
Clinical Potential of microRNA-7 in Cancer

Abstract
:1. Introduction
2. microRNA-7 Expression and Regulation
2.1. Biogenesis
2.2. Transcriptional and Post-Transcriptional Regulation
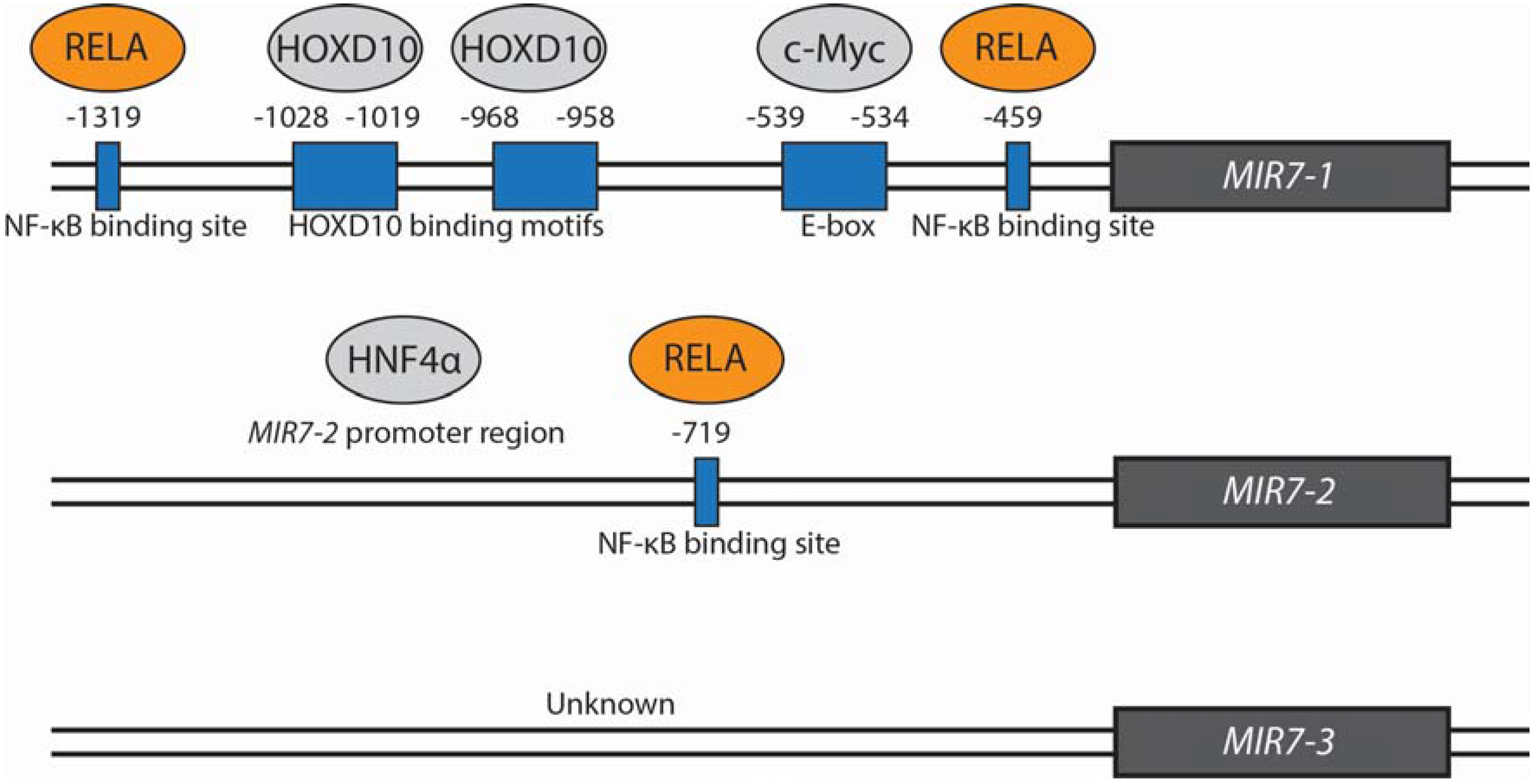

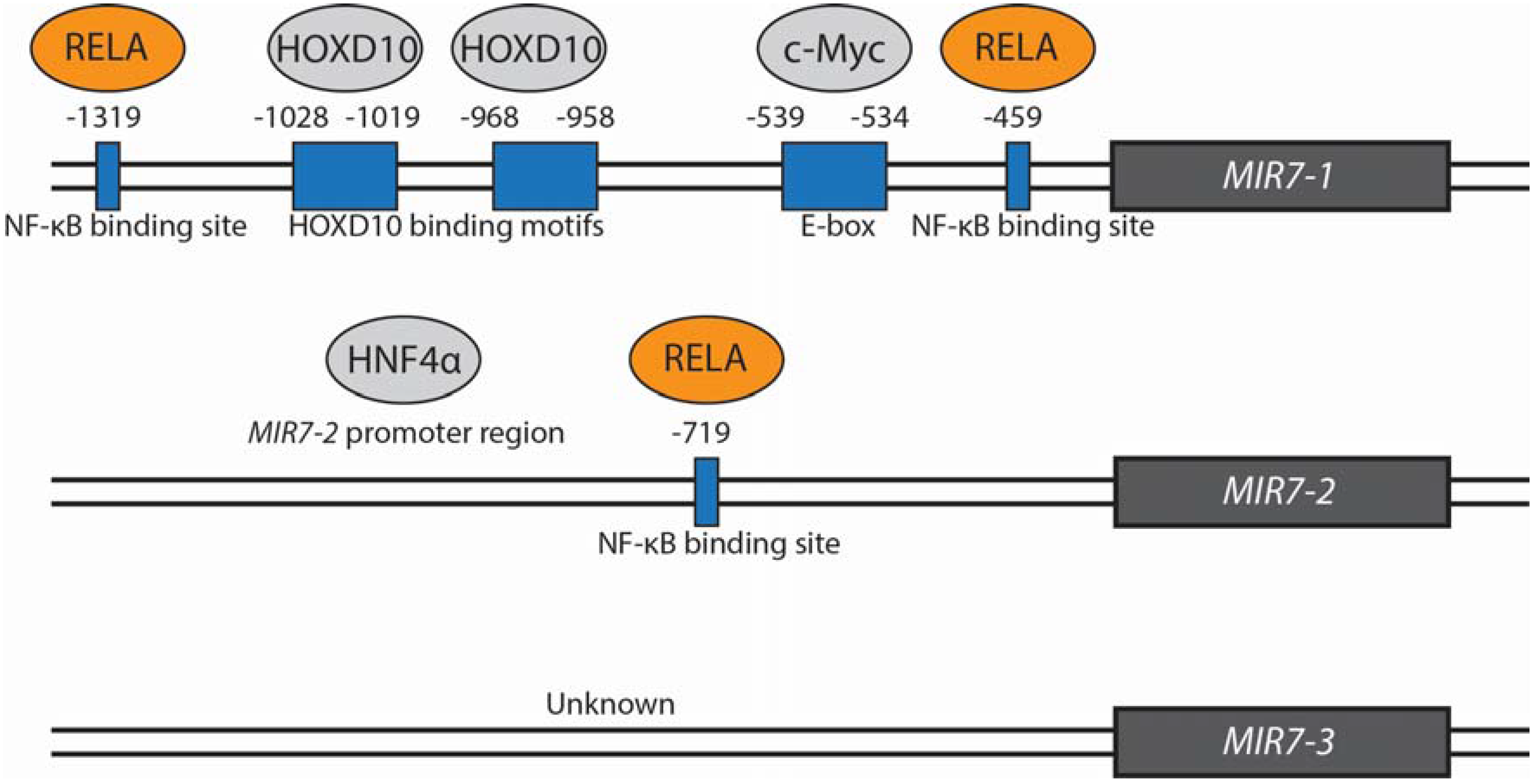{kind=link}
| Regulatory Molecule/Pathway | miR-7 Up- (↑)/Down- (↓) Regulation | Action | Direct/Indirect Interaction | Cancer Type | Reference |
|---|---|---|---|---|---|
| EGFR signaling | ↑ | Via Ras/ERK/Myc and additionally by PI3K/Akt pathways | Indirect | Lung | Chou et al. (2010) [8] |
| c-Myc | ↑ | Binds and stimulates expression from the MIR7-1 promoter | Direct | Lung | Chou et al. (2010) [8] |
| HOXD10 | ↑ | Binds and stimulates expression from the MIR7-1 promoter | Direct | Breast | Reddy et al. (2008) [2] |
| HNF4α | ↑ | Interacts with MIR7-2 promoter | Direct | Liver | Ning et al. (2014) [5] |
| FOXP3 | ↑ | Predicted binding regions in proximity to MIR7-1 and MIR7-2 loci | Not confirmed | Breast | McInnes et al. (2012) [28] |
| HBx | ↑ | Postulated to involve IKKα and IKK/NF-κB signaling | Indirect | Liver | Chen et al. (2013) [30] |
| RELA | ↓ | Binds to MIR7-1 and MIR7-2 promoter regions | Direct | Gastric | Zhao et al. (2015) [31] |
| Usp18 | ↓ | Mechanism not identified | Not confirmed | Cervical, Head and neck, Brain | Duex et al. (2011) [32] |
| HOTAIR | ↓ | Via inhibiting HOXD10 | Indirect | Breast | Zhang et al. (2014) [33] |
| SF2/ASF | ↑ | Binds to pri-miR-7 and promotes maturation via enhancing Drosha cleavage | Direct | Cervical | Wu et al. (2010) [34] |
| HuR | ↓ | Hypothesised to represses miR-7-1 processing which may involve HuR binding in the intron of hnRNPK which hosts the MIR7-1 gene | Not confirmed | Cervical, Lung | Lebedeva et al. (2011) [35], Li et al. (2013) [36] |
| TLR9 signaling | ↓ | Via HuR upregulation which is suggested to involve the PI3K/Akt pathway | Indirect | Lung | Li et al. (2013) [36] |
| MSI2 | ↓ | Binds to the terminal loop of the pri-miR-7 transcript in an HuR-dependent manner resulting in failure of the pri-miR-7-1 transcript to mature | Direct | Cervical, Brain | Choudhury et al. (2013) [25] |
| QKI 5 and QKI 6 | ↓ | Bind to QKI response elements in pri-miR-7-1 resulting in processing failure (binding sites also identified in pri-miR-7-2) | Direct | Brain | Wang et al. 2013 [37] |
| ciRS-7 | ↓ | Contains >70 seed-matched miR-7 binding sites that can sequester miR-7 | Direct | Proof of concept demonstrated in HeLa and HEK293 cells | Hansen et al. 2013 [39], Memczak et al. 2013 [40] |
3. The Role of microRNA-7 in Cancer
3.1. miR-7 is a Tumour Suppressor
3.2. miR-7: The Oncogene?
3.3. Genetic Influence on the Role of miR-7
4. microRNA-7 Has Biomarker Potential
5. Potential for microRNA-7 in Cancer Therapy
5.1. miR-7 Replacement Therapy Alone and in Combination with Current Therapeutic Agents
5.2. Potential for Small Molecule Activation of microRNA-7
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ha, M.; Kim, V.N. Regulation of microrna biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.D.; Ohshiro, K.; Rayala, S.K.; Kumar, R. Microrna-7, a homeobox d10 target, inhibits p21-activated kinase 1 and regulates its functions. Cancer Res. 2008, 68, 8195–8200. [Google Scholar] [CrossRef] [PubMed]
- Kefas, B.; Godlewski, J.; Comeau, L.; Li, Y.; Abounader, R.; Hawkinson, M.; Lee, J.; Fine, H.; Chiocca, E.A.; Lawler, S.; et al. Microrna-7 inhibits the epidermal growth factor receptor and the akt pathway and is down-regulated in glioblastoma. Cancer Res. 2008, 68, 3566–3572. [Google Scholar] [CrossRef] [PubMed]
- Kalinowski, F.C.; Giles, K.M.; Candy, P.A.; Ali, A.; Ganda, C.; Epis, M.R.; Webster, R.J.; Leedman, P.J. Regulation of epidermal growth factor receptor signaling and erlotinib sensitivity in head and neck cancer cells by mir-7. PLoS One 2012, 7, e47067. [Google Scholar] [CrossRef] [PubMed]
- Ning, B.F.; Ding, J.; Liu, J.; Yin, C.; Xu, W.P.; Cong, W.M.; Zhang, Q.; Chen, F.; Han, T.; Deng, X.; et al. Hepatocyte nuclear factor 4alpha-nuclear factor-kappab feedback circuit modulates liver cancer progression. Hepatology 2014, 60, 1607–1619. [Google Scholar] [CrossRef]
- Zhang, N.; Li, X.; Wu, C.W.; Dong, Y.; Cai, M.; Mok, M.T.; Wang, H.; Chen, J.; Ng, S.S.; Chen, M.; et al. Microrna-7 is a novel inhibitor of yy1 contributing to colorectal tumorigenesis. Oncogene 2013, 32, 5078–5088. [Google Scholar] [CrossRef] [PubMed]
- Giles, K.M.; Brown, R.A.; Epis, M.R.; Kalinowski, F.C.; Leedman, P.J. Mirna-7-5p inhibits melanoma cell migration and invasion. Biochem. Biophys. Res. Commun. 2013, 430, 706–710. [Google Scholar] [CrossRef] [PubMed]
- Chou, Y.T.; Lin, H.H.; Lien, Y.C.; Wang, Y.H.; Hong, C.F.; Kao, Y.R.; Lin, S.C.; Chang, Y.C.; Lin, S.Y.; Chen, S.J.; et al. Egfr promotes lung tumorigenesis by activating mir-7 through a ras/erk/myc pathway that targets the ets2 transcriptional repressor erf. Cancer Res. 2010, 70, 8822–8831. [Google Scholar] [CrossRef] [PubMed]
- Meza-Sosa, K.F.; Perez-Garcia, E.I.; Camacho-Concha, N.; Lopez-Gutierrez, O.; Pedraza-Alva, G.; Perez-Martinez, L. Mir-7 promotes epithelial cell transformation by targeting the tumor suppressor klf4. PLoS One 2014, 9, e103987. [Google Scholar] [CrossRef] [PubMed]
- Rai, K.; Takigawa, N.; Ito, S.; Kashihara, H.; Ichihara, E.; Yasuda, T.; Shimizu, K.; Tanimoto, M.; Kiura, K. Liposomal delivery of microrna-7-expressing plasmid overcomes epidermal growth factor receptor tyrosine kinase inhibitor-resistance in lung cancer cells. Mol. Cancer Ther. 2011, 10, 1720–1727. [Google Scholar] [CrossRef] [PubMed]
- Xiong, S.; Zheng, Y.; Jiang, P.; Liu, R.; Liu, X.; Chu, Y. Microrna-7 inhibits the growth of human non-small cell lung cancer a549 cells through targeting bcl-2. Int. J. Biol. Sci. 2011, 7, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.B.; Kjems, J.; Damgaard, C.K. Circular rna and mir-7 in cancer. Cancer Res. 2013, 73, 5609–5612. [Google Scholar] [CrossRef] [PubMed]
- Ncbi Database, Gene. Available online: http://www.ncbi.nlm.nih.gov/gene/?term=hsa-miR-7 (accessed on 11 June 2015).
- Meijer, H.A.; Smith, E.M.; Bushell, M. Regulation of mirna strand selection: Follow the leader? Biochem. Soc. Trans. 2014, 42, 1135–1140. [Google Scholar] [CrossRef] [PubMed]
- Baek, D.; Villen, J.; Shin, C.; Camargo, F.D.; Gygi, S.P.; Bartel, D.P. The impact of micrornas on protein output. Nature 2008, 455, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Ebert, M.S.; Sharp, P.A. Roles for micrornas in conferring robustness to biological processes. Cell 2012, 149, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Cloonan, N.; Wani, S.; Xu, Q.; Gu, J.; Lea, K.; Heater, S.; Barbacioru, C.; Steptoe, A.L.; Martin, H.C.; Nourbakhsh, E.; et al. Micrornas and their isomirs function cooperatively to target common biological pathways. Genome Biol. 2011, 12. [Google Scholar] [CrossRef] [PubMed]
- Landgraf, P.; Rusu, M.; Sheridan, R.; Sewer, A.; Iovino, N.; Aravin, A.; Pfeffer, S.; Rice, A.; Kamphorst, A.O.; Landthaler, M.; et al. A mammalian microrna expression atlas based on small rna library sequencing. Cell 2007, 129, 1401–1414. [Google Scholar] [CrossRef] [PubMed]
- Farh, K.K.; Grimson, A.; Jan, C.; Lewis, B.P.; Johnston, W.K.; Lim, L.P.; Burge, C.B.; Bartel, D.P. The widespread impact of mammalian micrornas on mrna repression and evolution. Science 2005, 310, 1817–1821. [Google Scholar] [CrossRef] [PubMed]
- Bravo-Egana, V.; Rosero, S.; Molano, R.D.; Pileggi, A.; Ricordi, C.; Dominguez-Bendala, J.; Pastori, R.L. Quantitative differential expression analysis reveals mir-7 as major islet microrna. Biochem. Biophys. Res. Commun. 2008, 366, 922–926. [Google Scholar] [CrossRef] [PubMed]
- Correa-Medina, M.; Bravo-Egana, V.; Rosero, S.; Ricordi, C.; Edlund, H.; Diez, J.; Pastori, R.L. Microrna mir-7 is preferentially expressed in endocrine cells of the developing and adult human pancreas. Gene Expr. Patterns GEP 2009, 9, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, J.; Liu, C.; Naji, A.; Stoffers, D.A. Microrna-7 regulates the mtor pathway and proliferation in adult pancreatic beta-cells. Diabetes 2013, 62, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Latreille, M.; Hausser, J.; Stutzer, I.; Zhang, Q.; Hastoy, B.; Gargani, S.; Kerr-Conte, J.; Pattou, F.; Zavolan, M.; Esguerra, J.L.; et al. Microrna-7a regulates pancreatic beta cell function. J. Clin. Investig. 2014, 124, 2722–2735. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Shalom-Feuerstein, R.; Riley, J.; Zhang, S.D.; Tucci, P.; Agostini, M.; Aberdam, D.; Knight, R.A.; Genchi, G.; Nicotera, P.; et al. Mir-7 and mir-214 are specifically expressed during neuroblastoma differentiation, cortical development and embryonic stem cells differentiation, and control neurite outgrowth in vitro. Biochem. Biophys. Res. Commun. 2010, 394, 921–927. [Google Scholar] [CrossRef]
- Choudhury, N.R.; de Lima Alves, F.; de Andres-Aguayo, L.; Graf, T.; Caceres, J.F.; Rappsilber, J.; Michlewski, G. Tissue-specific control of brain-enriched mir-7 biogenesis. Genes Dev. 2013, 27, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Ozsolak, F.; Poling, L.L.; Wang, Z.; Liu, H.; Liu, X.S.; Roeder, R.G.; Zhang, X.; Song, J.S.; Fisher, D.E. Chromatin structure analyses identify mirna promoters. Genes Dev. 2008, 22, 3172–3183. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.C.; Yu, D.; Lee, Y.S.; Wentzel, E.A.; Arking, D.E.; West, K.M.; Dang, C.V.; Thomas-Tikhonenko, A.; Mendell, J.T. Widespread microrna repression by myc contributes to tumorigenesis. Nat. Genet. 2008, 40, 43–50. [Google Scholar] [CrossRef] [PubMed]
- McInnes, N.; Sadlon, T.J.; Brown, C.Y.; Pederson, S.; Beyer, M.; Schultze, J.L.; McColl, S.; Goodall, G.J.; Barry, S.C. Foxp3 and foxp3-regulated micrornas suppress satb1 in breast cancer cells. Oncogene 2012, 31, 1045–1054. [Google Scholar] [CrossRef] [PubMed]
- Sadlon, T.J.; Wilkinson, B.G.; Pederson, S.; Brown, C.Y.; Bresatz, S.; Gargett, T.; Melville, E.L.; Peng, K.; D’Andrea, R.J.; Glonek, G.G.; et al. Genome-wide identification of human foxp3 target genes in natural regulatory t cells. J. Immunol. 2010, 185, 1071–1081. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Chien, P.H.; Chen, W.S.; Chien, Y.F.; Hsu, Y.Y.; Wang, L.Y.; Chen, J.Y.; Lin, C.W.; Huang, T.C.; Yu, Y.L.; et al. Hepatitis b virus-encoded x protein downregulates egfr expression via inducing microrna-7 in hepatocellular carcinoma cells. Evid.-Based Complement. Altern. Med. eCAM 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.-D.; Lu, Y.-Y.; Guo, H.; Xie, H.-H.; He, L.-J.; Shen, G.-F.; Zhou, J.-F.; Li, T.; Hu, S.-J.; Zhou, L.; et al. Microrna-7/nf-κb signaling regulatory feedback circuit regulates gastric carcinogenesis. J. Cell Biol. 2015, 210, 613–627. [Google Scholar] [CrossRef]
- Duex, J.E.; Comeau, L.; Sorkin, A.; Purow, B.; Kefas, B. Usp18 regulates epidermal growth factor (egf) receptor expression and cancer cell survival via microrna-7. J. Biol. Chem. 2011, 286, 25377–25386. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Cai, K.; Wang, J.; Wang, X.; Cheng, K.; Shi, F.; Jiang, L.; Zhang, Y.; Dou, J. Mir-7, inhibited indirectly by lincrna hotair, directly inhibits setdb1 and reverses the emt of breast cancer stem cells by downregulating the stat3 pathway. Stem Cells 2014, 32, 2858–2868. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Sun, S.; Tu, K.; Gao, Y.; Xie, B.; Krainer, A.R.; Zhu, J. A splicing-independent function of sf2/asf in microrna processing. Mol. Cell 2010, 38, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Lebedeva, S.; Jens, M.; Theil, K.; Schwanhausser, B.; Selbach, M.; Landthaler, M.; Rajewsky, N. Transcriptome-wide analysis of regulatory interactions of the rna-binding protein hur. Mol. Cell 2011, 43, 340–352. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.J.; Wang, C.H.; Zhou, Y.; Liao, Z.Y.; Zhu, S.F.; Hu, Y.; Chen, C.; Luo, J.M.; Wen, Z.K.; Xu, L.; et al. Tlr9 signaling repressed tumor suppressor mir-7 expression through up-regulation of hur in human lung cancer cells. Cancer Cell Int. 2013, 13. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Vogel, G.; Yu, Z.; Richard, S. The qki-5 and qki-6 rna binding proteins regulate the expression of microrna 7 in glial cells. Mol. Cell. Biol. 2013, 33, 1233–1243. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.B.; Wiklund, E.D.; Bramsen, J.B.; Villadsen, S.B.; Statham, A.L.; Clark, S.J.; Kjems, J. Mirna-dependent gene silencing involving ago2-mediated cleavage of a circular antisense rna. EMBO J. 2011, 30, 4414–4422. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural rna circles function as efficient microrna sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular rnas are a large class of animal rnas with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Kalinowski, F.C.; Brown, R.A.; Ganda, C.; Giles, K.M.; Epis, M.R.; Horsham, J.; Leedman, P.J. Microrna-7: A tumor suppressor mirna with therapeutic potential. Int. J. Biochem. Cell Biol. 2014, 54, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Gu, D.N.; Huang, Q.; Tian, L. The molecular mechanisms and therapeutic potential of microrna-7 in cancer. Expert opin. Ther. Targets 2015, 19, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Webster, R.J.; Giles, K.M.; Price, K.J.; Zhang, P.M.; Mattick, J.S.; Leedman, P.J. Regulation of epidermal growth factor receptor signaling in human cancer cells by microrna-7. J. Biol. Chem. 2009, 284, 5731–5741. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, N.; Gallagher, M.; Lao, N.; Gallagher, C.; Clarke, C.; Doolan, P.; Aherne, S.; Blanco, A.; Meleady, P.; Clynes, M.; et al. Mir-7 triggers cell cycle arrest at the g1/s transition by targeting multiple genes including skp2 and psme3. PLoS ONE 2013, 8, e65671. [Google Scholar] [CrossRef] [PubMed]
- Xiong, S.; Zheng, Y.; Jiang, P.; Liu, R.; Liu, X.; Qian, J.; Gu, J.; Chang, L.; Ge, D.; Chu, Y.; et al. Pa28gamma emerges as a novel functional target of tumour suppressor microrna-7 in non-small-cell lung cancer. Br. J. Cancer 2014, 110, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Luo, X.; Li, P.; Tan, J.; Wang, X.; Xiang, T.; Ren, G. Mir-7-5p suppresses cell proliferation and induces apoptosis of breast cancer cells mainly by targeting reggamma. Cancer Lett. 2015, 358, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Hu, S.; Zhang, X.; Wang, L.; Zhang, X.; Yan, B.; Zhao, J.; Yang, A.; Zhang, R. Microrna-7 arrests cell cycle in g1 phase by directly targeting ccne1 in human hepatocellular carcinoma cells. Biochem. Biophys. Res. Commun. 2014, 443, 1078–1084. [Google Scholar] [CrossRef] [PubMed]
- Suto, T.; Yokobori, T.; Yajima, R.; Morita, H.; Fujii, T.; Yamaguchi, S.; Altan, B.; Tsutsumi, S.; Asao, T.; Kuwano, H.; et al. Microrna-7 expression in colorectal cancer is associated with poor prognosis and regulates cetuximab sensitivity via egfr regulation. Carcinogenesis 2015, 36, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zheng, Y.; Sun, G.; Xiong, S. Restoration of mir-7 expression suppresses the growth of lewis lung cancer cells by modulating epidermal growth factor receptor signaling. Oncol. Rep. 2014, 32, 2511–2516. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.M.; Choi, E.J.; Kim, I.A. Microrna-7 increases radiosensitivity of human cancer cells with activated egfr-associated signaling. Radiother. Oncol. 2011, 101, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Saydam, O.; Senol, O.; Wurdinger, T.; Mizrak, A.; Ozdener, G.B.; Stemmer-Rachamimov, A.O.; Yi, M.; Stephens, R.M.; Krichevsky, A.M.; Saydam, N.; et al. Mirna-7 attenuation in schwannoma tumors stimulates growth by upregulating three oncogenic signaling pathways. Cancer Res. 2011, 71, 852–861. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Chen, M.; Zhou, J.; Mo, M.S.; Zhu, L.H.; Liu, Y.P.; Gui, Q.J.; Zhang, L.; Li, G.Q. Mir-7 inhibits the invasion and metastasis of gastric cancer cells by suppressing epidermal growth factor receptor expression. Oncol. Rep. 2014, 31, 1715–1722. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Jiang, Z.; Huang, J.; Huang, S.; Li, Y.; Yu, S.; Yu, S.; Liu, X. Mir-7 inhibits glioblastoma growth by simultaneously interfering with the pi3k/atk and raf/mek/erk pathways. Int. J. Oncol. 2014, 44, 1571–1580. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Xue, J.L.; Shen, Q.; Chen, J.; Tian, L. Microrna-7 inhibits tumor growth and metastasis by targeting the phosphoinositide 3-kinase/akt pathway in hepatocellular carcinoma. Hepatology 2012, 55, 1852–1862. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.G.; Wang, Y.Y.; Fan, L.G.; Luo, H.; Han, B.; Sun, L.H.; Wang, X.F.; Zhang, J.X.; Cao, L.; Wang, X.R.; et al. Microrna-7 regulates glioblastoma cell invasion via targeting focal adhesion kinase expression. Chin. Med. J. 2011, 124, 2616–2621. [Google Scholar] [PubMed]
- Kong, X.; Li, G.; Yuan, Y.; He, Y.; Wu, X.; Zhang, W.; Wu, Z.; Chen, T.; Wu, W.; Lobie, P.E.; et al. Microrna-7 inhibits epithelial-to-mesenchymal transition and metastasis of breast cancer cells via targeting fak expression. PLoS ONE 2012, 7, e41523. [Google Scholar] [CrossRef] [PubMed]
- Okuda, H.; Xing, F.; Pandey, P.R.; Sharma, S.; Watabe, M.; Pai, S.K.; Mo, Y.Y.; Iiizumi-Gairani, M.; Hirota, S.; Liu, Y.; et al. Mir-7 suppresses brain metastasis of breast cancer stem-like cells by modulating klf4. Cancer Res. 2013, 73, 1434–1444. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Dou, W.; He, L.; Liang, S.; Tie, J.; Liu, C.; Li, T.; Lu, Y.; Mo, P.; Shi, Y.; et al. Microrna-7 functions as an anti-metastatic microrna in gastric cancer by targeting insulin-like growth factor-1 receptor. Oncogene 2013, 32, 1363–1372. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Dai, L.X.; Zhang, S.; Yang, Y.; Yan, N.; Fan, P.; Dai, L.; Tian, H.W.; Cheng, L.; Zhang, X.M.; et al. Regulation of epidermal growth factor receptor signaling by plasmid-based microrna-7 inhibits human malignant gliomas growth and metastasis in vivo. Neoplasma 2013, 60, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Babae, N.; Bourajjaj, M.; Liu, Y.; van Beijnum, J.R.; Cerisoli, F.; Scaria, P.V.; Verheul, M.; van Berkel, M.P.; Pieters, E.H.; van Haastert, R.J.; et al. Systemic mirna-7 delivery inhibits tumor angiogenesis and growth in murine xenograft glioblastoma. Oncotarget 2014, 5, 6687–6700. [Google Scholar] [PubMed]
- Cao, D.; Jiang, J.; Tsukamoto, T.; Liu, R.; Ma, L.; Jia, Z.; Kong, F.; Oshima, M.; Cao, X. Canolol inhibits gastric tumors initiation and progression through cox-2/pge2 pathway in k19-c2me transgenic mice. PLoS ONE 2015, 10, e0120938. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhu, F.; Chen, P. Mir-7 and mir-218 epigenetically control tumor suppressor genes rassf1a and claudin-6 by targeting hoxb3 in breast cancer. Biochem. Biophys. Res. Commun. 2012, 424, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Foekens, J.A.; Sieuwerts, A.M.; Smid, M.; Look, M.P.; de Weerd, V.; Boersma, A.W.; Klijn, J.G.; Wiemer, E.A.; Martens, J.W. Four mirnas associated with aggressiveness of lymph node-negative, estrogen receptor-positive human breast cancer. Proc. Natl. Acad. Sci. USA 2008, 105, 13021–13026. [Google Scholar] [CrossRef] [PubMed]
- Veerla, S.; Lindgren, D.; Kvist, A.; Frigyesi, A.; Staaf, J.; Persson, H.; Liedberg, F.; Chebil, G.; Gudjonsson, S.; Borg, A.; et al. Mirna expression in urothelial carcinomas: Important roles of mir-10a, mir-222, mir-125b, mir-7 and mir-452 for tumor stage and metastasis, and frequent homozygous losses of mir-31. Int. J. Cancer 2009, 124, 2236–2242. [Google Scholar] [CrossRef] [PubMed]
- Rao, Q.; Shen, Q.; Zhou, H.; Peng, Y.; Li, J.; Lin, Z. Aberrant microrna expression in human cervical carcinomas. Med. Oncol. 2012, 29, 1242–1248. [Google Scholar] [CrossRef] [PubMed]
- Honegger, A.; Schilling, D.; Bastian, S.; Sponagel, J.; Kuryshev, V.; Sultmann, H.; Scheffner, M.; Hoppe-Seyler, K.; Hoppe-Seyler, F. Dependence of intracellular and exosomal micrornas on viral e6/e7 oncogene expression in hpv-positive tumor cells. PLoS Pathog. 2015, 11, e1004712. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, Y.; Akao, Y.; Taniguchi, K.; Kamatani, A.; Tahara, T.; Kamano, T.; Nakano, N.; Komura, N.; Ikuno, H.; Ohmori, T.; et al. Relationship between expression of onco-related mirnas and the endoscopic appearance of colorectal tumors. Int. J. Mol. Sci. 2015, 16, 1526–1543. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, F.E.; Ahmed, N.C.; Vos, P.W.; Bonnerup, C.; Atkins, J.N.; Casey, M.; Nuovo, G.J.; Naziri, W.; Wiley, J.E.; Mota, H.; et al. Diagnostic microrna markers to screen for sporadic human colon cancer in stool: I. Proof of principle. Cancer Genom. Proteom. 2013, 10, 93–113. [Google Scholar]
- Xu, K.; Chen, Z.; Qin, C.; Song, X. Mir-7 inhibits colorectal cancer cell proliferation and induces apoptosis by targeting xrcc2. OncoTargets Ther. 2014, 7, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.M.; Byrom, M.W.; Shelton, J.; Ford, L.P. Antisense inhibition of human mirnas and indications for an involvement of mirna in cell growth and apoptosis. Nucleic Acids Res. 2005, 33, 1290–1297. [Google Scholar] [CrossRef] [PubMed]
- Rowland, B.D.; Peeper, D.S. Klf4, p21 and context-dependent opposing forces in cancer. Nat. Rev. Cancer 2006, 6, 11–23. [Google Scholar] [CrossRef] [PubMed]
- De la Iglesia, N.; Konopka, G.; Puram, S.V.; Chan, J.A.; Bachoo, R.M.; You, M.J.; Levy, D.E.; Depinho, R.A.; Bonni, A. Identification of a pten-regulated stat3 brain tumor suppressor pathway. Genes Dev. 2008, 22, 449–462. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.F.; Lai, R. Stat3 in cancer-friend or foe? Cancers 2014, 6, 1408–1440. [Google Scholar] [CrossRef] [PubMed]
- Shahab, S.W.; Matyunina, L.V.; Hill, C.G.; Wang, L.; Mezencev, R.; Walker, L.D.; McDonald, J.F. The effects of microrna transfections on global patterns of gene expression in ovarian cancer cells are functionally coordinated. BMC Med. Genom. 2012, 5. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.Y.; Zhang, J.; Niu, W.; Guo, W.; Song, H.T.; Li, H.Y.; Fan, H.M.; Zhao, L.; Zhong, A.F.; Dai, Y.H.; et al. A preliminary analysis of microrna as potential clinical biomarker for schizophrenia. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2015, 163B, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Tandon, M.; Alevizos, I.; Illei, G.G. The majority of micrornas detectable in serum and saliva is concentrated in exosomes. PLoS ONE 2012, 7, e30679. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.L.; Cao, Y.; Liu, D.; Xu, M.; Liu, H.; Tang, R.N.; Ma, K.L.; Liu, B.C. Isolation and quantification of micrornas from urinary exosomes/microvesicles for biomarker discovery. Int. J. Biol. Sci. 2013, 9, 1021–1031. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating micrornas as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, S.; Li, L.; Li, M.; Guo, C.; Yao, J.; Mi, S. Exosome and exosomal microrna: Trafficking, sorting, and function. Genom. Proteom. Bioinform. 2015, 13, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, J.D.; Chevillet, J.R.; Kroh, E.M.; Ruf, I.K.; Pritchard, C.C.; Gibson, D.F.; Mitchell, P.S.; Bennett, C.F.; Pogosova-Agadjanyan, E.L.; Stirewalt, D.L.; et al. Argonaute2 complexes carry a population of circulating micrornas independent of vesicles in human plasma. Proc. Natl. Acad. Sci. USA 2011, 108, 5003–5008. [Google Scholar] [CrossRef] [PubMed]
- Vickers, K.C.; Palmisano, B.T.; Shoucri, B.M.; Shamburek, R.D.; Remaley, A.T. Micrornas are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nat. Cell Biol. 2011, 13, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, M.A.; Haj-Ahmad, Y. Promising candidate urinary microrna biomarkers for the early detection of hepatocellular carcinoma among high-risk hepatitis c virus egyptian patients. J. Cancer 2012, 3, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Haj-Ahmad, T.A.; Abdalla, M.A.; Haj-Ahmad, Y. Potential urinary mirna biomarker candidates for the accurate detection of prostate cancer among benign prostatic hyperplasia patients. J. Cancer 2014, 5, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Erbes, T.; Hirschfeld, M.; Rucker, G.; Jaeger, M.; Boas, J.; Iborra, S.; Mayer, S.; Gitsch, G.; Stickeler, E. Feasibility of urinary microrna detection in breast cancer patients and its potential as an innovative non-invasive biomarker. BMC Cancer 2015, 15. [Google Scholar] [CrossRef] [PubMed]
- Salazar, C.; Nagadia, R.; Pandit, P.; Cooper-White, J.; Banerjee, N.; Dimitrova, N.; Coman, W.B.; Punyadeera, C. A novel saliva-based microrna biomarker panel to detect head and neck cancers. Cell. Oncol. 2014, 37, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Yau, T.O.; Wu, C.W.; Dong, Y.; Tang, C.M.; Ng, S.S.; Chan, F.K.; Sung, J.J.; Yu, J. Microrna-221 and microrna-18a identification in stool as potential biomarkers for the non-invasive diagnosis of colorectal carcinoma. Br. J. Cancer 2014, 111, 1765–1771. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.W.; Ng, S.C.; Dong, Y.; Tian, L.; Ng, S.S.; Leung, W.W.; Law, W.T.; Yau, T.O.; Chan, F.K.; Sung, J.J.; et al. Identification of microrna-135b in stool as a potential noninvasive biomarker for colorectal cancer and adenoma. Clin. Cancer Res. 2014, 20, 2994–3002. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.A.; Baxter, D.H.; Zhang, S.; Huang, D.Y.; Huang, K.H.; Lee, M.J.; Galas, D.J.; Wang, K. The microrna spectrum in 12 body fluids. Clin. Chem. 2010, 56, 1733–1741. [Google Scholar] [CrossRef] [PubMed]
- Sieuwerts, A.M.; Mostert, B.; Bolt-de Vries, J.; Peeters, D.; de Jongh, F.E.; Stouthard, J.M.; Dirix, L.Y.; van Dam, P.A.; van Galen, A.; de Weerd, V.; et al. Mrna and microrna expression profiles in circulating tumor cells and primary tumors of metastatic breast cancer patients. Clin. Cancer Res. 2011, 17, 3600–3618. [Google Scholar] [CrossRef] [PubMed]
- Mostert, B.; Sieuwerts, A.M.; Martens, J.W.; Sleijfer, S. Diagnostic applications of cell-free and circulating tumor cell-associated mirnas in cancer patients. Expert Rev. Mol. Diagn. 2011, 11, 259–275. [Google Scholar] [PubMed]
- Rosenfeld, N.; Aharonov, R.; Meiri, E.; Rosenwald, S.; Spector, Y.; Zepeniuk, M.; Benjamin, H.; Shabes, N.; Tabak, S.; Levy, A.; et al. Micrornas accurately identify cancer tissue origin. Nat. Biotechnol. 2008, 26, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Xiang, J.; Li, Z.; Lu, S.; Hu, J.; Gao, X.; Yu, L.; Wang, L.; Wang, J.; Wu, Y.; et al. A plasma microrna panel for early detection of colorectal cancer. Int. J. Cancer. 2015, 136, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Kitano, M.; Rahbari, R.; Patterson, E.E.; Steinberg, S.M.; Prasad, N.B.; Wang, Y.; Zeiger, M.A.; Kebebew, E. Evaluation of candidate diagnostic micrornas in thyroid fine-needle aspiration biopsy samples. Thyroid 2012, 22, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.I.; Teixeira, A.L.; Dias, F.; Mauricio, J.; Lobo, F.; Morais, A.; Medeiros, R. Influence of peripheral whole-blood microrna-7 and microrna-221 high expression levels on the acquisition of castration-resistant prostate cancer: Evidences from in vitro and in vivo studies. Tumour Biol. 2014, 35, 7105–7113. [Google Scholar] [CrossRef] [PubMed]
- Pogribny, I.P.; Filkowski, J.N.; Tryndyak, V.P.; Golubov, A.; Shpyleva, S.I.; Kovalchuk, O. Alterations of micrornas and their targets are associated with acquired resistance of mcf-7 breast cancer cells to cisplatin. Int. J. Cancer. 2010, 127, 1785–1794. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Liu, X.; Zheng, Y.; Gu, J.; Xiong, S.; Jiang, P.; Jiang, X.; Huang, E.; Yang, Y.; Ge, D.; et al. Microrna-7 sensitizes non-small cell lung cancer cells to paclitaxel. Oncol. Lett. 2014, 8, 2193–2200. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Zheng, L.; Huang, M.; Wang, Y.; Bi, F. Microrna expression profiles associated with acquired gefitinib-resistance in human lung adenocarcinoma cells. Mol. Med. Rep. 2015, 11, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Fang, B.; Zeng, F.; Pang, H.; Zhang, J.; Shi, Y.; Wu, X.; Cheng, L.; Ma, C.; Xia, J.; et al. Curcumin inhibits cell growth and invasion through up-regulation of mir-7 in pancreatic cancer cells. Toxicol. Lett. 2014, 231, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Melo, S.; Villanueva, A.; Moutinho, C.; Davalos, V.; Spizzo, R.; Ivan, C.; Rossi, S.; Setien, F.; Casanovas, O.; Simo-Riudalbas, L.; et al. Small molecule enoxacin is a cancer-specific growth inhibitor that acts by enhancing tar rna-binding protein 2-mediated microrna processing. Proc. Natl. Acad. Sci. USA 2011, 108, 4394–4399. [Google Scholar] [CrossRef] [PubMed]
- Tu, C.Y.; Chen, C.H.; Hsia, T.C.; Hsu, M.H.; Wei, Y.L.; Yu, M.C.; Chen, W.S.; Hsu, K.W.; Yeh, M.H.; Liu, L.C.; et al. Trichostatin a suppresses egfr expression through induction of microrna-7 in an hdac-independent manner in lapatinib-treated cells. BioMed Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, S.; Meng, F.; Wang, J.; Zhang, Y.; Dai, E.; Yu, X.; Li, X.; Jiang, W. Sm2mir: A database of the experimentally validated small molecules’ effects on microrna expression. Bioinformatics 2013, 29, 409–411. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horsham, J.L.; Kalinowski, F.C.; Epis, M.R.; Ganda, C.; Brown, R.A.M.; Leedman, P.J. Clinical Potential of microRNA-7 in Cancer. J. Clin. Med. 2015, 4, 1668-1687. https://doi.org/10.3390/jcm4091668
Horsham JL, Kalinowski FC, Epis MR, Ganda C, Brown RAM, Leedman PJ. Clinical Potential of microRNA-7 in Cancer. Journal of Clinical Medicine. 2015; 4(9):1668-1687. https://doi.org/10.3390/jcm4091668
Chicago/Turabian StyleHorsham, Jessica L., Felicity C. Kalinowski, Michael R. Epis, Clarissa Ganda, Rikki A. M. Brown, and Peter J. Leedman. 2015. "Clinical Potential of microRNA-7 in Cancer" Journal of Clinical Medicine 4, no. 9: 1668-1687. https://doi.org/10.3390/jcm4091668