Common Variants in 22 Genes Regulate Response to Metformin Intervention in Children with Obesity: A Pharmacogenetic Study of a Randomized Controlled Trial

 , ,
, ,  ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Experimental Section
2.1. Study Design, Participants, and Intervention
2.2. Informed Consent and Ethics
2.3. Blood Samples Collection
2.4. Anthropometric and Biochemical Measurements
2.5. DNA Extraction and Genotyping
Candidate Gene and SNP Selection
2.6. Statistical Analysis
3. Results
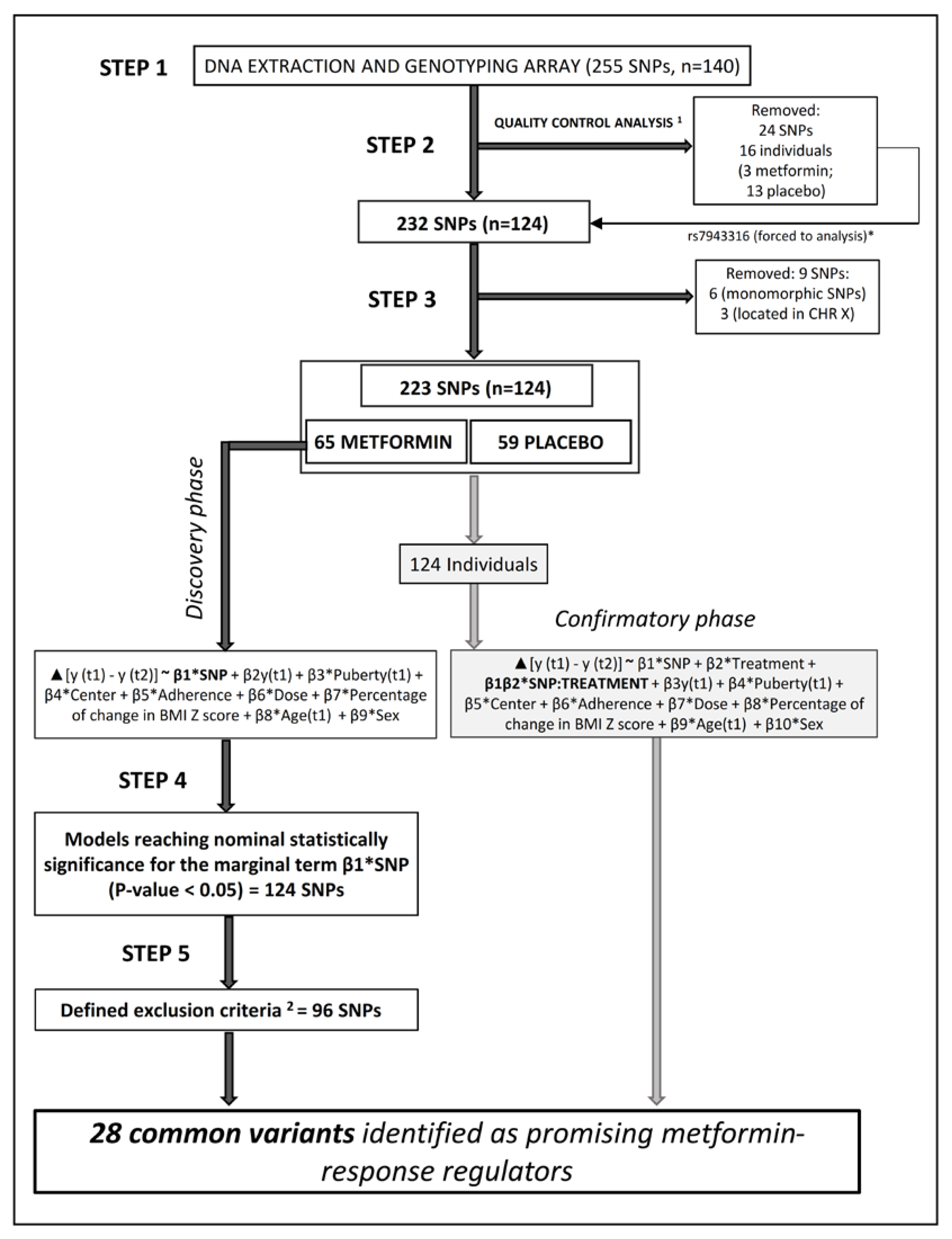
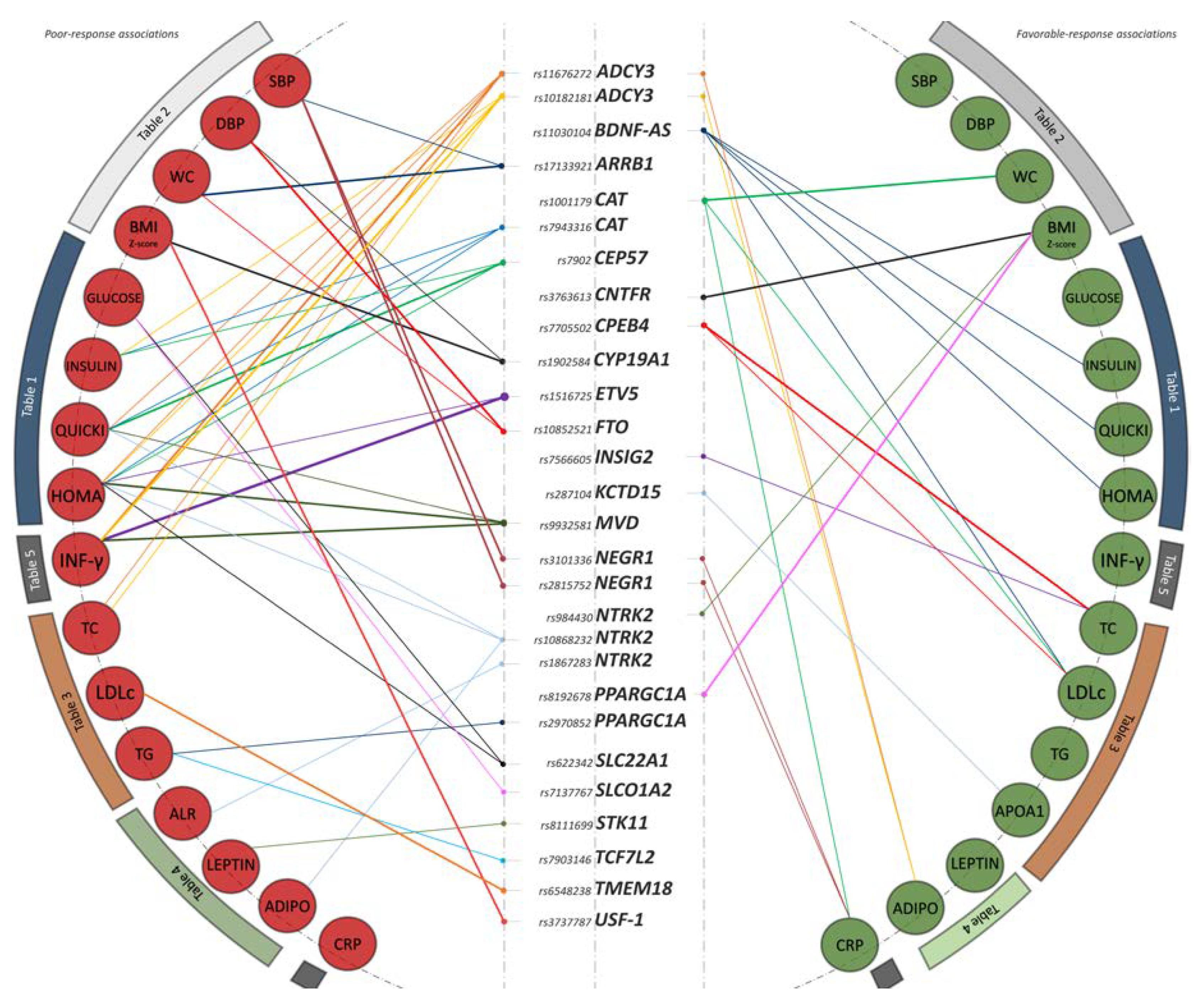
3.1. Identification of 28 Common Variants as Promising Metformin Pharmacogenetic Markers
3.2. Glucose Metabolism
3.3. Anthropometry and Blood Pressure
3.4. Lipid Metabolism
3.5. Adipokines and Inflammatory Biomarkers
3.6. Confirmatory Phase
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- GBD 2015 Obesity Collaborators; Afshin, A.; Forouzanfar, M.H.; Reitsma, M.B.; Sur, P.; Estep, K.; Lee, A.; Marczak, L.; Mokdad, A.H.; Moradi-Lakeh, M.; et al. Health Effects of Overweight and Obesity in 195 Countries over 25 Years. N. Engl. J. Med. 2017, 377, 13–27. [Google Scholar] [PubMed]
- Olza, J.; Gil-Campos, M.; Leis, R.; Bueno, G.; Aguilera, C.M.; Valle, M.; Cañete, R.; Tojo, R.; Moreno, L.A.; Gil, A. Presence of the metabolic syndrome in obese children at prepubertal age. Ann. Nutr. Metab. 2011, 58, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Knowler, W.C.; Barrett-Connor, E.; Fowler, S.E.; Hamman, R.F.; Lachin, J.M.; Walker, E.A.; Nathan, D.M.; Diabetes Prevention Program Research Group. Reduction in the incidence of type 2 diabetes with lifestyle intervention or metformin. N. Engl. J. Med. 2002, 346, 393–403. [Google Scholar] [PubMed]
- Mead, E.; Atkinson, G.; Richter, B.; Metzendorf, M.-I.; Baur, L.; Finer, N.; Corpeleijn, E.; O’Malley, C.; Ells, L.J. Drug interventions for the treatment of obesity in children and adolescents. Cochrane Database Syst. Rev. 2016, 11, CD012436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastor-Villaescusa, B.; Cañete, M.D.; Caballero-Villarraso, J.; Hoyos, R.; Latorre, M.; Vázquez-Cobela, R.; Plaza-Díaz, J.; Maldonado, J.; Bueno, G.; Leis, R.; et al. Metformin for Obesity in Prepubertal and Pubertal Children: A Randomized Controlled Trial. Pediatrics 2017, 140, e20164285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warnakulasuriya, L.S.; Fernando, M.M.A.; Adikaram, A.V.N.; Thawfeek, A.R.M.; Anurasiri, W.-M.L.; Silva, R.R.; Sirasa, M.S.F.; Rytter, E.; Forslund, A.H.; Samaranayake, D.L.; et al. Metformin in the Management of Childhood Obesity: A Randomized Control Trial. Child. Obes. 2018, 14, 553–565. [Google Scholar] [CrossRef] [PubMed]
- Zhou, K.; Donnelly, L.; Yang, J.; Li, M.; Deshmukh, H.; Van Zuydam, N.; Ahlqvist, E.; Spencer, C.C.; Groop, L.; Morris, A.D.; et al. Heritability of variation in glycaemic response to metformin: A genome-wide complex trait analysis. Lancet Diabetes Endocrinol. 2014, 2, 481–487. [Google Scholar] [CrossRef]
- Florez, J.C. The pharmacogenetics of metformin. Diabetologia 2017, 60, 1648–1655. [Google Scholar] [CrossRef] [Green Version]
- Sam, W.J.; Roza, O.; Hon, Y.Y.; Alfaro, R.M.; Calis, K.A.; Reynolds, J.C.; Yanovski, J.A. Effects of SLC22A1 Polymorphisms on Metformin-Induced Reductions in Adiposity and Metformin Pharmacokinetics in Obese Children with Insulin Resistance. J. Clin. Pharmacol. 2017, 57, 219–229. [Google Scholar] [CrossRef]
- van Rongen, A.; van der Aa, M.P.; Matic, M.; van Schaik, R.H.N.; Deneer, V.H.M.; van der Vorst, M.M.; Knibbe, C.A.J. Increased Metformin Clearance in Overweight and Obese Adolescents: A Pharmacokinetic Substudy of a Randomized Controlled Trial. Pediatr. Drugs 2018, 20, 365–374. [Google Scholar] [CrossRef] [Green Version]
- Goodarzi, M.O. Genetics of obesity: What genetic association studies have taught us about the biology of obesity and its complications. Lancet Diabetes Endocrinol. 2018, 6, 223–236. [Google Scholar] [CrossRef]
- Tanner, J.M.; Whitehouse, R.H. Clinical longitudinal standards for height, weight, height velocity, weight velocity, and stages of puberty. Arch. Dis. Child. 1976, 51, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Cole, T.J.; Bellizzi, M.C.; Flegal, K.M.; Dietz, W.H. Establishing a standard definition for child overweight and obesity worldwide: International survey. BMJ 2000, 320, 1240–1243. [Google Scholar] [CrossRef] [PubMed]
- Pastor-Villaescusa, B.; Caballero-Villarraso, J.; Cañete, M.D.; Hoyos, R.; Maldonado, J.; Bueno, G.; Leis, R.; Gil, Á.; Cañete, R.; Aguilera, C.M. Evaluation of differential effects of metformin treatment in obese children according to pubertal stage and genetic variations: Study protocol for a randomized controlled trial. Trials 2016, 17, 323. [Google Scholar] [CrossRef]
- Olza, J.; Gil-Campos, M.; Leis, R.; Rupérez, A.I.; Tojo, R.; Cañete, R.; Gil, A.; Aguilera, C.M. A gene variant of 11β-hydroxysteroid dehydrogenase type 1 is associated with obesity in children. Int. J. Obes. 2012, 36, 1558–1563. [Google Scholar] [CrossRef] [PubMed]
- Olza, J.; Gil-Campos, M.; Leis, R.; Rupérez, A.I.; Tojo, R.; Cañete, R.; Gil, Á.; Aguilera, C.M. Influence of variants in the NPY gene on obesity and metabolic syndrome features in Spanish children. Peptides 2013, 45, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Rupérez, A.I.; Olza, J.; Gil-Campos, M.; Leis, R.; Mesa, M.D.; Tojo, R.; Cañete, R.; Gil, Á.; Aguilera, C.M. Association of Genetic Polymorphisms for Glutathione Peroxidase Genes with Obesity in Spanish Children. J. Nutrigenet. Nutri. 2014, 7, 130–142. [Google Scholar] [CrossRef]
- Olza, J.; Rupérez, A.; Gil-Campos, M.; Leis, R.; Cañete, R.; Tojo, R.; Gil, Á.; Aguilera, C. Leptin Receptor Gene Variant rs11804091 Is Associated with BMI and Insulin Resistance in Spanish Female Obese Children: A Case-Control Study. Int. J. Mol. Sci. 2017, 18, 1690. [Google Scholar] [CrossRef]
- Rupérez, A.I.; Olza, J.; Gil-Campos, M.; Leis, R.; Mesa, M.D.; Tojo, R.; Cañete, R.; Gil, Á.; Aguilera, C.M. Are Catalase −844A/G Polymorphism and Activity Associated with Childhood Obesity? Antioxid. Redox Signal. 2013, 19, 1970–1975. [Google Scholar] [CrossRef]
- Aguilera, C.M.; Gomez-Llorente, C.; Tofe, I.; Gil-Campos, M.; Cañete, R.; Gil, Á. Genome-wide expression in visceral adipose tissue from obese prepubertal children. Int. J. Mol. Sci. 2015, 16, 7723–7737. [Google Scholar] [CrossRef]
- Deloukas, P.; Kanoni, S.; Willenborg, C.; Farrall, M.; Assimes, T.L.; Thompson, J.R.; Ingelsson, E.; Saleheen, D.; Erdmann, J.; Goldstein, B.A.; et al. Large-scale association analysis identifies new risk loci for coronary artery disease. Nat. Genet. 2013, 45, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Locke, A.E.; Kahali, B.; Berndt, S.I.; Justice, A.E.; Pers, T.H.; Day, F.R.; Powell, C.; Vedantam, S.; Buchkovich, M.L.; Yang, J.; et al. Genetic studies of body mass index yield new insights for obesity biology. Nature 2015, 518, 197–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warrington, N.M.; Howe, L.D.; Paternoster, L.; Kaakinen, M.; Herrala, S.; Huikari, V.; Wu, Y.Y.; Kemp, J.P.; Timpson, N.J.; Pourcain, B.S.; et al. A genome-wide association study of body mass index across early life and childhood. Int. J. Epidemiol. 2015, 44, 700–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jablonski, K.A.; McAteer, J.B.; De Bakker, P.I.W.; Franks, P.W.; Pollin, T.I.; Hanson, R.L.; Saxena, R.; Fowler, S.; Shuldiner, A.R.; Knowler, W.C.; et al. Common variants in 40 genes assessed for diabetes incidence and response to metformin and lifestyle intervention in the diabetes prevention program. Diabetes 2010, 59, 2672–2681. [Google Scholar] [CrossRef] [PubMed]
- Florez, J.C.; Jablonski, K.A.; Taylor, A.; Mather, K.; Horton, E.; White, N.H.; Barrett-Connor, E.; Knowler, W.C.; Shuldiner, A.R.; Pollin, T.I.; et al. The C allele of ATM rs11212617 does not associate with metformin response in the Diabetes Prevention Program. Diabetes Care 2012, 35, 1864–1867. [Google Scholar] [CrossRef]
- Zhou, K.; Yee, S.W.; Seiser, E.L.; van Leeuwen, N.; Tavendale, R.; Bennett, A.J.; Groves, C.J.; Coleman, R.L.; van der Heijden, A.A.; Beulens, J.W.; et al. Variation in the glucose transporter gene SLC2A2 is associated with glycemic response to metformin. Nat. Genet. 2016, 48, 1055–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rupérez, A.I.; Gil, A.; Aguilera, C.M. Genetics of oxidative stress in obesity. Int. J. Mol. Sci. 2014, 15, 3118–3144. [Google Scholar] [CrossRef]
- Wigginton, J.E.; Cutler, D.J.; Abecasis, G.R. A Note on Exact Tests of Hardy-Weinberg Equilibrium. Am. J. Hum. Genet. 2005, 76, 887–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Ojeda, F.J.; Anguita-Ruiz, A.; Rupérez, A.I.; Gomez-Llorente, C.; Olza, J.; Vázquez-Cobela, R.; Gil-Campos, M.; Bueno, G.; Leis, R.; Cañete, R.; et al. Effects of X-chromosome Tenomodulin Genetic Variants on Obesity in a Children’s Cohort and Implications of the Gene in Adipocyte Metabolism. Sci. Rep. 2019, 9, 3979. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Liu, Y.; Zhang, X. Validation of recommended definition in identifying elevated blood pressure in adolescents. J. Clin. Hypertens. 2019, 1–7. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Pawlyk, A.C.; Giacomini, K.M.; McKeon, C.; Shuldiner, A.R.; Florez, J.C. Metformin pharmacogenomics: Current status and future directions. Diabetes 2014, 63, 2590–2599. [Google Scholar] [CrossRef] [PubMed]
- Zhou, K.; Bellenguez, C.; Spencer, C.C.A.; Bennett, A.J.; Coleman, R.L.; Tavendale, R.; Hawley, S.A.; Donnelly, L.A.; Schofield, C.; Groves, C.J.; et al. Common variants near ATM are associated with glycemic response to metformin in type 2 diabetes. Nat. Genet. 2011, 43, 117–120. [Google Scholar] [PubMed]
- Clarson, C.; Mahmud, F.; Baker, J. Metformin in combination with structured lifestyle intervention improved body mass index in obese adolescents, but did not improve insulin resistance. Endocrine 2009, 36, 141–146. [Google Scholar] [CrossRef]
- Yanovski, J.A.; Krakoff, J.; Salaita, C.G.; McDuffie, J.R.; Kozlosky, M.; Sebring, N.G.; Reynolds, J.C.; Brady, S.M.; Calis, K.A. Effects of metformin on body weight and body composition in obese insulin-resistant children: A randomized clinical trial. Diabetes 2011, 60, 477–485. [Google Scholar] [CrossRef]
- Tong, T.; Shen, Y.; Lee, H.-W.; Yu, R.; Park, T. Adenylyl cyclase 3 haploinsufficiency confers susceptibility to diet-induced obesity and insulin resistance in mice. Sci. Rep. 2016, 6, 34179. [Google Scholar] [CrossRef]
- Pruunsild, P.; Kazantseva, A.; Aid, T.; Palm, K.; Timmusk, T. Dissecting the human BDNF locus: Bidirectional transcription, complex splicing, and multiple promoters. Genomics 2007, 90, 397–406. [Google Scholar] [CrossRef] [Green Version]
- Akbarian, S.A.; Salehi-Abargouei, A.; Pourmasoumi, M.; Kelishadi, R.; Nikpour, P.; Heidari-Beni, M. Association of Brain-derived neurotrophic factor gene polymorphisms with body mass index: A systematic review and meta-analysis. Adv. Med. Sci. 2018, 63, 43–56. [Google Scholar] [CrossRef]
- Delahanty, L.M.; Pan, Q.; Jablonski, K.A.; Watson, K.E.; McCaffery, J.M.; Shuldiner, A.; Kahn, S.E.; Knowler, W.C.; Florez, J.C.; Franks, P.W. Genetic predictors of weight loss and weight regain after intensive lifestyle modification, metformin treatment, or standard care in the Diabetes Prevention Program. Diabetes Care 2012, 35, 363–366. [Google Scholar] [CrossRef]
- Yamanaka, M.; Itakura, Y.; Tsuchida, A.; Nakagawa, T.; Taiji, M. Brain-derived neurotrophic factor (BDNF) prevents the development of diabetes in prediabetic mice. Biomed. Res. 2008, 29, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Yoo, D.Y.; Kim, W.; Nam, S.M.; Yoo, K.-Y.; Lee, C.H.; Choi, J.H.; Won, M.-H.; Hwang, I.K.; Yoon, Y.S. Reduced Cell Proliferation and Neuroblast Differentiation in the Dentate Gyrus of High Fat Diet-Fed Mice are Ameliorated by Metformin and Glimepiride Treatment. Neurochem. Res. 2011, 36, 2401–2408. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Liu, J.; Yu, H.; Chen, Y.; Wang, Q.; Xiang, L. Effect of metformin on Schwann cells under hypoxia condition. Int. J. Clin. Exp. Pathol. 2015, 8, 6748–6755. [Google Scholar] [PubMed]
- Eyileten, C.; Kaplon-Cieslicka, A.; Mirowska-Guzel, D.; Malek, L.; Postula, M. Antidiabetic Effect of Brain-Derived Neurotrophic Factor and Its Association with Inflammation in Type 2 Diabetes Mellitus. J. Diabetes Res. 2017, 2017, 2823671. [Google Scholar] [CrossRef] [PubMed]
- Díaz, M.; López-Bermejo, A.; Sánchez-Infantes, D.; Bassols, J.; De Zegher, F.; Ibáñez, L. Responsiveness to metformin in girls with androgen excess: Collective influence of genetic polymorphisms. Fertil. Steril. 2011, 96, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Willer, C.J.; Speliotes, E.K.; Loos, R.J.F.; Li, S.; Lindgren, C.M.; Heid, I.M.; Berndt, S.I.; Elliott, A.L.; Jackson, A.U.; Lamina, C.; et al. Six new loci associated with body mass index highlight a neuronal influence on body weight regulation. Nat. Genet. 2009, 41, 25–34. [Google Scholar]
- Thorleifsson, G.; Walters, G.B.; Gudbjartsson, D.F.; Steinthorsdottir, V.; Sulem, P.; Helgadottir, A.; Styrkarsdottir, U.; Gretarsdottir, S.; Thorlacius, S.; Jonsdottir, I.; et al. Genome-wide association yields new sequence variants at seven loci that associate with measures of obesity. Nat. Genet. 2009, 41, 18–24. [Google Scholar] [CrossRef]
- Gutierrez-Aguilar, R.; Kim, D.-H.; Casimir, M.; Dai, X.-Q.; Pfluger, P.T.; Park, J.; Haller, A.; Donelan, E.; Park, J.; D’Alessio, D.; et al. The role of the transcription factor ETV5 in insulin exocytosis. Diabetologia 2014, 57, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Arner, P.; Sahlqvist, A.-S.; Sinha, I.; Xu, H.; Yao, X.; Waterworth, D.; Rajpal, D.; Loomis, A.K.; Freudenberg, J.M.; Johnson, T.; et al. The epigenetic signature of systemic insulin resistance in obese women. Diabetologia 2016, 59, 2393–2405. [Google Scholar] [CrossRef] [Green Version]
- Hur, K.Y.; Lee, M.-S. New mechanisms of metformin action: Focusing on mitochondria and the gut. J. Diabetes Investig. 2015, 6, 600–609. [Google Scholar] [CrossRef]
- Becker, M.; Visser, L.; van Schaik, R.; Hofman, A.; Uitterlinden, A.; Stricker, B. Genetic variation in the multidrug and toxin extrusion 1 transporter protein influences the glucose-lowering effect of metformin in patients with diabetes: A preliminary study. Diabetes 2009, 58, 745–749. [Google Scholar] [CrossRef]
- Becker, M.L.; Visser, L.E.; van Schaik, R.H.N.; Hofman, A.; Uitterlinden, A.G.; Stricker, B.H.C. Interaction between polymorphisms in the OCT1 and MATE1 transporter and metformin response. Pharmacogenet. Genom. 2010, 20, 38–44. [Google Scholar] [CrossRef]
- Franks, P.W.; Christophi, C.A.; Jablonski, K.A.; Billings, L.K.; Delahanty, L.M.; Horton, E.S.; Knowler, W.C.; Florez, J.C. Common variation at PPARGC1A/B and change in body composition and metabolic traits following preventive interventions: The Diabetes Prevention Program. Diabetologia 2014, 57, 485–490. [Google Scholar] [CrossRef]
- Becker, M.L.; Visser, L.E.; van Schaik, R.H.N.; Hofman, A.; Uitterlinden, A.G.; Stricker, B.H.C. Genetic variation in the organic cation transporter 1 is associated with metformin response in patients with diabetes mellitus. Pharm. J. 2009, 9, 242–247. [Google Scholar] [CrossRef]
- Lopez-Bermejo, A.; Diaz, M.; Moran, E.; de Zegher, F.; Ibanez, L. A Single Nucleotide Polymorphism in STK11 Influences Insulin Sensitivity and Metformin Efficacy in Hyperinsulinemic Girls with Androgen Excess. Diabetes Care 2010, 33, 1544–1548. [Google Scholar] [CrossRef] [PubMed]
- Wing, M.R.; Ziegler, J.M.; Langefeld, C.D.; Roh, B.H.; Palmer, N.D.; Mayer-Davis, E.J.; Rewers, M.J.; Haffner, S.M.; Wagenknecht, L.E.; Bowden, D.W. Analysis of FTO gene variants with obesity and glucose homeostasis measures in the multiethnic Insulin Resistance Atherosclerosis Study cohort. Int. J. Obes. 2011, 35, 1173–1182. [Google Scholar] [CrossRef] [PubMed]
- Franks, P.W.; Jablonski, K.A.; Delahanty, L.M.; McAteer, J.B.; Kahn, S.E.; Knowler, W.C.; Florez, J.C. Assessing gene-treatment interactions at the FTO and INSIG2 loci on obesity-related traits in the Diabetes Prevention Program. Diabetologia 2008, 51, 2214–2223. [Google Scholar] [CrossRef] [PubMed]
- Christensen, M.M.H.; Brasch-Andersen, C.; Green, H.; Nielsen, F.; Damkier, P.; Beck-Nielsen, H.; Brosen, K. The pharmacogenetics of metformin and its impact on plasma metformin steady-state levels and glycosylated hemoglobin A1c. Pharmacogenet. Genom. 2011, 21, 837–850. [Google Scholar] [CrossRef]
- Tkáč, I.; Klimčáková, L.; Javorský, M.; Fabianová, M.; Schroner, Z.; Hermanová, H.; Babjaková, E.; Tkáčová, R. Pharmacogenomic association between a variant in SLC47A1gene and therapeutic response to metformin in type 2 diabetes. Diabetes Obes. Metab. 2013, 15, 189–191. [Google Scholar] [CrossRef]
- Mousavi, S.; Kohan, L.; Yavarian, M.; Habib, A. Pharmacogenetic variation of SLC47A1 gene and metformin response in type2 diabetes patients. Mol. Biol. Res. Commun. 2017, 6, 91–94. [Google Scholar]



{kind=link}
{kind=link}
| Δ Fasting Glucose | Δ Fasting Insulin | Δ HOMA-IR | Δ QUICKI | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| SNP | Nearest Gene | Effect (Other) Allele | Β (95%CI) | p-Value | Β (95%CI) | p-Value | Β (95%CI) | p-Value | Β (95%CI) | p-Value |
| rs11676272 | ADCY3 | A (G) | 0.08 (−2.57, 2.74) | 0.95 | −0.36 (−0.63, −0.08) | 0.01 | −0.31 (−0.58, −0.05) | 0.03 | 0.31 (0.04, 0.59) | 0.03 |
| rs10182181 | ADCY3 | A (G) | −0.32 (−3.03, 2.39) | 0.82 | −0.34 (−0.62, −0.06) | 0.02 | −0.31 (−0.58, −0.04) | 0.03 | 0.27 (−0.01, 0.55) | 0.07 |
| rs17133921 | ARRB1 | A (G) | 0.70 (−4.32, 5.71) | 0.79 | 0.33 (−0.2, 0.86) | 0.23 | 0.32 (−0.18, 0.83) | 0.22 | −0.02 (−0.54, 0.51) | 0.95 |
| rs11030104 | BDNF-AS | G (A) | 2.43 (−1.08, 5.95) | 0.18 | 0.48 (0.11, 0.85) | 0.01 | 0.48 (0.12, 0.83) | 0.01 | −0.47 (−0.82, −0.11) | 0.01 |
| rs1001179 | CAT | T (C) | 0.43 (−2.89, 3.75) | 0.80 | 0.27 (−0.1, 0.64) | 0.16 | 0.24 (−0.12, 0.59) | 0.20 | −0.29 (−0.65, 0.06) | 0.11 |
| rs7943316 | CAT | A (T) | −1.57 (−5.94, 2.8) | 0.49 | −0.63 (−1.07, −0.19) | 0.009 | −0.59 (−1.05, −0.13) | 0.02 | 0.46 (0.04, 0.88) | 0.04 |
| rs7902 | CEP57 | A (G) | −1.02 (−3.82, 1.78) | 0.49 | −0.37 (−0.7, −0.05) | 0.03* | −0.38 (−0.69, −0.08) | 0.02* | 0.49 (0.2, 0.78) | 0.002* |
| rs3763613 | CNTFR | T (G) | −0.78 (−4.35, 2.8) | 0.67 | 0.17 (−0.2, 0.55) | 0.36 | 0.15 (−0.2, 0.51) | 0.41 | −0.06 (−0.42, 0.3) | 0.73 |
| rs7705502 | CPEB4 | A (G) | −0.7 (−4.85, 3.47) | 0.75 | 0.08 (−0.38, 0.54) | 0.74 | 0.02 (−0.42, 0.47) | 0.92 | −0.06 (−0.51, 0.39) | 0.8 |
| rs1902584 | CYP19A1 | T (A) | 2.6 (−3.66, 8.85) | 0.42 | 0.45 (−0.29, 1.18) | 0.24 | 0.35 (−0.36, 1.06) | 0.34 | −0.66 (−1.35, 0.03) | 0.07 |
| rs1516725 | ETV5 | T (C) | −3.85 (−9.04, 1.35) | 0.15 | −0.58 (−1.24, 0.07) | 0.09 | −0.70 (−1.3, −0.1) | 0.03 | 0.57 (−0.05, 1.19) | 0.08 |
| rs10852521 | FTO | T (C) | −2.60 (−5.98, 0.78) | 0.14 | −0.25 (−0.61, 0.12) | 0.19 | −0.25 (−0.59, 0.1) | 0.17 | 0.15 (−0.21, 0.52) | 0.40 |
| rs7566605 | INSIG2 | C (G) | −2.03 (−5.13, 1.08) | 0.21 | −0.11 (−0.45, 0.23) | 0.53 | −0.19 (−0.51, 0.14) | 0.26 | −0.02 (−0.35, 0.32) | 0.93 |
| rs287104 | KCTD15 | G (A) | 0.69 (−2.11, 3.48) | 0.64 | −0.11 (−0.45, 0.23) | 0.53 | −0.17 (−0.46, 0.13) | 0.28 | 0.10 (−0.2, 0.4) | 0.53 |
| rs9932581 | MVD | T (C) | −1.86 (−4.82, 1.1) | 0.23 | −0.43 (−0.75, −0.12) | 0.01 | −0.45 (−0.74, −0.16) | 0.004 | 0.32 (0.01, 0.64) | 0.05 |
| rs3101336 | NEGR1 | T (C) | −0.90 (−3.85, 2.05) | 0.55 | −0.05 (−0.39, 0.28) | 0.76 | −0.08 (−0.4, 0.24) | 0.64 | −0.01 (−0.34, 0.31) | 0.94 |
| rs2815752 | NEGR1 | G (A) | −0.90 (−3.85, 2.05) | 0.55 | −0.05 (−0.39, 0.28) | 0.76 | −0.08 (−0.4, 0.24) | 0.64 | −0.01 (−0.34, 0.31) | 0.94 |
| rs984430 | NTRK2 | T (C) | 1.88 (−2.71, 6.48) | 0.43 | 0.06 (−0.46, 0.57) | 0.83 | 0.16 (−0.33, 0.65) | 0.52 | 0.10 (−0.38, 0.59) | 0.67 |
| rs10868232 | NTRK2 | G (A) | −2.51 (−6.78, 1.76) | 0.26 | −0.43 (−0.89, 0.02) | 0.07 | −0.52 (−0.94, −0.09) | 0.02 | 0.49 (0.06, 0.93) | 0.03 |
| rs1867283 | NTRK2 | G (A) | 0.84 (−2.08, 3.77) | 0.57 | −0.18 (−0.5, 0.13) | 0.26 | −0.18 (−0.49, 0.13) | 0.26 | 0.07 (−0.24, 0.39) | 0.64 |
| rs8192678 | PPARGC1A | T (C) | 2.01 (−1.17, 5.19) | 0.22 | 0.08 (−0.28, 0.44) | 0.66 | 0.11 (−0.23, 0.45) | 0.53 | 0.17 (−0.17, 0.52) | 0.33 |
| rs2970852 | PPARGC1A | T (C) | 1.01 (−1.92, 3.94) | 0.50 | −0.07 (−0.39, 0.26) | 0.70 | −0.05 (−0.36, 0.26) | 0.76 | 0.01 (−0.3,0. 32) | 0.95 |
| rs622342 | SLC22A1 | C (A) | −3.70 (−6.95, −0.46) | 0.03 | −0.31 (−0.69, 0.06) | 0.11 | −0.38 (−0.72, −0.04) | 0.03* | 0.11 (−0.27, 0.49) | 0.57 |
| rs7137767 | SLCO1A2 | A (C) | −3.75 (−6.57, −0.93) | 0.01 | −0.06 (−0.43, 0.3) | 0.74 | −0.14 (−0.48, 0.2) | 0.43 | −0.05 (−0.41, 0.3) | 0.77 |
| rs8111699 | STK11 | C (G) | −0.65 (−3.71, 2.42) | 0.68 | 0.09 (−0.26, 0.45) | 0.62 | 0.12 (−0.22, 0.46) | 0.48 | −0.04 (−0.39, 0.3) | 0.80 |
| rs7903146 | TCF7L2 | T (C) | −1.93 (−5.1, 1.24) | 0.24 | 0.18 (−0.18, 0.54) | 0.33 | 0.05 (−0.24, 0.4) | 0.77 | −0.06 (−0.41, 0.29) | 0.72 |
| rs6548238 | TMEM18 | T (C) | −0.83 (−4.93, 3.27) | 0.69 | 0.18 (−0.27, 0.64) | 0.43 | 0.08 (−0.36, 0.51) | 0.73 | −0.16 (−0.6, 0.28) | 0.47 |
| rs3737787 | USF-1 | A (G) | 0.37 (−2.84, 3.58) | 0.82 | 0.25 (−0.12, 0.61) | 0.19 | 0.22 (−0.12, 0.57) | 0.21 | −0.02 (−0.38, 0.34) | 0.92 |
| Δ BMI Z Score | Δ WC (cm) | Δ DBP | Δ SBP | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| SNP | Nearest Gene | Effect (other) Allele | Β (95%CI) | p-Value | Β (95%CI) | p-Value | Β (95%CI) | p-Value | Β (95%CI) | p-Value |
| rs11676272 | ADCY3 | A (G) | −0.27 (−0.58, 0.04) | 0.09 | −1.35 (−3.2, 0.5) | 0.16 | −2.58 (−5.39, 0.22) | 0.08 | −0.57 (−4.61, 3.47) | 0.78 |
| rs10182181 | ADCY3 | A (G) | −0.26 (−0.59, 0.07) | 0.13 | −1.43 (−3.31, 0.45) | 0.14 | −2.26 (−5.08, 0.56) | 0.12 | −1.13 (−5.29, 3.03) | 0.60 |
| rs17133921 | ARRB1 | A (G) | −0.14 (−0.76, 0.47) | 0.65 | −4.38 (−7.19, −1.56) | 0.004* | −4.06 (−9.56, 1.44) | 0.16 | −8.22 (−15.58, −0.86) | 0.03 |
| rs11030104 | BDNF−AS | G (A) | 0.40 (−0.02, 0.81) | 0.07 | 0.97 (−1.28, 3.22) | 0.40 | 2.47 (−1.32, 6.26) | 0.21 | −2.59 (−8.1, 2.92) | 0.36 |
| rs1001179 | CAT | T (C) | 0.08 (−0.32, 0.49) | 0.68 | 3.03 (1.24, 4.82) | 0.002* | 0.31 (−3.34, 3.96) | 0.87 | −1.59 (−6.51, 3.34) | 0.53 |
| rs7943316 | CAT | A (T) | 0.002 (−0.53, 0.54) | 0.99 | −1.92 (−4.58, 0.75) | 0.17 | 0.69 (−4.12, 5.50) | 0.78 | 1.68 (−4.66, 8.02) | 0.61 |
| rs7902 | CEP57 | A (G) | −0.21 (−0.57, 0.15) | 0.26 | 1.07 (−0.52, 2.66) | 0.19 | 0.79 (−2.43, 4.01) | 0.63 | −0.03 (−4.52, 4.46) | 0.99 |
| rs3763613 | CNTFR | T (G) | 0.55 (0.19, 0.91) | 0.004* | 0.87 (−1.17, 2.91) | 0.41 | 0.27 (−3.31, 3.84) | 0.88 | 2.27 (−2.81, 7.36) | 0.39 |
| rs7705502 | CPEB4 | A (G) | 0.25 (−0.82, 0.26) | 0.32 | 2.87 (0.02, 5.71) | 0.05 | −2.92 (−7.31, 1.47) | 0.20 | −2.62 (−8.99, 3.75) | 0.43 |
| rs1902584 | CYP19A1 | T (A) | −0.96 (−1.59, −0.32) | 0.005* | 1.63 (−2.14, 5.4) | 0.40 | −8.93 (−15.2, −2.67) | 0.008 | −5.67 (−15.70, 4.36) | 0.27 |
| rs1516725 | ETV5 | T (C) | 0.54 (−0.05, 1.14) | 0.08 | 0.52 (−2.51, 3.55) | 0.74 | −4.48 (−9.70, 0.73) | 0.10 | −8.49 (−15.99, −0.99) | 0.03 |
| rs10852521 | FTO | T (C) | −0.27 (−0.67, 0.11) | 0.17 | −2.41 (−4.44, −0.38) | 0.02 | −4.79 (−8.07, −1.52) | 0.007 | −0.08 (−5.2, 5.04) | 0.98 |
| rs7566605 | INSIG2 | C (G) | −0.08 (−0.47, 0.32) | 0.71 | −0.30 (−2.3, 1.7) | 0.77 | −1.05 (−4.51, 2.42) | 0.56 | −1.72 (−6.63, 3.20) | 0.50 |
| rs287104 | KCTD15 | G (A) | −0.01 (−0.36, 0.35) | 0.97 | 0.8 (−0.9, 2.5) | 0.36 | 1.56 (−1.38, 4.51) | 0.30 | 0.29 (−3.90, 4.47) | 0.89 |
| rs9932581 | MVD | T (C) | 0.12 (−0.22, 0.47) | 0.49 | −1.52 (−3.1, 0.06) | 0.07 | −0.64 (−4.01, 2.73) | 0.71 | −0.52 (−5.18, 4.15) | 0.83 |
| rs3101336 | NEGR1 | T (C) | −0.07 (−0.44, 0.3) | 0.72 | 0.33 (−1.51, 2.16) | 0.73 | −2.51 (−5.55, 0.53) | 0.11 | −6.22 (−10.28, −2.15) | 0.005 |
| rs2815752 | NEGR1 | G (A) | −0.07 (−0.44, 0.3) | 0.72 | 0.33 (−1.51, 2.16) | 0.73 | −2.51 (−5.55, 0.53) | 0.11 | −6.22 (−10.28, −2.15) | 0.005 |
| rs984430 | NTRK2 | T (C) | 0.56 (0.05, 1.08) | 0.03* | 0.28 (−2.57, 3.12) | 0.85 | −0.67 (−5.67, 4.33) | 0.80 | −0.93 (−8.16, 6.29) | 0.80 |
| rs10868232 | NTRK2 | G (A) | 0.10 (−0.4, 0.61) | 0.69 | 0.36 (−2.25, 2.96) | 0.79 | −3.73 (−8.38, 0.93) | 0.12 | 0.16 (−7.36, 7.68) | 0.97 |
| rs1867283 | NTRK2 | G (A) | −0.05 (−0.39, 0.3) | 0.78 | 0.80 (−0.95, 2.55) | 0.38 | −1.79 (−4.89, 1.31) | 0.27 | −2.38 (−6.87, 2.10) | 0.30 |
| rs8192678 | PPARGC1A | T (C) | 0.51 (0.16, 0.87) | 0.007* | −0.02 (−2.07, 2.04) | 0.99 | −0.26 (−3.76, 3.25) | 0.89 | 1.57 (−3.73, 6.86) | 0.57 |
| rs2970852 | PPARGC1A | T (C) | −0.33 (−0.68, 0.01) | 0.06 | −0.20 (−1.98, 1.59) | 0.83 | −0.90 (−4.02, 2.22) | 0.57 | −1.43 (−5.99, 3.12) | 0.54 |
| rs622342 | SLC22A1 | C (A) | 0.14 (−0.25, 0.54) | 0.47 | 2.14 (0.16, 4.12) | 0.05 | 1.05 (−2.53, 4.63) | 0.57 | −0.57 (−5.44, 4.30) | 0.82 |
| rs7137767 | SLCO1A2 | A (C) | 0.15 (−0.22, 0.52) | 0.44 | −0.44 (−2.46, 1.58) | 0.67 | 0.19 (−3.18, 3.56) | 0.91 | −4.21 (−8.74, 0.32) | 0.08 |
| rs8111699 | STK11 | C (G) | −0.26 (−0.62, 0.1) | 0.17 | −1.27 (−2.94, 0.4) | 0.14 | −0.94 (−4.24, 2.35) | 0.58 | 0.36 (−4.37, 5.09) | 0.88 |
| rs7903146 | TCF7L2 | T (C) | 0.24 (−0.14, 0.62) | 0.22 | −0.15 (−2.17, 1.86) | 0.88 | −1.20 (−4.68, 2.28) | 0.50 | −1.71 (−6.68, 3.26) | 0.50 |
| rs6548238 | TMEM18 | T (C) | −0.43 (−0.93, 0.06) | 0.09 | 1.57 (−0.9, 4.04) | 0.22 | 1.86 (−2.47, 6.18) | 0.41 | 0.40 (−5.84, 6.65) | 0.90 |
| rs3737787 | USF−1 | A (G) | −0.57 (−0.91, −0.24) | 0.001* | −0.1 (−2.03, 1.83) | 0.92 | −2.80 (−6.11, 0.51) | 0.11 | −3.59 (−8.36, 1.18) | 0.15 |
| Δ LDLc | Δ Total Cholesterol | Δ Triglycerides | Δ Apo A1 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| SNP | Nearest Gene | Effect (other) Allele | Β (95%CI) | p-Value | Β (95%CI) | p-Value | Β (95%CI) | p-Value | Β (95%CI) | p-Value |
| rs11676272 | ADCY3 | A (G) | −5.77 (−12.17, 0.63) | 0.08 | −8.57 (−14.91, −2.22) | 0.01* | 0.09 (−0.25, 0.42) | 0.61 | 2.28 (−6.84, 11.4) | 0.63 |
| rs10182181 | ADCY3 | A (G) | −5.77 (−12.4, 0.85) | 0.09 | −8.77 (−15.03, −2.51) | 0.009* | 0.09 (−0.24, 0.42) | 0.61 | −0.44 (−9.48, 8.59) | 0.92 |
| rs17133921 | ARRB1 | A (G) | −4.93 (−16.57, 6.71) | 0.41 | −7.44 (−18.77, 3.89) | 0.20 | −0.25 (−0.82, 0.32) | 0.40 | 2.33 (−11.17, 15.83) | 0.74 |
| rs11030104 | BDNF−AS | G (A) | 9.47 (1.07, 17.87) | 0.03 | 8.38 (0.12, 16.63) | 0.05 | −0.38 (−0.8, 0.04) | 0.09 | 0.58 (−11.45, 12.62) | 0.92 |
| rs1001179 | CAT | T (C) | 9.99 (2.34, 17.64) | 0.01 | 7.73 (−0.13, 15.6) | 0.06 | −0.16 (−0.6, 0.29) | 0.49 | −5.44 (−16.15, 5.27) | 0.33 |
| rs7943316 | CAT | A (T) | −0.98 (−11.82, 9.86) | 0.86 | 1.02 (−9.48, 11.51) | 0.85 | 0.12 (−0.41, 0.65) | 0.66 | 2.35 (−10.96, 15.66) | 0.73 |
| rs7902 | CEP57 | A (G) | 2.74 (−3.99, 9.47) | 0.43 | −0.62 (−7.37, 6.12) | 0.86 | 0.13 (−0.21, 0.47) | 0.47 | −0.59 (−10.2, 9.01) | 0.90 |
| rs3763613 | CNTFR | T (G) | 2.50 (−5.86, 10.86) | 0.56 | 3.33 (−4.8, 11.47) | 0.43 | −0.13 (−0.53, 0.27) | 0.52 | −3.52 (−14.83, 7.79) | 0.55 |
| rs7705502 | CPEB4 | A (G) | 12.87 (3.31, 22.43) | 0.01 | 13.81 (4.77, 22.85) | 0.005 | 0.30 (−0.19, 0.79) | 0.23 | 6.05 (−6.82, 18.91) | 0.36 |
| rs1902584 | CYP19A1 | T (A) | −1.27 (−16.78, 14.23) | 0.87 | 0.62 (−15.15, 16.4) | 0.94 | −0.37 (−1.12, 0.37) | 0.33 | −14.67 (−37.91, 8.56) | 0.23 |
| rs1516725 | ETV5 | T (C) | 7.95 (−4.99, 20.9) | 0.24 | 1.54 (−10.95, 14.02) | 0.81 | 0.25 (−0.32, 0.82) | 0.39 | −5.14 (−21.37, 11.08) | 0.54 |
| rs10852521 | FTO | T (C) | 5.27 (−2.78, 13.33) | 0.21 | 3.68 (−4.31, 11.66) | 0.37 | −0.10 (−0.49, 0.29) | 0.63 | −4.86 (−15.38, 5.65) | 0.37 |
| rs7566605 | INSIG2 | C (G) | 5.19 (−2.3, 12.67) | 0.18 | 7.74 (0.67, 14.82) | 0.03 | 0.26 (−0.1, 0.63) | 0.17 | 4.71 (−6.65, 16.07) | 0.42 |
| rs287104 | KCTD15 | G (A) | −3.35 (−10.17, 3.46) | 0.34 | −4.06 (−11.22, 3.09) | 0.27 | 0.08 (−0.26, 0.42) | 0.65 | −9.81 (−17.56, −2.06) | 0.02 |
| rs9932581 | MVD | T (C) | −5.45 (−12.19, 1.28) | 0.12 | −5.45 (−12.45, 1.54) | 0.13 | 0.12 (−0.25, 0.49) | 0.52 | −1.70 (−9.6, 6.2) | 0.67 |
| rs3101336 | NEGR1 | T (C) | 2.22 (−5.03, 9.47) | 0.55 | 1.45 (−5.63, 8.53) | 0.69 | −0.34 (−0.68, 0) | 0.06 | −2.85 (−12.17, 6.47) | 0.55 |
| rs2815752 | NEGR1 | G (A) | 2.22 (−5.03, 9.47) | 0.55 | 1.45 (−5.63, 8.53) | 0.69 | −0.34 (−0.68, 0) | 0.06 | −2.85 (−12.17, 6.47) | 0.55 |
| rs984430 | NTRK2 | T (C) | 2.61 (−9.08, 14.3) | 0.66 | 3.52 (−7.69, 14.73) | 0.54 | −0.03 (−0.58, 0.52) | 0.91 | 5.15 (−8.86, 19.15) | 0.48 |
| rs10868232 | NTRK2 | G (A) | −0.77 (−11.43, 9.89) | 0.89 | 2.05 (−8.37, 12.47) | 0.70 | 0.08 (−0.45, 0.62) | 0.76 | 11.95 (−0.08, 23.98) | 0.06 |
| rs1867283 | NTRK2 | G (A) | −0.47 (−8.03, 7.08) | 0.90 | −0.94 (−8.15, 6.27) | 0.80 | −0.10 (−0.45, 0.26) | 0.59 | −2.87 (−11.91, 6.17) | 0.54 |
| rs8192678 | PPARGC1A | T (C) | 2.96 (−5.14, 11.06) | 0.48 | 4.43 (−3.32, 12.18) | 0.27 | 0.06 (−0.33, 0.45) | 0.77 | 4.69 (−5.55, 14.93) | 0.38 |
| rs2970852 | PPARGC1A | T (C) | −0.60 (−8.01, 6.81) | 0.87 | −1.57 (−8.77, 5.64) | 0.67 | −0.36 (−0.7, −0.03) | 0.04 | 1.80 (−7.26, 10.85) | 0.70 |
| rs622342 | SLC22A1 | C (A) | −0.99 (−8.87, 6.89) | 0.81 | −1.27 (−9.3, 6.77) | 0.76 | 0.36 (−0.04, 0.76) | 0.08 | −2.15 (−11.77, 7.47) | 0.66 |
| rs7137767 | SLCO1A2 | A (C) | 3.45 (−4.22, 11.12) | 0.39 | 1.34 (−6.13, 8.8) | 0.73 | 0.23 (−0.15, 0.6) | 0.24 | −7.87 (−17.37, 1.62) | 0.12 |
| rs8111699 | STK11 | C (G) | −5.23 (−12.3, 1.83) | 0.15 | −4.07 (−11.02, 2.88) | 0.26 | 0.02 (−0.35, 0.39) | 0.93 | 6.43 (−2.94, 15.81) | 0.19 |
| rs7903146 | TCF7L2 | T (C) | 2.98 (−5.0, 11.0) | 0.47 | 1.49 (−6.74, 9.73) | 0.72 | −0.44 (−0.8, −0.07) | 0.02* | −10.90 (−20.81, −1) | 0.04 |
| rs6548238 | TMEM18 | T (C) | −14.44 (−23.88, −5) | 0.005 | −9.01 (−18.81, 0.8) | 0.08 | −0.08 (−0.59, 0.43) | 0.76 | 0.18 (−11.8, 12.16) | 0.98 |
| rs3737787 | USF−1 | A (G) | 2.51 (−5.4, 10.42) | 0.54 | 0.71 (−7.05, 8.47) | 0.86 | −0.07 (−0.46, 0.32) | 0.73 | −2.64 (−12.02, 6.75) | 0.59 |
| Δ Adiponectin | Δ Leptin | Δ ALR | Δ Fat Mass | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| SNP | Nearest Gene | Effect (other) Allele | Β (95%CI) | p-Value | Β (95%CI) | p-Value | Β (95%CI) | p-Value | Β (95%CI) | p-Value |
| rs11676272 | ADCY3 | A (G) | −0.43 (−0.76, −0.10) | 0.01* | −1.82 (−4.21, 0.57) | 0.14 | −0.12 (−0.47, 0.22) | 0.50 | −0.23 (−1.84, 1.38) | 0.78 |
| rs10182181 | ADCY3 | A (G) | −0.47 (−0.79, −0.14) | 0.008* | −2.17 (−4.59, 0.25) | 0.09 | −0.14 (−0.50, 0.22) | 0.44 | −0.21 (−1.86, 1.43) | 0.80 |
| rs17133921 | ARRB1 | A (G) | 0.20 (−0.42, 0.82) | 0.53 | −2.32 (−6.78, 2.15) | 0.32 | 0.36 (−0.27, 0.98) | 0.27 | −0.39 (−3.2, 2.43) | 0.79 |
| rs11030104 | BDNF−AS | G (A) | 0.14 (−0.33, 0.62) | 0.56 | −0.62 (−3.95, 2.71) | 0.72 | 0.10 (−0.36, 0.56) | 0.68 | −0.48 (−2.6, 1.64) | 0.66 |
| rs1001179 | CAT | T (C) | −0.16 (−0.61, 0.30) | 0.51 | 0.17 (−2.89, 3.23) | 0.91 | −0.23 (−0.66, 0.21) | 0.31 | 1.40 (−0.62, 3.41) | 0.18 |
| rs7943316 | CAT | A (T) | 0.24 (−0.34, 0.83) | 0.42 | 0.81 (−3.55, 5.17) | 0.72 | 0.36 (−0.26, 0.97) | 0.27 | −0.69 (−3.27, 1.89) | 0.60 |
| rs7902 | CEP57 | A (G) | −0.08 (−0.45, 0.30) | 0.69 | 0.25 (−2.22, 2.71) | 0.85 | −0.22 (−0.62, 0.17) | 0.28 | 0.22 (−1.52, 1.96) | 0.80 |
| rs3763613 | CNTFR | T (G) | −0.05 (−0.50, 0.40) | 0.84 | 0.92 (−2.37, 4.21) | 0.59 | 0.05 (−0.42, 0.53) | 0.83 | 0.19 (−1.79, 2.17) | 0.85 |
| rs7705502 | CPEB4 | A (G) | −0.51 (−1.05, 0.04) | 0.07 | −0.09 (−4.09, 3.92) | 0.97 | −0.21 (−0.76, 0.34) | 0.46 | −0.13 (−2.65, 2.4) | 0.92 |
| rs1902584 | CYP19A1 | T (A) | −0.21 (−1.05, 0.62) | 0.62 | −0.75 (−6.75, 5.24) | 0.81 | −0.53 (−1.38, 0.31) | 0.22 | 2.67 (−0.92, 6.27) | 0.15 |
| rs1516725 | ETV5 | T (C) | 0.06 (−0.74, 0.87) | 0.88 | −2.14 (−7.03, 2.75) | 0.40 | −0.03 (−0.82, 0.77) | 0.94 | 1.07 (−2.14, 4.29) | 0.52 |
| rs10852521 | FTO | T (C) | −0.06 (−0.50, 0.38) | 0.79 | −2.26 (−5.24, 0.72) | 0.14 | 0.09 (−0.37, 0.54) | 0.72 | −1.38 (−3.27, 0.52) | 0.16 |
| rs7566605 | INSIG2 | C (G) | −0.07 (−0.49, 0.35) | 0.75 | 0.06 (−2.85, 2.96) | 0.97 | 0.14 (−0.28, 0.56) | 0.52 | −0.03 (−1.86, 1.8) | 0.97 |
| rs287104 | KCTD15 | G (A) | 0.17 (−0.20, 0.54) | 0.36 | 0.46 (−2.17, 3.08) | 0.74 | 0.22 (−0.14, 0.58) | 0.24 | 0.62 (−0.59, 1.83) | 0.32 |
| rs9932581 | MVD | T (C) | 0.001 (−0.41, 0.40) | 0.99 | −2.08 (−4.60, 0.44) | 0.11 | 0.10 (−0.27, 0.47) | 0.59 | 0.50 (−1.22, 2.22) | 0.57 |
| rs3101336 | NEGR1 | T (C) | 0.16 (−0.23, 0.55) | 0.42 | −1.98 (−4.66, 0.69) | 0.15 | −0.02 (−0.41, 0.36) | 0.90 | 0.72 (−1, 2.45) | 0.41 |
| rs2815752 | NEGR1 | G (A) | 0.16 (−0.23, 0.55) | 0.42 | −1.98 (−4.66, 0.69) | 0.15 | −0.02 (−0.41, 0.36) | 0.90 | 0.72 (−1, 2.45) | 0.41 |
| rs984430 | NTRK2 | T (C) | −0.03 (−0.66, 0.61) | 0.94 | −4.09 (−8.31, 0.12) | 0.06 | 0.12 (−0.47, 0.71) | 0.70 | −0.08 (−2.77, 2.61) | 0.95 |
| rs10868232 | NTRK2 | G (A) | 0.60 (0.06, 1.13) | 0.03 | 1.39 (−2.60, 5.37) | 0.50 | 0.29 (−0.24, 0.82) | 0.29 | 1.63 (−0.87, 4.14) | 0.21 |
| rs1867283 | NTRK2 | G (A) | 0.33 (−0.04, 0.70) | 0.09 | 0.44 (−2.26, 3.14) | 0.75 | 0.43 (0.08, 0.77) | 0.02* | 0.52 (−1.19, 2.22) | 0.56 |
| rs8192678 | PPARGC1A | T (C) | −0.05 (−0.51, 0.42) | 0.85 | 2.87 (−0.07, 5.81) | 0.06 | −0.10 (−0.54, 0.34) | 0.67 | −0.15 (−2.05, 1.75) | 0.88 |
| rs2970852 | PPARGC1A | T (C) | 0.13 (−0.25, 0.52) | 0.50 | −0.36 (−3.16, 2.44) | 0.80 | 0.11 (−0.25, 0.48) | 0.54 | 1.04 (−0.65, 2.72) | 0.23 |
| rs622342 | SLC22A1 | C (A) | −0.17 (−0.64, 0.30) | 0.49 | 0.77 (−2.18, 3.72) | 0.61 | 0.03 (−0.39, 0.44) | 0.90 | −0.02 (−1.93, 1.89) | 0.98 |
| rs7137767 | SLCO1A2 | A (C) | 0.20 (−0.23, 0.64) | 0.37 | 0.12 (−2.90, 3.13) | 0.94 | 0.07 (−0.37, 0.50) | 0.77 | 0.56 (−1.17, 2.29) | 0.53 |
| rs8111699 | STK11 | C (G) | −0.30 (−0.69, 0.09) | 0.14 | −2.76 (−5.17, −0.34) | 0.03 | −0.16(−0.54, 0.23) | 0.43 | −0.09 (−1.87, 1.68) | 0.92 |
| rs7903146 | TCF7L2 | T (C) | 0.16 (−0.26, 0.59) | 0.46 | −1.26 (−4.22, 1.71) | 0.41 | 0.13 (−0.28, 0.55) | 0.54 | 0.98 (−0.9, 2.85) | 0.31 |
| rs6548238 | TMEM18 | T (C) | 0.03 (−0.53, 0.60) | 0.91 | 0.57 (−3.30, 4.44) | 0.77 | −0.33 (−0.86, 0.21) | 0.24 | 0.94 (−1.46, 3.35) | 0.45 |
| rs3737787 | USF−1 | A (G) | −0.25 (−0.71, 0.20) | 0.28 | 0.15 (−3.09, 3.39) | 0.93 | −0.45 (−0.86, −0.04) | 0.05 | 0.70 (−1.17, 2.58) | 0.46 |
| Δ INF−γ | Δ CRP | |||||
|---|---|---|---|---|---|---|
| SNP | Nearest Gene | Effect (other) Allele | Β (95%CI) | p-Value | Β (95%CI) | p-Value |
| rs11676272 | ADCY3 | A (G) | −0.45 (−0.73, −0.17) | 0.003 | −0.26 (−0.64, 0.12) | 0.19 |
| rs10182181 | ADCY3 | A (G) | −0.45 (−0.74, −0.16) | 0.004 | −0.18 (−0.57, 0.20) | 0.36 |
| rs17133921 | ARRB1 | A (G) | 0.21 (−0.33, 0.75) | 0.44 | −0.46 (−1.23, 0.31) | 0.24 |
| rs11030104 | BDNF−AS | G (A) | 0.11 (−0.34, 0.56) | 0.63 | 0.25 (−0.24, 0.75) | 0.32 |
| rs1001179 | CAT | T (C) | 0.12 (−0.26, 0.50) | 0.54 | 0.49 (0.05, 0.93) | 0.03* |
| rs7943316 | CAT | A (T) | −0.13 (−0.63, 0.36) | 0.61 | 0.12 (−0.51, 0.75) | 0.72 |
| rs7902 | CEP57 | A (G) | −0.13 (−0.46, 0.20) | 0.45 | 0.07 (−0.34, 0.49) | 0.72 |
| rs3763613 | CNTFR | T (G) | −0.08 (−0.50, 0.33) | 0.69 | 0.23 (−0.26, 0.71) | 0.37 |
| rs7705502 | CPEB4 | A (G) | −0.24 (−0.73, 0.24) | 0.33 | 0.05 (−0.56, 0.66) | 0.87 |
| rs1902584 | CYP19A1 | T (A) | −0.10 (−0.88, 0.69) | 0.81 | 0.50 (−0.44, 1.43) | 0.31 |
| rs1516725 | ETV5 | T (C) | −1.13 (−1.68, −0.59) | <0.001 | 0.06 (−0.73, 0.84) | 0.89 |
| rs10852521 | FTO | T (C) | −0.13 (−0.50, 0.24) | 0.51 | 0.18 (−0.27, 0.64) | 0.43 |
| rs7566605 | INSIG2 | C (G) | −0.27 (−0.63, 0.08) | 0.14 | −0.31 (−0.73, 0.11) | 0.16 |
| rs287104 | KCTD15 | G (A) | −0.14 (−0.47, 0.20) | 0.43 | 0.40 (0.02, 0.79) | 0.05 |
| rs9932581 | MVD | T (C) | −0.51 (−0.82, −0.21) | 0.002 | −0.11 (−0.52, 0.30) | 0.61 |
| rs3101336 | NEGR1 | T (C) | −0.06 (−0.40, 0.28) | 0.72 | 0.57 (0.15, 1.00) | 0.01 |
| rs2815752 | NEGR1 | G (A) | −0.06 (−0.40, 0.28) | 0.72 | 0.57 (0.15, 1.00) | 0.01 |
| rs984430 | NTRK2 | T (C) | 0.09 (−0.46, 0.64) | 0.75 | 0.46 (−0.19, 1.11) | 0.17 |
| rs10868232 | NTRK2 | G (A) | −0.24 (−0.74, 0.26) | 0.36 | −0.10 (−0.71, 0.50) | 0.75 |
| rs1867283 | NTRK2 | G (A) | −0.01 (−0.36, 0.34) | 0.96 | 0.24 (−0.18, 0.65) | 0.27 |
| rs8192678 | PPARGC1A | T (C) | 0.02 (−0.35, 0.40) | 0.91 | −0.09 (−0.54, 0.36) | 0.69 |
| rs2970852 | PPARGC1A | T (C) | −0.05 (−0.39, 0.29) | 0.77 | 0.02 (−0.38, 0.42) | 0.92 |
| rs622342 | SLC22A1 | C (A) | −0.27 (−0.68, 0.13) | 0.20 | 0.12 (−0.35, 0.60) | 0.62 |
| rs7137767 | SLCO1A2 | A (C) | −0.18 (−0.54, 0.19) | 0.35 | 0.08 (−0.36, 0.53) | 0.71 |
| rs8111699 | STK11 | C (G) | 0.21 (−0.14, 0.55) | 0.25 | −0.11 (−0.53, 0.31) | 0.61 |
| rs7903146 | TCF7L2 | T (C) | 0.06 (−0.32, 0.45) | 0.75 | 0.16 (−0.31, 0.62) | 0.51 |
| rs6548238 | TMEM18 | T (C) | 0.43 (−0.04, 0.89) | 0.08 | −0.10 (−0.74, 0.53) | 0.75 |
| rs3737787 | USF−1 | A (G) | 0.14 (−0.26, 0.53) | 0.50 | −0.37 (−0.83, 0.09) | 0.13 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anguita-Ruiz, A.; Pastor-Villaescusa, B.; Leis, R.; Bueno, G.; Hoyos, R.; Vázquez-Cobela, R.; Latorre-Millán, M.; Cañete, M.D.; Caballero-Villarraso, J.; Gil, Á.; et al. Common Variants in 22 Genes Regulate Response to Metformin Intervention in Children with Obesity: A Pharmacogenetic Study of a Randomized Controlled Trial. J. Clin. Med. 2019, 8, 1471. https://doi.org/10.3390/jcm8091471
Anguita-Ruiz A, Pastor-Villaescusa B, Leis R, Bueno G, Hoyos R, Vázquez-Cobela R, Latorre-Millán M, Cañete MD, Caballero-Villarraso J, Gil Á, et al. Common Variants in 22 Genes Regulate Response to Metformin Intervention in Children with Obesity: A Pharmacogenetic Study of a Randomized Controlled Trial. Journal of Clinical Medicine. 2019; 8(9):1471. https://doi.org/10.3390/jcm8091471
Chicago/Turabian StyleAnguita-Ruiz, Augusto, Belén Pastor-Villaescusa, Rosaura Leis, Gloria Bueno, Raúl Hoyos, Rocío Vázquez-Cobela, Miriam Latorre-Millán, M. Dolores Cañete, Javier Caballero-Villarraso, Ángel Gil, and et al. 2019. "Common Variants in 22 Genes Regulate Response to Metformin Intervention in Children with Obesity: A Pharmacogenetic Study of a Randomized Controlled Trial" Journal of Clinical Medicine 8, no. 9: 1471. https://doi.org/10.3390/jcm8091471