
Time-Resolved Study of Nanoparticle Induced Apoptosis Using Microfabricated Single Cell Arrays

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
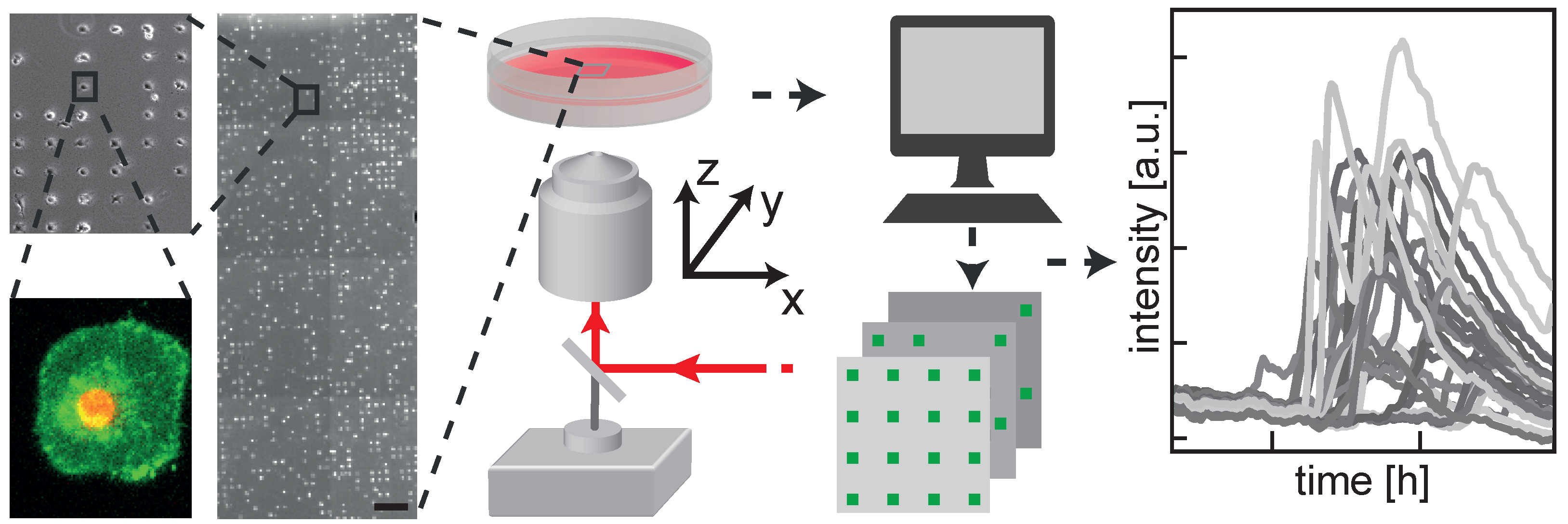
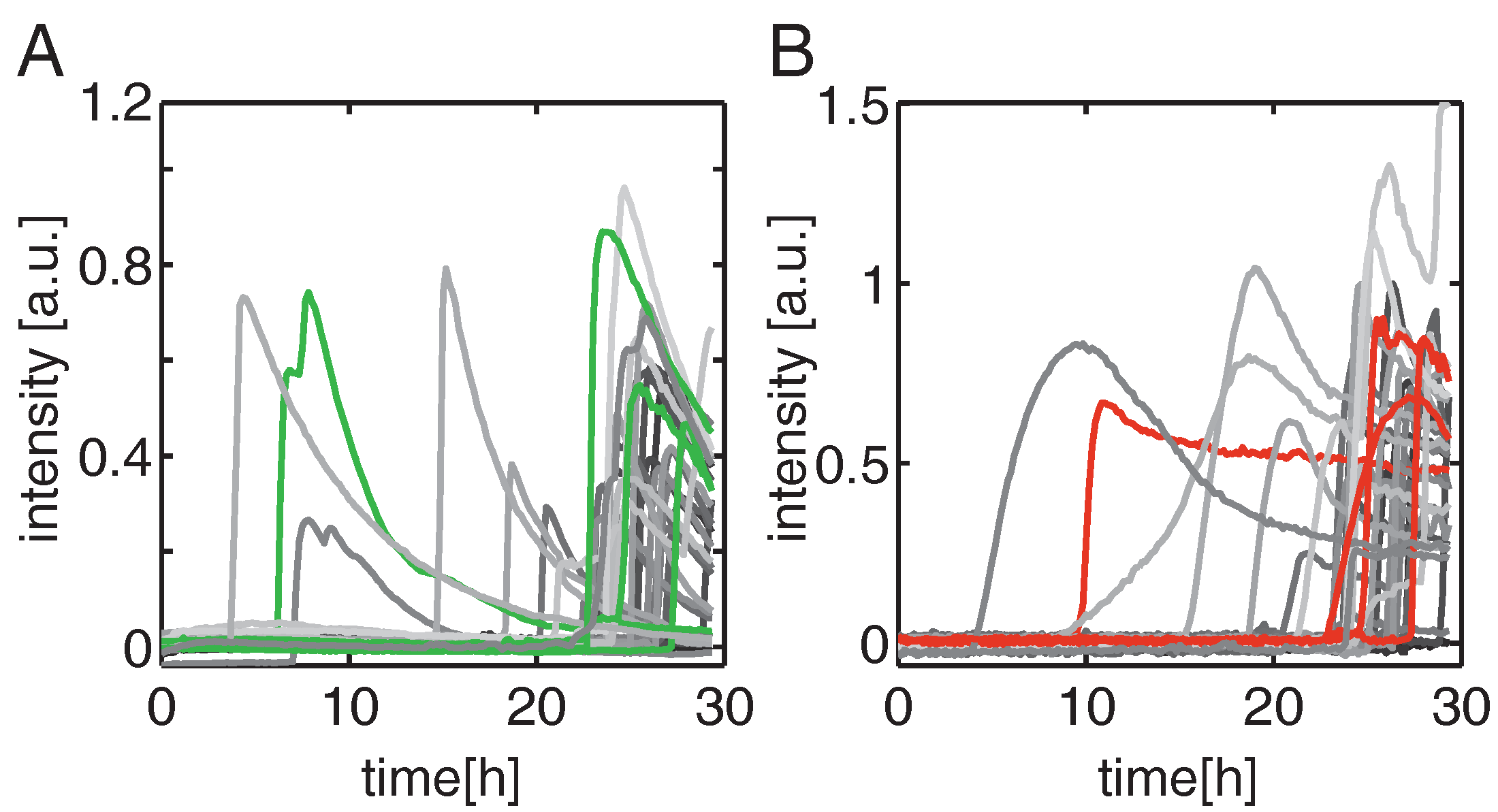
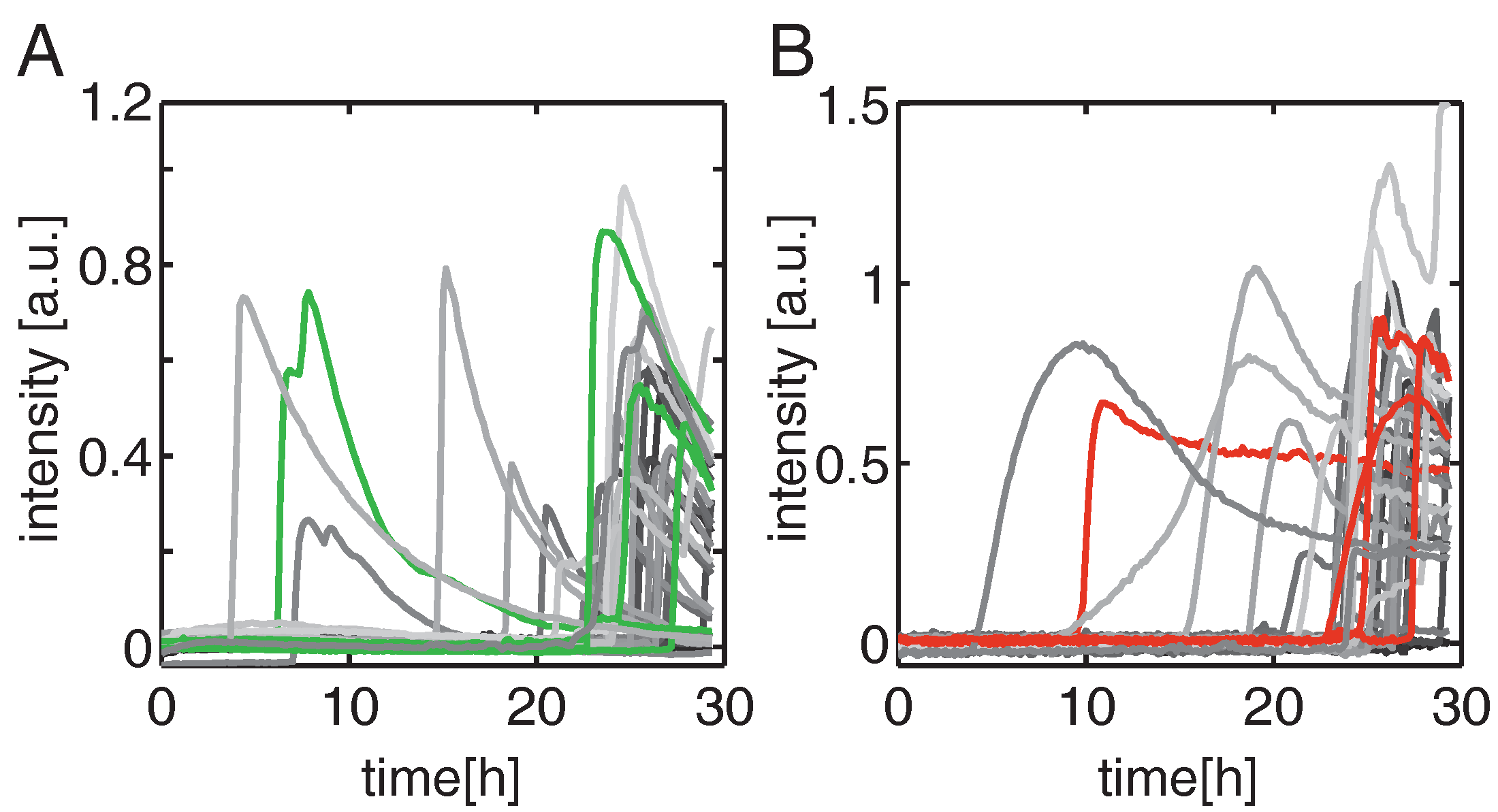
2.1. Highly Parallel Assessment of the Kinetics of Cell Death
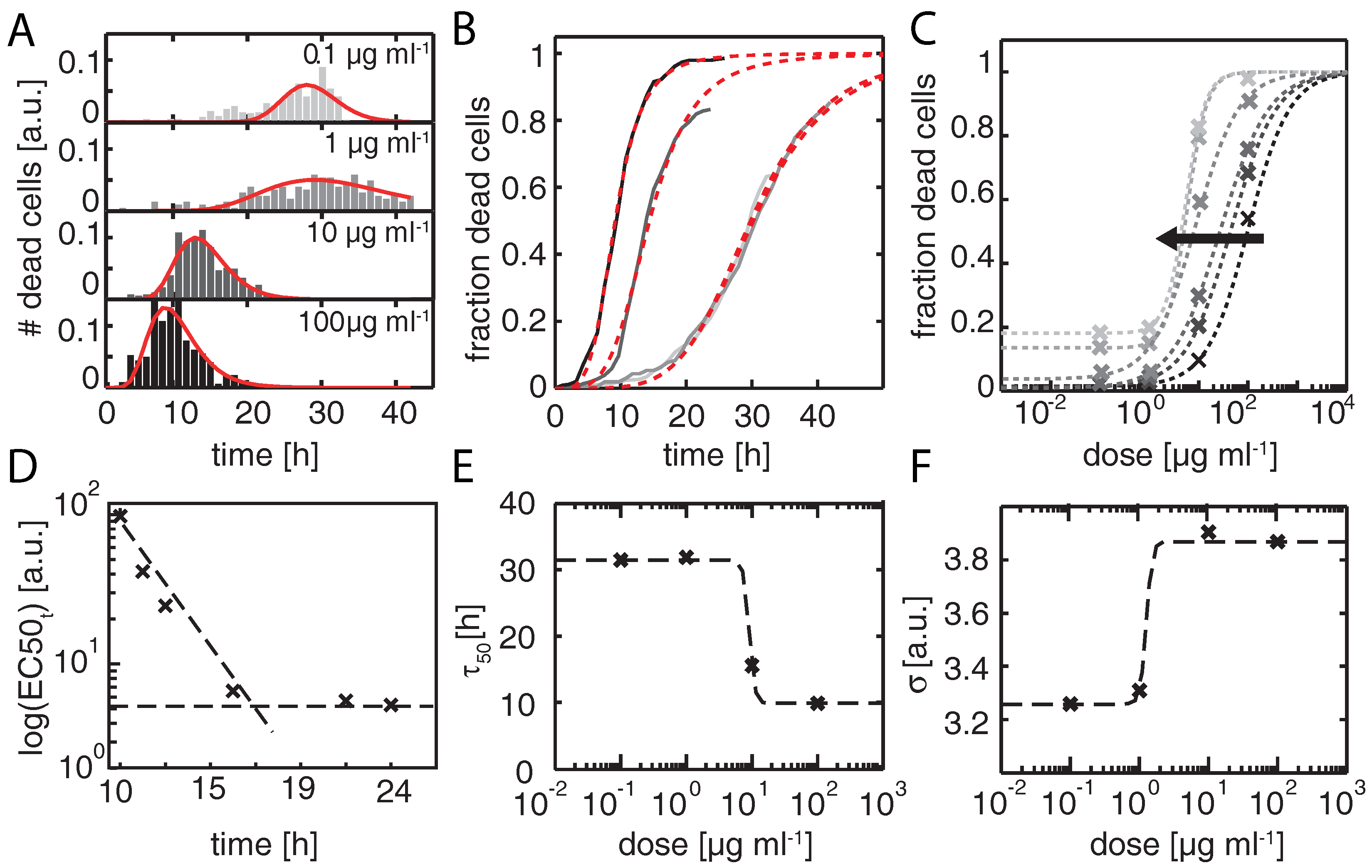
2.2. Time Dependent Dose Response Function
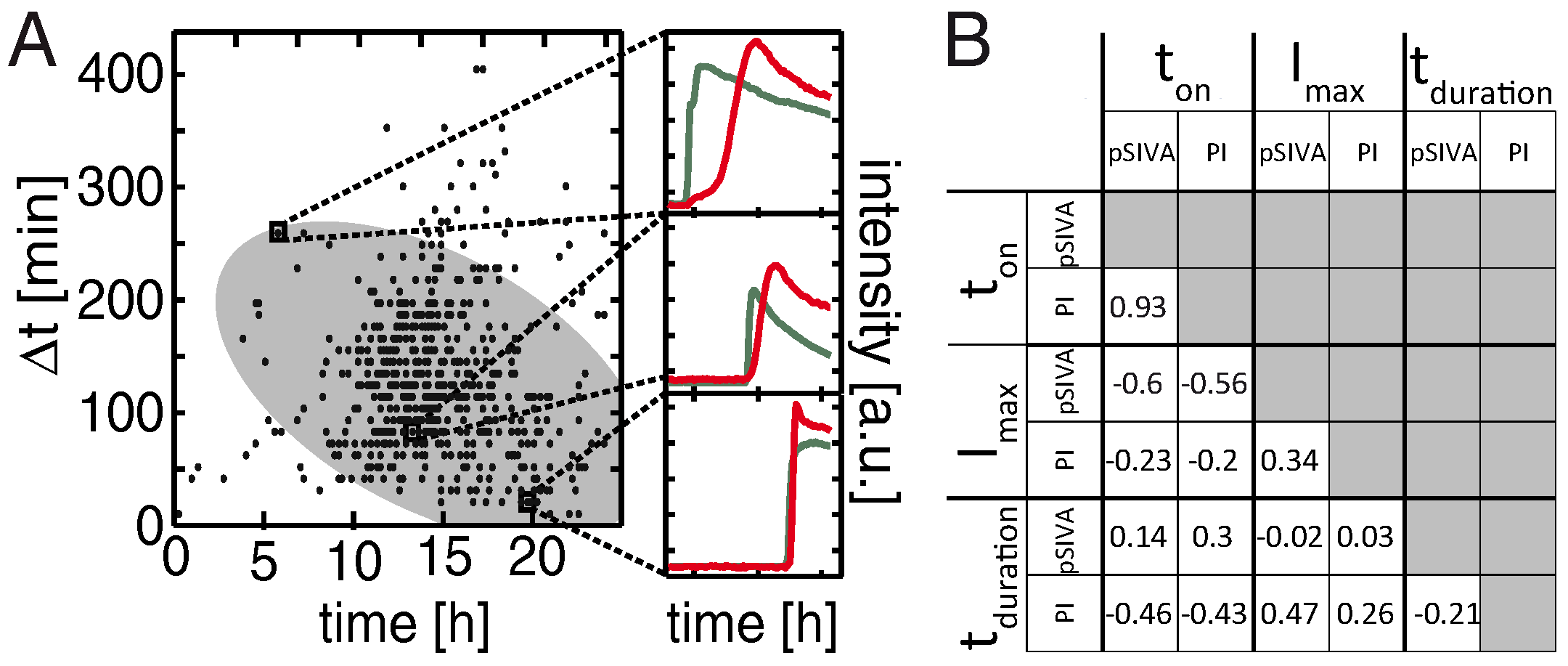
2.3. Two-Parameter Correlation of Cell Death
3. Discussion
4. Experimental Section
4.1. Cell Culture
4.2. NP Preparation and Characterization
4.3. Micropatterning
4.4. Time-Resolved Fluorescence Microscopy
4.5. Image and Data Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Colvin, V.L. The potential environmental impact of engineered nanomaterials. Nat. Biotech. 2003, 21, 1166–1170. [Google Scholar] [CrossRef] [PubMed]
- Nel, A.; Xia, T.; Madler, L.; Li, N. Toxic potential of materials at the nanolevel. Science 2006, 311, 622–627. [Google Scholar] [CrossRef] [PubMed]
- Anguissola, S.; Garry, D.; Salvati, A.; O’Brien, P.J.; Dawson, K.A. High content analysis provides mechanistic insights on the pathways of toxicity induced by amine-modified polystyrene nanoparticles. PLoS ONE 2014, 9, e108025. [Google Scholar] [CrossRef] [PubMed]
- Elsaesser, A.; Howard, C.V. Toxicology of nanoparticles. Adv. Drug Deliv. Rev. 2012, 64, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Love, S.A.; Maurer-Jones, M.A.; Thompson, J.W.; Lin, Y.S.; Haynes, C.L. Assessing nanoparticle toxicity. Annu. Rev. Anal. Chem. 2012, 5, 181–205. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Liu, Y.; Kong, X.; Lobie, P.E.; Chen, C.; Zhu, T. Nanotoxicity: A growing need for study in the endocrine system. Small 2013, 9, 1654–1671. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, C.; Simões, S.; Gaspar, R. Paclitaxel-loaded PLGA nanoparticles: preparation, physicochemical characterization and in vitro anti-tumoral activity. J. Control. Release 2002, 83, 273–286. [Google Scholar] [CrossRef]
- Kommareddy, S.; Amiji, M. Preparation and evaluation of thiol-modified gelatin nanoparticles for intracellular DNA delivery in response to glutathione. Bioconjugate Chem. 2005, 16, 1423–1432. [Google Scholar] [CrossRef] [PubMed]
- Jose, G.P.; Santra, S.; Mandal, S.K.; Sengupta, T.K. Singlet oxygen mediated DNA degradation by copper nanoparticles: potential towards cytotoxic effect on cancer cells. J. Nanobiotechnology 2011, 9, 9. [Google Scholar] [CrossRef] [PubMed]
- Bexiga, M.G.; Varela, J.A.; Wang, F.; Fenaroli, F.; Salvati, A.; Lynch, I.; Simpson, J.C.; Dawson, K.A. Cationic nanoparticles induce caspase 3-, 7- and 9-mediated cytotoxicity in a human astrocytoma cell line. Nanotoxicology 2011, 5, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Wlodkowic, D.; Skommer, J.; Darzynkiewicz, Z. Flow cytometry-based apoptosis detection. Methods Mol. Biol. 2009, 559, 19–32. [Google Scholar] [PubMed]
- Summers, H.D.; Rees, P.; Holton, M.D.; Rowan Brown, M.; Chappell, S.C.; Smith, P.J.; Errington, R.J. Statistical analysis of nanoparticle dosing in a dynamic cellular system. Nat. Nano 2011, 6, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Owen, M.S.; Lee, R.E.; Gaudet, S. Cell-to-cell variability in cell death: can systems biology help us make sense of it all? Cell Death Differ. 2014, 5, e1261. [Google Scholar] [CrossRef] [PubMed]
- Spencer, S.L.; Gaudet, S.; Albeck, J.G.; Burke, J.M.; Sorger, P.K. Non-genetic origins of cell-to-cell variability in TRAIL-induced apoptosis. Nature 2009, 459, 428–432. [Google Scholar] [CrossRef] [PubMed]
- Love, K.R.; Bagh, S.; Choi, J.; Love, J.C. Microtools for single-cell analysis in biopharmaceutical development and manufacturing. Trends Biotechnol. 2013, 31, 280–286. [Google Scholar] [CrossRef] [PubMed]
- Ware, M.J.; Godin, B.; Singh, N.; Majithia, R.; Shamsudeen, S.; Serda, R.E.; Meissner, K.E.; Rees, P.; Summers, H.D. Analysis of the Influence of Cell Heterogeneity on Nanoparticle Dose Response. ACS Nano 2014, 8, 6693–6700. [Google Scholar] [CrossRef] [PubMed]
- Aftab, O.; Nazir, M.; Fryknas, M.; Hammerling, U.; Larsson, R.; Gustafsson, M.G. Label free high throughput screening for apoptosis inducing chemicals using time-lapse microscopy signal processing. Apoptosis 2014, 19, 1411–1418. [Google Scholar] [CrossRef] [PubMed]
- Albeck, J.G.; Burke, J.M.; Aldridge, B.B.; Zhang, M.; Lauffenburger, D.A.; Sorger, P.K. Quantitative analysis of pathways controlling extrinsic apoptosis in single cells. Mol. Cell 2008, 30, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Forrester, H.B.; Albright, N.; Ling, C.C.; Dewey, W.C. Computerized video time-lapse analysis of apoptosis of REC:Myc cells X-irradiated in different phases of the cell cycle. Radiat. Res. 2000, 154, 625–639. [Google Scholar] [CrossRef]
- Milani, S.; Baldelli Bombelli, F.; Pitek, A.S.; Dawson, K.A.; Rädler, J. Reversible versus irreversible binding of transferrin to polystyrene nanoparticles: Soft and hard corona. ACS Nano 2012, 6, 2532–2541. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Yu, L.; Monopoli, M.P.; Sandin, P.; Mahon, E.; Salvati, A.; Dawson, K.A. The biomolecular corona is retained during nanoparticle uptake and protects the cells from the damage induced by cationic nanoparticles until degraded in the lysosomes. Nanomedicine 2013, 9, 1159–1168. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.T. Secondary necrosis: The natural outcome of the complete apoptotic program. FEBS Lett. 2010, 584, 4491–4499. [Google Scholar] [CrossRef] [PubMed]
- Vanden Berghe, T.; Grootjans, S.; Goossens, V.; Dondelinger, Y.; Krysko, D.V.; Takahashi, N.; Vandenabeele, P. Determination of apoptotic and necrotic cell death in vitro and in vivo. Methods 2013, 61, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Wlodkowic, D.; Faley, S.; Zagnoni, M.; Wikswo, J.P.; Cooper, J.M. Microfluidic single-cell array cytometry for the analysis of tumor apoptosis. Anal. Chem. 2009, 81, 5517–5523. [Google Scholar] [CrossRef] [PubMed]
- Röttgermann, P.J.; Hertrich, S.; Berts, I.; Albert, M.; Segerer, F.J.; Moulin, J.F.; Nickel, B.; Rädler, J.O. Cell motility on polyethylene glycol block copolymers correlates to fibronectin surface adsorption. Macromol. Biosci. 2014, 14, 1755–1763. [Google Scholar] [CrossRef] [PubMed]
- Röttgermann, P.J.F.; Piera Alberola, A.; Rädler, J.O. Cellular Self-Organization on Micro-Structured Surfaces. Soft Matter 2014, 10, 2397–2404. [Google Scholar] [CrossRef] [PubMed]
- Waring, M.J. Complex formation between ethidium bromide and nucleic acids. J. Mol. Biol. 1965, 13, 269–282. [Google Scholar] [CrossRef]
- Kim, J.A.; Åberg, C.; de Cárcer, G.; Malumbres, M.; Salvati, A.; Dawson, K.A. Low dose of amino-modified nanoparticles induces cell cycle arrest. ACS Nano 2013, 7, 7483–7494. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, C.; Suzuki, T.; Kashiwagi, A.; Yomo, T.; Kaneko, K. Ubiquity of log-normal distributions in intra-cellular reaction dynamics. Biophysics 2005, 1, 25–31. [Google Scholar] [CrossRef]
- Frasch, S.C.; Henson, P.M.; Kailey, J.M.; Richter, D.A.; Janes, M.S.; Fadok, V.A.; Bratton, D.L. Regulation of phospholipid scramblase activity during apoptosis and cell activation by protein kinase Cδ. J. Biol. Chem. 2000, 275, 23065–23073. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Meng, X.W.; Flatten, K.S.; Loegering, D.A.; Kaufmann, S.H. Phosphatidylserine exposure during apoptosis reflects bidirectional trafficking between plasma membrane and cytoplasm. Cell Death Differ. 2013, 20, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Edens, L.J.; White, K.H.; Jevtic, P.; Li, X.; Levy, D.L. Nuclear size regulation: from single cells to development and disease. Trends Cell Biol. 2013, 23, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Huber, M.D.; Gerace, L. The size-wise nucleus: Nuclear volume control in eukaryotes. J. Cell Biol. 2007, 179, 583–584. [Google Scholar] [CrossRef] [PubMed]
- Krippendorff, B.F.; Neuhaus, R.; Lienau, P.; Reichel, A.; Huisinga, W. Mechanism-based inhibition: Deriving K(I) and k(inact) directly from time-dependent IC(50) values. J. Biomol. Screen 2009, 14, 913–923. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, L.N.; Colborn, T.; Hayes, T.B.; Heindel, J.J.; Jacobs, D.R.J.; Lee, D.H.; Shioda, T.; Soto, A.M.; vom Saal, F.S.; Welshons, W.V.; et al. Hormones and endocrine-disrupting chemicals: Low-dose effects and nonmonotonic dose responses. Endocr Rev. 2012, 33, 378–455. [Google Scholar] [CrossRef] [PubMed]
- Yedjou, C.; Moore, P.; Tchounwou, P. Dose- and time-dependent response of human leukemia (HL-60) cells to arsenic trioxide treatment. Int. J. Environ. Res. Public Health 2006, 3, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Gregory, C.D.; Pound, J.D.; Devitt, A.; Wilson-Jones, M.; Ray, P.; Murray, R.J. Inhibitory effects of persistent apoptotic cells on monoclonal antibody production in vitro: Simple removal of non-viable cells improves antibody productivity by hybridoma cells in culture. mAbs 2009, 1, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.E.; Chen, J.; Chan, J.R.; Langen, R. Engineering a polarity-sensitive biosensor for time-lapse imaging of apoptotic processes and degeneration. Nat. Meth. 2010, 7, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.E.; Chen, J.; Langen, R.; Chan, J.R. Monitoring apoptosis and neuronal degeneration by real-time detection of phosphatidylserine externalization using a polarity-sensitive indicator of viability and apoptosis. Nat. Protoc. 2010, 5, 1396–1405. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Bexiga, M.G.; Anguissola, S.; Boya, P.; Simpson, J.C.; Salvati, A.; Dawson, K.A. Time resolved study of cell death mechanisms induced by amine-modified polystyrene nanoparticles. Nanoscale 2013, 5, 10868–10876. [Google Scholar] [CrossRef] [PubMed]




© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Röttgermann, P.J.F.; Dawson, K.A.; Rädler, J.O. Time-Resolved Study of Nanoparticle Induced Apoptosis Using Microfabricated Single Cell Arrays. Microarrays 2016, 5, 8. https://doi.org/10.3390/microarrays5020008
Röttgermann PJF, Dawson KA, Rädler JO. Time-Resolved Study of Nanoparticle Induced Apoptosis Using Microfabricated Single Cell Arrays. Microarrays. 2016; 5(2):8. https://doi.org/10.3390/microarrays5020008
Chicago/Turabian StyleRöttgermann, Peter J. F., Kenneth A. Dawson, and Joachim O. Rädler. 2016. "Time-Resolved Study of Nanoparticle Induced Apoptosis Using Microfabricated Single Cell Arrays" Microarrays 5, no. 2: 8. https://doi.org/10.3390/microarrays5020008