M.; Schotman, A.J.H.; Kroneman, J. Hyperlipoproteinaemia in ponies: Mechanisms and response to therapy. Clin. Chim. Acta 1975, 58, 1–15. [Google Scholar] [CrossRef] [Green Version]
- McKenzie, H.C., III. Equine hyperlipidemias. Vet. Clin. Equine Pract. 2011, 27, 59–72. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
(a) Diagram showing the relationship of the biochemical data in Experiment 1. Treatment explained 1% of the variability in the biochemical data. Vectors pointing in the same direction show characteristics with positive correlations; arrows heading towards a type indicate that this type positively affects these characteristics. (b) Diagram showing the relatedness of the haematological data of Experiment 1. Treatment explained 1.5% of the variability in the haematological data. Vectors pointing in the same direction show characteristics with positive correlations; arrows heading towards a type indicate that this type positively affects these characteristics.
Figure 1.
(a) Diagram showing the relationship of the biochemical data in Experiment 1. Treatment explained 1% of the variability in the biochemical data. Vectors pointing in the same direction show characteristics with positive correlations; arrows heading towards a type indicate that this type positively affects these characteristics. (b) Diagram showing the relatedness of the haematological data of Experiment 1. Treatment explained 1.5% of the variability in the haematological data. Vectors pointing in the same direction show characteristics with positive correlations; arrows heading towards a type indicate that this type positively affects these characteristics.

Figure 2.
Differences in the total protein content in equine blood between the beginning and the end of Experiment 2, shown for stallions of the three age groups (born in 2016, 2017 and 2018) fed and not fed knotweed. Means ± SEM. Statistically different values at p ≤ 0.05 are marked with *. N.S. = non-significant difference.
Figure 2.
Differences in the total protein content in equine blood between the beginning and the end of Experiment 2, shown for stallions of the three age groups (born in 2016, 2017 and 2018) fed and not fed knotweed. Means ± SEM. Statistically different values at p ≤ 0.05 are marked with *. N.S. = non-significant difference.

Figure 3.
Globulin content in the blood of stallions of all ages. The comparison was between warm-blooded and cold-blooded stallions fed with and without knotweed, Experiment 1. Means ± SEM. Statistically different values at p ≤ 0.05 are marked with *; N.S. = non-significant difference.
Figure 3.
Globulin content in the blood of stallions of all ages. The comparison was between warm-blooded and cold-blooded stallions fed with and without knotweed, Experiment 1. Means ± SEM. Statistically different values at p ≤ 0.05 are marked with *; N.S. = non-significant difference.
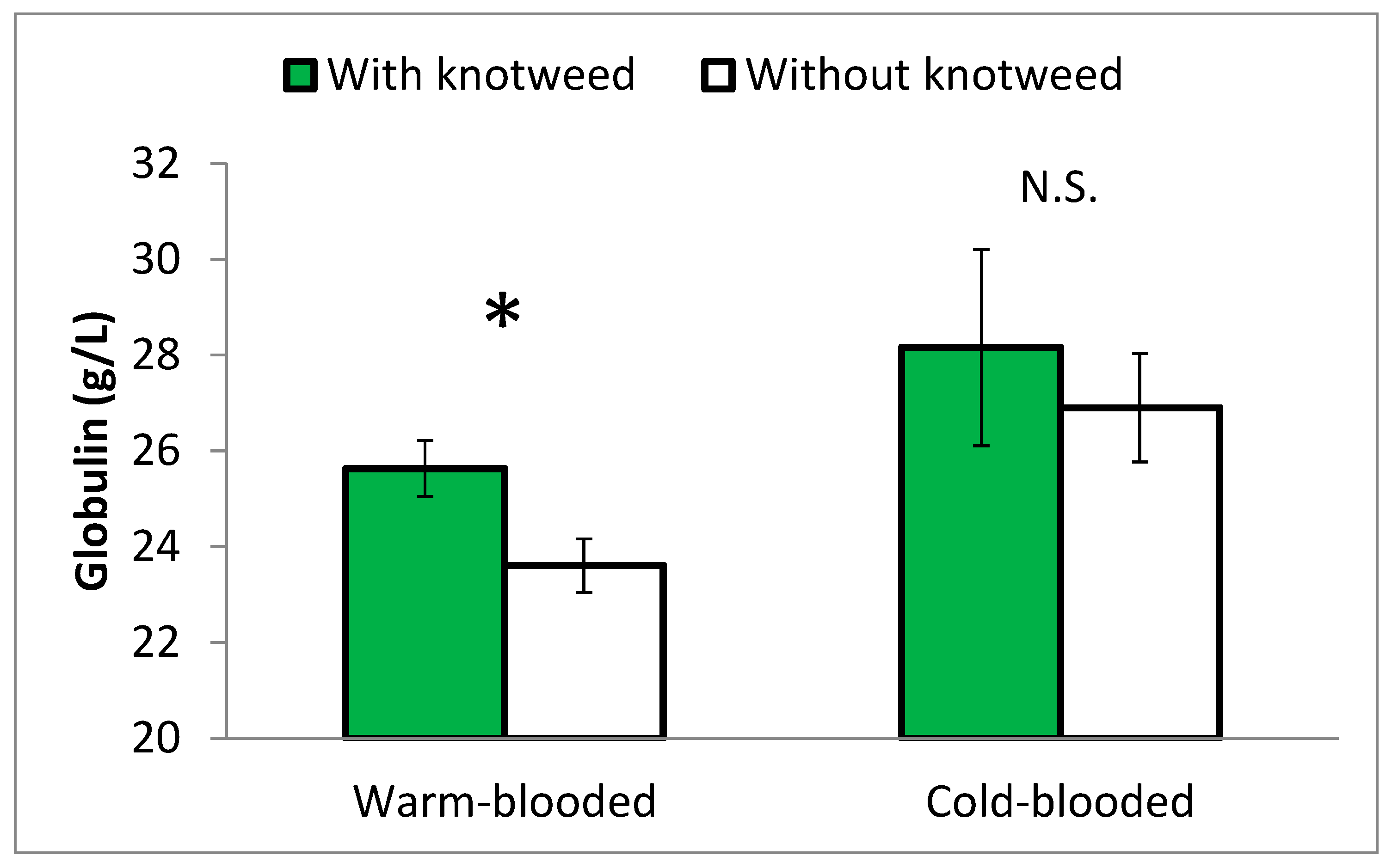
Figure 4.
Albumin/globulin ratio in the blood of warm-blooded and cold-blooded stallions fed with and without knotweed, Experiment 1. Means ± SEM. Statistically different values at p ≤ 0.05 are marked with *; N.S. = non-significant difference.
Figure 4.
Albumin/globulin ratio in the blood of warm-blooded and cold-blooded stallions fed with and without knotweed, Experiment 1. Means ± SEM. Statistically different values at p ≤ 0.05 are marked with *; N.S. = non-significant difference.

Figure 5.
Differences in equine ALP activity between the beginning and the end of Experiment 1 among horses fed and not fed knotweed. The warm-blooded and cold-blooded breeds were compared. Statistically different values at p ≤ 0.05 are marked with *. N.S. = non-significant difference.
Figure 5.
Differences in equine ALP activity between the beginning and the end of Experiment 1 among horses fed and not fed knotweed. The warm-blooded and cold-blooded breeds were compared. Statistically different values at p ≤ 0.05 are marked with *. N.S. = non-significant difference.
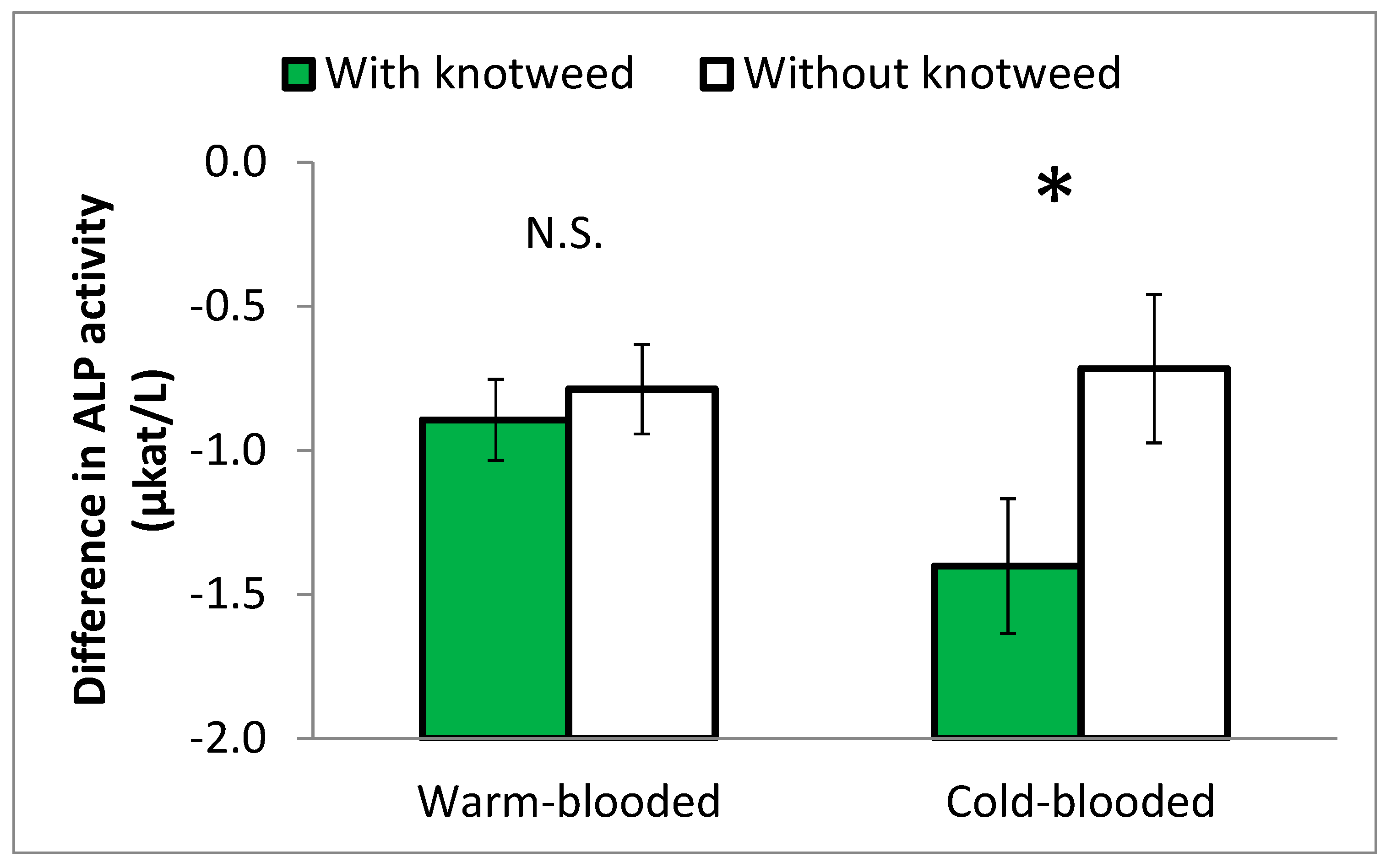
Figure 6.
Levels of triglycerides at the end of Experiment 2 in the blood of horses fed and not fed knotweed. Warm-blooded and cold-blooded horses were compared. Statistically different values at p ≤ 0.05 are marked with *. N.S. = non-significant difference.
Figure 6.
Levels of triglycerides at the end of Experiment 2 in the blood of horses fed and not fed knotweed. Warm-blooded and cold-blooded horses were compared. Statistically different values at p ≤ 0.05 are marked with *. N.S. = non-significant difference.
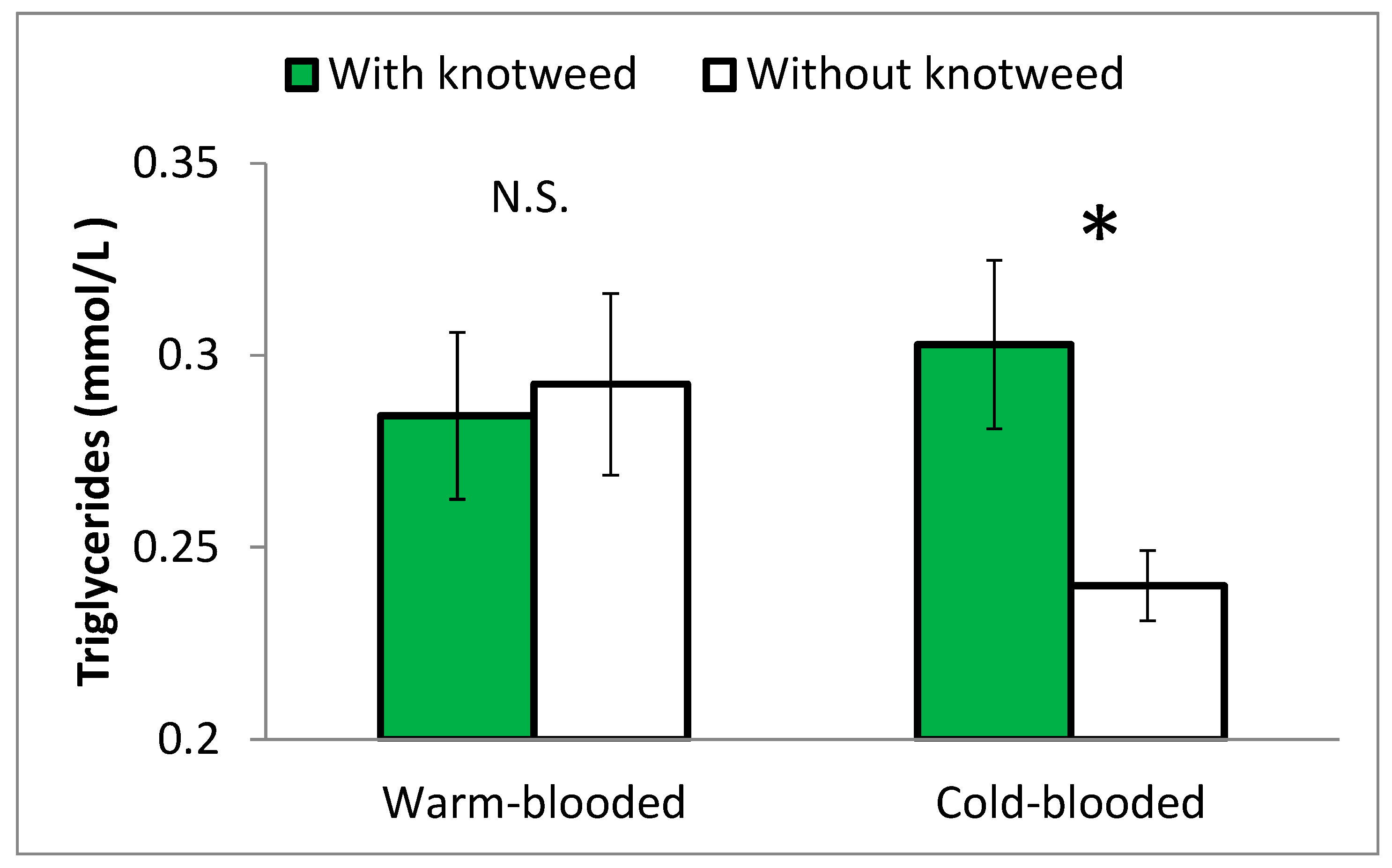
Figure 7.
The differences in the blood cholesterol levels of 2- and 3-year-old stallions fed with and without knotweed from the beginning to the end of Experiment 2, comparing warm-blooded and cold-blooded. Statistically different values at p ≤ 0.05 are marked with *. N.S. = non-significant values.
Figure 7.
The differences in the blood cholesterol levels of 2- and 3-year-old stallions fed with and without knotweed from the beginning to the end of Experiment 2, comparing warm-blooded and cold-blooded. Statistically different values at p ≤ 0.05 are marked with *. N.S. = non-significant values.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Amounts, composition and nutrient contents of the feed for 3-year-old warmblood horses.
| Unit | Grass Hay | Feed Supplement | Experimental Mixture for Control Horses | Experimental Mixture for Knotweed-Fed Horses | Salt | Control Horses, Total | Knotweed Fed Horses, Total | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Oat Grain | Supplementary Feed | KnotWeed | Rest of Mixture | |||||||
| INTAKE | kg/day | 8.00 | 0.25 | 2.50 | 0.50 | 0.15 | 0.35 | 0.03 | 11.28 | 11.28 |
| ENERGY | MJ/day | 56.90 | 3.10 | 28.20 | 6.50 | 1.18 | 4.63 | 0.00 | 94.70 | 94.01 |
| Protein | g/day | 752.00 | 29.50 | 432.50 | 51.50 | 18.97 | 35.6 | 0.00 | 1265.50 | 1268.57 |
| Lysine | g/day | 24.00 | 1.00 | 17.50 | 1.70 | 0.73 | 1.19 | 0.00 | 44.20 | 44.42 |
| Calcium | g/day | 26.40 | 0.20 | 22.50 | 0.22 | 1.2 | 0.15 | 0.00 | 49.32 | 50.45 |
| Phosphorus | g/day | 25.60 | 0.80 | 15.00 | 1.97 | 0.37 | 1.37 | 0.00 | 43.37 | 43.14 |
| Magnesium | g/day | 12.80 | 0.30 | 5.00 | 0.42 | 0.39 | 0.29 | 0.00 | 18.52 | 18.78 |
| Sodium | g/day | 1.60 | 0.10 | 4.00 | 0.04 | 0.07 | 0.03 | 10.00 | 15.74 | 15.80 |
| Selenium | mg/day | 0.80 | 0.05 | 0.71 | 0.01 | 0.01 | 0.01 | 0.00 | 1.57 | 1.58 |
| Copper | mg/day | 56.00 | 1.50 | 18.30 | 1.78 | 0.77 | 1.25 | 0.00 | 77.58 | 77.82 |
| Zinc | mg/day | 206.40 | 8.80 | 255.00 | 10.73 | 5.86 | 7.46 | 0.00 | 480.93 | 483.52 |
Table 2.
Biochemical and haematological characteristics of horse blood from the end of Experiment 1 and of Experiment 2. Mean ± SEM. Significant differences between control and knotweed-fed horses are marked by different letters in superscript.
Table 2.
Biochemical and haematological characteristics of horse blood from the end of Experiment 1 and of Experiment 2. Mean ± SEM. Significant differences between control and knotweed-fed horses are marked by different letters in superscript.
| Experiment 1 | Experiment 2 | ||||
|---|---|---|---|---|---|
| Biochemical Characteristic | Unit | Control Horses | Knotweed-Fed Horses | Control Horses | Knotweed-Fed Horses |
| Total protein | g/L | 60.696 ± 0.581 | 61.787 ± 0.710 | 61.281 ± 0.754 | 60.743 ± 1.274 |
| Albumin | g/L | 36.083 ± 0.313 | 35.405 ± 0.377 | 34.452 ± 0.668 | 33.823 ± 0.605 |
| Globulin | g/L | 24.612 ± 0.573 | 26.381 ± 0.744 | 26.829 ± 0.703 | 26.920 ± 1.258 |
| Urea | mmol/L | 5.500 ± 0.141 | 5.324 ± 0.169 | 5.171 ± 0.162 | 5.537 ± 0.279 |
| Creatinine | µmol/L | 110.482 ± 1.981 | 108.787 ± 2.264 | 87.484 ± 1.734 | 88.467 ± 1.718 |
| Cholesterol | mmol/L | 2.307 ± 0.045 | 2.307 ± 0.054 | 2.061 ± 0.053 | 2.110 ± 0.056 |
| Glucose | mmol/L | 6.344 ± 0.192 | 6.399 ± 0.226 | 5.842 ± 0.207 | 5.804 ± 0.182 |
| Triglyceride | mmol/L | 0.328 ± 0.019 | 0.337 ± 0.018 | 0.274 ± 0.016 | 0.291 ± 0.016 |
| AST activity | µkat/L | 5.168 ± 0.062 | 5.053 ± 0.092 | 5.755 ± 0.189 | 5.703 ± 0.319 |
| ALT activity | µkat/L | 0.213 ± 0.006 | 0.215 ± 0.011 | 0.202 ± 0.010 | 0.210 ± 0.009 |
| ALP activity | µkat/L | 3.042 ± 0.148 | 3.077 ± 0.153 | 3.575 ± 0.219 | 3.774 ± 0.292 |
| Haematological Characteristic | Unit | Control Horses | Knotweed-Fed Horses | Control Horses | Knotweed-Fed Horses |
| Haemoglobin | g/L | 108.500 ± 2.015 | 108.649 ± 2.178 | 113.194 ± 2.940 | 111.233 ± 2.721 |
| PCT | % | 0.084 ± 0.003 b | 0.100 ± 0.004 a | ||
| MCH | pg | 13.150 ± 0.139 | 13.095 ± 0.148 | 14.074 ± 0.117 | 13.910 ± 0.148 |
| Erythrocyte | 1012/L | 10.471 ± 2.278 | 8.267 ± 0.124 | 8.001 ± 0.173 | 7.950 ± 0.152 |
| Thrombocyte | 109/L | 161.839 ± 5.665 b | 189.367 ± 7.769 a | ||
| Leucocyte | 109/L | 8.228 ± 0.227 | 8.749 ± 0.293 | 8.781 ± 0.319 | 9.047 ± 0.351 |
| Lymphocyte | 109/L | 4.604 ± 0.188 | 4.680 ± 0.186 | 5.591 ± 0.286 | 5.756 ± 0.317 |
| Neutrophil | 109/L | 3.362 ± 0.130 | 3.771 ± 0.223 | 2.830 ± 0.261 | 2.871 ± 0.223 |
| Monocyte | 109/L | 0.028 ± 0.007 b | 0.055 ± 0.007 a | 0.187 ± 0.022 | 0.157 ± 0.023 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kovářová, M.; Maděra, P.; Frantík, T.; Novák, J.; Vencl, Š. Effects of Knotweed-Enriched Feed on the Blood Characteristics and Fitness of Horses. Agriculture 2022, 12, 109. https://doi.org/10.3390/agriculture12010109
AMA Style
Kovářová M, Maděra P, Frantík T, Novák J, Vencl Š. Effects of Knotweed-Enriched Feed on the Blood Characteristics and Fitness of Horses. Agriculture. 2022; 12(1):109. https://doi.org/10.3390/agriculture12010109
Chicago/Turabian StyleKovářová, Marcela, Petr Maděra, Tomáš Frantík, Jan Novák, and Štěpán Vencl. 2022. "Effects of Knotweed-Enriched Feed on the Blood Characteristics and Fitness of Horses" Agriculture 12, no. 1: 109. https://doi.org/10.3390/agriculture12010109
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.