SEPT14 Mutations and Teratozoospermia: Genetic Effects on Sperm Head Morphology and DNA Integrity


Abstract
:1. Introduction
1.1. Male Infertility and Sperm DNA Damage
1.2. Septins
1.3. SEPT14
2. Materials and Methods
2.1. Participants and Semen Analysis
2.2. Sperm Genomic DNA Extraction, Polymerase Chain Reaction, and Sequencing
2.3. Transmission Electron Microscopy
2.4. Toluidine Blue Staining
2.5. Comet Assay
2.6. Cloning, Transfection, and Immunofluorescence Staining
2.7. Statistical Analysis
3. Results
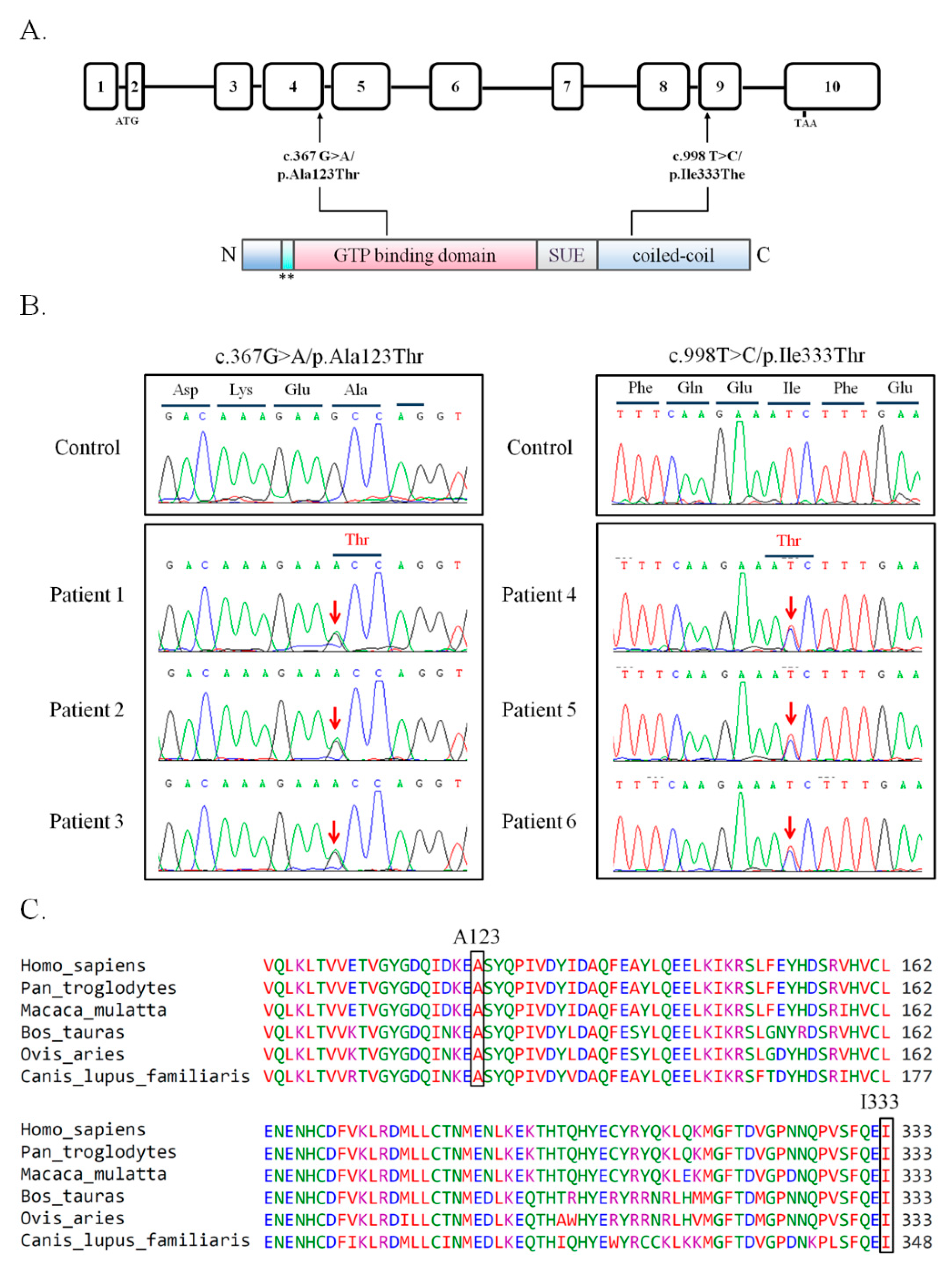
3.1. Two Novel Missense Mutations in the SEPT14 Gene
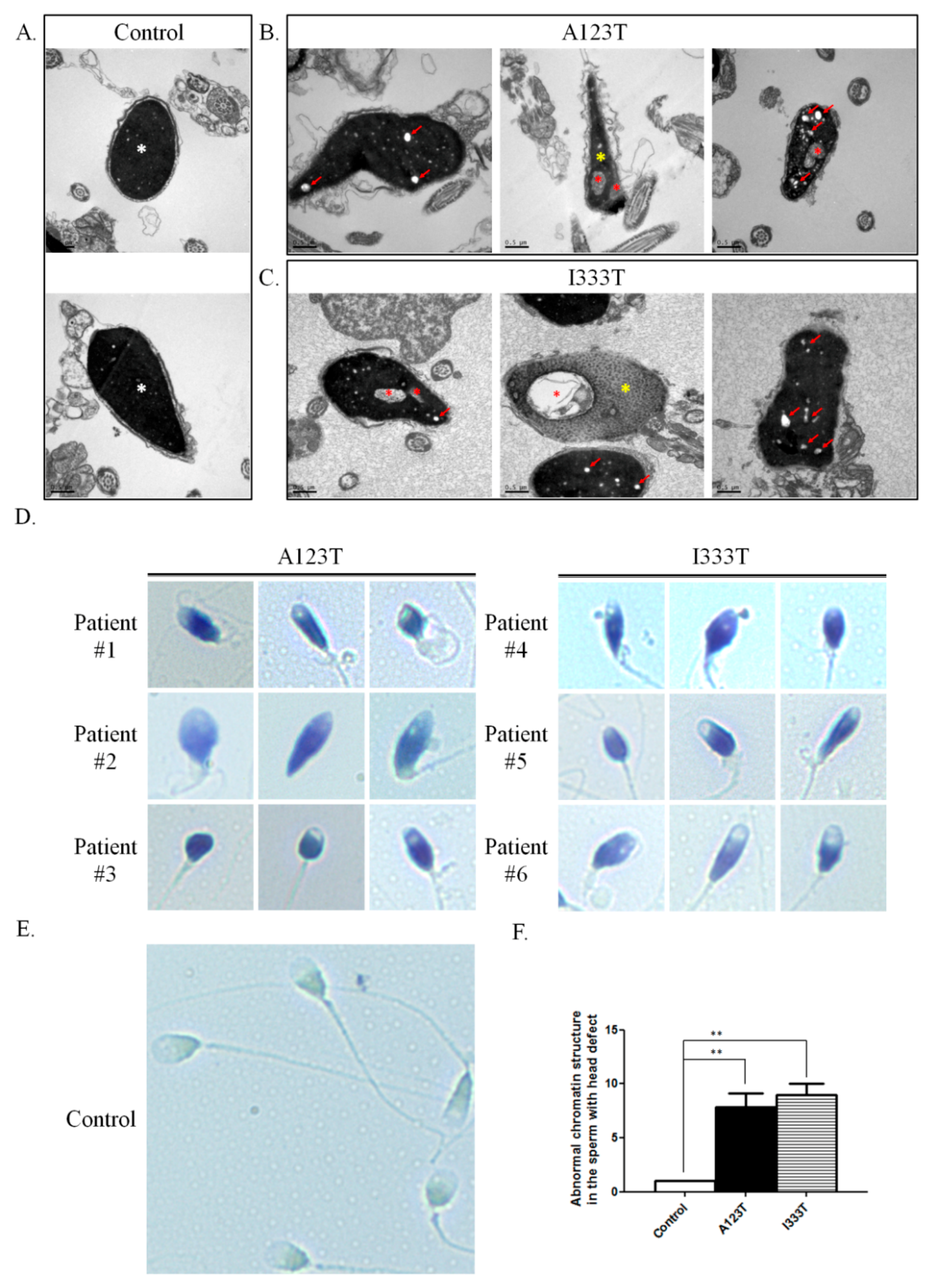
3.2. Severely Malformed Heads and Diminished Chromatin Integrity in Sperm with SEPT14 Mutations
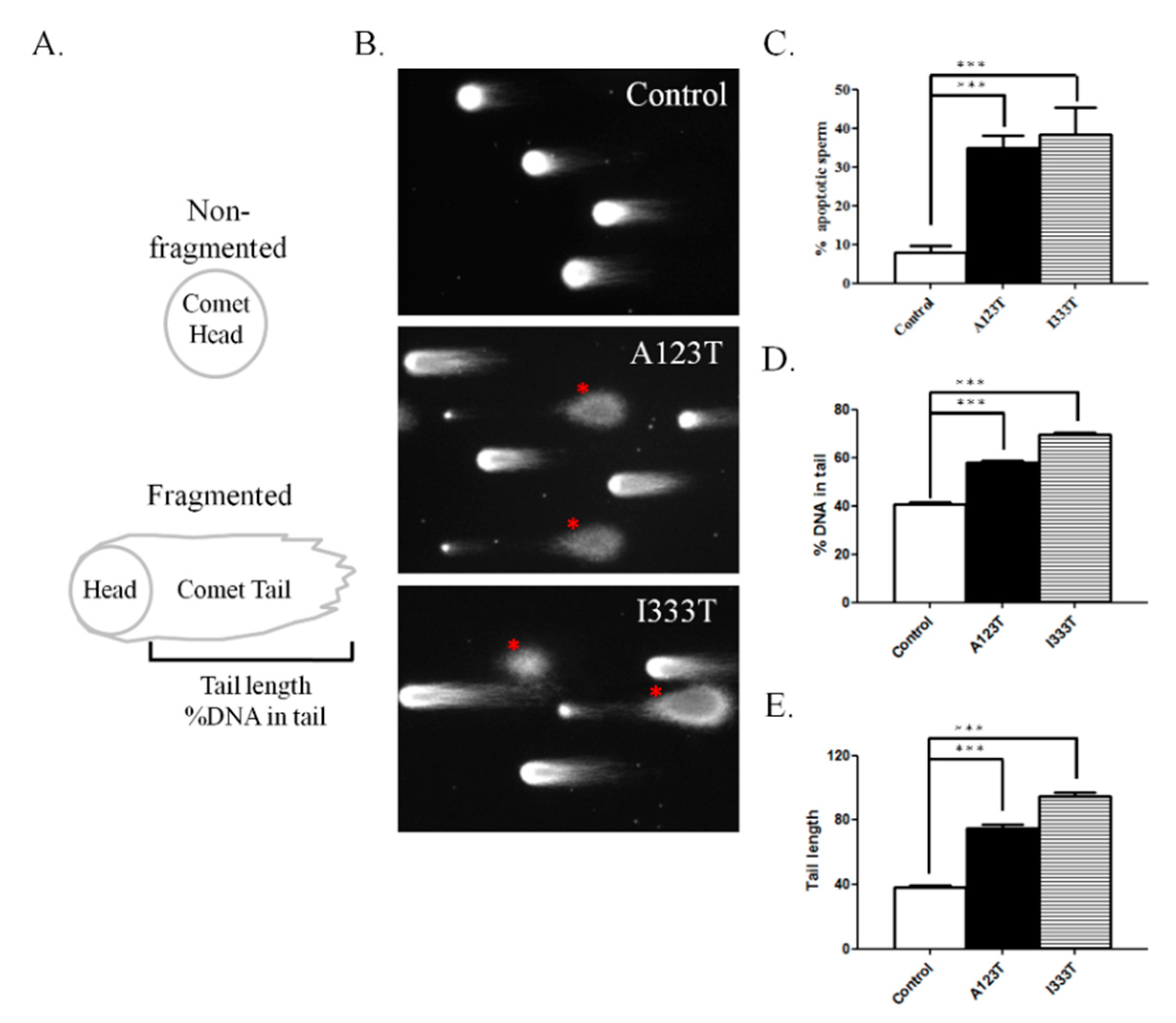
3.3. DNA Integrity in Sperm with SEPT14 Mutations
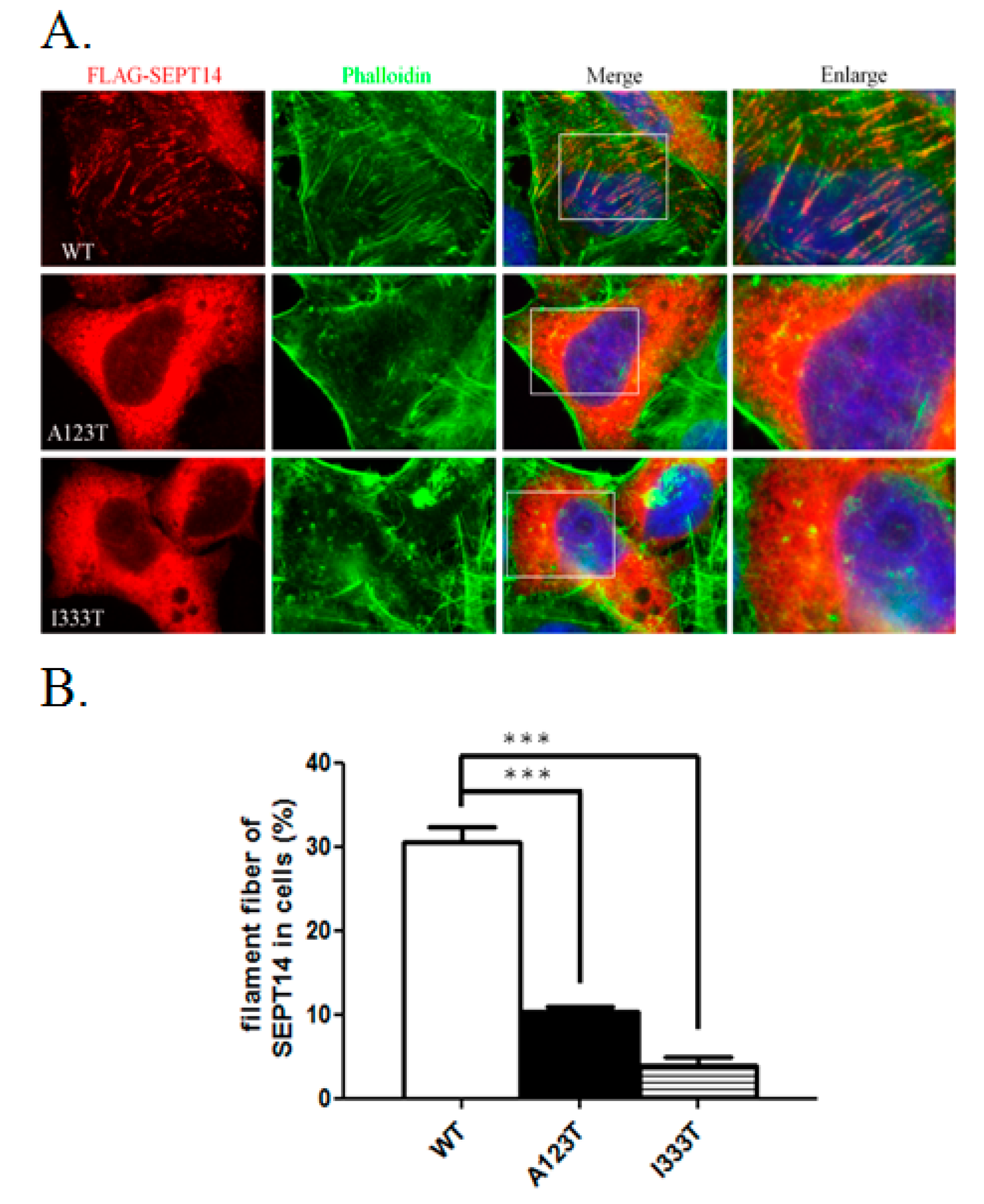
3.4. SEPT14 Mutations Damage the Forming Ability of the Filamentous Structure
4. Discussion
4.1. SEPTs and Sperm DNA Damage
4.2. Sperm DNA Damage and ART
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Boivin, J.; Bunting, L.; Collins, J.A.; Nygren, K.G. International estimates of infertility prevalence and treatment-seeking: Potential need and demand for infertility medical care. Hum. Reprod. 2007, 22, 1506–1512. [Google Scholar] [CrossRef] [PubMed]
- Vayena, E.R.P.; Griffin, P.D. Medical, Ethical & Social Aspects of Assisted Reproduction Current Practices & Controversies in Assisted Reproduction: Report of a WHO Meeting; WHO: Geneva, Switzerland, 2001. [Google Scholar]
- Brugh, V.M., 3rd; Lipshultz, L.I. Male factor infertility: Evaluation and management. Med. Clin. N. Am. 2004, 88, 367–385. [Google Scholar] [CrossRef]
- Brahem, S.; Mehdi, M.; Elghezal, H.; Saad, A. Detection of DNA fragmentation and meiotic segregation in human with isolated teratozoospermia. J. Assist. Reprod. Genet. 2011, 28, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Palermo, G.D.; Neri, Q.V.; Cozzubbo, T.; Rosenwaks, Z. Perspectives on the assessment of human sperm chromatin integrity. Fertil. Steril. 2014, 102, 1508–1517. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhang, Q.; Wang, Y.; Li, Y. Whether sperm deoxyribonucleic acid fragmentation has an effect on pregnancy and miscarriage after in vitro fertilization/intracytoplasmic sperm injection: A systematic review and meta-analysis. Fertil. Steril. 2014, 102, 998–1005 e8. [Google Scholar] [CrossRef] [PubMed]
- Osman, A.; Alsomait, H.; Seshadri, S.; El-Toukhy, T.; Khalaf, Y. The effect of sperm DNA fragmentation on live birth rate after IVF or ICSI: A systematic review and meta-analysis. Reprod. Biomed. Online 2015, 30, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Wdowiak, A.; Bakalczuk, S.; Bakalczuk, G. The effect of sperm DNA fragmentation on the dynamics of the embryonic development in intracytoplasmatic sperm injection. Reprod. Biol. 2015, 15, 94–100. [Google Scholar] [CrossRef]
- Agarwal, A.; Cho, C.L.; Esteves, S.C. Should we evaluate and treat sperm DNA fragmentation? Curr. Opin. Obstet. Gynecol. 2016, 28, 164–171. [Google Scholar] [CrossRef] [Green Version]
- Zini, A.; Boman, J.M.; Belzile, E.; Ciampi, A. Sperm DNA damage is associated with an increased risk of pregnancy loss after IVF and ICSI: Systematic review and meta-analysis. Hum. Reprod. 2008, 23, 2663–2668. [Google Scholar] [CrossRef]
- Evenson, D.; Wixon, R. Meta-analysis of sperm DNA fragmentation using the sperm chromatin structure assay. Reprod. Biomed. Online 2006, 12, 466–472. [Google Scholar] [CrossRef]
- Schulte, R.T.; Ohl, D.A.; Sigman, M.; Smith, G.D. Sperm DNA damage in male infertility: Etiologies, assays, and outcomes. J. Assist. Reprod. Genet. 2010, 27, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Evenson, D.P.; Larson, K.L.; Jost, L.K. Sperm chromatin structure assay: Its clinical use for detecting sperm DNA fragmentation in male infertility and comparisons with other techniques. J. Androl. 2002, 23, 25–43. [Google Scholar] [CrossRef] [PubMed]
- Evenson, D.P.; Darzynkiewicz, Z.; Melamed, M.R. Relation of mammalian sperm chromatin heterogeneity to fertility. Science 1980, 210, 1131–1133. [Google Scholar] [CrossRef] [PubMed]
- Evenson, D.P.; Jost, L.K.; Baer, R.K.; Turner, T.W.; Schrader, S.M. Individuality of DNA denaturation patterns in human sperm as measured by the sperm chromatin structure assay. Reprod. Toxicol. 1991, 5, 115–125. [Google Scholar] [CrossRef]
- De Braekeleer, M.; Nguyen, M.H.; Morel, F.; Perrin, A. Genetic aspects of monomorphic teratozoospermia: A review. J. Assist. Reprod. Genet. 2015, 32, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Coutton, C.; Escoffier, J.; Martinez, G.; Arnoult, C.; Ray, P.F. Teratozoospermia: Spotlight on the main genetic actors in the human. Hum. Reprod. Update 2015, 21, 455–485. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.H.; Wang, Y.Y.; Chen, H.I.; Kuo, Y.C.; Chiou, Y.W.; Lin, H.H.; Wu, C.M.; Hsu, C.C.; Chiang, H.S.; Kuo, P.L. SEPTIN12 Genetic Variants Confer Susceptibility to Teratozoospermia. PLoS ONE 2012, 7, e34011. [Google Scholar] [CrossRef]
- Ravel, C.; Chantot-Bastaraud, S.; El Houate, B.; Berthaut, I.; Verstraete, L.; De Larouziere, V.; Lourenco, D.; Dumaine, A.; Antoine, J.M.; Mandelbaum, J.; et al. Mutations in the protamine 1 gene associated with male infertility. Mol. Hum. Reprod. 2007, 13, 461–464. [Google Scholar] [CrossRef] [Green Version]
- Mostowy, S.; Cossart, P. Septins: The fourth component of the cytoskeleton. Nat. Rev. Mol. Cell Biol. 2012, 13, 183–194. [Google Scholar] [CrossRef]
- Kinoshita, M.; Noda, M. Roles of septins in the mammalian cytokinesis machinery. Cell Struct. Funct. 2001, 26, 667–670. [Google Scholar] [CrossRef]
- Kinoshita, M.; Field, C.M.; Coughlin, M.L.; Straight, A.F.; Mitchison, T.J. Self- and actin-templated assembly of Mammalian septins. Dev. Cell 2002, 3, 791–802. [Google Scholar] [CrossRef]
- Bai, X.; Bowen, J.R.; Knox, T.K.; Zhou, K.; Pendziwiat, M.; Kuhlenbaumer, G.; Sindelar, C.V.; Spiliotis, E.T. Novel septin 9 repeat motifs altered in neuralgic amyotrophy bind and bundle microtubules. J. Cell Biol. 2013, 203, 895–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, C.; Dolat, L.; Angelis, D.; Forgacs, E.; Spiliotis, E.T.; Galkin, V.E. Septin 9 Exhibits Polymorphic Binding to F-Actin and Inhibits Myosin and Cofilin Activity. J. Mol. Biol. 2015, 427, 3273–3284. [Google Scholar] [CrossRef] [Green Version]
- Hall, P.A.; Jung, K.; Hillan, K.J.; Russell, S.E. Expression profiling the human septin gene family. J. Pathol. 2005, 206, 269–278. [Google Scholar] [CrossRef]
- Peterson, E.A.; Petty, E.M. Conquering the complex world of human septins: Implications for health and disease. Clin. Genet. 2010, 77, 511–524. [Google Scholar] [CrossRef] [PubMed]
- Sugino, Y.; Ichioka, K.; Soda, T.; Ihara, M.; Kinoshita, M.; Ogawa, O.; Nishiyama, H. Septins as diagnostic markers for a subset of human asthenozoospermia. J. Urol. 2008, 180, 2706–2709. [Google Scholar] [CrossRef]
- Lhuillier, P.; Rode, B.; Escalier, D.; Lores, P.; Dirami, T.; Bienvenu, T.; Gacon, G.; Dulioust, E.; Toure, A. Absence of annulus in human asthenozoospermia: Case report. Hum. Reprod. 2009, 24, 1296–1303. [Google Scholar] [CrossRef] [PubMed]
- Ihara, M.; Kinoshita, A.; Yamada, S.; Tanaka, H.; Tanigaki, A.; Kitano, A.; Goto, M.; Okubo, K.; Nishiyama, H.; Ogawa, O.; et al. Cortical organization by the septin cytoskeleton is essential for structural and mechanical integrity of mammalian spermatozoa. Dev. Cell 2005, 8, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Kissel, H.; Georgescu, M.M.; Larisch, S.; Manova, K.; Hunnicutt, G.R.; Steller, H. The Sept4 septin locus is required for sperm terminal differentiation in mice. Dev. Cell 2005, 8, 353–364. [Google Scholar] [CrossRef]
- Kuo, Y.C.; Lin, Y.H.; Chen, H.I.; Wang, Y.Y.; Chiou, Y.W.; Lin, H.H.; Pan, H.A.; Wu, C.M.; Su, S.M.; Hsu, C.C.; et al. SEPT12 mutations cause male infertility with defective sperm annulus. Hum. Mutat. 2012, 33, 710–719. [Google Scholar] [CrossRef]
- Lin, Y.H.; Lin, Y.M.; Wang, Y.Y.; Yu, I.S.; Lin, Y.W.; Wang, Y.H.; Wu, C.M.; Pan, H.A.; Chao, S.C.; Yen, P.H.; et al. The expression level of septin12 is critical for spermiogenesis. Am. J. Pathol. 2009, 174, 1857–1868. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.H.; Chou, C.K.; Hung, Y.C.; Yu, I.S.; Pan, H.A.; Lin, S.W.; Kuo, P.L. SEPT12 deficiency causes sperm nucleus damage and developmental arrest of preimplantation embryos. Fertil. Steril. 2011, 95, 363–365. [Google Scholar] [CrossRef]
- Peterson, E.A.; Kalikin, L.M.; Steels, J.D.; Estey, M.P.; Trimble, W.S.; Petty, E.M. Characterization of a SEPT9 interacting protein, SEPT14, a novel testis-specific septin. Mamm. Genome 2007, 18, 796–807. [Google Scholar] [CrossRef] [PubMed]
- Shinoda, T.; Ito, H.; Sudo, K.; Iwamoto, I.; Morishita, R.; Nagata, K. Septin 14 is involved in cortical neuronal migration via interaction with Septin 4. Mol. Biol. Cell 2010, 21, 1324–1334. [Google Scholar] [CrossRef]
- Rozenkrantz, L.; Gan-Or, Z.; Gana-Weisz, M.; Mirelman, A.; Giladi, N.; Bar-Shira, A.; Orr-Urtreger, A. SEPT14 Is Associated with a Reduced Risk for Parkinson’s Disease and Expressed in Human Brain. J. Mol. Neurosci. MN 2016, 59, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Shafipour, M.; Sabbaghian, M.; Shahhoseini, M.; Sadighi Gilani, M.A. Comparative expression analysis of Septin 14 in testes of infertile men with normal spermatogenesis and spermatogenic failure. Iran. J. Reprod. Med. 2014, 12, 205–208. [Google Scholar]
- Vahabi Barzi, N.; Hosseinifar, H.; Nickhah Kalashami, Z.; Sadighi Gilani, M.A.; Sabbaghian, M. Localization of septin 14 protein in sperm. Iran. J. Reprod. Med. 2015, 13, 70. [Google Scholar]
- Wang, Y.Y.; Lin, Y.H.; Wu, Y.N.; Chen, Y.L.; Lin, Y.C.; Cheng, C.Y.; Chiang, H.S. Loss of SLC9A3 decreases CFTR protein and causes obstructed azoospermia in mice. PLoS Genet. 2017, 13, e1006715. [Google Scholar] [CrossRef]
- Erenpreiss, J.; Bars, J.; Lipatnikova, V.; Erenpreisa, J.; Zalkalns, J. Comparative study of cytochemical tests for sperm chromatin integrity. J. Androl. 2001, 22, 45–53. [Google Scholar] [PubMed]
- Haines, G.A.; Hendry, J.H.; Daniel, C.P.; Morris, I.D. Germ cell and dose-dependent DNA damage measured by the comet assay in murine spermatozoaa after testicular X-irradiation. Biol. Reprod. 2002, 67, 854–861. [Google Scholar] [CrossRef]
- Huang, C.Y.; Wang, Y.Y.; Chen, Y.L.; Chen, M.F.; Chiang, H.S.; Kuo, P.L.; Lin, Y.H. CDC42 Negatively Regulates Testis-Specific SEPT12 Polymerization. Int. J. Mol. Sci. 2018, 19, 2627. [Google Scholar] [CrossRef] [PubMed]
- Sakkas, D.; Alvarez, J.G. Sperm DNA fragmentation: Mechanisms of origin, impact on reproductive outcome, and analysis. Fertil. Steril. 2010, 93, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Zini, A.; Libman, J. Sperm DNA damage: Clinical significance in the era of assisted reproduction. CMAJ Can. Med. Assoc. J. 2006, 175, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Avendano, C.; Franchi, A.; Duran, H.; Oehninger, S. DNA fragmentation of normal spermatozoa negatively impacts embryo quality and intracytoplasmic sperm injection outcome. Fertil. Steril. 2010, 94, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Larson-Cook, K.L.; Brannian, J.D.; Hansen, K.A.; Kasperson, K.M.; Aamold, E.T.; Evenson, D.P. Relationship between the outcomes of assisted reproductive techniques and sperm DNA fragmentation as measured by the sperm chromatin structure assay. Fertil. Steril. 2003, 80, 895–902. [Google Scholar] [CrossRef]
- Lopes, S.; Sun, J.G.; Jurisicova, A.; Meriano, J.; Casper, R.F. Sperm deoxyribonucleic acid fragmentation is increased in poor-quality semen samples and correlates with failed fertilization in intracytoplasmic sperm injection. Fertil. Steril. 1998, 69, 528–532. [Google Scholar] [CrossRef]
- Virro, M.R.; Larson-Cook, K.L.; Evenson, D.P. Sperm chromatin structure assay (SCSA) parameters are related to fertilization, blastocyst development, and ongoing pregnancy in in vitro fertilization and intracytoplasmic sperm injection cycles. Fertil. Steril. 2004, 81, 1289–1295. [Google Scholar] [CrossRef] [PubMed]
- Hamidi, J.; Frainais, C.; Amar, E.; Bailly, E.; Clement, P.; Menezo, Y. A double-blinded comparison of in situ TUNEL and aniline blue versus flow cytometry acridine orange for the determination of sperm DNA fragmentation and nucleus decondensation state index. Zygote 2015, 23, 556–562. [Google Scholar] [CrossRef] [PubMed]
- Bungum, M.; Humaidan, P.; Axmon, A.; Spano, M.; Bungum, L.; Erenpreiss, J.; Giwercman, A. Sperm DNA integrity assessment in prediction of assisted reproduction technology outcome. Hum. Reprod. 2007, 22, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Borini, A.; Tarozzi, N.; Bizzaro, D.; Bonu, M.A.; Fava, L.; Flamigni, C.; Coticchio, G. Sperm DNA fragmentation: Paternal effect on early post-implantation embryo development in ART. Hum. Reprod. 2006, 21, 2876–2881. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nucleotide Sequence/Amino Acid Variation | ||||||
|---|---|---|---|---|---|---|
| c.367G>A/p.Ala123Thr | c.998T>C/p.Ile333Thr | |||||
| Ratio | Patients: 3/254; Controls: 0/116 | Patients: 3/254; Controls: 0/116 | ||||
| Exon | 4 | 9 | ||||
| Case No. | Patient 1 | Patient 2 | Patient 3 | Patient 4 | Patient 5 | Patient 6 |
| Clinical feature | Oligoteratozoospermia | Teratozoospermia | Teratozoospermia | Teratozoospermia | Teratozoospermia | Teratozoospermia |
| Abnormal morphology (%) (Krueger criteria) | 90 | 88 | 95 | 91 | 90 | 95 |
| Head defect (%) | 91 | 83 | 91 | 88 | 93 | 94 |
| Neck defect (%) | 1 | 9 | 3 | 9 | 4 | 3 |
| Tail defect (%) | 9 | 11 | 10 | 11 | 5 | 18 |
| Immature (%) | 11 | 13 | 1 | 3 | 3 | 5 |
| Sperm concentration (106/ml) (>15 × 106/mL) | 13 | 145 | 70 | 28 | 68 | 20 |
| Progressive motility (%) (>32%) | 49 | 55 | 37 | 48 | 54 | 45 |
| Non-progressive motility (%) | 15 | 11 | 16 | 17 | 14 | 30 |
| Karyotype | 46, XY | 46, XY | 46, XY | 46, XY | 46, XY | 46, XY |
| Age (years) | 31 | 39 | 35 | 32 | 35 | 38 |
| Cigarette smoking, alcohol consumption, genital disease | No | No | No | No | No | No |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-Y.; Lai, T.-H.; Chen, M.-F.; Lee, H.-L.; Kuo, P.-L.; Lin, Y.-H. SEPT14 Mutations and Teratozoospermia: Genetic Effects on Sperm Head Morphology and DNA Integrity. J. Clin. Med. 2019, 8, 1297. https://doi.org/10.3390/jcm8091297
Wang Y-Y, Lai T-H, Chen M-F, Lee H-L, Kuo P-L, Lin Y-H. SEPT14 Mutations and Teratozoospermia: Genetic Effects on Sperm Head Morphology and DNA Integrity. Journal of Clinical Medicine. 2019; 8(9):1297. https://doi.org/10.3390/jcm8091297
Chicago/Turabian StyleWang, Ya-Yun, Tsung-Hsuan Lai, Mei-Feng Chen, Hui-Ling Lee, Pao-Lin Kuo, and Ying-Hung Lin. 2019. "SEPT14 Mutations and Teratozoospermia: Genetic Effects on Sperm Head Morphology and DNA Integrity" Journal of Clinical Medicine 8, no. 9: 1297. https://doi.org/10.3390/jcm8091297