
REM-Enriched Naps Are Associated with Memory Consolidation for Sad Stories and Enhance Mood-Related Reactivity

 ,
, 
Abstract
:1. Introduction
2. Experimental Section
2.1. Participants
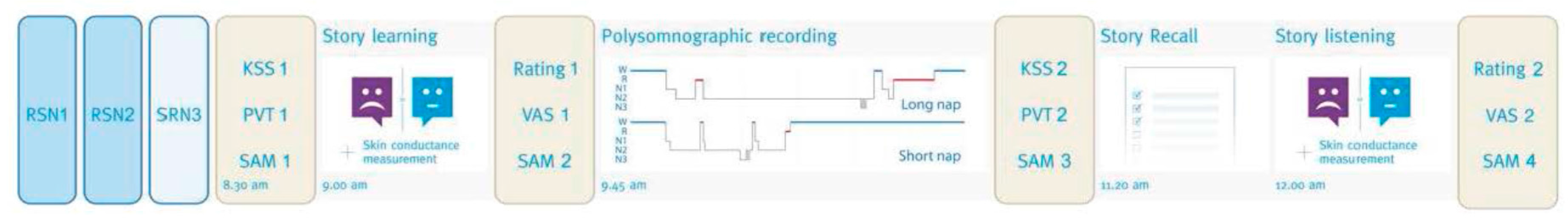
2.2. Procedure
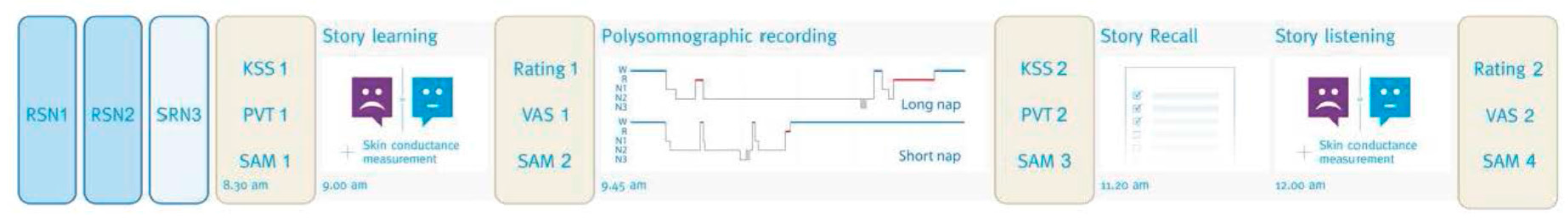
2.3. Material
2.3.1. Stories and Questionnaires
2.3.2. Skin Conductance Measurement
2.3.3. Polysomnography
2.4. Data Analysis
2.4.1. Actigraphic Data
2.4.2. Sleep Data
2.4.3. Statistical Analyses
3. Results
3.1. Sleep and Vigilance
3.1.1. Nocturnal Sleep Prior to Experimental Sessions

{kind=link}
{kind=link}
{kind=link}
| Group | Story | Night 1 (RSN) | Night 2 (RSN) | Night 3 (SRN) | |
|---|---|---|---|---|---|
| TST (agenda, in hours) | Short nap | Neutral story | 8.21 ± 1.2 | 8.08 ± 1 | 5.83 ± 1 * |
| Sad story | 7.67 ± 1.7 | 7.75 ± 1.7 | 5.58 ± 1.2 * | ||
| Long nap | Neutral story | 7.89 ± 1.1 | 7.25 ± 0.8 | 5 ± 0.9 * | |
| Sad story | 7.5 ± 1.5 | 7.89 ± 1.7 | 5.78 ± 1 * | ||
| TST (actimetry, in hours) | Short nap | Neutral story | 8.58 ± 0.8 | 8.47 ± 1.1 | 6.26 ± 0.7 * |
| Sad story | 8.82 ± 2.9 | 8.69 ± 2 | 6.11 ± 1 * | ||
| Long nap | Neutral story | 8.15 ± 2.1 | 7.87 ± 1 | 6.14 ± 2.3 * | |
| Sad story | 8.31 ± 1.7 | 8.33 ± 1.4 | 6.47 ± 1.7 * |
3.1.2. Sleep during Long and Short Naps
| Long Nap Condition (N = 12) | Short Nap Condition (N = 12) | Nap | Story | Interaction | |||
|---|---|---|---|---|---|---|---|
| Sad Story | Neutral Story | Sad Story | Neutral Story | p Value | p Value | p Value | |
| W (min) | 24.41 (± 11.9) | 21.25 (± 15.2) | 30.29 (± 35.6) | 27.25 (± 24.7) | 0.496 | 0.498 | 0.989 |
| N1 (min) | 11 (± 6.1) | 11.71 (± 5.9) | 5.5 (± 4) | 7.66 (± 4.3) | 0.008 | 0.290 | 0.588 |
| N2 (min) | 48.96 (± 16.2) | 51.83 ± 14.5) | 29.25 (± 11.6) | 34.5 (± 9.2) | 0.001 | 0.156 | 0.672 |
| N3 (min) | 11.58 (± 15) | 6.21 (± 8.4) | 7.92 (± 9.5) | 1.66 (± 2.7) | 0.225 | 0.022 | 0.855 |
| REM (min) | 17.67 (± 9.3) | 21.33 (± 11.3) | 3.42 (± 7.2) | 0.87 (± 1.6) | 0.000 | 0.815 | 0.204 |
| SPT (min) | 113.62 (± 12.6) | 112.33 (± 8) | 76.375 (± 30.9) | 71.96 (± 22.4) | 0.000 | 0.524 | 0.726 |
| TST (min) | 89.21 (± 11) | 91.08 (± 11.9) | 46.08 (± 15.3) | 44.71 (± 10.2) | 0.000 | 0.935 | 0.578 |
| SOL (min) | 13.5 (± 8.9) | 9.17 (± 5.1) | 9.42 (± 11.3) | 8 (± 6.4) | 0.404 | (0.052) | 0.308 |
| WASO (min) | 10.92 (± 9.5) | 12.125 (± 10.74) | 20.87 (± 30.6) | 16.27 (± 17.7) | 0.365 | 0.922 | 0.726 |
| Micro arousals | 3.41 (± 0.9) | 3.25 (± 0.7) | 2.17 (± 0.9) | 1.92 (± 0.7) | 0.175 | 0.719 | 0.942 |
| SE (%) | 78.88 (± 9.6) | 81.54 (± 12.1) | 67.82 (± 26.2) | 66.56 (± 21) | (0.07) | 0.827 | 0.544 |
| Total Nr of REMs | 139.25 (± 114.7) | 142.67 (± 74.4) | 30 (± 59.9) | 9.08 (± 14.6) | 0.000 | 0.684 | 0.572 |
| REM sleep density | 6.83 (± 3.8) | 8.27 (± 6.2) | 2.56 (± 4.4) | 2.94 (± 3.9) | 0.005 | 0.433 | 0.645 |
| Total Nr of Slow Spindles | 47.5 (± 21.6) | 59 (± 40.4) | 17.67 (± 15.9) | 20.92 (± 18.4) | 0.001 | 0.213 | 0.480 |
| Total Nr of Fast Spindles | 77.83 (± 52.5) | 99.42 (± 68.8) | 38.58 (± 44.8) | 55.42 (± 68.4) | (0.061) | 0.125 | 0.845 |
| Total Nr of Slow & Fast spindles | 125.33 (± 67.4) | 158.42 (± 100.2) | 57.08 (± 56.3) | 76.42 (± 81.4) | 0.012 | 0.125 | 0.679 |
| Density of slow spindles | 1.09 (± 0.6) | 1.28 (± 1.3) | 0.58 (± 0.4) | 0.61 (± 0.5) | 0.019 | 0.633 | 0.735 |
| Density of fast spindles | 1.82 (± 2.4) | 1.99 (± 1.6) | 1.32 (± 1.1) | 1.44 (± 1.3) | (0.096) | 0.194 | 0.431 |
| Density of slow and fast spindles | 3.11 (± 2.9) | 3.27 (± 2.8) | 1.72 (± 1.2) | 2.06 (± 1.6) | (0.051) | 0.714 | 0.895 |
3.1.3. Sleepiness and Vigilance before and after the Nap (Table 3)
| Group | Story | Before Nap | After Nap | |
|---|---|---|---|---|
| KSS mean scores | Short nap | Neutral story | 5.58 ± 2.2 | 2.75 ± 1.5 |
| Sad story | 5.58 ± 2.1 | 3.33 ± 1.9 | ||
| Long nap | Neutral story | 5.58 ± 1.7 | 3.33 ± 1.3 | |
| Sad story | 5.66 ± 2.2 | 3.75 ± 1.5 | ||
| PVT mean 1/reaction time | Short nap | Neutral story | 0.0030 ± 0.0003 | 0.0031 ± 0.0004 |
| Sad story | 0.0029 ± 0.0003 | 0.0029 ± 0.0005 | ||
| Long nap | Neutral story | 0.0031 ± 0.0005 | 0.0033 ± 0.0004 | |
| Sad story | 0.0031 ± 0.0004 | 0.0032 ± 0.0004 | ||
| PVT Coefficient of variation (%) | Short nap | Neutral story | 17.17 ± 4.4 | 15.11 ± 4.1 |
| Sad story | 17.87 ± 3.7 | 18.40 ± 5.9 | ||
| Long nap | Neutral story | 17.51 ± 6.3 | 15.77 ± 3.5 | |
| Sad story | 18.95 ± 8.8 | 16.28 ± 4.1 |
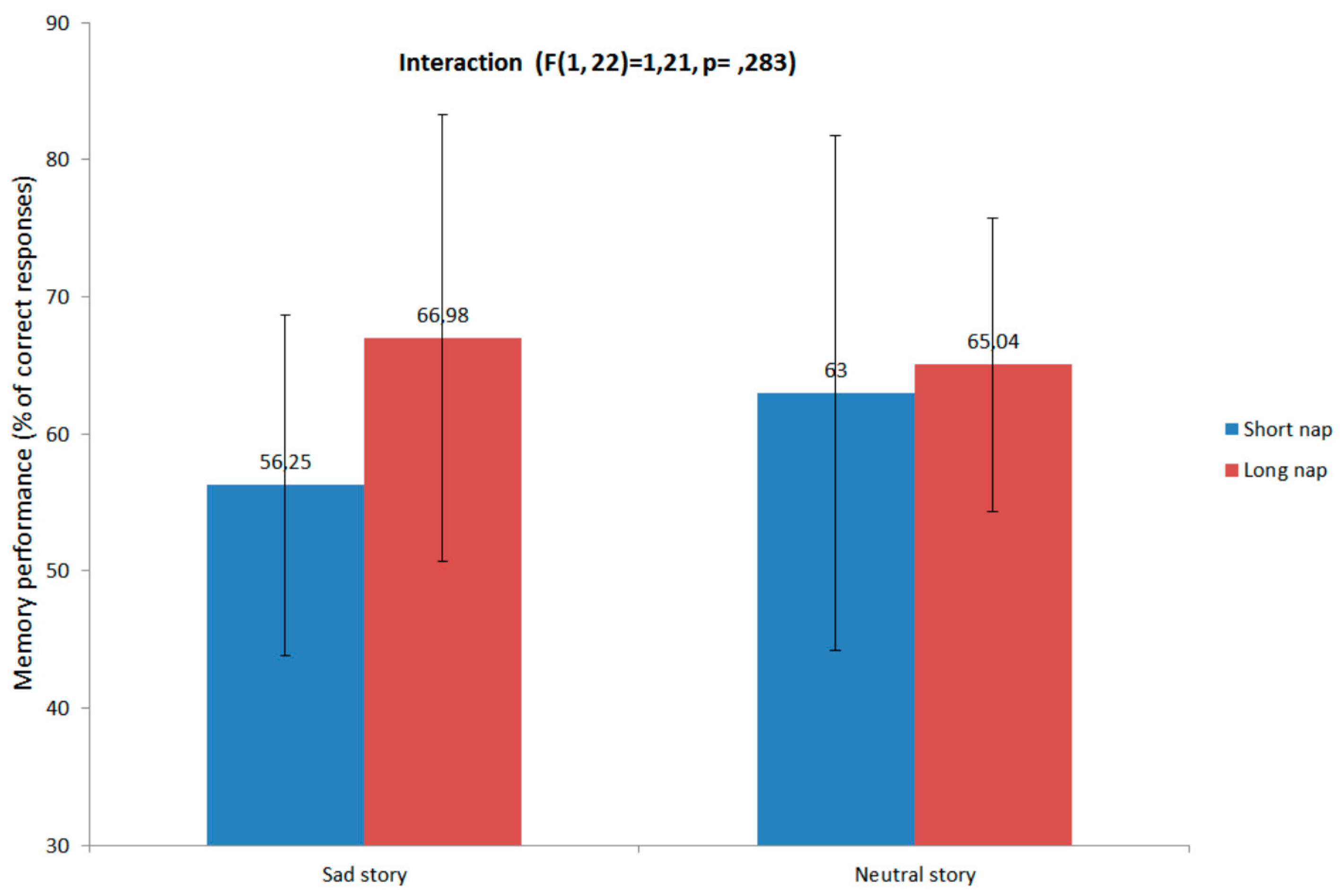
3.2. Memory Performance and Relation with Sleep
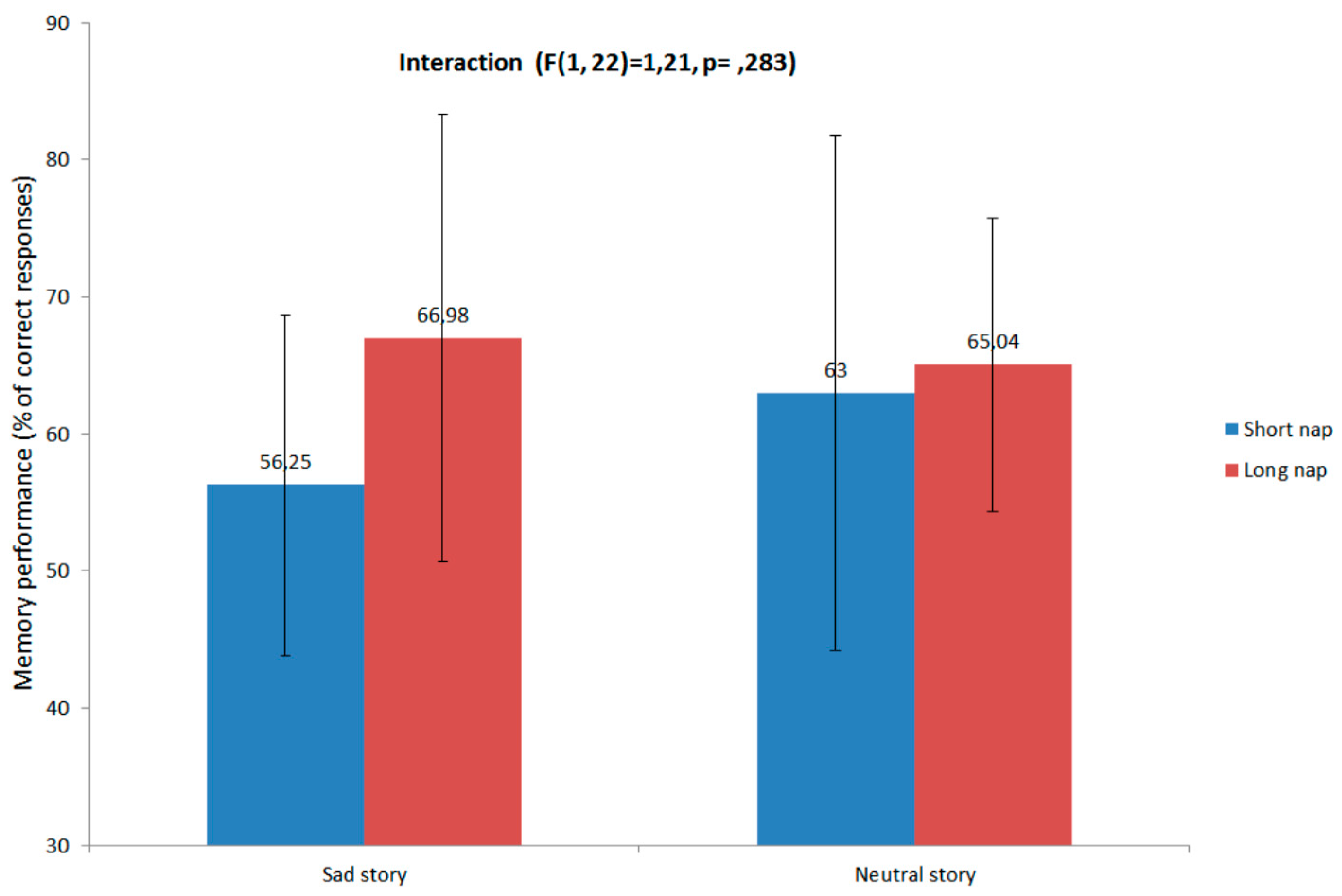
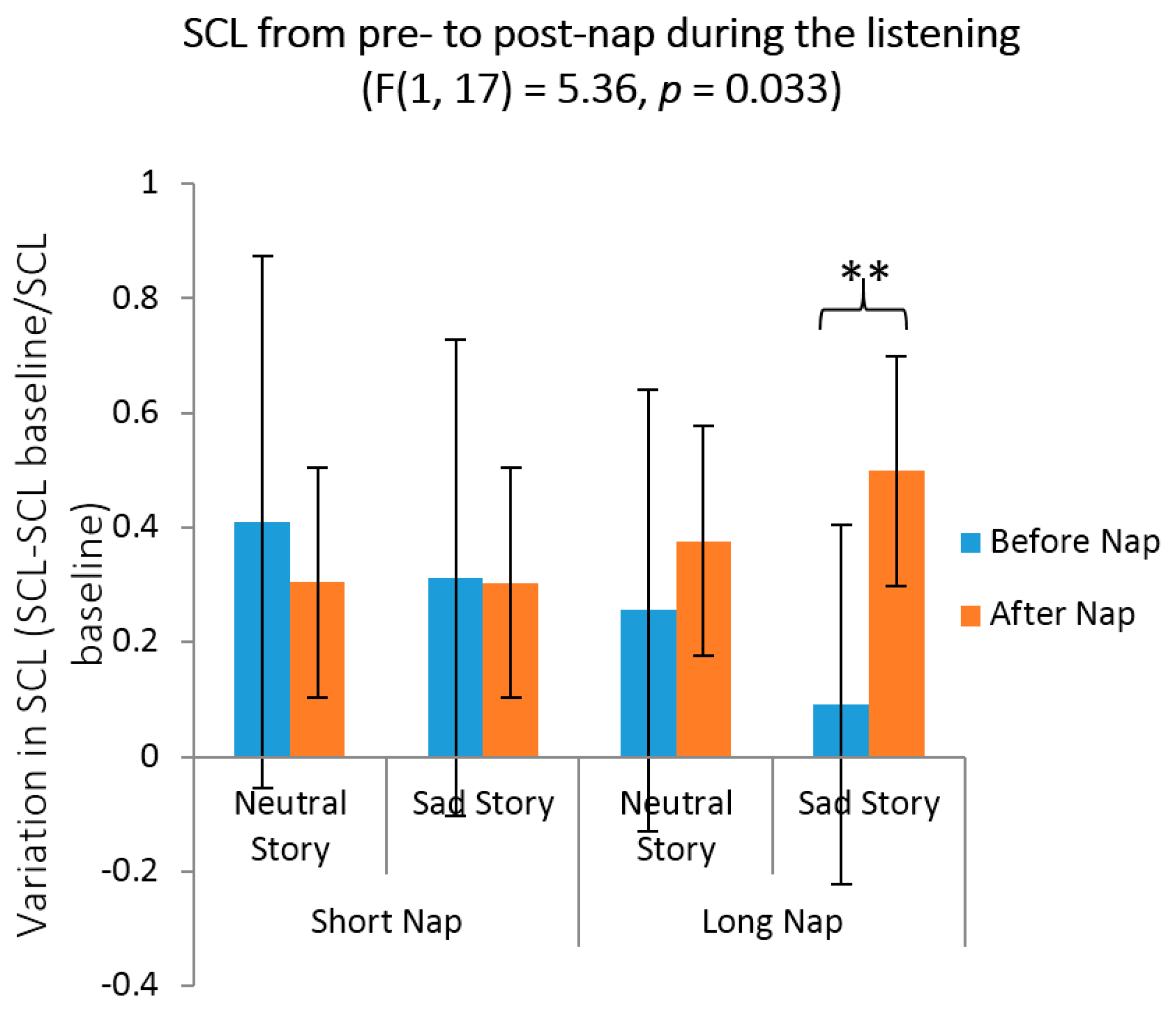
3.3. Emotional Reactivity, Mood and Arousal Measures
| Questionnaire | Nap | Story | SAM 1 | SAM 2 | SAM 3 | SAM 4 |
|---|---|---|---|---|---|---|
| Self-Assessment Manikin mean mood scores | Short nap | Neutral story | 0.83 ± 1 | 0.5 ± 1.7 | 2.08 ± 1.1 | 1.33 ± 1.3 |
| Sad story | 1.4 ± 1 | 0.08 ± 1.9 | 0.42 ± 1.7 | 0.83 ± 1.6 | ||
| Long nap | Neutral story | 1.16 ± 1.6 | 1.33 ± 1.7 | 2 ± 1.3 | 1.66 ± 1.6 | |
| Sad story | 1.5 ± 0.9 | −0.58 ± 1.2 | 1.58 ± 0.8 | 0 ± 1.8 | ||
| Self-Assessment Manikin mean arousal scores | Short nap | Neutral story | 1.66 ± 0.8 | 1.5 ± 0.8 | 1.75 ± 0.8 | 1.66 ± 0.7 |
| Sad story | 0.75 ± 1.2 | 1.58 ± 1 | 0.16 ± 1.3 | 0.25 ± 1.4 | ||
| Long nap | Neutral story | 2.33 ± 1 | 1.91 ± 0.9 | 2.2 ± 1 | 2.1 ± 1 | |
| Sad story | 0.83 ± 1.7 | 1.25 ± 1.2 | 0.16 ± 1.8 | 1 ± 1.5 |
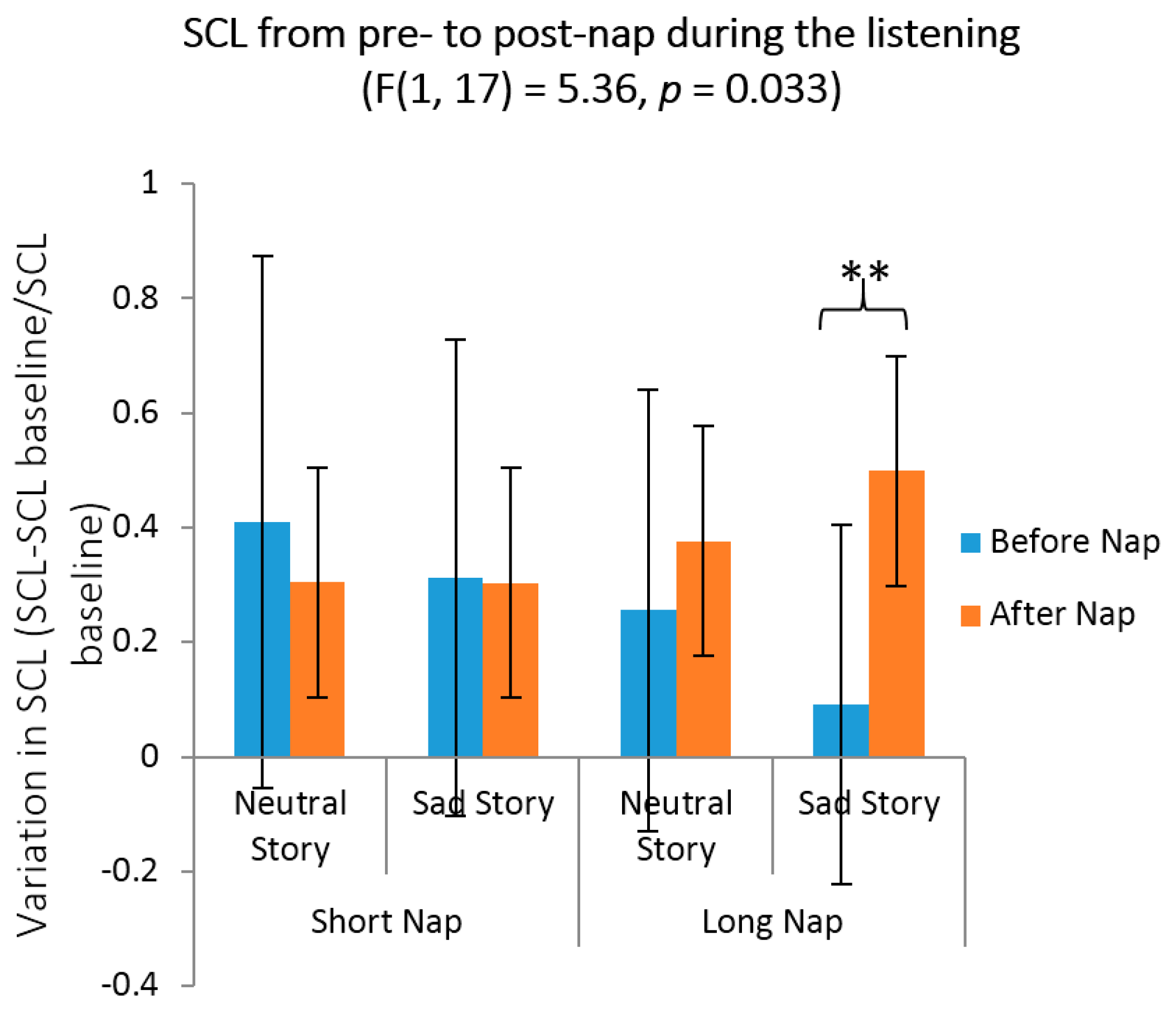
3.4. Affective Rating for Neutral and Sad Stories before vs. after the Nap
4. Discussion
4.1. The Influence of Sleep on Memory Consolidation
4.2. Objective and Self-Reported Emotional Reactivity
4.3. Emotional Rating of Neutral and Sad Stories
4.4. Limitations
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- McGaugh, J.L. Time-dependent processes in memory storage. Science 1966, 153, 1351–1358. [Google Scholar] [CrossRef] [PubMed]
- Cahill, L. Neurobiological mechanisms of emotionally influenced, long-term memory. Prog. Brain Res. 2000, 126, 29–37. [Google Scholar] [PubMed]
- McGaugh, J.L. The amygdala modulates the consolidation of memories of emotionally arousing experiences. Annu. Rev. Neurosci. 2004, 27, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Ochsner, K.N. Are affective events richly recollected or simply familiar? The experience and process of recognizing feelings past. J. Exp. Psychol. Gen. 2000, 129, 242–261. [Google Scholar] [CrossRef] [PubMed]
- Peigneux, P.; Smith, C. Memory processing in relation to sleep. In Principles and Practice of Sleep Medicine, 5th ed.; Kryger, M., Ed.; Elsevier: Philadelphia, PA, USA, 2010; pp. 335–347. [Google Scholar]
- Rasch, B.; Born, J. About sleep’s role in memory. Physiol. Rev. 2013, 93, 681–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, M.P. Sleep, memory and emotion. Prog. Brain Res. 2010, 185, 49–68. [Google Scholar] [PubMed]
- Deliens, G.; Gilson, M.; Peigneux, P. Sleep and the processing of emotions. Exp. Brain Res. 2014, 232, 1403–1414. [Google Scholar] [CrossRef] [PubMed]
- Genzel, L.; Spoormaker, V.I.; Konrad, B.N.; Desler, M. The role of rapid eye movement sleep for amygdala-related memory processing. Neurobiol. Learn. Mem. 2015, 122, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Baglioni, C.; Spiegelhalder, K.; Lombardo, C.; Riemann, D. Sleep and emotions: A focus on insomnia. Sleep Med. Rev. 2010, 14, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Bower, B.; Bylsma, L.M.; Morris, B.H.; Rottenberg, J. Poor reported sleep quality predicts low positive affect in daily life among healthy and mood-disordered persons. J. Sleep Res. 2010, 19, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Mauss, I.B.; Troy, A.S.; LeBourgeois, M.K. Poorer sleep quality is associated with lower emotion-regulation ability in a laboratory paradigm. Cogn. Emot. 2013, 27, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Franzen, P.L.; Buysse, D.J.; Dahl, R.E.; Thompson, W.; Siegle, G.J. Sleep deprivation alters pupillary reactivity to emotional stimuli in healthy young adults. Biol. Psychol. 2009, 80, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.S.; Gujar, N.; Hu, P.; Jolesz, F.A.; Walker, M.P. The human emotional brain without sleep—A prefrontal amygdala disconnect. Curr. Biol. 2007, 17, R877–R878. [Google Scholar] [CrossRef] [PubMed]
- Lang, P.J.; Bradley, M.M.; Cuthbert, B.N. International Affective Picture System (IAPS): Affective Ratings of Pictures and Instruction Manual; Technical Report A-6; University of Florida: Gainesville, FL, USA, 2005. [Google Scholar]
- Rosales-Lagarde, A.; Armony, J.L.; del Río-Portilla, Y.; Trejo-Martínez, D.; Conde, R.; Corsi-Cabrera, M. Enhanced emotional reactivity after selective REM sleep deprivation in humans: An fMRI study. Front. Behav. Neurosci. 2012, 6, 25. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, T.J.; Crowell, C.R.; Alger, S.E.; Kensinger, E.A.; Villano, M.; Mattingly, S.M.; Payne, J.D. Psychophysiological arousal at encoding leads to reduced reactivity but enhanced emotional memory following sleep. Neurobiol. Learn. Mem. 2014, 114, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Wagner, U.; Fischer, S.; Born, J. Changes in emotional responses to aversive pictures across periods rich in slow-wave sleep versus rapid eye movement sleep. Psychosom. Med. 2002, 64, 627–634. [Google Scholar] [PubMed]
- Werner, G.G.; Schabus, M.; Blechert, J.; Kolodyazhniy, V.; Wilhelm, F.H. Pre- to postsleep change in psychophysiological reactivity to emotional films: Late-night REM sleep is associated with attenuated emotional processing. Psychophysiology 2015, 52, 813–825. [Google Scholar] [CrossRef] [PubMed]
- Talamini, L.M.; Bringmann, L.F.; de Boer, M.; Hofman, W.F. Sleeping worries away or worrying away sleep? Physiological evidence on sleep-emotion interactions. PLoS ONE 2013, 8, e62480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pace-Schott, E.F.; Shepherd, E.; Spencer, R.; Marcello, M.; Tucker, M.; Propper, R.E.; Stickgold, R. Napping promotes inter-session habituation to emotional stimuli. Neurobiol. Learn. Mem. 2011, 95, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Lara Carrasco, J.; Nielsen, T.A.; Solomonova, E.; Levrier, K.; Popova, A. Overnight emotional adaptation to negative stimuli is altered by REM sleep deprivation and is correlated with intervening dream emotions. J. Sleep Res. 2009, 18, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Stylos-Allan, M.; Walker, M.P. Sleep facilitates consolidation of emotional declarative memory. Psychol. Sci. 2006, 17, 891–898. [Google Scholar] [CrossRef] [PubMed]
- Payne, J.D.; Stickgold, R.; Swanberg, K.; Kensinger, E.A. Sleep preferentially enhances memory for emotional components of scenes. Psychol. Sci. 2008, 19, 781–788. [Google Scholar] [CrossRef] [PubMed]
- Wagner, U.; Hallschmid, M.; Rasch, B.; Born, J. Brief sleep after learning keeps emotional memories alive for years. Biol. Psychiatry 2006, 60, 788–790. [Google Scholar] [CrossRef] [PubMed]
- Atienza, M.; Cantero, J.L. Modulatory effects of emotion and sleep on recollection and familiarity. J. Sleep Res. 2008, 17, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Van Heugten-van der Kloet, D.; Giesbrecht, T.; Merckelbach, H. Sleep loss increases dissociation and affects memory for emotional stimuli. J. Behav. Ther. Exp. Psychiatry 2014, 47, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Sterpenich, V.; Albouy, G.; Boly, M.; Vandewalle, G.; Darsaud, A.; Balteau, E.; Dang-Vu, T.T.; Desseilles, M.; D’Argembeau, A.; Gais, S.; et al. Sleep-related hippocampo-cortical interplay during emotional memory recollection. PLoS Biol. 2007, 5, e282. [Google Scholar] [CrossRef] [PubMed]
- Wagner, U.; Gais, S.; Born, J. Emotional memory formation is enhanced across sleep intervals with high amounts of rapid eye movement sleep. Learn. Mem. 2001, 8, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Nishida, M.; Pearsall, J.; Buckner, R.L.; Walker, M.P. REM sleep, prefrontal theta, and the consolidation of human emotional memory. Cereb. Cortex 2009, 19, 1158–1166. [Google Scholar] [CrossRef] [PubMed]
- Wagner, U.; Degirmenci, M.; Drosopoulos, S.; Perras, B.; Born, J. Effects of cortisol suppression on sleep-associated consolidation of neutral and emotional memory. Biol. Psychiatry 2005, 58, 885–893. [Google Scholar] [CrossRef] [PubMed]
- Baran, B.; Pace-Schott, E.F.; Ericson, C.; Spencer, R.M.C. Processing of emotional reactivity and emotional memory over sleep. J. Neurosci. 2012, 32, 1035–1042. [Google Scholar] [CrossRef] [PubMed]
- Groch, S.; Wilhelm, I.; Diekelmann, S.; Born, J. The role of REM sleep in the processing of emotional memories: Evidence from behavior and event-related potentials. Neurobiol. Learn. Mem. 2013, 99, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Benca, R.M.; Obermeyer, W.H.; Thisted, R.A.; Gillin, J.C. Sleep and psychiatric disorders. A meta-analyses. Arch. Gen. Psychiatry 1992, 49, 651–670. [Google Scholar] [CrossRef] [PubMed]
- Cartwright, R.; Luten, A.; Young, M.; Mercer, P.; Bears, M. Role of REM sleep and dream affect in overnight mood regulation: A study of normal volunteers. Psychiatry Res. 1998, 81, 1–8. [Google Scholar] [CrossRef]
- Tinguely, G.; Huber, R.; Borbely, A.A.; Achermann, P. Non-rapid eye movement sleep with low muscle tone as a marker of rapid eye movement sleep regulation. BMC Neurosci. 2006, 7, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deliens, G.; Leproult, R.; Neu, D.; Peigneux, P. Rapid eye movement and non-rapid eye movement sleep contributions in memory consolidation and resistance to retroactive interference for verbal material. Sleep 2013, 36, 1875–1883. [Google Scholar] [CrossRef] [PubMed]
- Buysse, D.J.; Reynolds, C.F.; Monk, T.H.; Berman, S.R.; Kupfer, D.J. The Pittsburgh Sleep Quality Index: A new instrument for psychiatric practice and research. Psychiatry Res. 1989, 28, 193–213. [Google Scholar] [CrossRef]
- Johns, M.W. A new method for measuring daytime sleepiness: The Epworth Sleepiness Scale. Sleep 1991, 14, 50–55. [Google Scholar]
- Beck, A.T.; Steer, R.A.; Garbin, M.G. Psychometric properties of Beck Depression Inventory: Twenty-five years of evaluation. Clin. Psychol. 1988, 8, 77–100. [Google Scholar] [CrossRef]
- Zigmond, A.S.; Snaith, R.P. The hospital anxiety and depression scale. Acta Psychiatr. Scand. 1983, 67, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Deltour, J.J. Echelle de Vocabulaire Mill Hill de J. C. Raven: Adaptation Française et normes Comparées du Mill Hill et du Standard Progressive Matrices (PM38). Manuel et Annexes; Application des Techniques Modernes: Braine le Château, Belgique, 1933. [Google Scholar]
- Bagby, R.M.; Parker, J.D.A.; Taylor, G.J. The twenty-item Toronto Alexithymia Scale-I. Item selection and cross-validation of the factor structure. J. Psychosom. Res. 1994, 38, 23–32. [Google Scholar] [CrossRef]
- Horne, J.A.; Ostberg, O.A. Self-assessment questionnaire to determine morningness-eveningness in human circadian rhythms. Int. J. Chronobiol. 1976, 4, 97–110. [Google Scholar] [PubMed]
- Akerstedt, T.; Gillberg, M. Subjective and objective sleepiness in the active individual. Int. J. Neurosci. 1990, 52, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Dinges, D.I.; Powell, J.W. Microcomputer analysis of performance on a portable, simple visual RT task sustained operations. Behav. Res. Methods Instrum. Comput. 1985, 17, 652–655. [Google Scholar] [CrossRef]
- Bradley, M.M.; Lang, P.J. Measuring emotion: The self-assessment Manikin and the semantic differential. J. Behav. Ther. Exp. Psychiatry 1994, 25, 49–59. [Google Scholar] [CrossRef]
- Ekman, P.; Friesen, W.V.; Ellsworth, P. What emotion categories or dimensions can observers judge from facial behavior? In Emotion in the Human Face; Ekman, P., Ed.; Cambridge University Press: New York, NY, USA, 1972; pp. 39–55. [Google Scholar]
- Bertels, J.; Deliens, G.; Peigneux, P.; Destrebecqz, A. The Brussels Mood Inductive Audio Stories (MIAS) database. Behav. Res. Methods 2014, 46, 1098–1107. [Google Scholar] [CrossRef] [PubMed]
- Boucsein, W.; Fowles, D.C.; Grimnes, S.; Ben-Shakhar, G.; Roth, W.T.; Dawson, M.E.; Filion, D.L. Publication recommendations for electrodermal measurements. Psychophysiology 2012, 49, 1017–1034. [Google Scholar] [PubMed]
- Iber, C.; Ancoli-Israel, S.; Chesson, A.; Quand, S.F. The AASM Manual for the Scoring of Sleep and Associated Events: Rules, Terminology, and Technical Specification, 1st ed.; American Academy of Sleep Medicine: Westchester, IL, USA, 2007. [Google Scholar]
- Basner, M.; Dinges, D.F. Maximizing sensitivity of the psychomotor vigilance test (PVT) to sleep loss. Sleep 2011, 34, 581–591. [Google Scholar] [PubMed]
- Hornung, O.P.; Regen, F.; Danker-Hopfe, H.; Schredl, M.; Heuser, I. The relationship between REM sleep and memory consolidation in old age and effects of cholinergic medication. Biol. Psychiatry 2007, 61, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Datta, S. PGO Wave Generation: Mechanisms and functional significance. In Rapid Eye Movement Sleep; Mallick, B.N., Inoue, S., Eds.; Narosa Publishing House: New Delhi, India, 1999; pp. 91–106. [Google Scholar]
- Datta, S. Avoidance task training potentiates phasic pontine-wave density in the rat: A mechanism for sleep-dependent plasticity. J. Neurosci. 2000, 20, 8607–8613. [Google Scholar] [PubMed]
- Mavanji, V.; Datta, S. Activation of the phasic pontine-wave generator enhances improvement of learning performance: A mechanism for sleep-dependent plasticity. Eur. J. Neurosci. 2003, 17, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.; Mavanji, V.; Ulloor, J.; Patterson, E.H. Activation of phasic pontine- wave generator prevents rapid eye movement sleep deprivation-induced learning impairment in the rat: A mechanism for sleep-dependent plasticity. J. Neurosci. 2004, 24, 1416–1427. [Google Scholar] [CrossRef] [PubMed]
- Peigneux, P.; Laureys, S.; Fuchs, S.; Delbeuck, X.; Degueldre, C.; Aerts, J.; Delfiore, G.; Luxen, A.; Maquet, P. Generation of rapid eye movements during paradoxical sleep in humans. Neuroimage 2001, 14, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Lim, A.S.; Lozano, A.M.; Moro, E.; Hamani, C.; Hutchison, W.D.; Dostrovsky, J.O.; Lang, A.E.; Wennberg, R.A.; Murray, B.J. Characterization of REM-sleep associated ponto-geniculo-occipital waves in the human pons. Sleep 2007, 30, 823–827. [Google Scholar] [PubMed]
- Ioannides, A.A.; Corsi-Cabrera, M.; Fenwick, P.B.; del Rio Portilla, Y.; Laskaris, N.A.; Khurshudyan, A.; Theofilou, D.; Shibata, T.; Uchida, S.; Nakabayashi, T.; et al. MEG tomography of human cortex and brainstem activity in waking and REM sleep saccades. Cereb. Cortex 2004, 14, 56–72. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Salado, I.; Cruz-Aguilar, M.A. The origin and functions of dreams based on PGO activity. Salud Ment. 2014, 37, 49–58. [Google Scholar]
- Van der Helm, E.; Walker, M.P. Overnight therapy? The role of sleep in emotional brain processing. Psychol. Bull. 2009, 135, 731–748. [Google Scholar] [CrossRef] [PubMed]
- Talbot, L.S.; Hairston, I.S.; Eidelman, P.; Gruber, J.; Harvey, A.G. The effect of mood on sleep onset latency and REM sleep in interepisode bipolar disorder. J. Abnorm. Psychol. 2009, 118, 448–458. [Google Scholar] [CrossRef] [PubMed]
- Gujar, N.; McDonald, S.A.; Nishida, M.; Walker, M.P. A role for REM sleep in recalibrating the sensitivity of the human brain to specific emotions. Cereb. Cortex 2011, 21, 1115–1123. [Google Scholar] [CrossRef] [PubMed]
- Deliens, G.; Peigneux, P. One night of sleep is insufficient to achieve sleep-to-forget emotional decontextualisation processes. Cogn. Emot. 2014, 28, 698–706. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gilson, M.; Deliens, G.; Leproult, R.; Bodart, A.; Nonclercq, A.; Ercek, R.; Peigneux, P. REM-Enriched Naps Are Associated with Memory Consolidation for Sad Stories and Enhance Mood-Related Reactivity. Brain Sci. 2016, 6, 1. https://doi.org/10.3390/brainsci6010001
Gilson M, Deliens G, Leproult R, Bodart A, Nonclercq A, Ercek R, Peigneux P. REM-Enriched Naps Are Associated with Memory Consolidation for Sad Stories and Enhance Mood-Related Reactivity. Brain Sciences. 2016; 6(1):1. https://doi.org/10.3390/brainsci6010001
Chicago/Turabian StyleGilson, Médhi, Gaétane Deliens, Rachel Leproult, Alice Bodart, Antoine Nonclercq, Rudy Ercek, and Philippe Peigneux. 2016. "REM-Enriched Naps Are Associated with Memory Consolidation for Sad Stories and Enhance Mood-Related Reactivity" Brain Sciences 6, no. 1: 1. https://doi.org/10.3390/brainsci6010001