The Role of Mesenchymal Stem Cells in Atherosclerosis: Prospects for Therapy via the Modulation of Inflammatory Milieu


 ,
, 

Abstract
1. Introduction
2. Characteristics of MSCs
3. Origin of MSCs
4. Colonization and Migration of MSCs
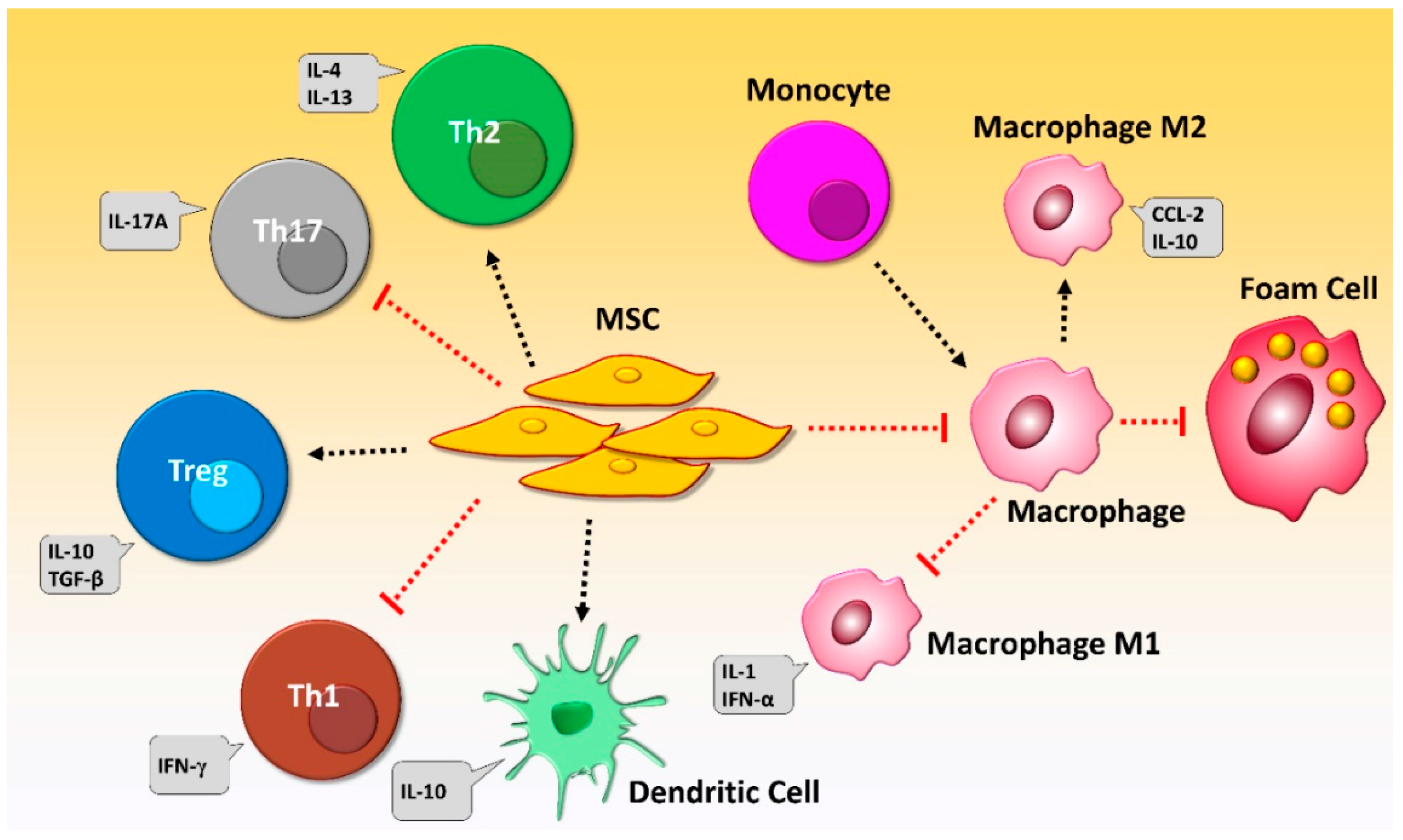
5. Immunomodulatory Properties of MSCs
6. Pathophysiology of Atherosclerosis and the Role of Inflammation
7. Modulation of Inflammatory Mediators by MSCs during Atherosclerosis
8. Prospects for MSC-Based Therapy of Atherosclerosis
8.1. MSCs and the Improvement of Endothelial Function during Atherosclerosis
8.2. MSCs and Regulatory T Cell Development during Atherosclerosis
8.3. Recruitment of MSCs into Atherosclerotic Plaques
8.4. The Role of MSCs in Modulating Lipid Levels
8.5. Stability of Atherosclerotic Plaque and MSCs
9. Drawbacks of MSCs Therapy in Atherosclerosis
10. Investigation of MSC Potential to Treat Atherosclerosis in the Clinical Setting
11. Conclusions and Future Challenges and Perspectives
Author Contributions
Conflicts of Interest
Abbreviations
| MSC | mesenchymal stem cell |
| EC | endothelial cell |
| ox-LDL | oxidized low-density lipoprotein |
| PRRs | pattern recognition receptors |
| IL | interleukin |
| BM-MSC | bone marrow-derived mesenchymal stem cell |
| UC-MSC | umbilical cord—derived mesenchymal stem cell |
| GPCR | G-protein coupled receptor |
| VEGF/VEGFR | vascular endothelial growth factor/vascular endothelial growth factor receptor |
| SCF-c-Kit | stem cell factor—tyrosine kinase receptor |
| SDF-1 | stromal cell-derived factor-1 |
| CXCR4 | CXC chemokine receptor-4 |
| HGF/c-Met | hepatocyte growth factor |
| PDGF/PDGFR | platelet-derived growth factor/platelet-derived growth factor receptor |
| MCP-1/CCR2 | monocyte chemoattractant protein-1/CC chemokine receptor 2 |
| HMGB1/RAGE | high mobility group box 1/receptor of advanced glycation end products |
| TGF-β | transforming growth factor-β |
| CXCL | chemokine (C-X-C motif) ligand |
| CCL | chemokine (C-C motif) ligand |
| EGFR | epidermal growth factor receptor |
| MMP | matrix metalloproteinase |
| GVHD | graft versus host disease |
| IDO | indoleamine-pyrrole-2-3-dioxygenase |
| DC | dendritic cell |
| NK | natural killer |
| IFN | interferon |
| TNF-α | tumor necrosis factor α |
| DC1 | dendritic cell type 1 |
| TSG-6 | TNF-α-stimulated gene-6 |
| TLR | toll-like receptor |
| ApoE | apolipoprotein E |
| LDLR | low-density lipoprotein receptor |
| S-MSC | skin-derived MSC |
| Treg cell | regulatory T cell |
| PGE2 | prostaglandin E2 |
| hAMSC | human amnion mesenchymal stem cell |
| NO | nitric oxide |
| eNOS | endothelial nitric oxide synthase |
| cAMP | cyclic guanosine monophosphate |
| A-MSC | amnion-derived mesenchymal stem cell |
| MIP-2 | macrophage inflammatory protein 2 |
| FOXP3 | forkhead box transcription factor |
| mRNA | messenger RNA |
| BrdU | 5-bromo-2′-deoxyuridine |
| VLDL | very low density lipoprotein |
| iPSC-MSC/iMSC | induced pluripotent stem cells- derived mesenchymal stem cell |
| SREBP-1c | sterol response element binding protein-1c |
| CRP | C-reactive protein |
| PAI-1 | plasminogen activator inhibitor-1 |
| CVD | cardiovascular disease |
| PAD | peripheral arterial disease |
| CLI | critical limb ischemia |
| ERC | endometrial regenerative cell |
| SLI | severe limb ischemia |
| PAOD | peripheral arterial occlusive disease |
| ASC | adipose-derived stem/stroma cell |
| ECM | extracellular matrix |
| YAP | Yes-associated protein |
| hs-CRP | high-sensitivity C-reactive protein |
| iNOS | inducible nitric oxide synthase |
| SRA | class A scavenger receptor |
References
- Mahmoudi, M.; Aslani, S.; Fadaei, R.; Jamshidi, A.R. New insights to the mechanisms underlying atherosclerosis in rheumatoid arthritis. Int. J. Rheum. Dis. 2017, 20, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Sanjadi, M.; Rezvanie Sichanie, Z.; Totonchi, H.; Karami, J.; Rezaei, R.; Aslani, S. Atherosclerosis and autoimmunity: A growing relationship. Int. J. Rheum. Dis. 2018, 21, 908–921. [Google Scholar] [CrossRef] [PubMed]
- Glass, C.K.; Witztum, J.L. Atherosclerosis: The road ahead. Cell 2001, 104, 503–516. [Google Scholar] [CrossRef]
- Baldassarre, D.; Castelnuovo, S.; Frigerio, B.; Amato, M.; Werba, J.P.; De Jong, A.; Ravani, A.L.; Tremoli, E.; Sirtori, C.R. Effects of timing and extent of smoking, type of cigarettes, and concomitant risk factors on the association between smoking and subclinical atherosclerosis. Stroke 2009, 40, 1991–1998. [Google Scholar] [CrossRef] [PubMed]
- Dzubur, A.; Dzubur, A.; Mekic, M. Tobacco smoking and obesity as risk factors of polyvascular atherosclerosis. Med. Arch. 2009, 63, 90–93. [Google Scholar]
- Moran, A.E.; Roth, G.A.; Narula, J.; Mensah, G.A. 1990–2010 global cardiovascular disease atlas. Glob. Heart 2014, 9, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Tall, A.R. Plasma high density lipoproteins. Metabolism and relationship to atherogenesis. J. Clin. Investig. 1990, 86, 379–384. [Google Scholar] [CrossRef]
- Jonasson, L.; Holm, J.; Skalli, O.; Bondjers, G.; Hansson, G.K. Regional accumulations of T cells, macrophages, and smooth muscle cells in the human atherosclerotic plaque. Arteriosclerosis 1986, 6, 131–138. [Google Scholar] [CrossRef]
- Antohe, F. Endothelial cells and macrophages, partners in atherosclerotic plaque progression. Arch. Physiol. Biochem. 2006, 112, 245–253. [Google Scholar] [CrossRef]
- Libby, P. Inflammation in atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 2045–2051. [Google Scholar] [CrossRef]
- Tietge, U.J. Hyperlipidemia and cardiovascular disease: Inflammation, dyslipidemia, and atherosclerosis. Curr. Opin. Lipidol. 2014, 25, 94–95. [Google Scholar] [CrossRef] [PubMed]
- Hermansson, A.; Ketelhuth, D.F.; Strodthoff, D.; Wurm, M.; Hansson, E.M.; Nicoletti, A.; Paulsson-Berne, G.; Hansson, G.K. Inhibition of T cell response to native low-density lipoprotein reduces atherosclerosis. J. Exp. Med. 2010, 207, 1081–1093. [Google Scholar] [CrossRef] [PubMed]
- Abdolmaleki, F.; Gheibi Hayat, S.M.; Bianconi, V.; Johnston, T.P.; Sahebkar, A. Atherosclerosis and immunity: A perspective. Trends Cardiovasc. Med. 2018. [Google Scholar] [CrossRef] [PubMed]
- Hansson, G.K.; Libby, P. The immune response in atherosclerosis: A double-edged sword. Nat. Rev. Immunol. 2006, 6, 508. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, R.; Hansson, G.K. Treating inflammation in atherosclerotic cardiovascular disease: Emerging therapies. Eur. Heart J. 2009, 30, 2838–2844. [Google Scholar] [CrossRef]
- Chruściel, P.; Sahebkar, A.; Rembek-Wieliczko, M.; Serban, M.C.; Ursoniu, S.; Mikhailidis, D.P.; Jones, S.R.; Mosteoru, S.; Blaha, M.J.; Martin, S.S.; et al. Impact of statin therapy on plasma adiponectin concentrations: A systematic review and meta-analysis of 43 randomized controlled trial arms. Atherosclerosis 2016, 253, 194–208. [Google Scholar] [CrossRef] [PubMed]
- Parizadeh, S.M.R.; Azarpazhooh, M.R.; Moohebati, M.; Nematy, M.; Ghayour-Mobarhan, M.; Tavallaie, S.; Rahsepar, A.A.; Amini, M.; Sahebkar, A.; Mohammadi, M.; et al. Simvastatin therapy reduces prooxidant-antioxidant balance: Results of a placebo-controlled cross-over trial. Lipids 2011, 46, 333–340. [Google Scholar] [CrossRef]
- Sahebkar, A.; Kotani, K.; Serban, C.; Ursoniu, S.; Mikhailidis, D.P.; Jones, S.R.; Ray, K.K.; Blaha, M.J.; Rysz, J.; Toth, P.P.; et al. Statin therapy reduces plasma endothelin-1 concentrations: A meta-analysis of 15 randomized controlled trials. Atherosclerosis 2015, 241, 433–442. [Google Scholar] [CrossRef]
- Sahebkar, A.; Serban, C.; Mikhailidis, D.P.; Undas, A.; Lip, G.Y.H.; Muntner, P.; Bittner, V.; Ray, K.K.; Watts, G.F.; Hovingh, G.K.; et al. Association between statin use and plasma d-dimer levels: A systematic review and meta-analysis of randomised controlled trials. Thromb. Haemost. 2015, 114, 546–557. [Google Scholar] [CrossRef]
- Sahebkar, A.; Serban, C.; Ursoniu, S.; Mikhailidis, D.P.; Undas, A.; Lip, G.Y.H.; Bittner, V.; Ray, K.K.; Watts, G.F.; Kees Hovingh, G.; et al. The impact of statin therapy on plasma levels of von Willebrand factor antigen: Systematic review and meta-analysis of Randomised placebo-controlled trials. Thromb. Haemost. 2016, 115, 520–532. [Google Scholar] [CrossRef]
- Serban, C.; Sahebkar, A.; Ursoniu, S.; Mikhailidis, D.P.; Rizzo, M.; Lip, G.Y.H.; Kees Hovingh, G.; Kastelein, J.J.P.; Kalinowski, L.; Rysz, J.; et al. A systematic review and meta-analysis of the effect of statins on plasma asymmetric dimethylarginine concentrations. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Parsamanesh, N.; Moossavi, M.; Bahrami, A.; Fereidouni, M.; Barreto, G.; Sahebkar, A. NLRP3 inflammasome as a treatment target in atherosclerosis: A focus on statin therapy. Int. Immunopharmacol. 2019, 73, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Sahebkar, A.; Rathouska, J.; Simental-Mendía, L.E.; Nachtigal, P. Statin therapy and plasma cortisol concentrations: A systematic review and meta-analysis of randomized placebo-controlled trials. Pharmacol. Res. 2016, 103, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Ridker, P.M.; Everett, B.M.; Thuren, T.; MacFadyen, J.G.; Chang, W.H.; Ballantyne, C.; Fonseca, F.; Nicolau, J.; Koenig, W.; Anker, S.D.; et al. Antiinflammatory Therapy with Canakinumab for Atherosclerotic Disease. N. Engl. J. Med. 2017, 377, 1119–1131. [Google Scholar] [CrossRef] [PubMed]
- Fathollahi, A.; Gabalou, N.; Aslani, S. Mesenchymal stem cell transplantation in systemic lupus erythematous, a mesenchymal stem cell disorder. Lupus 2018, 27, 1053–1064. [Google Scholar] [CrossRef] [PubMed]
- Corsten, M.F.; Shah, K. Therapeutic stem-cells for cancer treatment: Hopes and hurdles in tactical warfare. Lancet Oncol. 2008, 9, 376–384. [Google Scholar] [CrossRef]
- Teo, A.K.; Vallier, L. Emerging use of stem cells in regenerative medicine. Biochem. J. 2010, 428, 11–23. [Google Scholar] [CrossRef]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef]
- Börger, V.; Bremer, M.; Ferrer-Tur, R.; Gockeln, L.; Stambouli, O.; Becic, A.; Giebel, B. Mesenchymal stem/stromal cell-derived extracellular vesicles and their potential as novel immunomodulatory therapeutic agents. Int. J. Mol. Sci. 2017, 18, 1450. [Google Scholar] [CrossRef]
- Li, J.; Xue, H.; Li, T.; Chu, X.; Xin, D.; Xiong, Y.; Qiu, W.; Gao, X.; Qian, M.; Xu, J. Exosomes derived from mesenchymal stem cells attenuate the progression of atherosclerosis in ApoE−/-mice via miR-let7 mediated infiltration and polarization of M2 macrophage. Biochem. Biophys. Res. Commun. 2019, 510, 565–572. [Google Scholar] [CrossRef]
- Friedenstein, A.; Piatetzky-Shapiro, I.; Petrakova, K. Osteogenesis in transplants of bone marrow cells. Development 1966, 16, 381–390. [Google Scholar]
- KV, P.; AI, K. Heterotopic of bone marrow. Analysis of precursor cells for osteogenic and hematopoietic tissues. Transplantation 1968, 6, 230–247. [Google Scholar]
- Mildmay-White, A.; Khan, W. Cell surface markers on adipose-derived stem cells: A systematic review. Curr. Stem Cell Res. Ther. 2017, 12, 484–492. [Google Scholar] [CrossRef] [PubMed]
- Mushahary, D.; Spittler, A.; Kasper, C.; Weber, V.; Charwat, V. Isolation, cultivation, and characterization of human mesenchymal stem cells. Cytom. Part A 2018, 93, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Kalervo Väänänen, H. Mesenchymal stem cells. Ann. Med. 2005, 37, 469–479. [Google Scholar] [CrossRef]
- Uccelli, A.; Moretta, L.; Pistoia, V. Mesenchymal stem cells in health and disease. Nat. Rev. Immunol. 2008, 8, 726. [Google Scholar] [CrossRef] [PubMed]
- Mortada, I.; Mortada, R. Epigenetic changes in mesenchymal stem cells differentiation. Eur. J. Med. Genet. 2018, 61, 114–118. [Google Scholar] [CrossRef]
- Caplan, A.I. Mesenchymal stem cells. J. Orthop. Res. 1991, 9, 641–650. [Google Scholar] [CrossRef]
- Bexell, D.; Scheding, S.; Bengzon, J. Toward brain tumor gene therapy using multipotent mesenchymal stromal cell vectors. Mol. Ther. 2010, 18, 1067–1075. [Google Scholar] [CrossRef]
- Samsonraj, R.M.; Raghunath, M.; Nurcombe, V.; Hui, J.H.; van Wijnen, A.J.; Cool, S.M. Concise review: Multifaceted characterization of human mesenchymal stem cells for use in regenerative medicine. Stem Cells Transl. Med. 2017, 6, 2173–2185. [Google Scholar] [CrossRef]
- Fiorina, P.; Jurewicz, M.; Augello, A.; Vergani, A.; Dada, S.; La Rosa, S.; Selig, M.; Godwin, J.; Law, K.; Placidi, C. Immunomodulatory function of bone marrow-derived mesenchymal stem cells in experimental autoimmune type 1 diabetes. J. Immunol. 2009, 183, 993–1004. [Google Scholar] [CrossRef]
- Nauta, A.J.; Fibbe, W.E. Immunomodulatory properties of mesenchymal stromal cells. Blood 2007, 110, 3499–3506. [Google Scholar] [CrossRef] [PubMed]
- Volarevic, V.; Gazdic, M.; Markovic, B.S.; Jovicic, N.; Djonov, V.; Arsenijevic, N. Mesenchymal stem cell-derived factors: Immuno-modulatory effects and therapeutic potential. Biofactors 2017, 43, 633–644. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.J.; Gage, F.H.; Weissman, I.L. Can stem cells cross lineage boundaries? Nat. Med. 2001, 7, 393–396. [Google Scholar] [CrossRef]
- Jiang, Y.; Jahagirdar, B.N.; Reinhardt, R.L.; Schwartz, R.E.; Keene, C.D.; Ortiz-Gonzalez, X.R.; Reyes, M.; Lenvik, T.; Lund, T.; Blackstad, M. Pluripotency of mesenchymal stem cells derived from adult marrow. Nature 2002, 418, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Bentzon, J.F.; Stenderup, K.; Hansen, F.; Schroder, H.; Abdallah, B.; Jensen, T.A.; Kassem, M. Tissue distribution and engraftment of human mesenchymal stem cells immortalized by human telomerase reverse transcriptase gene. Biochem. Biophys. Res. Commun. 2005, 330, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.M.; Glowacki, J. Age-related decline in the osteogenic potential of human bone marrow cells cultured in three-dimensional collagen sponges. J. Cell. Biochem. 2001, 82, 583–590. [Google Scholar] [CrossRef]
- Arutyunyan, I.; Elchaninov, A.; Makarov, A.; Fatkhudinov, T. Umbilical cord as prospective source for mesenchymal stem cell-based therapy. Stem Cells Int. 2016, 2016. [Google Scholar] [CrossRef]
- Cao, Y.; Gang, X.; Sun, C.; Wang, G. Mesenchymal stem cells improve healing of diabetic foot ulcer. J. Diabetes Res. 2017, 2017, 9328347. [Google Scholar] [CrossRef]
- Stewart, M.C.; Stewart, A.A. Mesenchymal stem cells: Characteristics, sources, and mechanisms of action. Vet. Clin. N. Am. Equine Pract. 2011, 27, 243–261. [Google Scholar] [CrossRef]
- Kern, S.; Eichler, H.; Stoeve, J.; Klüter, H.; Bieback, K. Comparative analysis of mesenchymal stem cells from bone marrow, umbilical cord blood, or adipose tissue. Stem Cells 2006, 24, 1294–1301. [Google Scholar] [CrossRef] [PubMed]
- Pierdomenico, L.; Bonsi, L.; Calvitti, M.; Rondelli, D.; Arpinati, M.; Chirumbolo, G.; Becchetti, E.; Marchionni, C.; Alviano, F.; Fossati, V. Multipotent mesenchymal stem cells with immunosuppressive activity can be easily isolated from dental pulp. Transplantation 2005, 80, 836–842. [Google Scholar] [CrossRef] [PubMed]
- Prindull, G.; Ben-Ishay, Z.; Ebell, W.; Bergholz, M.; Dirk, T.; Prindull, B. CFU-F circulating in cord blood. Ann. Hematol. 1987, 54, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Joerger-Messerli, M.S.; Marx, C.; Oppliger, B.; Mueller, M.; Surbek, D.V.; Schoeberlein, A. Mesenchymal stem cells from Wharton’s jelly and amniotic fluid. Best Pract. Res. Clin. Obstet. Gynaecol. 2016, 31, 30–44. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Wan, C.; Li, G. Concise review: Multipotent mesenchymal stromal cells in blood. Stem Cells 2007, 25, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.S.; Lee, J.; Lee, E.; Kwon, Y.S.; Lee, E.; Ahn, W.; Jiang, M.H.; Kim, J.C.; Son, Y. A new role of substance P as an injury-inducible messenger for mobilization of CD29+ stromal-like cells. Nat. Med. 2009, 15, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Spaeth, E.L.; Kidd, S.; Marini, F.C. Tracking inflammation-induced mobilization of mesenchymal stem cells. Stem Cell Mobil. Methods Protoc. 2012, 904, 173–190. [Google Scholar]
- Momin, E.N.; Vela, G.; Zaidi, H.A.; Quiñones-Hinojosa, A. The oncogenic potential of mesenchymal stem cells in the treatment of cancer: Directions for future research. Curr. Immunol. Rev. 2010, 6, 137–148. [Google Scholar] [CrossRef]
- Mishra, P.J.; Banerjee, D. Activation and Differentiation of Mesenchymal Stem Cells. In Signal Transduction Immunohistochemistry; Springer: Berlin/Heidelberg, Germany, 2017; pp. 201–209. [Google Scholar]
- Son, B.R.; Marquez-Curtis, L.A.; Kucia, M.; Wysoczynski, M.; Turner, A.R.; Ratajczak, J.; Ratajczak, M.Z.; Janowska-Wieczorek, A. Migration of bone marrow and cord blood mesenchymal stem cells in vitro is regulated by stromal-derived factor-1-CXCR4 and hepatocyte growth factor-c-met axes and involves matrix metalloproteinases. Stem Cells 2006, 24, 1254–1264. [Google Scholar] [CrossRef]
- Nakamizo, A.; Marini, F.; Amano, T.; Khan, A.; Studeny, M.; Gumin, J.; Chen, J.; Hentschel, S.; Vecil, G.; Dembinski, J. Human bone marrow–derived mesenchymal stem cells in the treatment of gliomas. Cancer Res. 2005, 65, 3307–3318. [Google Scholar] [CrossRef]
- Imitola, J.; Raddassi, K.; Park, K.I.; Mueller, F.J.; Nieto, M.; Teng, Y.D.; Frenkel, D.; Li, J.; Sidman, R.L.; Walsh, C.A. Directed migration of neural stem cells to sites of CNS injury by the stromal cell-derived factor 1α/CXC chemokine receptor 4 pathway. Proc. Natl. Acad. Sci. USA 2004, 101, 18117–18122. [Google Scholar] [CrossRef] [PubMed]
- Quante, M.; Tu, S.P.; Tomita, H.; Gonda, T.; Wang, S.S.; Takashi, S.; Baik, G.H.; Shibata, W.; DiPrete, B.; Betz, K.S. Bone marrow-derived myofibroblasts contribute to the mesenchymal stem cell niche and promote tumor growth. Cancer Cell 2011, 19, 257–272. [Google Scholar] [CrossRef] [PubMed]
- Schelker, R.C.; Iberl, S.; Müller, G.; Hart, C.; Herr, W.; Grassinger, J. TGF-β1 and CXCL12 modulate proliferation and chemotherapy sensitivity of acute myeloid leukemia cells co-cultured with multipotent mesenchymal stromal cells. Hematology 2018, 23, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Huang, W.; Chen, X.; Lian, Y.; Wang, J.; Cai, C.; Huang, L.; Wang, T.; Ren, J.; Xiang, A.P. CXCR5-overexpressing mesenchymal stromal cells exhibit enhanced homing and can decrease contact hypersensitivity. Mol. Ther. 2017, 25, 1434–1447. [Google Scholar] [CrossRef][Green Version]
- Smith, H.; Whittall, C.; Weksler, B.; Middleton, J. Chemokines stimulate bidirectional migration of human mesenchymal stem cells across bone marrow endothelial cells. Stem Cells Dev. 2011, 21, 476–486. [Google Scholar] [CrossRef]
- Liu, L.; Chen, J.X.; Zhang, X.W.; Sun, Q.; Yang, L.; Liu, A.; Hu, S.; Guo, F.; Liu, S.; Huang, Y. Chemokine receptor 7 overexpression promotes mesenchymal stem cell migration and proliferation via secreting Chemokine ligand 12. Sci. Rep. 2018, 8, 204. [Google Scholar] [CrossRef]
- Jagadeesha, D.K.; Takapoo, M.; Banfi, B.; Bhalla, R.C.; Miller, F.J. Nox1 transactivation of epidermal growth factor receptor promotes N-cadherin shedding and smooth muscle cell migration. Cardiovasc. Res. 2012, 93, 406–413. [Google Scholar] [CrossRef]
- Maretzky, T.; Evers, A.; Zhou, W.; Swendeman, S.L.; Wong, P.M.; Rafii, S.; Reiss, K.; Blobel, C.P. Migration of growth factor-stimulated epithelial and endothelial cells depends on EGFR transactivation by ADAM17. Nat. Commun. 2010, 2, 229. [Google Scholar] [CrossRef]
- Yahata, Y.; Shirakata, Y.; Tokumaru, S.; Yang, L.; Dai, X.; Tohyama, M.; Tsuda, T.; Sayama, K.; Iwai, M.; Horiuchi, M. A novel function of angiotensin II in skin wound healing Induction of fibroblast and keratinocyte migration by angiotensin II via heparin-binding epidermal growth factor (EGF)-like growth factor-mediated EGF receptor transactivation. J. Biol. Chem. 2006, 281, 13209–13216. [Google Scholar] [CrossRef]
- Rayego-Mateos, S.; Rodrigues-Diez, R.; Morgado-Pascual, J.L.; Valentijn, F.; Valdivielso, J.M.; Goldschmeding, R.; Ruiz-Ortega, M. Role of Epidermal Growth Factor Receptor (EGFR) and Its Ligands in Kidney Inflammation and Damage. Mediat. Inflamm. 2018, 2018. [Google Scholar] [CrossRef]
- Porcile, C.; Bajetto, A.; Barbieri, F.; Barbero, S.; Bonavia, R.; Biglieri, M.; Pirani, P.; Florio, T.; Schettini, G. Stromal cell-derived factor-1α (SDF-1α/CXCL12) stimulates ovarian cancer cell growth through the EGF receptor transactivation. Exp. Cell Res. 2005, 308, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Roelle, S.; Grosse, R.; Aigner, A.; Krell, H.W.; Czubayko, F.; Gudermann, T. Matrix metalloproteinases 2 and 9 mediate epidermal growth factor receptor transactivation by gonadotropin-releasing hormone. J. Biol. Chem. 2003, 278, 47307–47318. [Google Scholar] [CrossRef] [PubMed]
- Zuo, C.; Li, X.; Huang, J.; Chen, D.; Ji, K.; Yang, Y.; Xu, T.; Zhu, D.; Yan, C.; Gao, P. Osteoglycin attenuates cardiac fibrosis by suppressing cardiac myofibroblast proliferation and migration through antagonizing lysophosphatidic acid 3/matrix metalloproteinase 2/epidermal growth factor receptor signalling. Cardiovasc. Res. 2018, 114, 703–712. [Google Scholar] [CrossRef] [PubMed]
- De Becker, A.; Van Hummelen, P.; Bakkus, M.; Broek, I.V.; De Wever, J.; De Waele, M.; Van Riet, I. Migration of culture-expanded human mesenchymal stem cells through bone marrow endothelium is regulated by matrix metalloproteinase-2 and tissue inhibitor of metalloproteinase-3. Haematologica 2007, 92, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Merchant, N.; Nagaraju, G.P.; Rajitha, B.; Lammata, S.; Jella, K.K.; Buchwald, Z.S.; Lakka, S.S.; Ali, A.N. Matrix metalloproteinases: Their functional role in lung cancer. Carcinogenesis 2017, 38, 766–780. [Google Scholar] [CrossRef]
- Franco, C.; Patricia, H.R.; Timo, S.; Claudia, B.; Marcela, H. Matrix metalloproteinases as regulators of periodontal inflammation. Int. J. Mol. Sci. 2017, 18, 440. [Google Scholar] [CrossRef]
- Qiu, Y.; Marquez-Curtis, L.A.; Janowska-Wieczorek, A. Mesenchymal stromal cells derived from umbilical cord blood migrate in response to complement C1q. Cytotherapy 2012, 14, 285–295. [Google Scholar] [CrossRef]
- François, S.; Bensidhoum, M.; Mouiseddine, M.; Mazurier, C.; Allenet, B.; Semont, A.; Frick, J.; Saché, A.; Bouchet, S.; Thierry, D. Local irradiation not only induces homing of human mesenchymal stem cells at exposed sites but promotes their widespread engraftment to multiple organs: A study of their quantitative distribution after irradiation damage. Stem Cells 2006, 24, 1020–1029. [Google Scholar] [CrossRef]
- Bouchlaka, M.N.; Moffitt, A.B.; Kim, J.; Kink, J.A.; Bloom, D.D.; Love, C.; Dave, S.; Hematti, P.; Capitini, C.M. Human Mesenchymal Stem Cell–Educated Macrophages Are a Distinct High IL-6–Producing Subset that Confer Protection in Graft-versus-Host-Disease and Radiation Injury Models. Biol. Blood Marrow Transplant. 2017, 23, 897–905. [Google Scholar] [CrossRef]
- Di Nicola, M.; Carlo-Stella, C.; Magni, M.; Milanesi, M.; Longoni, P.D.; Matteucci, P.; Grisanti, S.; Gianni, A.M. Human bone marrow stromal cells suppress T-lymphocyte proliferation induced by cellular or nonspecific mitogenic stimuli. Blood 2002, 99, 3838–3843. [Google Scholar] [CrossRef]
- William, T.T.; Pendleton, J.D.; Beyer, W.M.; Egalka, M.C.; Guinan, E.C. Suppression of allogeneic T-cell proliferation by human marrow stromal cells: Implications in transplantation. Transplantation 2003, 75, 389–397. [Google Scholar]
- Ghosh, T.; Barik, S.; Bhuniya, A.; Dhar, J.; Dasgupta, S.; Ghosh, S.; Sarkar, M.; Guha, I.; Sarkar, K.; Chakrabarti, P. Tumor-associated mesenchymal stem cells inhibit naïve T cell expansion by blocking cysteine export from dendritic cells. Int. J. Cancer 2016, 139, 2068–2081. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, J.A.; Hettiaratchi, M.H.; McDevitt, T.C. Enhanced Immunosuppression of T Cells by Sustained Presentation of Bioactive Interferon-γ Within Three-Dimensional Mesenchymal Stem Cell Constructs. Stem Cells Transl. Med. 2017, 6, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Von Bubnoff, D.; Bieber, T. The indoleamine 2, 3-dioxygenase (IDO) pathway controls allergy. Allergy 2012, 67, 718–725. [Google Scholar] [CrossRef] [PubMed]
- Munn, D.H.; Mellor, A.L. Indoleamine 2, 3 dioxygenase and metabolic control of immune responses. Trends Immunol. 2013, 34, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Ge, W.; Li, C.; You, S.; Liao, L.; Han, Q.; Deng, W.; Zhao, R.C. Effects of mesenchymal stem cells on differentiation, maturation, and function of human monocyte-derived dendritic cells. Stem Cells Dev. 2004, 13, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Spaggiari, G.M.; Capobianco, A.; Abdelrazik, H.; Becchetti, F.; Mingari, M.C.; Moretta, L. Mesenchymal stem cells inhibit natural killer–cell proliferation, cytotoxicity, and cytokine production: Role of indoleamine 2, 3-dioxygenase and prostaglandin E2. Blood 2008, 111, 1327–1333. [Google Scholar] [CrossRef]
- Petri, R.M.; Hackel, A.; Hahnel, K.; Dumitru, C.A.; Bruderek, K.; Flohe, S.B.; Paschen, A.; Lang, S.; Brandau, S. Activated tissue-resident mesenchymal stromal cells regulate natural killer cell immune and tissue-regenerative function. Stem Cell Rep. 2017, 9, 985–998. [Google Scholar] [CrossRef]
- Aggarwal, S.; Pittenger, M.F. Human mesenchymal stem cells modulate allogeneic immune cell responses. Blood 2005, 105, 1815–1822. [Google Scholar] [CrossRef]
- Ghannam, S.; Pène, J.; Torcy-Moquet, G.; Jorgensen, C.; Yssel, H. Mesenchymal stem cells inhibit human Th17 cell differentiation and function and induce a T regulatory cell phenotype. J. Immunol. 2010, 185, 302–312. [Google Scholar] [CrossRef]
- Augello, A.; Tasso, R.; Negrini, S.M.; Amateis, A.; Indiveri, F.; Cancedda, R.; Pennesi, G. Bone marrow mesenchymal progenitor cells inhibit lymphocyte proliferation by activation of the programmed death 1 pathway. Eur. J. Immunol. 2005, 35, 1482–1490. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.H.; Pulin, A.A.; Seo, M.J.; Kota, D.J.; Ylostalo, J.; Larson, B.L.; Semprun-Prieto, L.; Delafontaine, P.; Prockop, D.J. Intravenous hMSCs improve myocardial infarction in mice because cells embolized in lung are activated to secrete the anti-inflammatory protein TSG-6. Cell Stem Cell 2009, 5, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Rashedi, I.; Gómez-Aristizábal, A.; Wang, X.H.; Viswanathan, S.; Keating, A. TLR3 or TLR4 activation enhances mesenchymal stromal cell-mediated Treg induction via notch signaling. Stem Cells 2017, 35, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Members:, A.T.F.; Perk, J.; De Backer, G.; Gohlke, H.; Graham, I.; Reiner, Ž.; Verschuren, M.; Albus, C.; Benlian, P.; Boysen, G. European Guidelines on cardiovascular disease prevention in clinical practice (version 2012) The Fifth Joint Task Force of the European Society of Cardiology and Other Societies on Cardiovascular Disease Prevention in Clinical Practice (constituted by representatives of nine societies and by invited experts) Developed with the special contribution of the European Association for Cardiovascular Prevention & Rehabilitation (EACPR). Eur. Heart J. 2012, 33, 1635–1701. [Google Scholar]
- Katakami, N. Mechanism of development of atherosclerosis and cardiovascular disease in diabetes mellitus. J. Atheroscler. Thromb. 2017. [Google Scholar] [CrossRef]
- Hurtubise, J.; McLellan, K.; Durr, K.; Onasanya, O.; Nwabuko, D.; Ndisang, J.F. The different facets of dyslipidemia and hypertension in atherosclerosis. Curr. Atheroscler. Rep. 2016, 18, 82. [Google Scholar] [CrossRef] [PubMed]
- Pirro, M.; Schillaci, G.; Savarese, G.; Gemelli, F.; Mannarino, M.R.; Siepi, D.; Bagaglia, F.; Mannarino, E. Attenuation of inflammation with short-term dietary intervention is associated with a reduction of arterial stiffness in subjects with hypercholesterolaemia. Eur. J. Cardiovasc. Prev. Rehabil. 2004, 11, 497–502. [Google Scholar] [CrossRef]
- Gisterå, A.; Hansson, G.K. The immunology of atherosclerosis. Nat. Rev. Nephrol. 2017, 13, 368. [Google Scholar] [CrossRef]
- Marchesi, S.; Lupattelli, G.; Lombardini, R.; Roscini, A.R.; Siepi, D.; Vaudo, G.; Pirro, M.; Sinzinger, H.; Schillaci, G.; Mannarino, E. Effects of fenofibrate on endothelial function and cell adhesion molecules during post-prandial lipemia in hypertriglyceridemia. J. Clin. Pharm. Ther. 2003, 28, 419–424. [Google Scholar] [CrossRef]
- Schillaci, G.; Pucci, G.; Pirro, M.; Monacelli, M.; Scarponi, A.M.; Manfredelli, M.R.; Rondelli, F.; Avenia, N.; Mannarino, E. Large-artery stiffness: A reversible marker of cardiovascular risk in primary hyperparathyroidism. Atherosclerosis 2011, 218, 96–101. [Google Scholar] [CrossRef]
- Hansson, G.K. Inflammation, atherosclerosis, and coronary artery disease. N. Engl. J. Med. 2005, 352, 1685–1695. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Cybulsky, M.I.; Gimbrone, M.A., Jr.; Libby, P. An atherogenic diet rapidly induces VCAM-1, a cytokine-regulatable mononuclear leukocyte adhesion molecule, in rabbit aortic endothelium. Arterioscler. Thromb. 1993, 13, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Wagner, D.D.; Frenette, P.S. The vessel wall and its interactions. Blood 2008, 111, 5271–5281. [Google Scholar] [CrossRef] [PubMed]
- Weber, C.; Zernecke, A.; Libby, P. The multifaceted contributions of leukocyte subsets to atherosclerosis: Lessons from mouse models. Nat. Rev. Immunol. 2008, 8, 802. [Google Scholar] [CrossRef] [PubMed]
- Chistiakov, D.A.; Melnichenko, A.A.; Myasoedova, V.A.; Grechko, A.V.; Orekhov, A.N. Mechanisms of foam cell formation in atherosclerosis. J. Mol. Med. 2017, 95, 1153–1165. [Google Scholar] [CrossRef]
- Mannarino, E.; Pirro, M. Endothelial injury and repair: A novel theory for atherosclerosis. Angiology 2008, 59, 69S–72S. [Google Scholar] [CrossRef]
- Emeson, E.E.; Shen, M.L.; Bell, C.; Qureshi, A. Inhibition of atherosclerosis in CD4 T-cell-ablated and nude (nu/nu) C57BL/6 hyperlipidemic mice. Am. J. Pathol. 1996, 149, 675–685. [Google Scholar]
- Frostegård, J.; Ulfgren, A.K.; Nyberg, P.; Hedin, U.; Swedenborg, J.; Andersson, U.; Hansson, G.K. Cytokine expression in advanced human atherosclerotic plaques: Dominance of pro-inflammatory (Th1) and macrophage-stimulating cytokines. Atherosclerosis 1999, 145, 33–43. [Google Scholar] [CrossRef]
- Zhou, X.; Paulsson, G.; Stemme, S.; Hansson, G.K. Hypercholesterolemia is associated with a T helper (Th) 1/Th2 switch of the autoimmune response in atherosclerotic apo E-knockout mice. J. Clin. Investig. 1998, 101, 1717–1725. [Google Scholar] [CrossRef]
- Voloshyna, I.; Littlefield, M.J.; Reiss, A.B. Atherosclerosis and interferon-γ: New insights and therapeutic targets. Trends Cardiovasc. Med. 2014, 24, 45–51. [Google Scholar] [CrossRef]
- Tournois, C.; Pignon, B.; Sevestre, M.A.; Al-Rifai, R.; Creuza, V.; Poitevin, G.; François, C.; Nguyen, P. Cell therapy in critical limb ischemia: A comprehensive analysis of two cell therapy products. Cytotherapy 2017, 19, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Stoltz, J.F.; Bensoussan, D.; Zhang, L.; Decot, V.; De Isla, N.; Li, Y.; Huselstein, C.; Benkirane-Jessel, N.; Li, N.; Reppel, L. Stem cells and applications: A survey. Bio Med. Mater. Eng. 2015, 25, 3–26. [Google Scholar]
- Wei, X.; Sun, G.; Zhao, X.; Wu, Q.; Chen, L.; Xu, Y.; Pang, X.; Qi, G. Human amnion mesenchymal stem cells attenuate atherosclerosis by modulating macrophage function to reduce immune response. Int. J. Mol. Med. 2019, 44, 1425–1435. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.X.; Wang, C.Q.; Li, X.Y.; Feng, G.K.; Zhu, H.L.; Ding, Y.; Jiang, X.J. Mesenchymal stem cells alleviate atherosclerosis by elevating number and function of CD4+ CD25+ FOXP3+ regulatory T-cells and inhibiting macrophage foam cell formation. Mol. Cell. Biochem. 2015, 400, 163–172. [Google Scholar] [CrossRef] [PubMed]
- English, K.; Ryan, J.; Tobin, L.; Murphy, M.; Barry, F.; Mahon, B.P. Cell contact, prostaglandin E2 and transforming growth factor beta 1 play non-redundant roles in human mesenchymal stem cell induction of CD4+ CD25Highforkhead box P3+ regulatory T cells. Clin. Exp. Immunol. 2009, 156, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Sotiropoulou, P.A.; Perez, S.A.; Gritzapis, A.D.; Baxevanis, C.N.; Papamichail, M. Interactions between human mesenchymal stem cells and natural killer cells. Stem Cells 2006, 24, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Frodermann, V.; Van Duijn, J.; Van Pel, M.; Van Santbrink, P.J.; Bot, I.; Kuiper, J.; De Jager, S.C. Mesenchymal stem cells reduce murine atherosclerosis development. Sci. Rep. 2015, 5, 15559. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Sun, W.; Wang, X.; Zhang, K.; Xi, W.; Gao, P. Skin-Derived Mesenchymal Stem Cells Alleviate Atherosclerosis via Modulating Macrophage Function. Stem Cells Transl. Med. 2015, 4, 1294–1301. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Zhou, S.; Zhou, T.; Su, H.; Pan, H.; Du, W.; Liu, B.; Liu, Q. Immuno-inflammatory regulation effect of mesenchymal stem cell transplantation in a rat model of myocardial infarction. Cytotherapy 2008, 10, 469–478. [Google Scholar] [CrossRef]
- Van Buul, G.; Villafuertes, E.; Bos, P.; Waarsing, J.; Kops, N.; Narcisi, R.; Weinans, H.; Verhaar, J.; Bernsen, M.; Van Osch, G. Mesenchymal stem cells secrete factors that inhibit inflammatory processes in short-term osteoarthritic synovium and cartilage explant culture. Osteoarthr. Cartil. 2012, 20, 1186–1196. [Google Scholar] [CrossRef]
- Jungebluth, P.; Luedde, M.; Ferrer, E.; Luedde, T.; Vucur, M.; Peinado, V.I.; Go, T.; Schreiber, C.; Von Richthofen, M.; Bader, A. Mesenchymal stem cells restore lung function by recruiting resident and nonresident proteins. Cell Transplant. 2011, 20, 1561–1574. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.S.; Hu, S.W.; Zhang, Q.H.; Xia, A.X.; Jiang, Z.X.; Chen, X.M. Mesenchymal stem cells stabilize atherosclerotic vulnerable plaque by anti-inflammatory properties. PLoS ONE 2015, 10, e0136026. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.M.; Du, D.Y.; Li, Y.T.; Ge, X.L.; Qin, P.T.; Zhang, Q.H.; Liu, Y. Allogeneic bone marrow mesenchymal stem cells transplantation for stabilizing and repairing of atherosclerotic ruptured plaque. Thromb. Res. 2013, 131, e253–e257. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.L.; Yet, S.F.; Hsu, Y.T.; Wang, G.J.; Hung, S.C. Mesenchymal stem cells ameliorate atherosclerotic lesions via restoring endothelial function. Stem Cells Transl. Med. 2015, 4, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Kawi, S.H.; Hashem, K.S. Possible therapeutic effect of stem cell in atherosclerosis in albino rats. A histological and immunohistochemical study. Int. J. Stem Cells 2015, 8, 200. [Google Scholar] [CrossRef] [PubMed]
- Prockop, D.J.; Oh, J.Y. Mesenchymal stem/stromal cells (MSCs): Role as guardians of inflammation. Mol. Ther. 2012, 20, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; Kanzler, H.; Soumelis, V.; Gilliet, M. Dendritic cell lineage, plasticity and cross-regulation. Nat. Immunol. 2001, 2, 585. [Google Scholar] [CrossRef]
- Baratelli, F.; Lin, Y.; Zhu, L.; Yang, S.C.; Heuzé-Vourc’h, N.; Zeng, G.; Reckamp, K.; Dohadwala, M.; Sharma, S.; Dubinett, S.M. Prostaglandin E2 induces FOXP3 gene expression and T regulatory cell function in human CD4+ T cells. J. Immunol. 2005, 175, 1483–1490. [Google Scholar] [CrossRef]
- Choi, H.; Lee, R.H.; Bazhanov, N.; Oh, J.Y.; Prockop, D.J. Anti-inflammatory protein TSG-6 secreted by activated MSCs attenuates zymosan-induced mouse peritonitis by decreasing TLR2/NF-κB signaling in resident macrophages. Blood 2011, 118, 330–338. [Google Scholar] [CrossRef]
- Wisniewski, H.G.; Maier, R.; Lotz, M.; Lee, S.; Klampfer, L.; Lee, T.; Vilcek, J. TSG-6: A TNF-, IL-1-, and LPS-inducible secreted glycoprotein associated with arthritis. J. Immunol. 1993, 151, 6593–6601. [Google Scholar]
- Wisniewski, H.G.; Hua, J.; Poppers, D.M.; Naime, D.; Vilcek, J.; Cronstein, B.N. TNF/IL-1-inducible protein TSG-6 potentiates plasmin inhibition by inter-alpha-inhibitor and exerts a strong anti-inflammatory effect in vivo. J. Immunol. 1996, 156, 1609–1615. [Google Scholar] [PubMed]
- Cao, T.V.; La, M.; Getting, S.J.; Day, A.J.; Perretti, M. Inhibitory effects of TSG-6 link module on leukocyte–endothelial cell interactions in vitro and in vivo. Microcirculation 2004, 11, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Lüscher, T.; Barton, M. Biology of the endothelium. Clin. Cardiol. 1997, 20, II-3-10. [Google Scholar] [PubMed]
- Forstermann, U.; Munzel, T. Endothelial nitric oxide synthase in vascular disease: From marvel to menace. Circulation 2006, 113, 1708–1714. [Google Scholar] [CrossRef] [PubMed]
- Félétou, M. The endothelium, Part I: Multiple functions of the endothelial cells—Focus on endothelium-derived vasoactive mediators. In Colloquium Series on Integrated Systems Physiology: From Molecule to Function; Morgan & Claypool Life Sciences Publishers: London, UK, 2009–2011; pp. 1–306.
- De Caterina, R.; Libby, P.; Peng, H.B.; Thannickal, V.J.; Rajavashisth, T.; Gimbrone, M.A.; Shin, W.S.; Liao, J.K. Nitric oxide decreases cytokine-induced endothelial activation. Nitric oxide selectively reduces endothelial expression of adhesion molecules and proinflammatory cytokines. J. Clin. Investig. 1995, 96, 60–68. [Google Scholar] [CrossRef] [PubMed]
- König, J.; Huppertz, B.; Desoye, G.; Parolini, O.; Fröhlich, J.D.; Weiss, G.; Dohr, G.; Sedlmayr, P.; Lang, I. Amnion-derived mesenchymal stromal cells show angiogenic properties but resist differentiation into mature endothelial cells. Stem Cells Dev. 2011, 21, 1309–1320. [Google Scholar] [CrossRef] [PubMed]
- Fu, R.; Wang, Q.; Guo, Q.; Xu, J.; Wu, X. XJP-1 protects endothelial cells from oxidized low-density lipoprotein-induced apoptosis by inhibiting NADPH oxidase subunit expression and modulating the PI3K/Akt/eNOS pathway. Vasc. Pharmacol. 2013, 58, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Chu, L.; Hao, H.; Luo, M.; Huang, Y.; Chen, Z.; Lu, T.; Zhao, X.; Verfaillie, C.M.; Zweier, J.L.; Liu, Z. Ox-LDL modifies the behaviour of bone marrow stem cells and impairs their endothelial differentiation via inhibition of Akt phosphorylation. J. Cell. Mol. Med. 2011, 15, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Guo, X.; Chen, S.Y. Function and therapeutic potential of mesenchymal stem cells in atherosclerosis. Front. Cardiovasc. Med. 2017, 4, 32. [Google Scholar] [CrossRef] [PubMed]
- Azimi, M.; Aslani, S.; Mortezagholi, S.; Salek, A.; Javan, M.R.; Rezaiemanesh, A.; Ghaedi, M.; Gholamzad, M.; Salehi, E. Identification, isolation, and functional assay of regulatory T cells. Immunol. Investig. 2016, 45, 584–602. [Google Scholar] [CrossRef] [PubMed]
- Hori, S.; Nomura, T.; Sakaguchi, S. Control of regulatory T cell development by the transcription factor Foxp3. Science 2003, 299, 1057–1061. [Google Scholar] [CrossRef] [PubMed]
- Shevach, E.M. From vanilla to 28 flavors: Multiple varieties of T regulatory cells. Immunity 2006, 25, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Chistiakov, D.A.; Sobenin, I.A.; Orekhov, A.N. Regulatory T cells in atherosclerosis and strategies to induce the endogenous atheroprotective immune response. Immunol. Lett. 2013, 151, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Sardella, G.; De Luca, L.; Francavilla, V.; Accapezzato, D.; Mancone, M.; Sirinian, M.I.; Fedele, F.; Paroli, M. Frequency of naturally-occurring regulatory T cells is reduced in patients with ST-segment elevation myocardial infarction. Thromb. Res. 2007, 120, 631–634. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, R.; Gerdes, N.; Badeau, R.M.; Gisterå, A.; Strodthoff, D.; Ketelhuth, D.F.; Lundberg, A.M.; Rudling, M.; Nilsson, S.K.; Olivecrona, G. Depletion of FOXP3+ regulatory T cells promotes hypercholesterolemia and atherosclerosis. J. Clin. Investig. 2013, 123, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Ait-Oufella, H.; Salomon, B.L.; Potteaux, S.; Robertson, A.K.L.; Gourdy, P.; Zoll, J.; Merval, R.; Esposito, B.; Cohen, J.L.; Fisson, S. Natural regulatory T cells control the development of atherosclerosis in mice. Nat. Med. 2006, 12, 178. [Google Scholar] [CrossRef]
- Mallat, Z.; Besnard, S.; Duriez, M.; Deleuze, V.; Emmanuel, F.; Bureau, M.F.; Soubrier, F.; Esposito, B.; Duez, H.; Fievet, C. Protective role of interleukin-10 in atherosclerosis. Circ. Res. 1999, 85, e17–e24. [Google Scholar] [CrossRef] [PubMed]
- Pinderski, L.J.; Fischbein, M.P.; Subbanagounder, G.; Fishbein, M.C.; Kubo, N.; Cheroutre, H.; Curtiss, L.K.; Berliner, J.A.; Boisvert, W.A. Overexpression of interleukin-10 by activated T lymphocytes inhibits atherosclerosis in LDL receptor–deficient mice by altering lymphocyte and macrophage phenotypes. Circ. Res. 2002, 90, 1064–1071. [Google Scholar] [CrossRef]
- Robertson, A.K.L.; Rudling, M.; Zhou, X.; Gorelik, L.; Flavell, R.A.; Hansson, G.K. Disruption of TGF-β signaling in T cells accelerates atherosclerosis. J. Clin. Investig. 2003, 112, 1342–1350. [Google Scholar] [CrossRef]
- Gojova, A.; Brun, V.; Esposito, B.; Cottrez, F.; Gourdy, P.; Ardouin, P.; Tedgui, A.; Mallat, Z.; Groux, H. Specific abrogation of transforming growth factor-β signaling in T cells alters atherosclerotic lesion size and composition in mice. Blood 2003, 102, 4052–4058. [Google Scholar] [CrossRef][Green Version]
- Foks, A.; Frodermann, V.; ter Borg, M.; Habets, K.; Bot, I.; Zhao, Y.; van Eck, M.; van Berkel, T.J.; Kuiper, J.; van Puijvelde, G. Differential effects of regulatory T cells on the initiation and regression of atherosclerosis. Atherosclerosis 2011, 218, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Melief, S.M.; Schrama, E.; Brugman, M.H.; Tiemessen, M.M.; Hoogduijn, M.J.; Fibbe, W.E.; Roelofs, H. Multipotent stromal cells induce human regulatory T cells through a novel pathway involving skewing of monocytes toward anti-inflammatory macrophages. Stem Cells 2013, 31, 1980–1991. [Google Scholar] [CrossRef] [PubMed]
- Cahill, E.F.; Tobin, L.M.; Carty, F.; Mahon, B.P.; English, K. Jagged-1 is required for the expansion of CD4+ CD25+ FoxP3+ regulatory T cells and tolerogenic dendritic cells by murine mesenchymal stromal cells. Stem Cell Res. Ther. 2015, 6, 19. [Google Scholar] [CrossRef] [PubMed]
- Alhadlaq, A.; Mao, J.J. Mesenchymal stem cells: Isolation and therapeutics. Stem Cells Dev. 2004, 13, 436–448. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.R.; Zanella, C.A.; Zucchi, F.C.; Dombroski, T.C.; Costa, E.T.; Guethe, L.M.; Oliveira, A.O.; Donatti, A.L.; Neder, L.; Chimelli, L. Immunolocalization of nitric oxide synthase isoforms in human archival and rat tissues, and cultured cells. J. Neurosci. Methods 2011, 198, 16–22. [Google Scholar] [CrossRef] [PubMed]
- De Boer, H.; Verseyden, C.; Ulfman, L.; Zwaginga, J.; Bot, I.; Biessen, E.; Rabelink, T.; Van Zonneveld, A. Fibrin and activated platelets cooperatively guide stem cells to a vascular injury and promote differentiation towards an endothelial cell phenotype. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 1653–1659. [Google Scholar] [CrossRef] [PubMed]
- Babaev, V.R.; Fazio, S.; Gleaves, L.A.; Carter, K.J.; Semenkovich, C.F.; Linton, M.F. Macrophage lipoprotein lipase promotes foam cell formation and atherosclerosis in vivo. J. Clin. Investig. 1999, 103, 1697–1705. [Google Scholar] [CrossRef]
- Van Eck, M.; Zimmermann, R.; Groot, P.H.; Zechner, R.; Van Berkel, T.J. Role of macrophage-derived lipoprotein lipase in lipoprotein metabolism and atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2000, 20, e53–e62. [Google Scholar] [CrossRef]
- Chin-Dusting, J.P.; Shaw, J.A. Lipids and atherosclerosis: Clinical management of hypercholesterolaemia. Expert Opin. Pharmacother. 2001, 2, 419–430. [Google Scholar] [CrossRef]
- Reardon, C.A.; Blachowicz, L.; Lukens, J.; Nissenbaum, M.; Getz, G.S. Genetic background selectively influences innominate artery atherosclerosis: Immune system deficiency as a probe. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 1449–1454. [Google Scholar] [CrossRef]
- Bartolomé, N.; Arteta, B.; Martínez, M.J.; Chico, Y.; Ochoa, B. Kupffer cell products and interleukin 1? directly promote VLDL secretion and apoB mRNA up-regulation in rodent hepatocytes. Innate Immun. 2008, 14, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Ruan, H.; Miles, P.D.; Ladd, C.M.; Ross, K.; Golub, T.R.; Olefsky, J.M.; Lodish, H.F. Profiling gene transcription in vivo reveals adipose tissue as an immediate target of tumor necrosis factor-α: Implications for insulin resistance. Diabetes 2002, 51, 3176–3188. [Google Scholar] [CrossRef]
- Von Der Thusen, J.H.; Kuiper, J.; Fekkes, M.L.; de Vos, P.; van Berkel, T.J.; Biessen, E.A. Attenuation of atherogenesis by systemic and local adenovirus-mediated gene transfer of interleukin-10 in LDLr−/− mice. FASEB J. 2001, 15, 2730–2732. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Liang, M.; Chen, W.; Sun, X.; Wang, X.; Li, C.; Yang, Y.; Yang, Z.; Zeng, W. Human induced pluripotent stem cell-derived mesenchymal stem cells alleviate atherosclerosis by modulating inflammatory responses. Mol. Med. Rep. 2018, 17, 1461–1468. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Z.; Cao, T.H.; Han, J.C.; Qu, H.; Jiang, S.Q.; Xie, B.D.; Yan, X.L.; Wu, H.; Liu, X.L.; Zhang, F. Comparison of adipose-and bone marrow-derived stem cells in protecting against ox-LDL-induced inflammation in M1-macrophage-derived foam cells. Mol. Med. Rep. 2019, 19, 2660–2670. [Google Scholar] [CrossRef]
- Silvestre-Roig, C.; de Winther, M.P.; Weber, C.; Daemen, M.J.; Lutgens, E.; Soehnlein, O. Atherosclerotic plaque destabilization: Mechanisms, models, and therapeutic strategies. Circ. Res. 2014, 114, 214–226. [Google Scholar] [CrossRef]
- Virmani, R.; Kolodgie, F.D.; Burke, A.P.; Farb, A.; Schwartz, S.M. Lessons from sudden coronary death: A comprehensive morphological classification scheme for atherosclerotic lesions. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 1262–1275. [Google Scholar] [CrossRef]
- Spagnoli, L.G.; Mauriello, A.; Sangiorgi, G.; Fratoni, S.; Bonanno, E.; Schwartz, R.S.; Piepgras, D.G.; Pistolese, R.; Ippoliti, A.; Holmes, D.R. Extracranial thrombotically active carotid plaque as a risk factor for ischemic stroke. JAMA 2004, 292, 1845–1852. [Google Scholar] [CrossRef] [PubMed]
- Burke, A.P.; Farb, A.; Malcom, G.T.; Liang, Y.H.; Smialek, J.; Virmani, R. Coronary risk factors and plaque morphology in men with coronary disease who died suddenly. N. Engl. J. Med. 1997, 336, 1276–1282. [Google Scholar] [CrossRef]
- Redgrave, J.; Lovett, J.; Gallagher, P.; Rothwell, P. Histological assessment of 526 symptomatic carotid plaques in relation to the nature and timing of ischemic symptoms: The Oxford plaque study. Circulation 2006, 113, 2320–2328. [Google Scholar] [CrossRef]
- Guo, S.; Meng, S.; Chen, B.; Liu, J.; Gao, L.; Wu, Y. C-reactive protein can influence the proliferation, apoptosis, and monocyte chemotactic protein-1 production of human umbilical vein endothelial cells. DNA Cell Biol. 2011, 30, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Forte, L.; Cimmino, G.; Loffredo, F.; De Palma, R.; Abbate, G.; Calabrò, P.; Ingrosso, D.; Galletti, P.; Carangio, C.; Casillo, B. C-reactive protein is released in the coronary circulation and causes endothelial dysfunction in patients with acute coronary syndromes. Int. J. Cardiol. 2011, 152, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Haverkate, F. Levels of haemostatic factors, arteriosclerosis and cardiovascular disease. Vasc. Pharmacol. 2002, 39, 109–112. [Google Scholar] [CrossRef]
- Kleemann, R.; Zadelaar, S.; Kooistra, T. Cytokines and atherosclerosis: A comprehensive review of studies in mice. Cardiovasc. Res. 2008, 79, 360–376. [Google Scholar] [CrossRef] [PubMed]
- Heeschen, C.; Dimmeler, S.; Hamm, C.W.; Fichtlscherer, S.; Boersma, E.; Simoons, M.L.; Zeiher, A.M. Serum level of the antiinflammatory cytokine interleukin-10 is an important prognostic determinant in patients with acute coronary syndromes. Circulation 2003, 107, 2109–2114. [Google Scholar] [CrossRef] [PubMed]
- Potteaux, S.; Esposito, B.; van Oostrom, O.; Brun, V.; Ardouin, P.; Groux, H.; Tedgui, A.; Mallat, Z. Leukocyte-derived interleukin 10 is required for protection against atherosclerosis in low-density lipoprotein receptor knockout mice. Arterioscler. Thromb. Vasc. Boil. 2004, 24, 1474–1478. [Google Scholar] [CrossRef] [PubMed]
- Rössig, L.; Dimmeler, S.; Zeiher, A.M. Apoptosis in the vascular wall and atherosclerosis. Basic Res. Cardiol. 2001, 96, 11–22. [Google Scholar] [CrossRef]
- Mallat, Z.; Tedgui, A. Apoptosis in the vasculature: Mechanisms and functional importance. Br. J. Pharmacol. 2000, 130, 947–962. [Google Scholar] [CrossRef]
- Li, Y.; Gerbod-Giannone, M.C.; Seitz, H.; Cui, D.; Thorp, E.; Tall, A.R.; Matsushima, G.K.; Tabas, I. Cholesterol-induced apoptotic macrophages elicit an inflammatory response in phagocytes, which is partially attenuated by the Mer receptor. J. Biol. Chem. 2006, 281, 6707–6717. [Google Scholar] [CrossRef]
- Li, X.; Sun, W.; Xi, W.; Shen, W.; Wei, T.; Chen, W.; Gao, P.; Li, Q. Transplantation of skin mesenchymal stem cells attenuated AngII-induced hypertension and vascular injury. Biochem. Biophys. Res. Commun. 2018, 497, 1068–1075. [Google Scholar] [CrossRef]
- Reinders, M.E.; de Fijter, J.W.; Roelofs, H.; Bajema, I.M.; de Vries, D.K.; Schaapherder, A.F.; Claas, F.H.; van Miert, P.P.; Roelen, D.L.; van Kooten, C. Autologous bone marrow-derived mesenchymal stromal cells for the treatment of allograft rejection after renal transplantation: Results of a phase I study. Stem Cells Transl. Med. 2013, 2, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Ke, M.; Xu, L.; Liu, L.; Chen, X.; Xia, W.; Li, X.; Chen, Z.; Ma, J.; Liao, D. Donor-derived mesenchymal stem cells combined with low-dose tacrolimus prevent acute rejection after renal transplantation: A clinical pilot study. Transplantation 2013, 95, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Mudrabettu, C.; Kumar, V.; Rakha, A.; Yadav, A.K.; Ramachandran, R.; Kanwar, D.B.; Nada, R.; Minz, M.; Sakhuja, V.; Marwaha, N. Safety and efficacy of autologous mesenchymal stromal cells transplantation in patients undergoing living donor kidney transplantation: A pilot study. Nephrology 2015, 20, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Subbanna, P.K.T. Mesenchymal stem cells for treating GVHD: In-vivo fate and optimal dose. Med. Hypotheses 2007, 69, 469–470. [Google Scholar] [CrossRef] [PubMed]
- Koç, O.N.; Gerson, S.L.; Cooper, B.W.; Dyhouse, S.M.; Haynesworth, S.E.; Caplan, A.I.; Lazarus, H.M. Rapid hematopoietic recovery after coinfusion of autologous-blood stem cells and culture-expanded marrow mesenchymal stem cells in advanced breast cancer patients receiving high-dose chemotherapy. J. Clin. Oncol. 2000, 18, 307. [Google Scholar] [CrossRef]
- Miura, M.; Miura, Y.; Padilla-Nash, H.M.; Molinolo, A.A.; Fu, B.; Patel, V.; Seo, B.M.; Sonoyama, W.; Zheng, J.J.; Baker, C.C. Accumulated chromosomal instability in murine bone marrow mesenchymal stem cells leads to malignant transformation. Stem Cells 2006, 24, 1095–1103. [Google Scholar] [CrossRef] [PubMed]
- Houghton, J.; Stoicov, C.; Nomura, S.; Rogers, A.B.; Carlson, J.; Li, H.; Cai, X.; Fox, J.G.; Goldenring, J.R.; Wang, T.C. Gastric cancer originating from bone marrow-derived cells. Science 2004, 306, 1568–1571. [Google Scholar] [CrossRef]
- Barkholt, L.; Flory, E.; Jekerle, V.; Lucas-Samuel, S.; Ahnert, P.; Bisset, L.; Büscher, D.; Fibbe, W.; Foussat, A.; Kwa, M. Risk of tumorigenicity in mesenchymal stromal cell–based therapies—Bridging scientific observations and regulatory viewpoints. Cytotherapy 2013, 15, 753–759. [Google Scholar] [CrossRef]
- Liao, J.; Chen, X.; Li, Y.; Ge, Z.; Duan, H.; Zou, Y.; Ge, J. Transfer of bone-marrow-derived mesenchymal stem cells influences vascular remodeling and calcification after balloon injury in hyperlipidemic rats. BioMed Res. Int. 2012, 2012, 165296. [Google Scholar] [CrossRef]
- Drela, K.; Stanaszek, L.; Nowakowski, A.; Kuczynska, Z.; Lukomska, B. Experimental strategies of mesenchymal stem cell propagation: Adverse events and potential risk of functional changes. Stem Cells Int. 2019, 2019, 7012692. [Google Scholar] [CrossRef]
- Huselstein, C.; Rahouadj, R.; De Isla, N.; Bensoussan, D.; Stoltz, J.F.; Li, Y. Mechanobiology of mesenchymal stem cells: Which interest for cell-based treatment? Bio Med. Mater. Eng. 2017, 28, S47–S56. [Google Scholar] [CrossRef] [PubMed]
- Benayahu, D.; Wiesenfeld, Y.; Sapir-Koren, R. How is mechanobiology involved in mesenchymal stem cell differentiation toward the osteoblastic or adipogenic fate? J. Cell. Physiol. 2019, 234, 12133–12141. [Google Scholar] [CrossRef] [PubMed]
- Stanton, A.E.; Tong, X.; Yang, F. Extracellular matrix type modulates mechanotransduction of stem cells. Acta Biomater. 2019, 96, 310–320. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Animal Model | Cell Source | Administration Route | Consequence | Reference |
|---|---|---|---|---|
| New Zealand rabbits | Bone marrow | Intravenous | Increased: TSG-6, IL-10, hs-CRP, TNF-α, IL-6, NF-κB Decreased: Apoptosis, MMPs | [123] |
| New Zealand rabbits | Bone marrow | Intra-arterial | Increased: Collagen fibers Decreased: MMPs, PAI-1, hs-CRP | [124] |
| ApoE−/− mice | Bone marrow | Intravenous | Increased: Tregs Decreased: SRA, CD36 | [115] |
| ApoE−/− mice | Bone marrow | Intravenous | Increased: eNOS, IL8, MIP-2 | [125] |
| ApoE−/− mice | Skin | Intravenous | Increased: IL-10, PGE2 Decreased: NF-κB, TNF-α | [119] |
| LDLR−/− mice | Bone marrow | Intravenous | Increased: Tregs Decreased: CD4+ T cells, CCL2, IFN-γ, monocytes, TNF-α, serum cholesterol | [118] |
| Albino rats | Cord blood | Intravenous | Increased: iNOS | [126] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahdavi Gorabi, A.; Banach, M.; Reiner, Ž.; Pirro, M.; Hajighasemi, S.; Johnston, T.P.; Sahebkar, A. The Role of Mesenchymal Stem Cells in Atherosclerosis: Prospects for Therapy via the Modulation of Inflammatory Milieu. J. Clin. Med. 2019, 8, 1413. https://doi.org/10.3390/jcm8091413
Mahdavi Gorabi A, Banach M, Reiner Ž, Pirro M, Hajighasemi S, Johnston TP, Sahebkar A. The Role of Mesenchymal Stem Cells in Atherosclerosis: Prospects for Therapy via the Modulation of Inflammatory Milieu. Journal of Clinical Medicine. 2019; 8(9):1413. https://doi.org/10.3390/jcm8091413
Chicago/Turabian StyleMahdavi Gorabi, Armita, Maciej Banach, Željko Reiner, Matteo Pirro, Saeideh Hajighasemi, Thomas P. Johnston, and Amirhossein Sahebkar. 2019. "The Role of Mesenchymal Stem Cells in Atherosclerosis: Prospects for Therapy via the Modulation of Inflammatory Milieu" Journal of Clinical Medicine 8, no. 9: 1413. https://doi.org/10.3390/jcm8091413
APA StyleMahdavi Gorabi, A., Banach, M., Reiner, Ž., Pirro, M., Hajighasemi, S., Johnston, T. P., & Sahebkar, A. (2019). The Role of Mesenchymal Stem Cells in Atherosclerosis: Prospects for Therapy via the Modulation of Inflammatory Milieu. Journal of Clinical Medicine, 8(9), 1413. https://doi.org/10.3390/jcm8091413