Effects of the Japanese Encephalitis Virus Genotype V-Derived Sub-Viral Particles on the Immunogenicity of the Vaccine Characterized by a Novel Virus-Like Particle-Based Assay

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
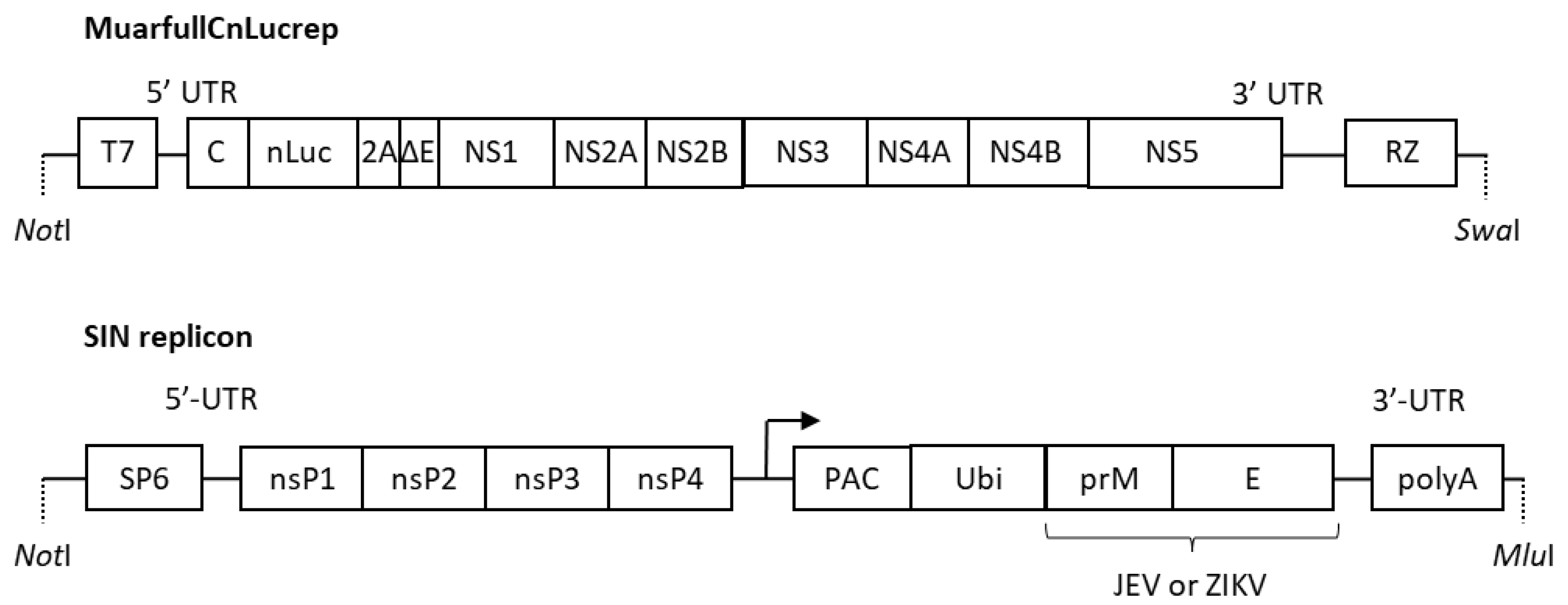
2.2. Preparation of VLPs and SVPs
2.3. Immunization of Mice
2.4. Neutralization Test
2.5. Enzyme-Linked ImmunoSorbent Assay (ELISA)
3. Results
3.1. Preparation of the Luciferase Gene-Transducing VLP Bearing the E Protein of JEV and ZKV
3.2. Evaluation of the VLP for Application in the Neutralization Test
3.3. Immunogenicity of JEVAX and JEV Muar Strain-Derived SVP Against JEV GV
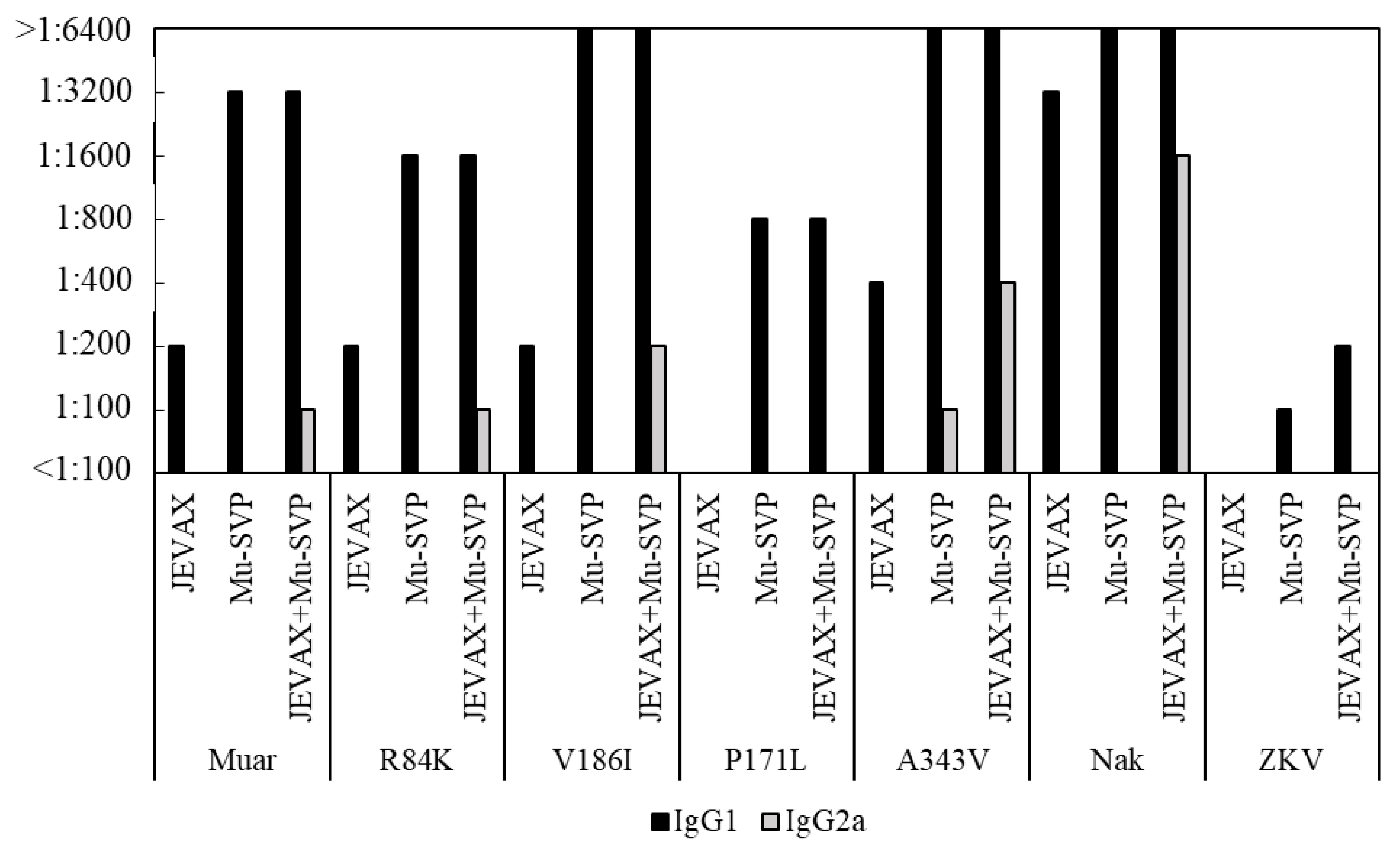
3.4. Th1/Th2 Immune Profile Induced by JEVAX and Mu-SVP
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Solomon, T.; Dung, N.M.; Kneen, R.; Gainsborough, M.; Vaughn, D.W.; Khanh, V.T. Japanese encephalitis. J. Neurol. Neurosurg. Psychiatry 2000, 68, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Pierson, T.C.; Diamond, M.S. Flaviviruses. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 747–794. [Google Scholar]
- Ishikawa, T.; Konishi, E. Combating Japanese encephalitis: Vero-cell derived inactivated vaccines and the situation in Japan. Future Virol. 2010, 5, 785–799. [Google Scholar] [CrossRef]
- Van den Hurk, A.F.; Ritchie, S.A.; Mackenzie, J.S. Ecology and geographical expansion of Japanese encephalitis virus. Annu. Rev. Entomol. 2009, 54, 17–35. [Google Scholar] [CrossRef] [PubMed]
- Lindenbach, B.D.; Murray, C.; Thiel, H.J.; Rice, C.M. Flaviviridae. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 712–746. [Google Scholar]
- Hasegawa, H.; Yoshida, M.; Fujita, S.; Kobayashi, Y. Comparison of structural proteins among antigenically different Japanese encephalitis virus strains. Vaccine 1994, 12, 841–844. [Google Scholar] [CrossRef]
- Ishikawa, T.; Konishi, E. Japanese encephalitis: Epidemiology, prevention and current status of antiviral drug development. Expert Opin. Orphan Drugs 2014, 2, 923–936. [Google Scholar] [CrossRef]
- Solomon, T.; Ni, H.; Beasley, D.W.; Ekkelenkamp, M.; Cardosa, M.J.; Barrett, A.D. Origin and evolution of Japanese encephalitis virus in southeast Asia. J. Virol. 2003, 77, 3091–3098. [Google Scholar] [CrossRef]
- Mohammed, M.A.; Galbraith, S.E.; Radford, A.D.; Dove, W.; Takasaki, T.; Kurane, I.; Solomon, T. Molecular phylogenetic and evolutionary analyses of Muar strain of Japanese encephalitis virus reveal it is the missing fifth genotype. Infect. Genet. Evol. 2011, 11, 855–862. [Google Scholar] [CrossRef]
- Li, M.H.; Fu, S.H.; Chen, W.X.; Wang, H.Y.; Guo, Y.H.; Liu, Q.Y.; Li, Y.X.; Luo, H.M.; Da, W.; Duo Ji, D.Z.; et al. Genotype v Japanese encephalitis virus is emerging. PLoS Negl. Trop. Dis. 2011, 5, e1231. [Google Scholar] [CrossRef]
- Takhampunya, R.; Kim, H.C.; Tippayachai, B.; Kengluecha, A.; Klein, T.A.; Lee, W.J.; Grieco, J.; Evans, B.P. Emergence of Japanese encephalitis virus genotype V in the Republic of Korea. Virol. J. 2011, 8, 449. [Google Scholar] [CrossRef]
- Kim, H.; Cha, G.W.; Jeong, Y.E.; Lee, W.G.; Chang, K.S.; Roh, J.Y.; Yang, S.C.; Park, M.Y.; Park, C.; Shin, E.H. Detection of Japanese encephalitis virus genotype V in Culex orientalis and Culex pipiens (Diptera: Culicidae) in Korea. PLoS ONE 2015, 10, e0116547. [Google Scholar] [CrossRef]
- De Wispelaere, M.; Frenkiel, M.P.; Despres, P. A Japanese encephalitis virus genotype 5 molecular clone is highly neuropathogenic in a mouse model: Impact of the structural protein region on virulence. J. Virol. 2015, 89, 5862–5875. [Google Scholar] [CrossRef]
- Tajima, S.; Yagasaki, K.; Kotaki, A.; Tomikawa, T.; Nakayama, E.; Moi, M.L.; Lim, C.K.; Saijo, M.; Kurane, I.; Takasaki, T. In vitro growth, pathogenicity and serological characteristics of the Japanese encephalitis virus genotype V Muar strain. J. Gen. Virol. 2015, 96, 2661–2669. [Google Scholar] [CrossRef]
- Ishikawa, T.; Yamanaka, A.; Konishi, E. A review of successful flavivirus vaccines and the problems with those flaviviruses for which vaccines are not yet available. Vaccine 2014, 32, 1326–1337. [Google Scholar] [CrossRef]
- Erra, E.O.; Askling, H.H.; Yoksan, S.; Rombo, L.; Riutta, J.; Vene, S.; Lindquist, L.; Vapalahti, O.; Kantele, A. Cross-protective capacity of Japanese encephalitis (JE) vaccines against circulating heterologous JE virus genotypes. Clin. Infect. Dis. 2013, 56, 267–270. [Google Scholar] [CrossRef]
- Ishikawa, T.; Abe, M.; Masuda, M. Construction of an infectious molecular clone of Japanese encephalitis virus genotype V and its derivative subgenomic replicon capable of expressing a foreign gene. Virus Res. 2015, 195, 153–161. [Google Scholar] [CrossRef]
- Hall, M.P.; Unch, J.; Binkowski, B.F.; Valley, M.P.; Butler, B.L.; Wood, M.G.; Otto, P.; Zimmerman, K.; Vidugiris, G.; Machleidt, T.; et al. Engineered luciferase reporter from a deep sea shrimp utilizing a novel imidazopyrazinone substrate. ACS Chem. Biol. 2012, 7, 1848–1857. [Google Scholar] [CrossRef]
- Ishikawa, T.; Widman, D.G.; Bourne, N.; Konishi, E.; Mason, P.W. Construction and evaluation of a chimeric pseudoinfectious virus vaccine to prevent Japanese encephalitis. Vaccine 2008, 26, 2772–2781. [Google Scholar] [CrossRef]
- Whitton, J.L.; Oldstone, M.B.A. The immune response to flaviviruses. In Fields Virology, 4th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott-Raven Publisher: New York, NY, USA, 2001; pp. 285–320. [Google Scholar]
- Yoshii, K.; Ikawa, A.; Chiba, Y.; Omori, Y.; Maeda, J.; Murata, R.; Kariwa, H.; Takashima, I. Establishment of a neutralization test involving reporter gene-expressing virus-like particles of tick-borne encephalitis virus. J. Virol. Methods 2009, 161, 173–176. [Google Scholar] [CrossRef] [Green Version]
- Mercier-Delarue, S.; Durier, C.; de Verdiere, N.C.; Poveda, J.D.; Meiffredy, V.; Garcia, M.D.F.; Lastere, S.; Cesaire, R.; Manuggera, J.C.; Molina, J.M.; et al. Screening test for neutralizing antibodies against yellow fever virus, based on a flavivirus pseudotype. PLoS ONE 2017, 12, e0177882. [Google Scholar] [CrossRef]
- Mattia, K.; Puffer, B.A.; Williams, K.L.; Gonzalez, R.; Murray, M.; Sluzas, E.; Pagano, D.; Ajith, S.; Bower, M.; Berdougo, E.; et al. Dengue reporter virus particles for measuring neutralizing antibodies against each of the four dengue serotypes. PLoS ONE 2011, 6, e27252. [Google Scholar] [CrossRef]
- Garg, H.; Sedano, M.; Plata, G.; Punke, E.B.; Joshi, A. Development of Virus-Like-Particle Vaccine and Reporter Assay for Zika Virus. J. Virol. 2017, 91, e00834-17. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Fu, S.; Gao, X.; Li, M.; Cui, S.; Li, X.; Cao, Y.; Lei, W.; Lu, Z.; He, Y.; et al. Low Protective Efficacy of the Current Japanese Encephalitis Vaccine against the Emerging Genotype 5 Japanese Encephalitis Virus. PLoS Negl. Trop. Dis. 2016, 10, e0004686. [Google Scholar] [CrossRef]
- Morita, K.; Tadano, M.; Nakaji, S.; Kosai, K.; Mathenge, E.G.; Pandey, B.D.; Hasebe, F.; Inoue, S.; Igarashi, A. Locus of a virus neutralization epitope on the Japanese encephalitis virus envelope protein determined by use of long PCR-based region-specific random mutagenesis. Virology 2001, 287, 417–426. [Google Scholar] [CrossRef]
- Lin, C.W.; Wu, S.C. A functional epitope determinant on domain III of the Japanese encephalitis virus envelope protein interacted with neutralizing-antibody combining sites. J. Virol. 2003, 77, 2600–2606. [Google Scholar] [CrossRef]
- Mason, P.W.; Pincus, S.; Fournier, M.J.; Mason, T.L.; Shope, R.E.; Paoletti, E. Japanese encephalitis virus-vaccinia recombinants produce particulate forms of the structural membrane proteins and induce high levels of protection against lethal JEV infection. Virology 1991, 180, 294–305. [Google Scholar] [CrossRef]
- Pierson, T.C.; Xu, Q.; Nelson, S.; Oliphant, T.; Nybakken, G.E.; Fremont, D.H.; Diamond, M.S. The stoichiometry of antibody-mediated neutralization and enhancement of West Nile virus infection. Cell Host Microbe 2007, 1, 135–145. [Google Scholar] [CrossRef]
- Kimura-Kuroda, J.; Yasui, K. Protection of mice against Japanese encephalitis virus by passive administration with monoclonal antibodies. J. Immunol. 1988, 141, 3606–3610. [Google Scholar]
- Manolova, V.; Flace, A.; Bauer, M.; Schwarz, K.; Saudan, P.; Bachmann, M.F. Nanoparticles target distinct dendritic cell populations according to their size. Eur. J. Immunol. 2008, 38, 1404–1413. [Google Scholar] [CrossRef]
- Aberle, J.H.; Aberle, S.W.; Kofler, R.M.; Mandl, C.W. Humoral and cellular immune response to RNA immunization with flavivirus replicons derived from tick-borne encephalitis virus. J. Virol. 2005, 79, 15107–15113. [Google Scholar] [CrossRef]
- Coutelier, J.P.; van der Logt, J.T.; Heessen, F.W.; Warnier, G.; Van Snick, J. IgG2a restriction of murine antibodies elicited by viral infections. J. Exp. Med. 1987, 165, 64–69. [Google Scholar] [CrossRef]
- Chen, H.W.; Huang, H.W.; Hu, H.M.; Chung, H.H.; Wu, S.H.; Chong, P.; Tao, M.H.; Pan, C.H. A poorly neutralizing IgG2a/c response elicited by a DNA vaccine protects mice against Japanese encephalitis virus. J. Gen. Virol. 2014, 95, 1983–1990. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, E.; Coban, C.; Ishii, K.J. Particulate adjuvant and innate immunity: Past achievements, present findings, and future prospects. Int. Rev. Immunol. 2013, 32, 209–220. [Google Scholar] [CrossRef]
- Levast, B.; Awate, S.; Babiuk, L.; Mutwiri, G.; Gerdts, V.; van Drunen Littel-van den Hurk, S. Vaccine Potentiation by Combination Adjuvants. Vaccines 2014, 2, 297–322. [Google Scholar] [CrossRef] [Green Version]
- Ramakrishna, C.; Ravi, V.; Desai, A.; Subbakrishna, D.K.; Shankar, S.K.; Chandramuki, A. T helper responses to Japanese encephalitis virus infection are dependent on the route of inoculation and the strain of mouse used. J. Gen. Virol. 2003, 84, 1559–1567. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Accession number * | Designation ** | Amino Acid Position in E Protein | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 52 | 58 | 84 | 129 | 156 | 171 | 186 | 208 | 240 | 292 | 343 | |||
| Muar | HM596272 | Muar | Q | T | R | I | S | P | V | T | L | D | A*** |
| K12AS1148 | KJ420590 | R84K | E | T | K | I | T | P | V | S | M | E | A |
| K12AS1151 | KJ420591 | R84K | E | T | K | I | T | P | V | S | M | E | A |
| 10-1827 | JN587258 | R84K | E | T | K | I | T | P | V | S | M | E | A |
| XZ0934 | JF915894 | V186I | E | T | R | I | T | P | I | S | M | E | A |
| K12YJ1203 | KJ420592 | P171L | E | T | K | I | T | L | V | S | M | E | A |
| K12HC959 | KJ420589 | A343V | E | A | K | T | T | P | V | S | M | E | V |
| Beijing 1 | AB920347 | - | Q | S | K | T | S | P | V | S | M | D | A |
| Nakayama | AB920348 | - | Q | S | K | T | S | P | V | S | M | D | A |
| Immunogen | Virus like particles Used in the Neutralization Test * | ||||||
|---|---|---|---|---|---|---|---|
| Muar | R84K | V186I | P171L | A343V | Nak | ZKV | |
| 1/10 dose of JEVAX | <1:10 | <1:10 | <1:10 | <1:10 | <1:10 | 1:10 | <1:10 |
| Mu-SVP** (800 ng) | 1:10 | <1:10 | 1:10 | 1:10 | 1:10 | 1:20 | <1:10 |
| 1/10 dose of JEVAX and Mu-SVP (400 ng) | 1:40 | 1:40 | 1:40 | 1:40 | 1:40 | 1:80 | <1:10 |
| Saline | <1:10 | <1:10 | <1:10 | <1:10 | <1:10 | <1:10 | <1:10 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Honjo, S.; Masuda, M.; Ishikawa, T. Effects of the Japanese Encephalitis Virus Genotype V-Derived Sub-Viral Particles on the Immunogenicity of the Vaccine Characterized by a Novel Virus-Like Particle-Based Assay. Vaccines 2019, 7, 81. https://doi.org/10.3390/vaccines7030081
Honjo S, Masuda M, Ishikawa T. Effects of the Japanese Encephalitis Virus Genotype V-Derived Sub-Viral Particles on the Immunogenicity of the Vaccine Characterized by a Novel Virus-Like Particle-Based Assay. Vaccines. 2019; 7(3):81. https://doi.org/10.3390/vaccines7030081
Chicago/Turabian StyleHonjo, Sarah, Michiaki Masuda, and Tomohiro Ishikawa. 2019. "Effects of the Japanese Encephalitis Virus Genotype V-Derived Sub-Viral Particles on the Immunogenicity of the Vaccine Characterized by a Novel Virus-Like Particle-Based Assay" Vaccines 7, no. 3: 81. https://doi.org/10.3390/vaccines7030081
APA StyleHonjo, S., Masuda, M., & Ishikawa, T. (2019). Effects of the Japanese Encephalitis Virus Genotype V-Derived Sub-Viral Particles on the Immunogenicity of the Vaccine Characterized by a Novel Virus-Like Particle-Based Assay. Vaccines, 7(3), 81. https://doi.org/10.3390/vaccines7030081