
Prenatal Ethanol Exposure and Whisker Clipping Disrupt Ultrasonic Vocalizations and Play Behavior in Adolescent Rats


Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Prenatal Exposure
2.3. Postnatal Manipulations
2.4. Ultrasonic Vocalization Recording
2.5. Experimental Design and Data Analysis
3. Results
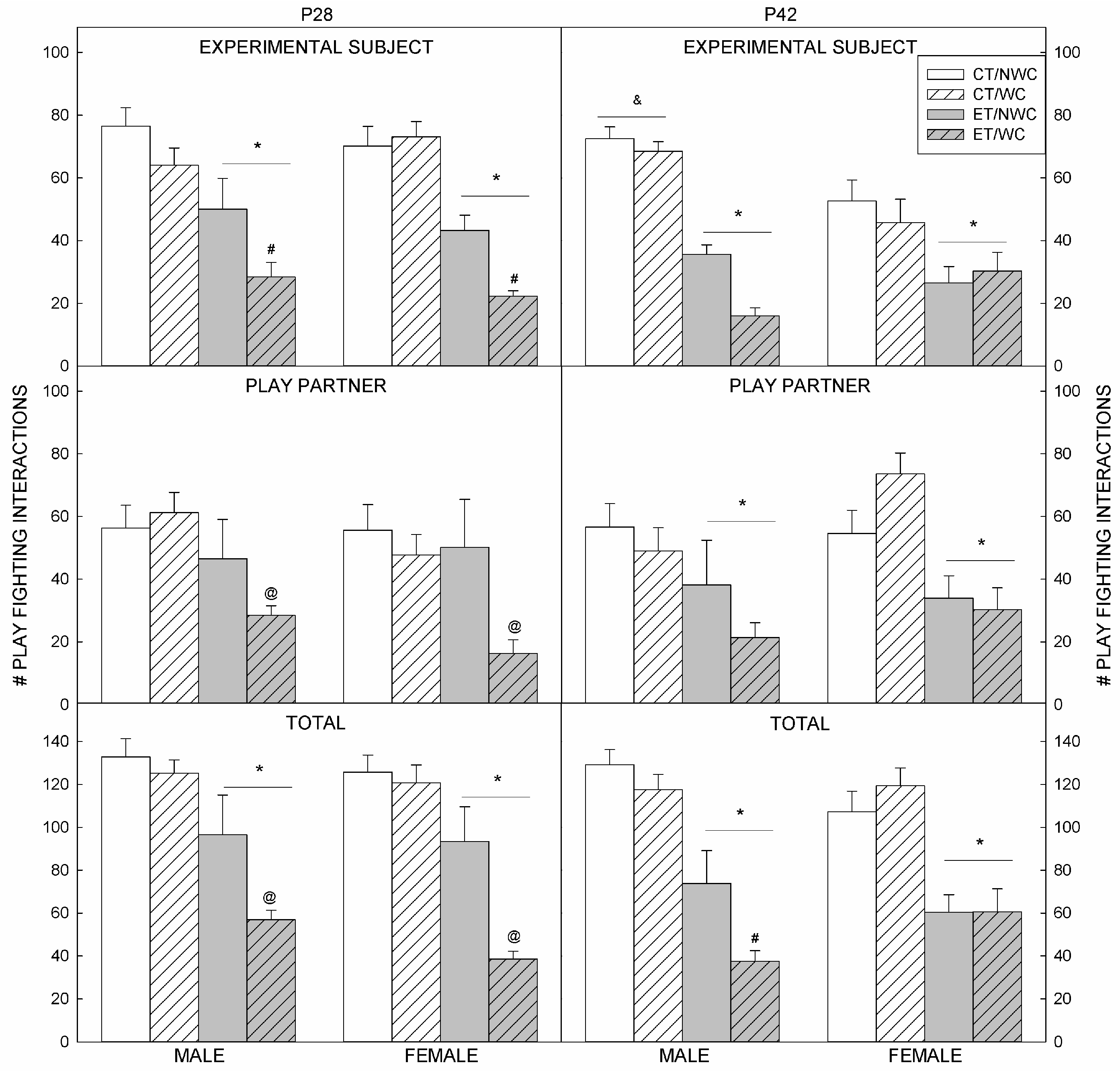
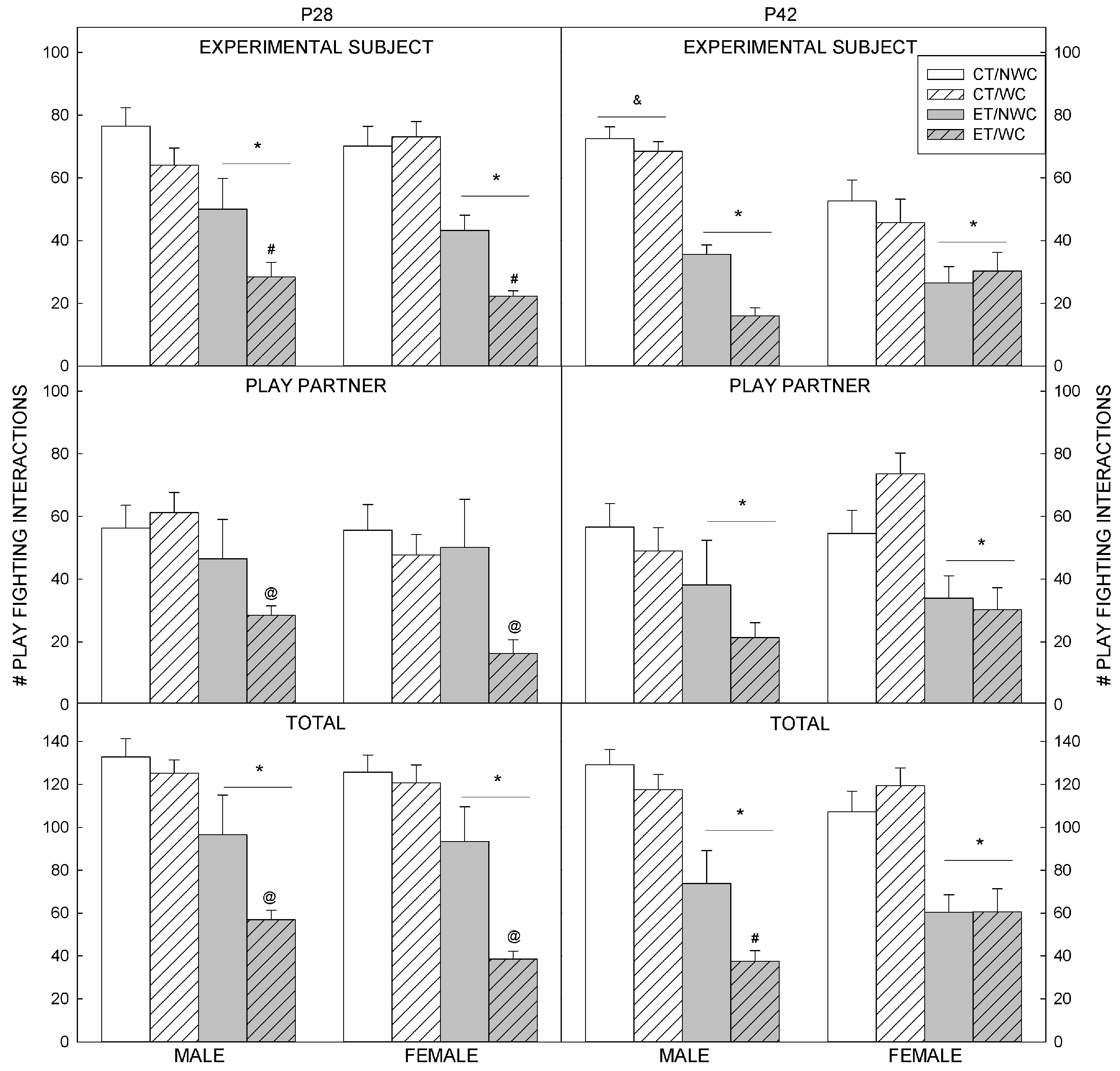
3.1. Play Fighting
3.2. Play Fighting Outcomes at P28
3.3. Play Fighting Outcomes at P42
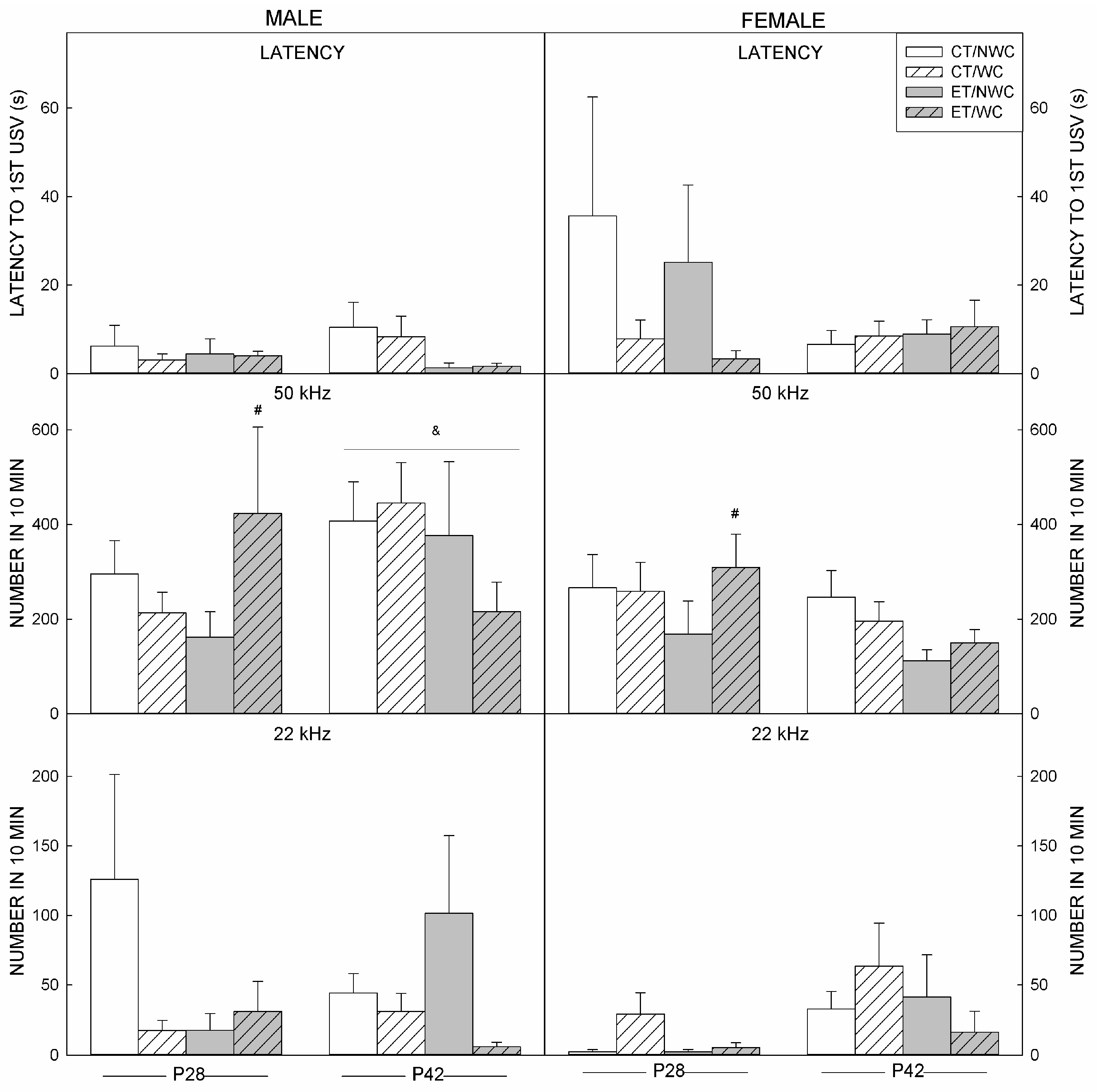
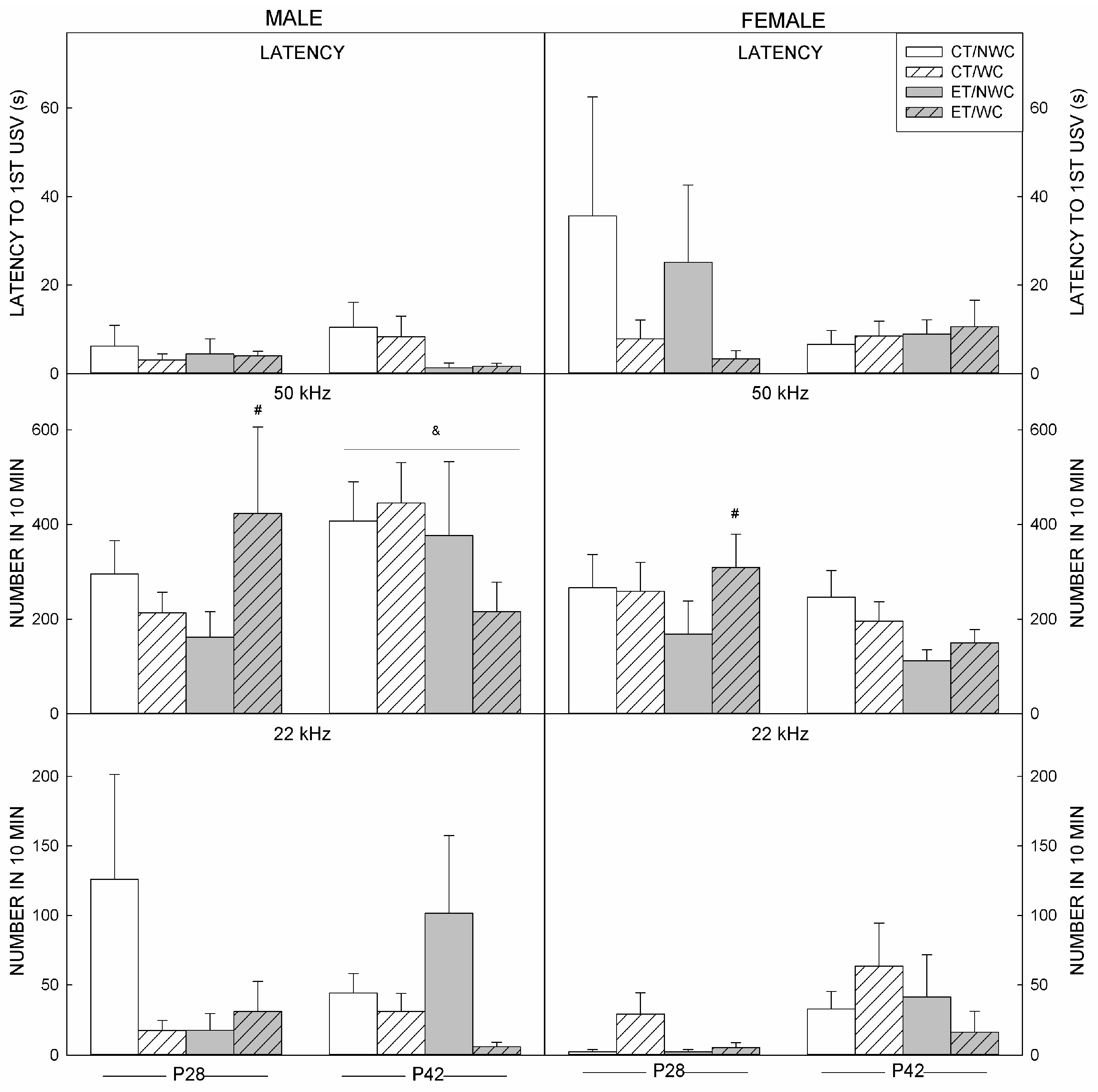
3.4. USV Outcomes at P28
3.5. USV Outcomes at P42
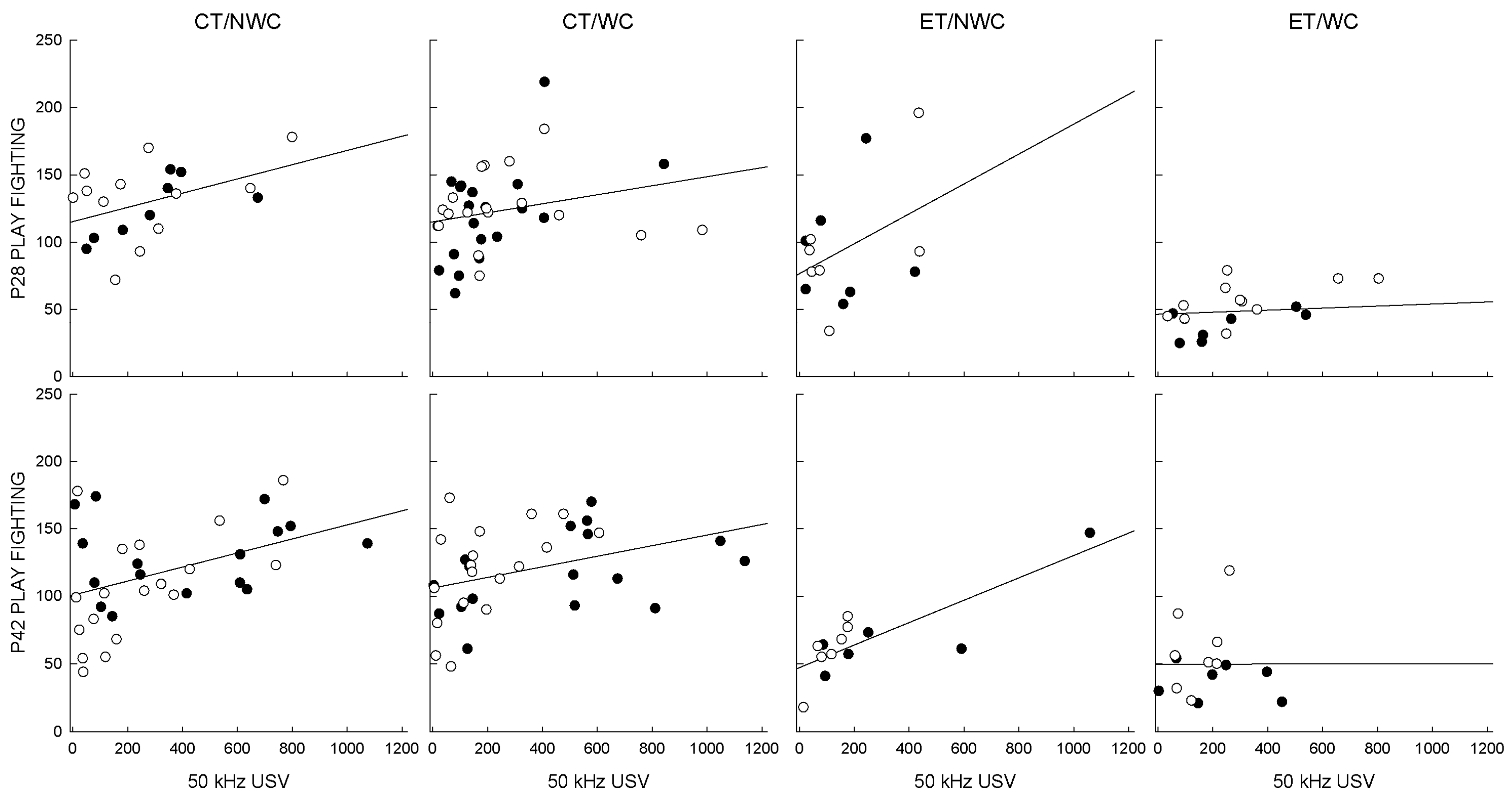
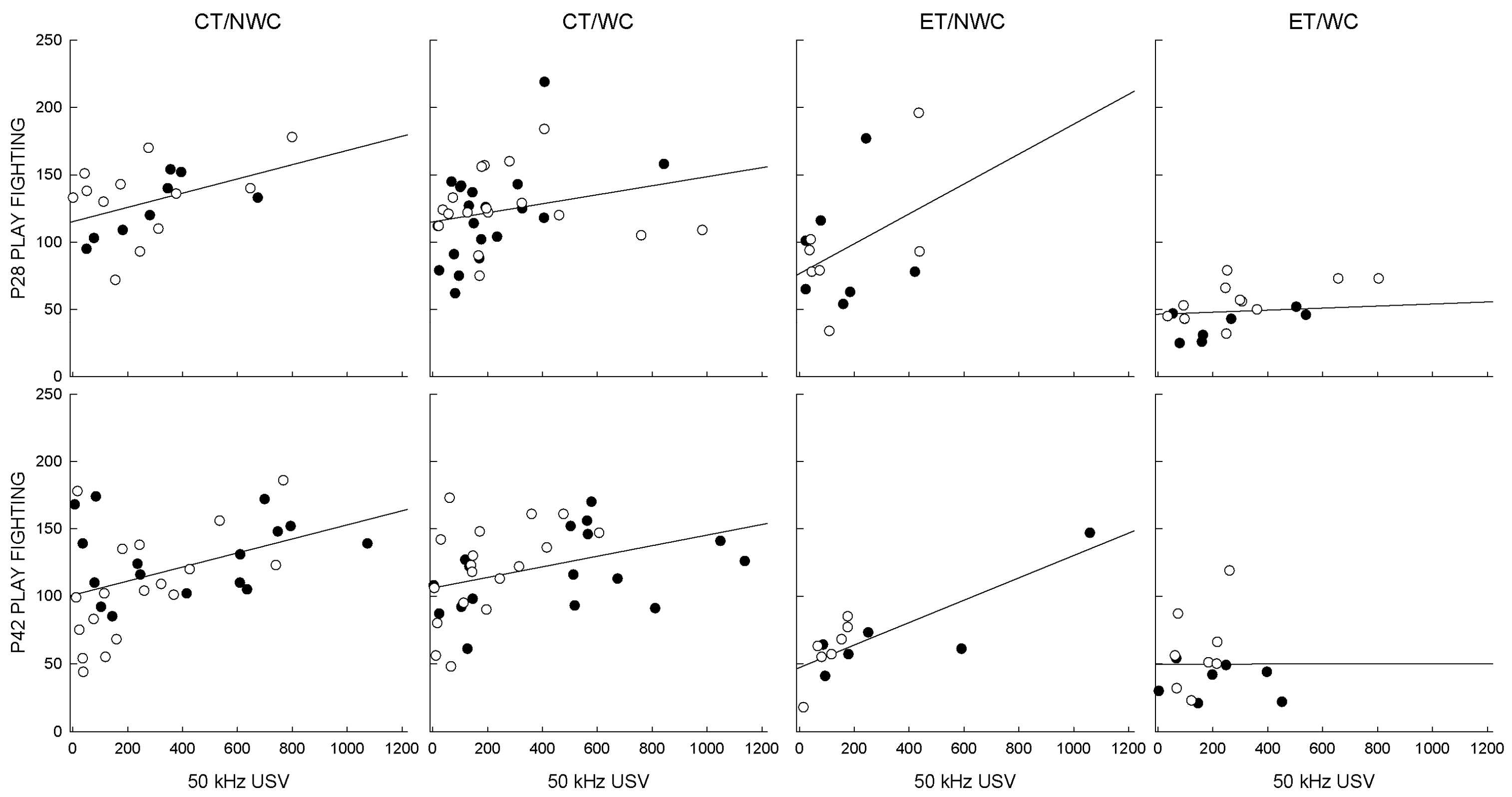
3.6. Correlations
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Brudzynski, S.M. Communication of adult rats by ultrasonic vocalization: Biological, sociobiological, and neuroscience approaches. ILAR J. 2009, 50, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Barfield, R.J.; Thomas, D.A. The role of ultrasonic vocalizations in the regulation of reproduction in rats. Ann. N. Y. Acad. Sci. 1986, 474, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Knutson, B.; Burgdorf, J.; Panksepp, J. Anticipation of play elicits high-frequency ultrasonic vocalizations in young rats. J. Comp. Psychol. 1998, 112, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Burgdorf, J.; Knutson, B.; Panksepp, J. Anticipation of rewarding electrical brain stimulation evokes ultrasonic vocalization in rats. Behav. Neurosci. 2000, 114, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Brudzynski, S.M. Principles of rat communication: Quantitative parameters of ultrasonic calls in rats. Behav. Genet. 2005, 35, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Vivian, J.A.; Miczek, K.A. Ultrasounds during morphine withdrawal in rats. Psychopharmacology 1991, 104, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Mutschler, N.H.; Miczek, K.A. Withdrawal from iv cocaine "binges" in rats: Ultrasonic distress calls and startle. Psychopharmacology 1998, 135, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Sales, G.D. Ultrasound and aggressive behaviour in rats and other small mammals. Anim. Behav. 1972, 20, 88–100. [Google Scholar] [CrossRef]
- Himmler, B.T.; Kisko, T.M.; Euston, D.R.; Kolb, B.; Pellis, S.M. Are 50-kHz calls used as play signals in the playful interactions of rats? I. Evidence from the timing and context of their use. Behav. Process. 2014, 106, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Brunelli, S.A.; Nie, R.; Whipple, C.; Winiger, V.; Hofer, M.A.; Zimmerberg, B. The effects of selective breeding for infant ultrasonic vocalizations on play behavior in juvenile rats. Physiol. Behav. 2006, 87, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Burgdorf, J.; Moskal, J.R.; Brudzynski, S.M.; Panksepp, J. Rats selectively bred for low levels of play-induced 50 kHz vocalizations as a model for Autism Spectrum Disorders: A role for NMDA receptors. Behav. Brain Res. 2013, 251, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Pellis, S.M.; Pellis, V.C. On knowing it’s only play: The role of play signals in play fighting. Aggress. Violent Behav. 1996, 1, 249–268. [Google Scholar] [CrossRef]
- Pellis, S.M.; Pellis, V.C. Play fighting of rats in comparative perspective: A schema for neurobehavioral analyses. Neurosci. Biobehav. Rev. 1998, 23, 87–101. [Google Scholar] [CrossRef]
- Wilson, S. The development of social behaviour in the vole (Microtus agrestis). Zool. J. Linn. Soc. 1973, 52, 45–62. [Google Scholar] [CrossRef]
- Meyer, L.S.; Riley, E.P. Social play in juvenile rats prenatally exposed to alcohol. Teratology 1986, 34, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Marino, M.D.; Cronise, K.; Lugo, J.N., Jr.; Kelly, S.J. Ultrasonic vocalizations and maternal-infant interactions in a rat model of fetal alcohol syndrome. Dev. Psychobiol. 2002, 41, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Wellmann, K.A.; George, F.; Brnouti, F.; Mooney, S.M. Docosahexaenoic acid partially ameliorates deficits in social behavior and ultrasonic vocalizations caused by prenatal ethanol exposure. Behav. Brain Res. 2015, 286, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, D.A.; Barto, D.; Rodriguez, C.I.; Magcalas, C.M.; Fink, B.C.; Rice, J.P.; Bird, C.W.; Davies, S.; Savage, D.D. Effects of moderate prenatal ethanol exposure and age on social behavior, spatial response perseveration errors and motor behavior. Behav. Brain Res. 2014, 269, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Lugo, J.N.; Marino, M.D.; Gass, J.T.; Wilson, M.A.; Kelly, S.J. Ethanol exposure during development reduces resident aggression and testosterone in rats. Physiol. Behav. 2006, 87, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Zimmerberg, B.; McDonald, B.C. Prenatal alcohol exposure influences the effects of neuroactive steroids on separation-induced ultrasonic vocalizations in rat pups. Pharmacol. Biochem. Behav. 1996, 55, 541–547. [Google Scholar] [CrossRef]
- Wellmann, K.; Lewis, B.; Barron, S. Agmatine reduces ultrasonic vocalization deficits in female rat pups exposed neonatally to ethanol. Neurotoxicol. Teratol. 2010, 32, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Wellmann, K.A.; Mooney, S.M. Unilateral whisker clipping exacerbates ethanol-induced social and somatosensory behavioral deficits in a sex- and age-dependent manner. Physiol. Behav. 2014, 148, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Willey, A.R.; Varlinskaya, E.I.; Spear, L.P. Social interactions and 50 kHz ultrasonic vocalizations in adolescent and adult rats. Behav. Brain Res. 2009, 202, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.W. Circadian rhythm of cell proliferation in the telencephalic ventricular zone: Effect of in utero exposure to ethanol. Brain Res. 1992, 595, 17–24. [Google Scholar] [CrossRef]
- Youngentob, S.L.; Kent, P.F.; Sheehe, P.R.; Molina, J.C.; Spear, N.E.; Youngentob, L.M. Experience-induced fetal plasticity: The effect of gestational ethanol exposure on the behavioral and neurophysiologic olfactory response to ethanol odor in early postnatal and adult rats. Behav. Neurosci. 2007, 121, 1293–1305. [Google Scholar] [CrossRef] [PubMed]
- Burghardt, G.M. The comparative reach of play and brain: Perspective, evidence, and implications. Am. J. Play 2010, 2, 338–356. [Google Scholar]
- Trezza, V.; Damsteegt, R.; Vanderschuren, L.J. Conditioned place preference induced by social play behavior: Parametrics, extinction, reinstatement and disruption by methylphenidate. Eur. Neuropsychopharmacol. 2009, 19, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Vanderschuren, L.J.; Trezza, V. What the laboratory rat has taught us about social play behavior: Role in behavioral development and neural mechanisms. Curr. Top. Behav. Neurosci. 2014, 16, 189–212. [Google Scholar] [PubMed]
- Baarendse, P.J.; Counotte, D.S.; O’Donnell, P.; Vanderschuren, L.J. Early social experience is critical for the development of cognitive control and dopamine modulation of prefrontal cortex function. Neuropsychopharmacology 2013, 38, 1485–1494. [Google Scholar] [CrossRef] [PubMed]
- Zeeb, F.D.; Robbins, T.W.; Winstanley, C.A. Serotonergic and dopaminergic modulation of gambling behavior as assessed using a novel rat gambling task. Neuropsychopharmacology 2009, 34, 2329–2343. [Google Scholar] [CrossRef] [PubMed]
- Einon, D.F.; Morgan, M.J.; Kibbler, C.C. Brief periods of socialization and later behavior in the rat. Dev. Psychobiol. 1978, 11, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Einon, D.F.; Morgan, M.J. Early isolation produces enduring hyperactivity in the rat, but no effect upon spontaneous alternation. Q. J. Exp. Psychol. 1978, 30, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Marquardt, K.; Brigman, J.L. The impact of prenatal alcohol exposure on social, cognitive and affective behavioral domains: Insights from rodent models. Alcohol 2016, 51, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kisko, T.M.; Euston, D.R.; Pellis, S.M. Are 50-kHz calls used as play signals in the playful interactions of rats? III. The effects of devocalization on play with unfamiliar partners as juveniles and as adults. Behav. Process. 2015, 113, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Royalty, J. Effects of prenatal ethanol exposure on juvenile play-fighting and postpubertal aggression in rats. Psychol. Rep. 1990, 66, 551–560. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, T.K.; Barfield, R.J.; Geyer, L.A. Ultrasonic vocalisations facilitate sexual behaviour of female rats. Nature 1978, 272, 163–164. [Google Scholar] [CrossRef] [PubMed]
- Willadsen, M.; Seffer, D.; Schwarting, R.K.; Wohr, M. Rodent ultrasonic communication: Male prosocial 50-kHz ultrasonic vocalizations elicit social approach behavior in female rats (Rattus norvegicus). J. Comp. Psychol. 2014, 128, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Ward, I.L.; Ward, O.B.; Winn, R.J.; Bielawski, D. Male and female sexual behavior potential of male rats prenatally exposed to the influence of alcohol, stress, or both factors. Behav. Neurosci. 1994, 108, 1188–1195. [Google Scholar] [CrossRef] [PubMed]
- Simons, D.J.; Land, P.W. Early experience of tactile stimulation influences organization of somatic sensory cortex. Nature 1987, 326, 694–697. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.J.; Chen, W.J.; Chuang, Y.W.; Wang, Y.C. Neonatal whisker trimming causes long-lasting changes in structure and function of the somatosensory system. Exp. Neurol. 2009, 219, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, J.L.; Salazar, E. Effects of whisker trimming on GABAA receptor binding in the barrel cortex of developing and adult rats. J. Comp. Neurol. 1998, 395, 209–216. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| - | CT/NWC | CT/WC | ET/NWC | ET/WC |
|---|---|---|---|---|
| P28 M | 8 (3 CH/5 PF) | 19 (10 CH/9 PF) | 7 | 8 |
| P28 F | 12 (5 CH/7 PF) | 18 (10 CH/8 PF) | 7 | 11 |
| P42 M | 16 (8 CH/8 PF) | 17 (9 CH/8 PF) | 6 | 7 |
| P42 F | 18 (9 CH/9 PF) | 18 (9 CH/9 PF) | 7 | 8 |
| Age | Condition | n | Pearson’s r | p |
|---|---|---|---|---|
| P28 | CT/NWC | 20 | 0.441 | 0.051 * |
| CT/WC | 37 | 0.240 | 0.152 | |
| ET/NWC | 14 | 0.396 | 0.162 | |
| ET/WC | 19 | 0.180 | 0.461 | |
| P42 | CT/NWC | 34 | 0.416 | 0.014 * |
| CT/WC | 35 | 0.367 | 0.030 * | |
| ET/NWC | 13 | 0.808 | 0.001 * | |
| ET/WC | 15 | 0.001 | 0.998 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Waddell, J.; Yang, T.; Ho, E.; Wellmann, K.A.; Mooney, S.M. Prenatal Ethanol Exposure and Whisker Clipping Disrupt Ultrasonic Vocalizations and Play Behavior in Adolescent Rats. Brain Sci. 2016, 6, 43. https://doi.org/10.3390/brainsci6040043
Waddell J, Yang T, Ho E, Wellmann KA, Mooney SM. Prenatal Ethanol Exposure and Whisker Clipping Disrupt Ultrasonic Vocalizations and Play Behavior in Adolescent Rats. Brain Sciences. 2016; 6(4):43. https://doi.org/10.3390/brainsci6040043
Chicago/Turabian StyleWaddell, Jaylyn, Tianqi Yang, Eric Ho, Kristen A. Wellmann, and Sandra M. Mooney. 2016. "Prenatal Ethanol Exposure and Whisker Clipping Disrupt Ultrasonic Vocalizations and Play Behavior in Adolescent Rats" Brain Sciences 6, no. 4: 43. https://doi.org/10.3390/brainsci6040043