
Hemodynamic and Light-Scattering Changes of Rat Spinal Cord and Primary Somatosensory Cortex in Response to Innocuous and Noxious Stimuli

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Preparation
2.2. Data Collection
2.3. Mechanical, Electrical and Chemical Stimuli
2.4 Statistical Analysis
3. Results
3.1. Basal Levels of Hemodynamic Parameters and Light-Scattering Coefficients

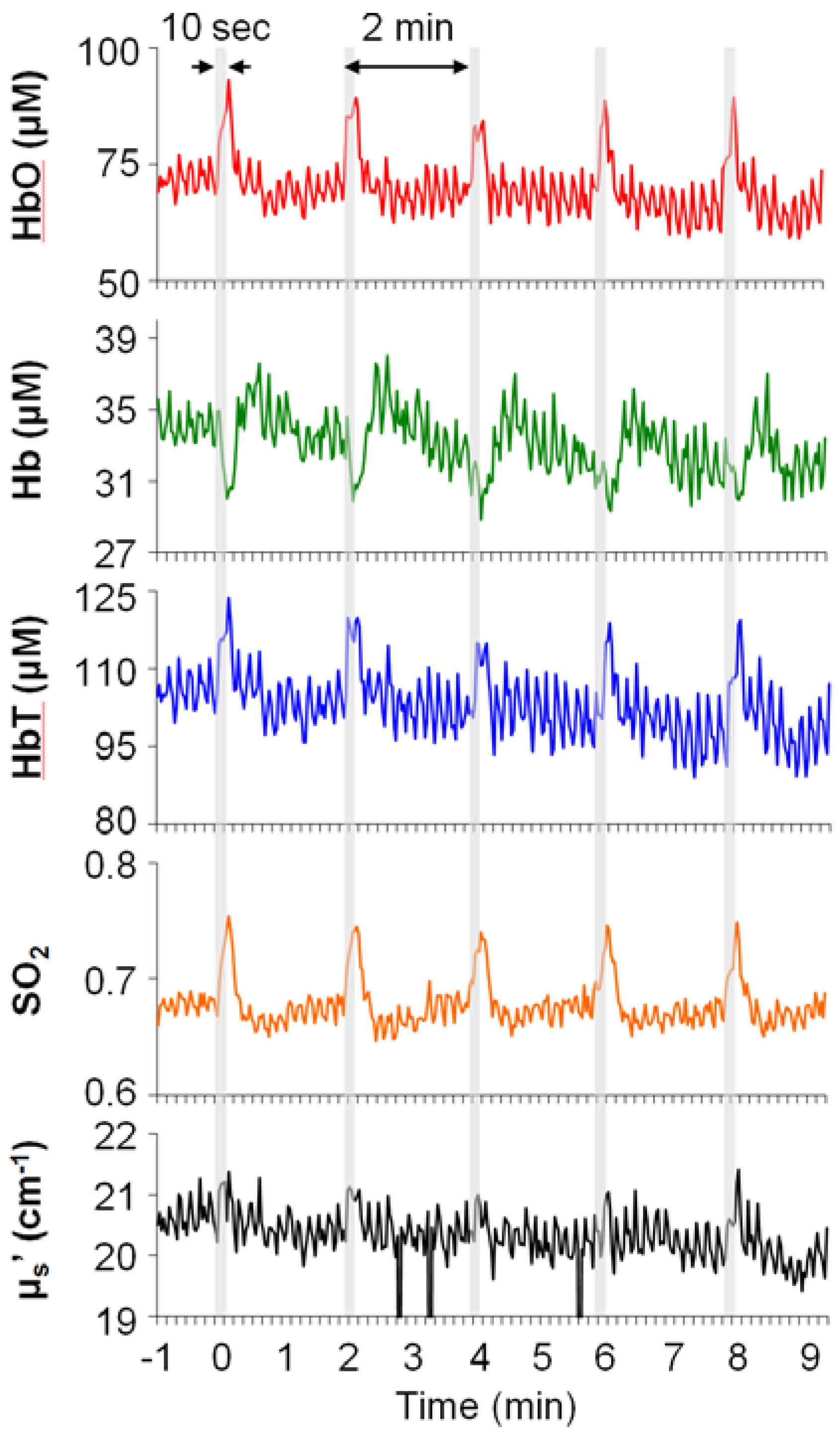{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spinal Cord | SI | SP vs SI | |||
|---|---|---|---|---|---|
| Ipsi | Cont | Ipsi | Cont | ||
| HbO (µM) | 31.1 ± 18.7 | 14.5 ± 4.5 | 112.1 ± 21.7 | 71.8 ± 15.6 | p = 0.002 (*) |
| Hb (µM) | 50.5 ± 28.7 | 18.8 ± 7.9 | 46.7 ± 16.7 | 68.1 ± 18.9 | p = 0.079 |
| SO2 | 0.40 ± 0.07 | 0.52 ± 0.07 | 0.70 ± 0.08 | 0.51 ± 0.07 | p = 0.044 (*) |
| HbT (µM) | 81.3 ± 47.0 | 33.4 ± 11.0 | 160.1 ± 24.7 | 140.0 ± 30.7 | p = 0.014 (*) |
| µs′ (cm-1) | 29.5 ± 3.6 | 35.1 ± 2.8 | 13.5 ± 2.9 | 11.5 ± 0.6 | p < 0.001 (*) |
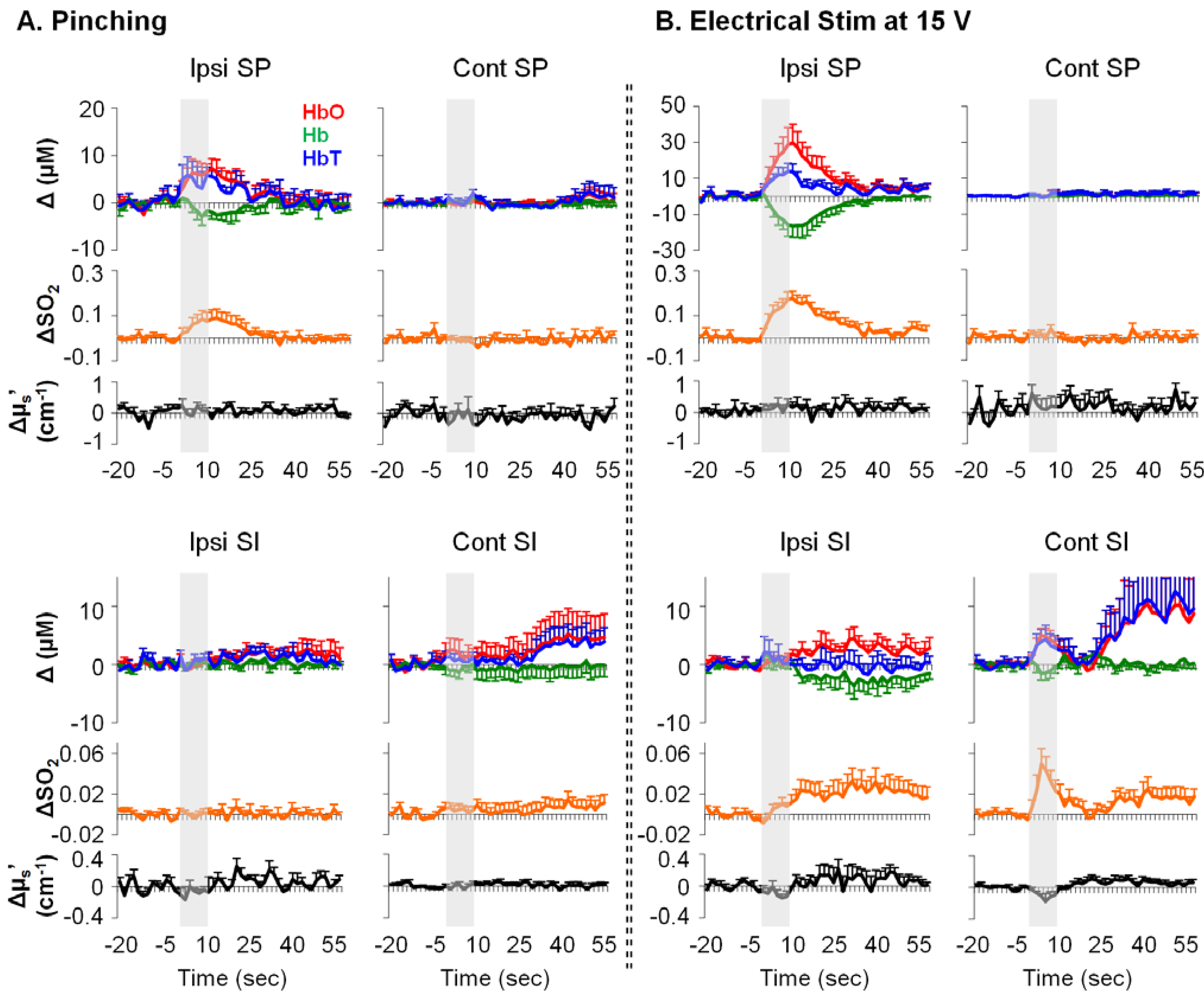
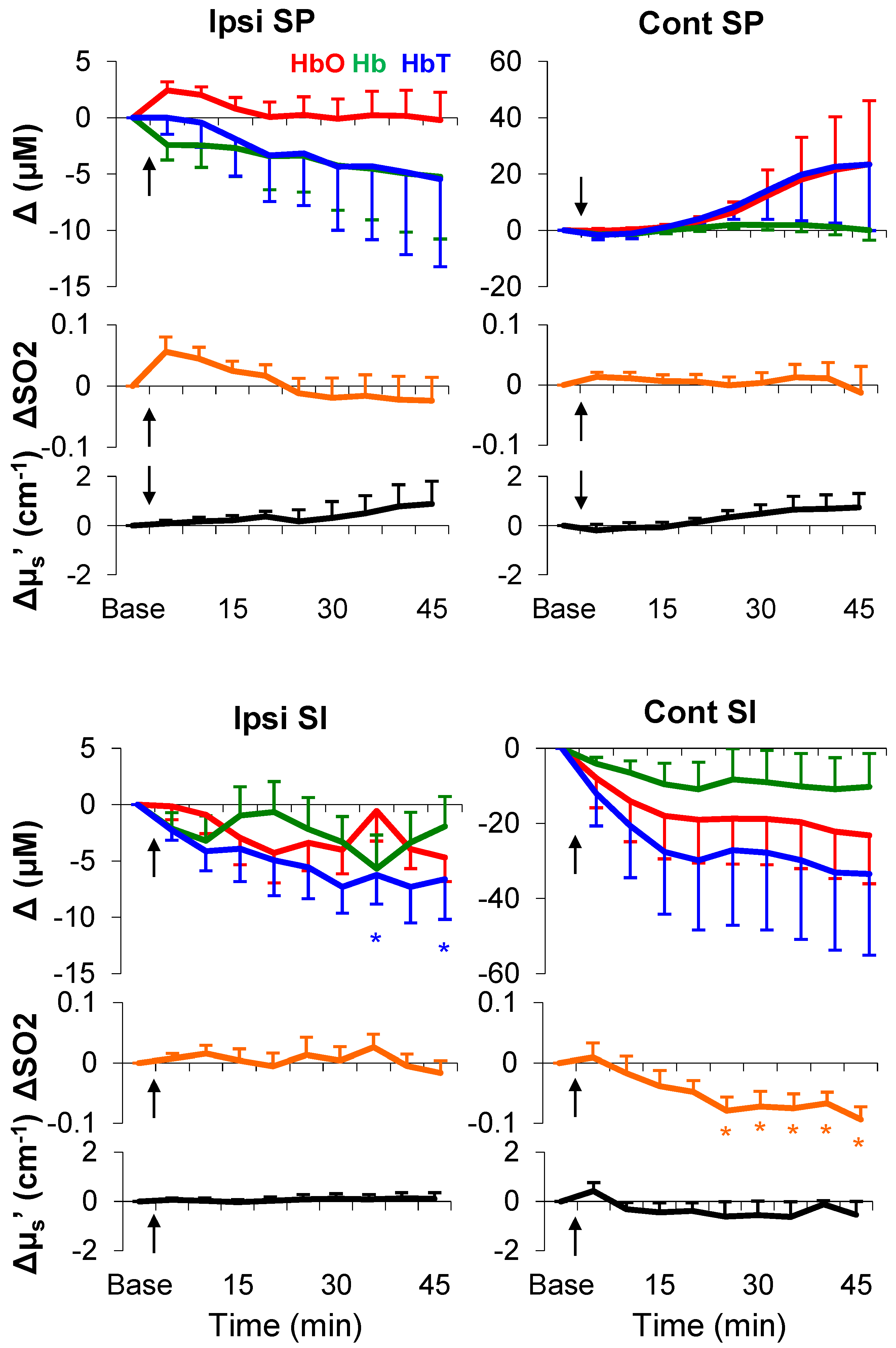
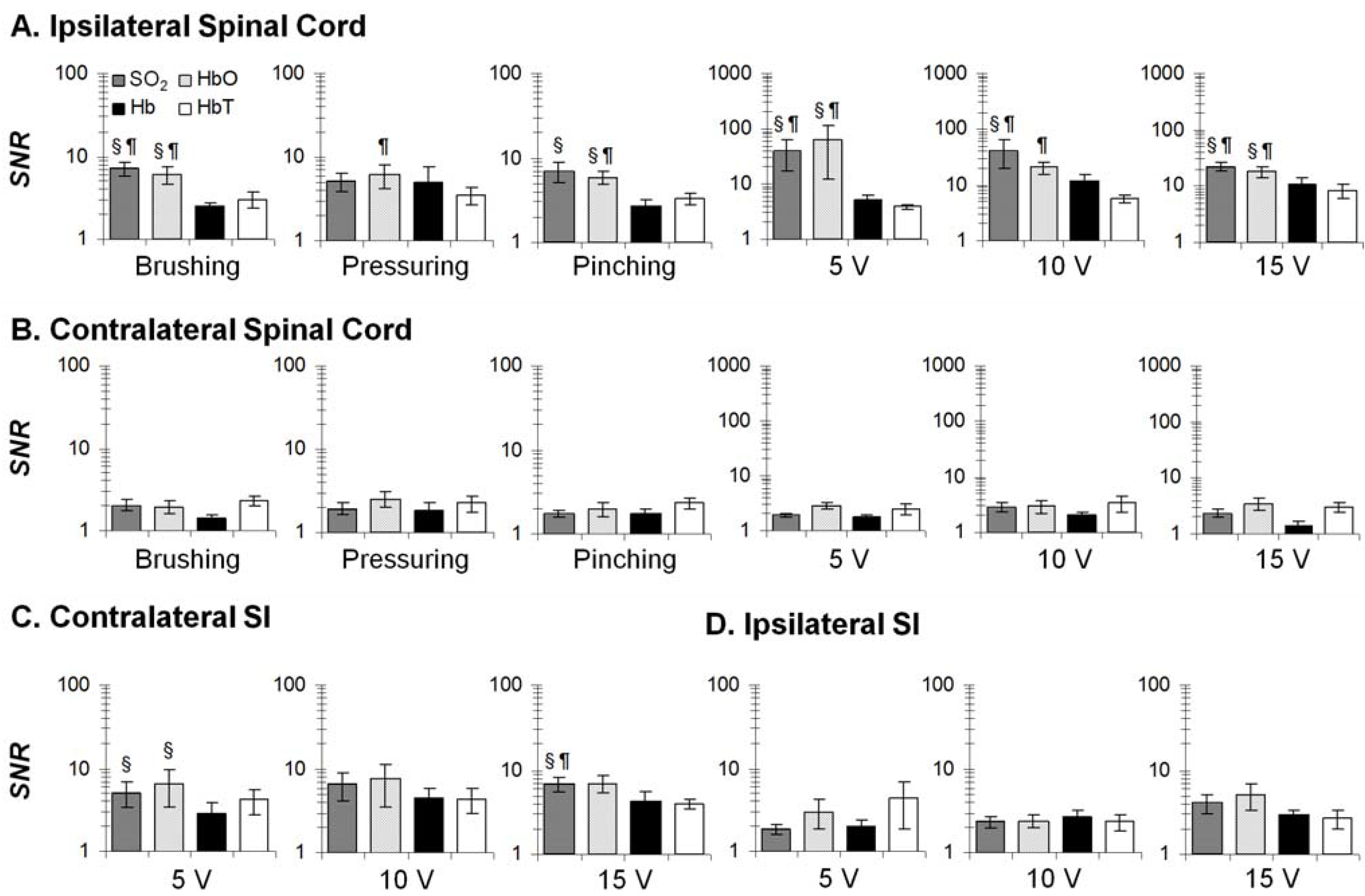
3.2. Mechanical- and Electrical-Stimulation-Induced Hemodynamic and Light-Scattering Changes
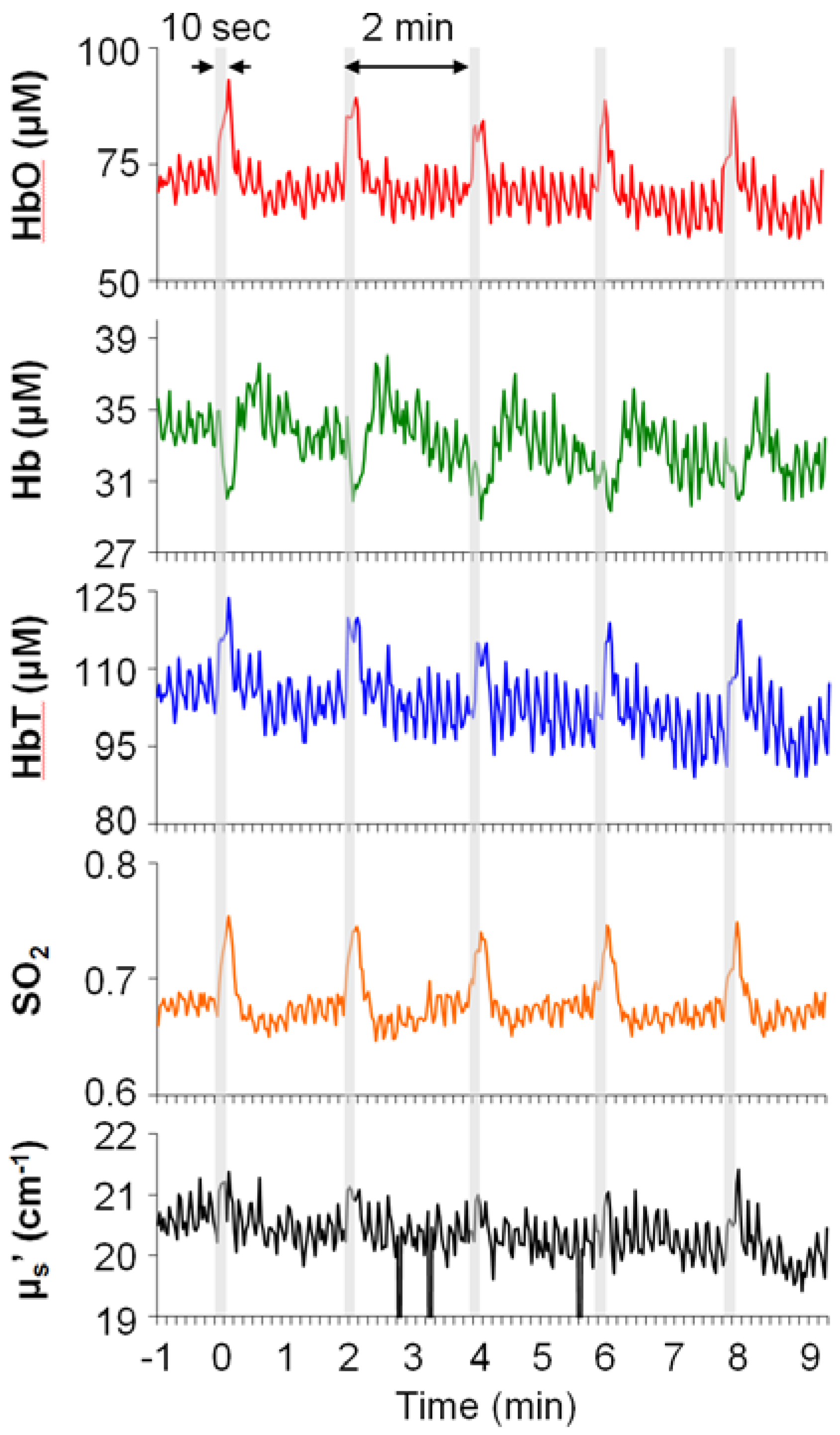

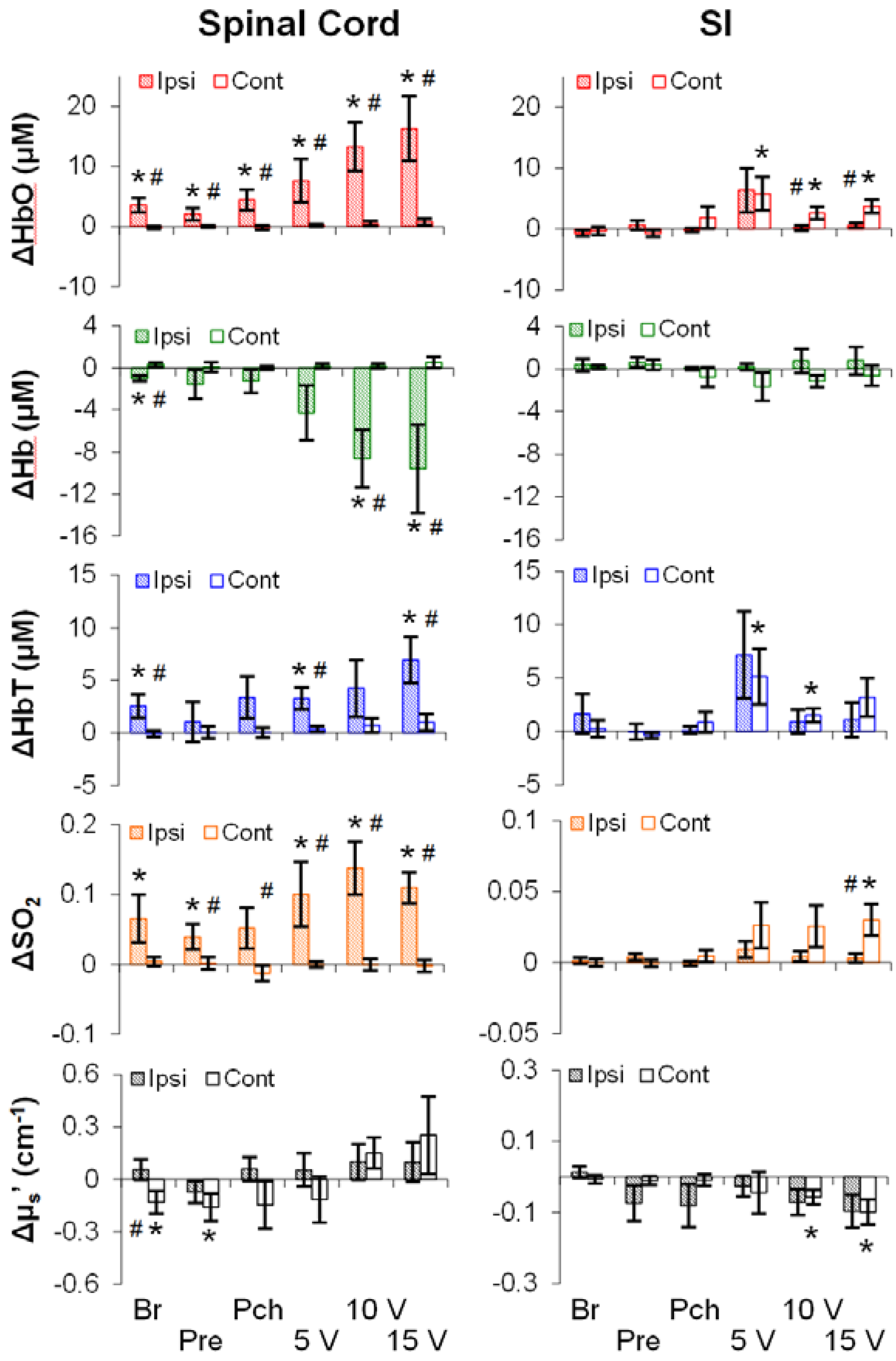
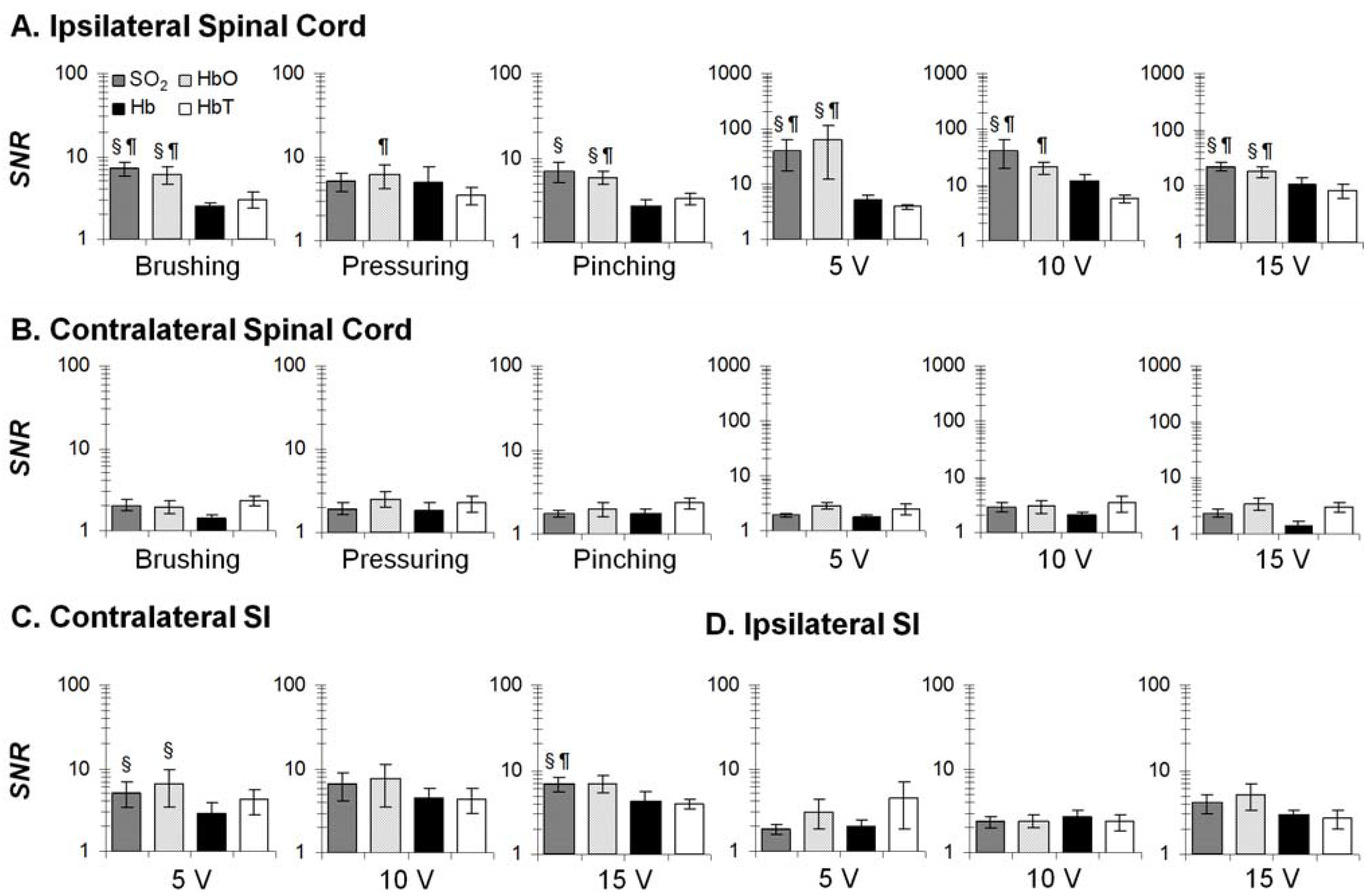
3.3. Temporal Characteristics of Hemodynamics in the Spinal Cord and SI
| HbO | SO2 | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Onset | Peak | Duration | n | Onset | Peak | Duration | n | ||
| Ipsi SP | Br | 4.8 ± 1.1 (‡) | 9.8 ± 0.9 | 15.6 ± 3.1 | 8 | 7.3 ± 1.6 | 13.8 ± 1.4 | 19.5 ± 3.6 | 10 |
| Pre | 4.6 ± 1.4 | 13.7 ± 2.6 | 17.4 ± 4.0 | 9 | 5.0 ± 1.0 | 13.7 ± 2.6 | 17.2 ± 3.7 | 9 | |
| Pch | 4.0 ± 1.0 (‡) | 10.0 ± 1.1 | 15.6 ± 3.3 | 8 | 3.5 ± 0.5 | 13.1 ± 1.8 | 19.8 ± 3.2 | 8 | |
| 5V | 3.2 ± 0.7 (‡) | 10.8 ± 0.9 (†) | 23.5 ± 4.1 (†) | 10 | 4.2 ± 0.7 (‡†) | 12.3 ± 0.8 (†) | 30.8 ± 4.5 (†) | 10 | |
| 10V | 4.0 ± 1.0 (‡) | 11.5 ± 0.9 (†) | 29.2 ± 3.8 (†) | 10 | 2.8 ± 0.5 (‡) | 11.0 ± 0.4 (†) | 35.3 ± 3.8 (†) | 10 | |
| 15V | 3.8 ± 1.5 | 12.3 ± 1.1 (†) | 33.7 ± 5.2 (†) | 10 | 4.2 ± 1.5 (‡†) | 13.3 ± 1.0 (†) | 30.7 ± 6.2 (†) | 10 | |
| Cont SI | Br | 2.2 ± 1.5 | 5.0 ± 1.9 | 15.0 ± 13.3 | 3 | 3.3 ± 1.7 | 3.3 ± 1.7 | 2.5 ± 0.8 | 2 |
| Pre | 5.0 ± 3.3 | 5.0 ± 3.3 | 1.7 ± 0 | 2 | 8.3 | 8.3 | 1.7 | 1 | |
| Pch | 4.2 ± 4.2 | 6.7 ± 1.7 | 4.2 ± 2.5 | 2 | 2.8 ± 2.8 | 4.4 ± 2.4 | 3.3 ± 1.7 | 3 | |
| 5V | 1.9 ± 0.8 | 4.1 ± 1.1 | ± 2.1 | 7 | 1.1 ± 0.6 (‡) | 4.4 ± 1.2 | 9.2 ± 2.3 | 6 | |
| 10V | 3.3 ± 0.7 (‡) | 4.8 ± 0.7 | 4.5 ± 1.6 | 7 | 2.5 ± 0.5 | 4.0 ± 0.6 | 5.8 ± 1.7 | 8 | |
| 15V | 2.9 ± 0.9 | 5.4 ± 0.8 | 13.8 ± 4.6 | 8 | 1.9 ± 0.5 (‡) | 3.7 ± 0.2 | 10.4 ± 2.1 | 9 | |
| Ipsi SI | Br | 23.8 ± 6.2 | 38.3 ± 9.1 | 19.0 ± 8.7 | 8 | 15.3 ± 7.3 | 33.1 ± 12.3 | 22.8 ± 10.8 | 6 |
| Pre | 9.8 ± 3.1 | 21.9 ± 9.8 | 19.5 ± 9.1 | 7 | 13.7 ± 9.7 | 33.0 ± 13.4 | 24.0 ± 15.9 | 5 | |
| Pch | 27.4 ± 9.5 | 47.6 ± 8.4 | 18.8 ± 5.6 | 7 | 22.8 ± 8.5 | 52.5 ± 12.7 | 32.8 ± 11.6 | 6 | |
| 5V | 20.2 ± 8.8 | 30.5 ± 6.7 | 21.4 ± 12.1 | 7 | 23.3 ± 6.5 | 35.4 ± 5.8 | 16.3 ± 9.5 | 8 | |
| 10V | 15.0 ± 4.4 | 33.0 ± 5.4 | 26.5 ± 5.9 | 9 | 15.6 ± 5.2 | 26.7 ± 6.4 | 13.2 ± 4.0 | 9 | |
| 15V | 7.6 ± 2.6 | 23.5 ± 7.5 | 21.7 ± 8.8 | 9 | 18.5 ± 6.1 | 29.8 ± 6.2 | 31.3 ± 9.8 | 8 | |
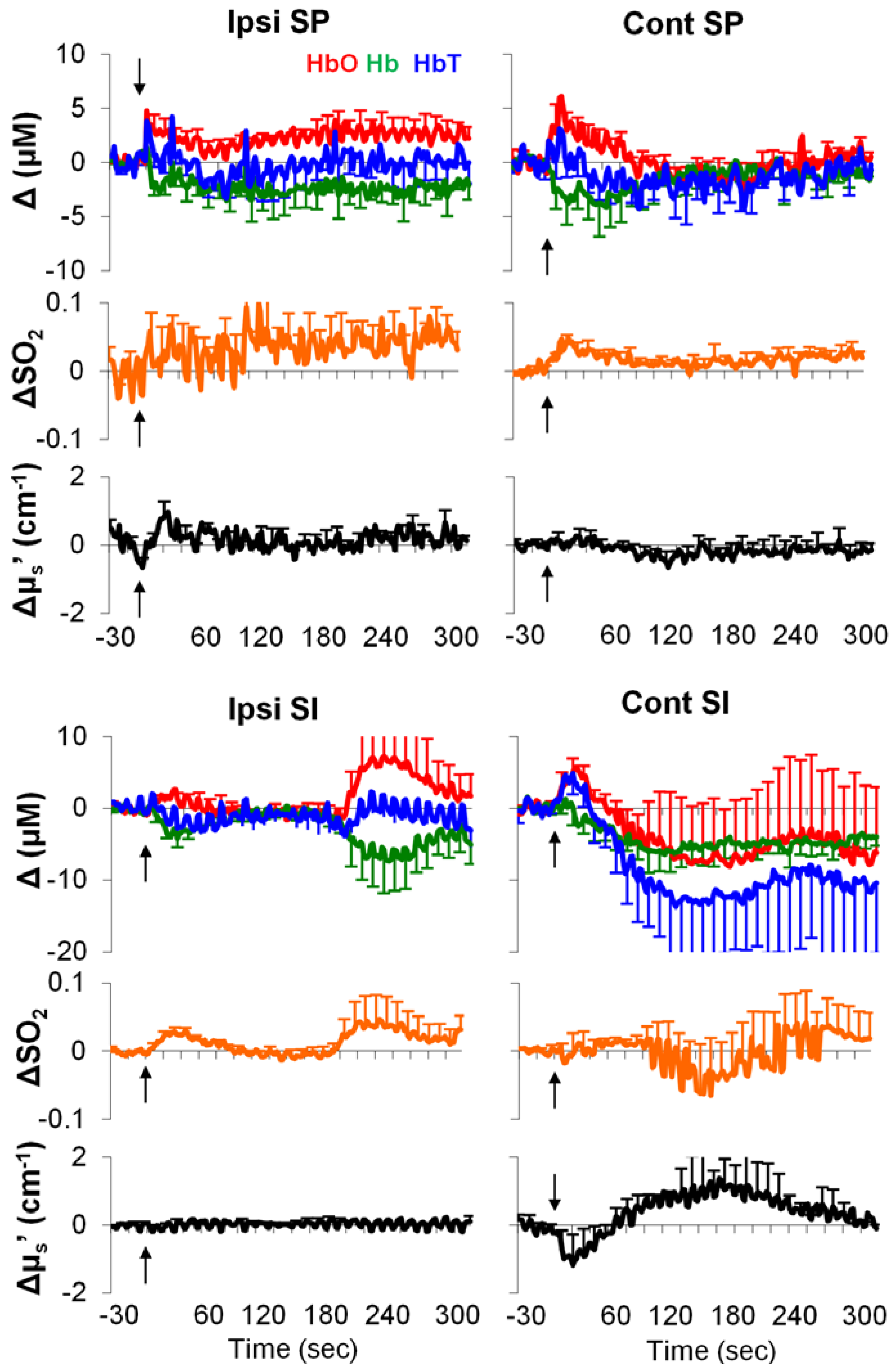
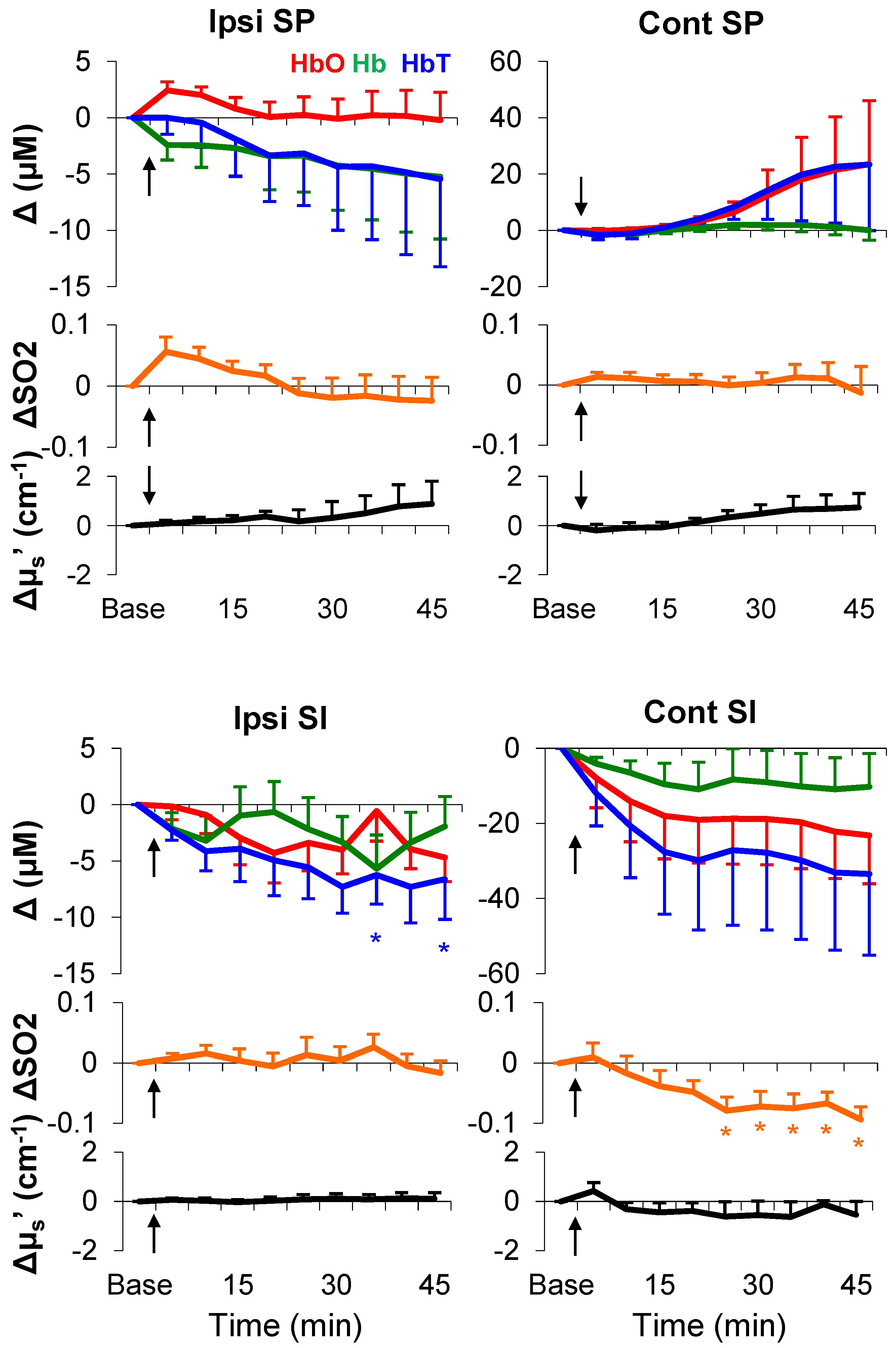
3.4. Formalin-Induced Hemodynamic and Light-Scattering Changes
| Ipsi SP | Cont SP | Ipsi SI | Cont SI | |
|---|---|---|---|---|
| ΔHbO (μM) | 2.2 ± 0.8 (*) | 2.5 ± 1.8 | 0.8 ± 0.3 (*) | 2.1 ± 1.3 |
| ΔHb (μM) | −1.6 ± 0.7 (*) | −2.8 ± 1.8 | −2.3 ± 0.6 (*) | −1.5 ± 1.3 |
| ΔSO2 | 0.02 ± 0.03 | 0.03 ± 0.007 (*) | 0.02 ± 0.002 (*) | 0.003 ± 0.1 |
| ΔHbT (μM) | 0.3 ± 0.5 | −0.1 ± 0.8 | −1.3 ± 0.6 (*) | 0.5 ± 2.2 |
| Δµs′ (cm−1) | 0.3 ± 0.2 | 0.02 ± 0.2 | −0.002 ± 0.02 | −0.5 ± 0.5 |

| Ipsi SP | Cont SP | Ipsi SI | Cont SI | |
|---|---|---|---|---|
| SO2 | 0.024 (a,*) | 0.98 | 0.61 | 0.001 (a,*) |
| HbO | 0.48 | 0.38 | 0.08 | 0.012 (*) |
| Hb | 0.71 | 0.33 | 0.40 | 0.45 |
| HbT | 0.79 | 0.33 | 0.046 (a, *) | 0.046 (*) |
| μs′ | 0.80 | 0.35 | 0.98 | 0.35 |
4. Discussions
4.1. Electrical, but Not Mechanical, Stimuli Produced an Intensity-Dependent HbO Increase
4.2. Regional Characteristics of Hemodynamic Responses
4.3. Hemodynamic Signatures of Spinal Cord and SI in Response to a Long-Lasting Noxious Stimulus
4.4. Regional Characteristics of Light Scattering
4.5. Anesthesia
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Raichle, M.E. The restless brain: How intrinsic activity organizes brain function. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2015, 370. [Google Scholar] [CrossRef]
- Borsook, D.; Becerra, L.; Hargreaves, R. Biomarkers for chronic pain and analgesia. Part 1: The need, reality, challenges, and solutions. Discov. Med. 2011, 11, 197–207. [Google Scholar] [PubMed]
- Tracey, I. Imaging pain. Br. J. Anaesth. 2008, 101, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Woo, C.W.; Wager, T.D. Neuroimaging-based biomarker discovery and validation. Pain 2015, 156, 1379–1381. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.E.; Chatterjee, N.; Younger, J.; Mackey, S. Towards a physiology-based measure of pain: Patterns of human brain activity distinguish painful from non-painful thermal stimulation. PLoS ONE 2011, 6, e24124. [Google Scholar] [CrossRef] [PubMed]
- Wager, T.D.; Atlas, L.Y.; Lindquist, M.A.; Roy, M.; Woo, C.W.; Kross, E. An fmri-based neurologic signature of physical pain. N. Engl. J. Med. 2013, 368, 1388–1397. [Google Scholar] [CrossRef] [PubMed]
- Becerra, L.; Harris, W.; Grant, M.; George, E.; Boas, D.; Borsook, D. Diffuse optical tomography activation in the somatosensory cortex: Specific activation by painful vs. Non-painful thermal stimuli. PLoS ONE 2009, 4, e8016. [Google Scholar] [CrossRef] [PubMed]
- Yennu, A.; Tian, F.; Liu, H.; Rawat, R.; Manry, M.T.; Gatchel, R. A preliminary investigation of human frontal cortex under noxious thermal stimulation over the temporomandibular joint using functional near infrared spectroscopy. J. Appl. Biobehav. Res. 2013, 18, 134–155. [Google Scholar] [CrossRef]
- Yucel, M.A.; Aasted, C.M.; Petkov, M.P.; Borsook, D.; Boas, D.A.; Becerra, L. Specificity of hemodynamic brain responses to painful stimuli: A functional near-infrared spectroscopy study. Sci. Rep. 2015, 5, 9469. [Google Scholar] [CrossRef] [PubMed]
- He, J.W.; Kashyap, D.; Trevino, L.A.; Liu, H.; Peng, Y.B. Simultaneous absolute measures of glabrous skin hemodynamic and light-scattering change in response to formalin injection in rats. Neurosci. Lett. 2011, 492, 59–63. [Google Scholar] [CrossRef]
- Sharma, V.; He, J.W.; Narvenkar, S.; Peng, Y.B.; Liu, H. Quantification of light reflectance spectroscopy and its application: Determination of hemodynamics on the rat spinal cord and brain induced by electrical stimulation. NeuroImage 2011, 56, 1316–1328. [Google Scholar] [CrossRef] [PubMed]
- Hill, D.K.; Keynes, R.D. Opacity changes in stimulated nerve. J. Physiol. 1949, 108, 278–281. [Google Scholar] [CrossRef] [PubMed]
- Cohen, L.B.; Keynes, R.D.; Landowne, D. Changes in axon light scattering that accompany the action potential: Current-dependent components. J. Physiol. 1972, 224, 727–752. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Swanson, E.A.; Lin, C.P.; Schuman, J.S.; Stinson, W.G.; Chang, W.; Hee, M.R.; Flotte, T.; Gregory, K.; Puliafito, C.A.; et al. Optical coherence tomography. Science 1991, 254, 1178–1181. [Google Scholar] [CrossRef] [PubMed]
- Maheswari, R.U.; Takaoka, H.; Kadono, H.; Homma, R.; Tanifuji, M. Novel functional imaging technique from brain surface with optical coherence tomography enabling visualization of depth resolved functional structure in vivo. J. Neurosci. Methods 2003, 124, 83–92. [Google Scholar] [CrossRef]
- Srinivasan, V.J.; Wojtkowski, M.; Fujimoto, J.G.; Duker, J.S. In vivo measurement of retinal physiology with high-speed ultrahigh-resolution optical coherence tomography. Opt. Lett. 2006, 31, 2308–2310. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, A.D.; Chen, Y.; Fujimoto, J.G.; Ruvinskaya, L.; Devor, A.; Boas, D.A. Depth-resolved imaging of functional activation in the rat cerebral cortex using optical coherence tomography. Opt. Lett. 2006, 31, 3459–3461. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rajagopalan, U.M.; Tanifuji, M. Functional optical coherence tomography reveals localized layer-specific activations in cat primary visual cortex in vivo. Opt. Lett. 2007, 32, 2614–2616. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Radhakrishnan, H.; Senapati, A.K.; Hagains, C.E.; Peswani, D.; Mathker, A.; Peng, Y.B. Near infrared and visible spectroscopic measurements to detect changes in light scattering and hemoglobin oxygen saturation from rat spinal cord during peripheral stimulation. NeuroImage 2008, 40, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.B.; Lin, Q.; Willis, W.D. Effects of gaba and glycine receptor antagonists on the activity and pag-induced inhibition of rat dorsal horn neurons. Brain Res. 1996, 736, 189–201. [Google Scholar] [CrossRef]
- Zimmermann, M. Ethical guidelines for investigations of experimental pain in conscious animals. Pain 1983, 16, 109–110. [Google Scholar] [CrossRef]
- Peng, Y.B.; Lin, Q.; Willis, W.D. The role of 5-ht3 receptors in periaqueductal gray-induced inhibition of nociceptive dorsal horn neurons in rats. J. Pharmacol. Exp. Ther. 1996, 276, 116–124. [Google Scholar] [PubMed]
- Ativanichayaphong, T.; He, J.W.; Hagains, C.E.; Peng, Y.B.; Chiao, J.C. A combined wireless neural stimulating and recording system for study of pain processing. J. Neurosci. Methods 2008, 170, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Plesnila, N.; Putz, C.; Rinecker, M.; Wiezorrek, J.; Schleinkofer, L.; Goetz, A.E.; Kuebler, W.M. Measurement of absolute values of hemoglobin oxygenation in the brain of small rodents by near infrared reflection spectrophotometry. J. Neurosci. Methods 2002, 114, 107–117. [Google Scholar] [CrossRef]
- Nihashi, T.; Naganawa, S.; Sato, C.; Kawai, H.; Nakamura, T.; Fukatsu, H.; Ishigaki, T.; Aoki, I. Contralateral and ipsilateral responses in primary somatosensory cortex following electrical median nerve stimulation—An fmri study. Clin. Neurophysiol. 2005, 116, 842–848. [Google Scholar] [CrossRef] [PubMed]
- Hlushchuk, Y.; Hari, R. Transient suppression of ipsilateral primary somatosensory cortex during tactile finger stimulation. J. Neurosci. 2006, 26, 5819–5824. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Disbrow, E.A.; Zumer, J.M.; McGonigle, D.J.; Nagarajan, S.S. Spatiotemporal integration of tactile information in human somatosensory cortex. BMC Neurosci. 2007, 8, 21. [Google Scholar] [CrossRef] [PubMed]
- Killackey, H.P.; Gould, H.J., III; Cusick, C.G.; Pons, T.P.; Kaas, J.H. The relation of corpus callosum connections to architectonic fields and body surface maps in sensorimotor cortex of new and old world monkeys. J. Comp. Neurol. 1983, 219, 384–419. [Google Scholar] [CrossRef] [PubMed]
- Gardner, E.P.; Martin, J.H. Coding of sensory information. In Principles of Neural Science; Kandel, E.R., Schwartz, J.H., Jessell, T.M., Eds.; McGraw-Hill: New York, NY, USA, 2000; pp. 411–429. [Google Scholar]
- Georgopoulos, A.P. Functional properties of primary afferent units probably related to pain mechanisms in primate glabrous skin. J. Neurophysiol. 1976, 39, 71–83. [Google Scholar] [PubMed]
- Peng, Y.B.; Ringkamp, M.; Campbell, J.N.; Meyer, R.A. Electrophysiological assessment of the cutaneous arborization of adelta-fiber nociceptors. J. Neurophysiol. 1999, 82, 1164–1177. [Google Scholar] [PubMed]
- Chung, J.M.; Surmeier, D.J.; Lee, K.H.; Sorkin, L.S.; Honda, C.N.; Tsong, Y.; Willis, W.D. Classification of primate spinothalamic and somatosensory thalamic neurons based on cluster analysis. J. Neurophysiol. 1986, 56, 308–327. [Google Scholar] [PubMed]
- Takano, T.; Tian, G.F.; Peng, W.; Lou, N.; Libionka, W.; Han, X.; Nedergaard, M. Astrocyte-mediated control of cerebral blood flow. Nat. Neurosci. 2006, 9, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Hendrich, K.S.; Masamoto, K.; Kim, S.G. Arterial versus total blood volume changes during neural activity-induced cerebral blood flow change: Implication for bold fmri. J. Cereb. Blood Flow Metab. 2007, 27, 1235–1247. [Google Scholar] [CrossRef] [PubMed]
- Dubuisson, D.; Dennis, S.G. The formalin test: A quantitative study of the analgesic effects of morphine, meperidine, and brain stem stimulation in rats and cats. Pain 1977, 4, 161–174. [Google Scholar] [CrossRef]
- Dickenson, A.H.; Sullivan, A.F. Peripheral origins and central modulation of subcutaneous formalin-induced activity of rat dorsal horn neurones. Neurosci. Lett. 1987, 83, 207–211. [Google Scholar] [CrossRef]
- He, J.W.; Tian, F.; Liu, H.; Peng, Y.B. Cerebrovascular responses of the rat brain to noxious stimuli as examined by functional near-infrared whole brain imaging. J. Neurophysiol. 2012, 107, 2853–2865. [Google Scholar] [CrossRef] [PubMed]
- Iadecola, C. Neurovascular regulation in the normal brain and in Alzheimer’s disease. Nat. Rev. Neurosci. 2004, 5, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Fox, P.T.; Raichle, M.E. Focal physiological uncoupling of cerebral blood flow and oxidative metabolism during somatosensory stimulation in human subjects. Proc. Natl. Acad. Sci. USA 1986, 83, 1140–1144. [Google Scholar] [CrossRef] [PubMed]
- Vretzakis, G.; Georgopoulou, S.; Stamoulis, K.; Stamatiou, G.; Tsakiridis, K.; Zarogoulidis, P.; Katsikogianis, N.; Kougioumtzi, I.; Machairiotis, N.; Tsiouda, T.; et al. Cerebral oximetry in cardiac anesthesia. J. Thorac. Dis. 2014, 6 (Suppl. S1), S60–S69. [Google Scholar] [PubMed]
- Scheeren, T.W.; Schober, P.; Schwarte, L.A. Monitoring tissue oxygenation by near infrared spectroscopy (nirs): Background and current applications. J. Clin. Monit. Comput. 2012, 26, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Yildiz, S.; Cimsit, M.; Ilgezdi, S.; Uzun, G.; Gumus, T.; Qyrdedi, T.; Dalci, D. Hyperbaric oxygen therapy used to treat radiation injury: Two case reports. Ostomy/Wound Manag. 2006, 52, 14–16, 18, 20. [Google Scholar]
- Thompson, C.D.; Uhelski, M.L.; Wilson, J.R.; Fuchs, P.N. Hyperbaric oxygen treatment decreases pain in two nerve injury models. Neurosci. Res. 2010, 66, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Gebhart, S.C.; Lin, W.C.; Mahadevan-Jansen, A. In vitro determination of normal and neoplastic human brain tissue optical properties using inverse adding-doubling. Phys. Med. Biol. 2006, 51, 2011–2027. [Google Scholar] [CrossRef]
- Tremoleda, J.L.; Kerton, A.; Gsell, W. Anaesthesia and physiological monitoring during in vivo imaging of laboratory rodents: Considerations on experimental outcomes and animal welfare. EJNMMI Res. 2012, 2, 44. [Google Scholar] [CrossRef] [PubMed]
- Masamoto, K.; Kanno, I. Anesthesia and the quantitative evaluation of neurovascular coupling. J. Cereb. Blood Flow Metab. 2012, 32, 1233–1247. [Google Scholar] [CrossRef] [PubMed]
- Franceschini, M.A.; Radhakrishnan, H.; Thakur, K.; Wu, W.; Ruvinskaya, S.; Carp, S.; Boas, D.A. The effect of different anesthetics on neurovascular coupling. NeuroImage 2010, 51, 1367–1377. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.N.; Purdon, P.L.; van Dort, C.J. General anesthesia and altered states of arousal: A systems neuroscience analysis. Annu. Rev. Neurosci. 2011, 34, 601–628. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, J.-W.; Liu, H.; Peng, Y.B. Hemodynamic and Light-Scattering Changes of Rat Spinal Cord and Primary Somatosensory Cortex in Response to Innocuous and Noxious Stimuli. Brain Sci. 2015, 5, 400-418. https://doi.org/10.3390/brainsci5040400
He J-W, Liu H, Peng YB. Hemodynamic and Light-Scattering Changes of Rat Spinal Cord and Primary Somatosensory Cortex in Response to Innocuous and Noxious Stimuli. Brain Sciences. 2015; 5(4):400-418. https://doi.org/10.3390/brainsci5040400
Chicago/Turabian StyleHe, Ji-Wei, Hanli Liu, and Yuan Bo Peng. 2015. "Hemodynamic and Light-Scattering Changes of Rat Spinal Cord and Primary Somatosensory Cortex in Response to Innocuous and Noxious Stimuli" Brain Sciences 5, no. 4: 400-418. https://doi.org/10.3390/brainsci5040400
APA StyleHe, J.-W., Liu, H., & Peng, Y. B. (2015). Hemodynamic and Light-Scattering Changes of Rat Spinal Cord and Primary Somatosensory Cortex in Response to Innocuous and Noxious Stimuli. Brain Sciences, 5(4), 400-418. https://doi.org/10.3390/brainsci5040400