Insights into Intraspecies Variation in Primate Prosocial Behavior: Capuchins (Cebus apella) Fail to Show Prosociality on a Touchscreen Task

Abstract
:1. Introduction
2. Method
2.1. Subjects
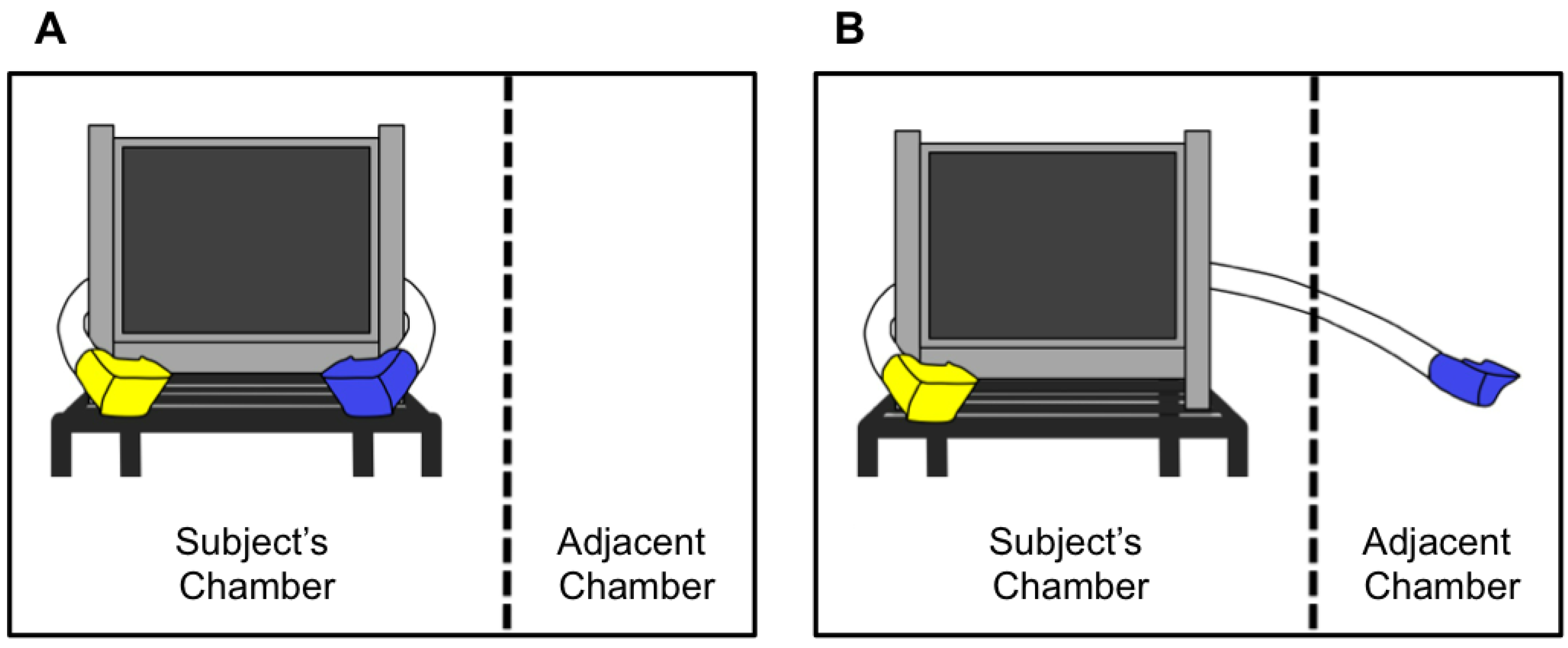
2.2. Testing Apparatus
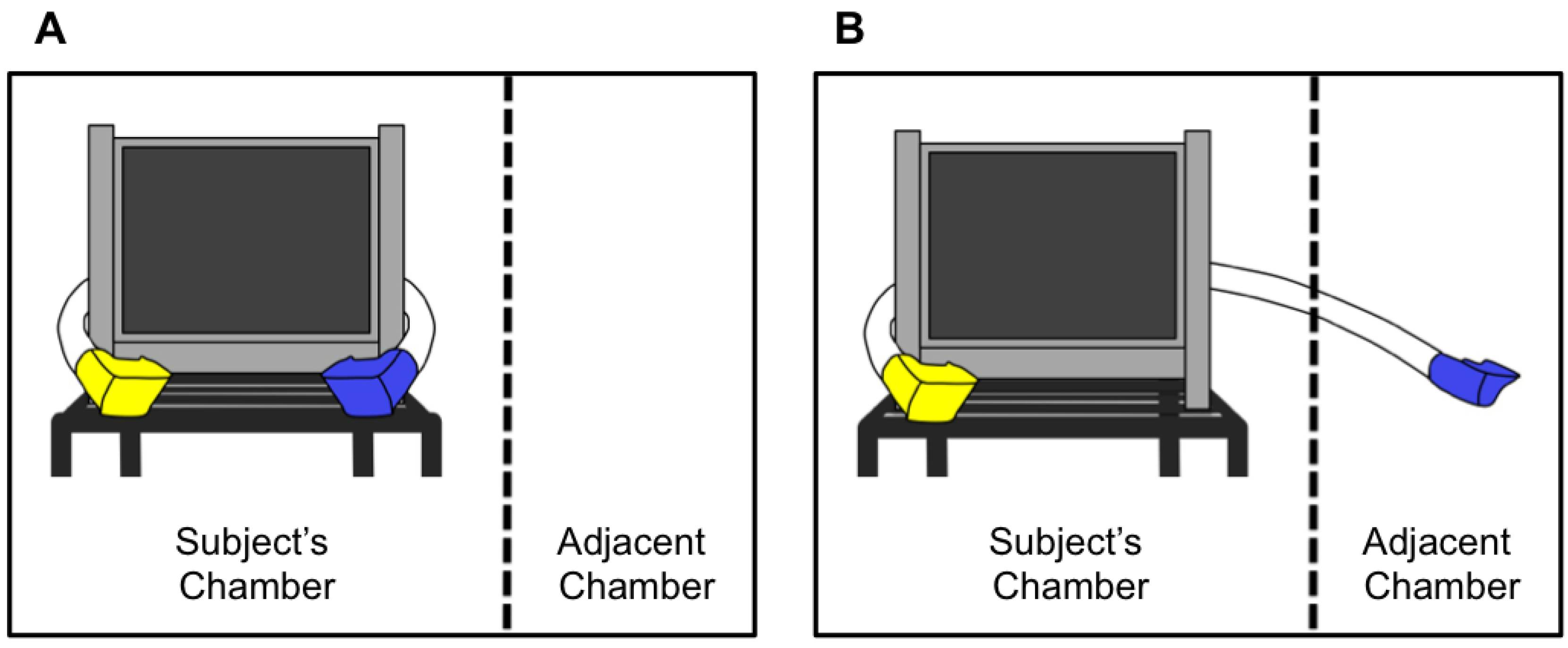
2.3. Stimuli

2.4. Procedure
2.4.1. Stimulus Preference Control Session
2.4.2. Training on Reward Contingencies
2.4.3. Test Sessions
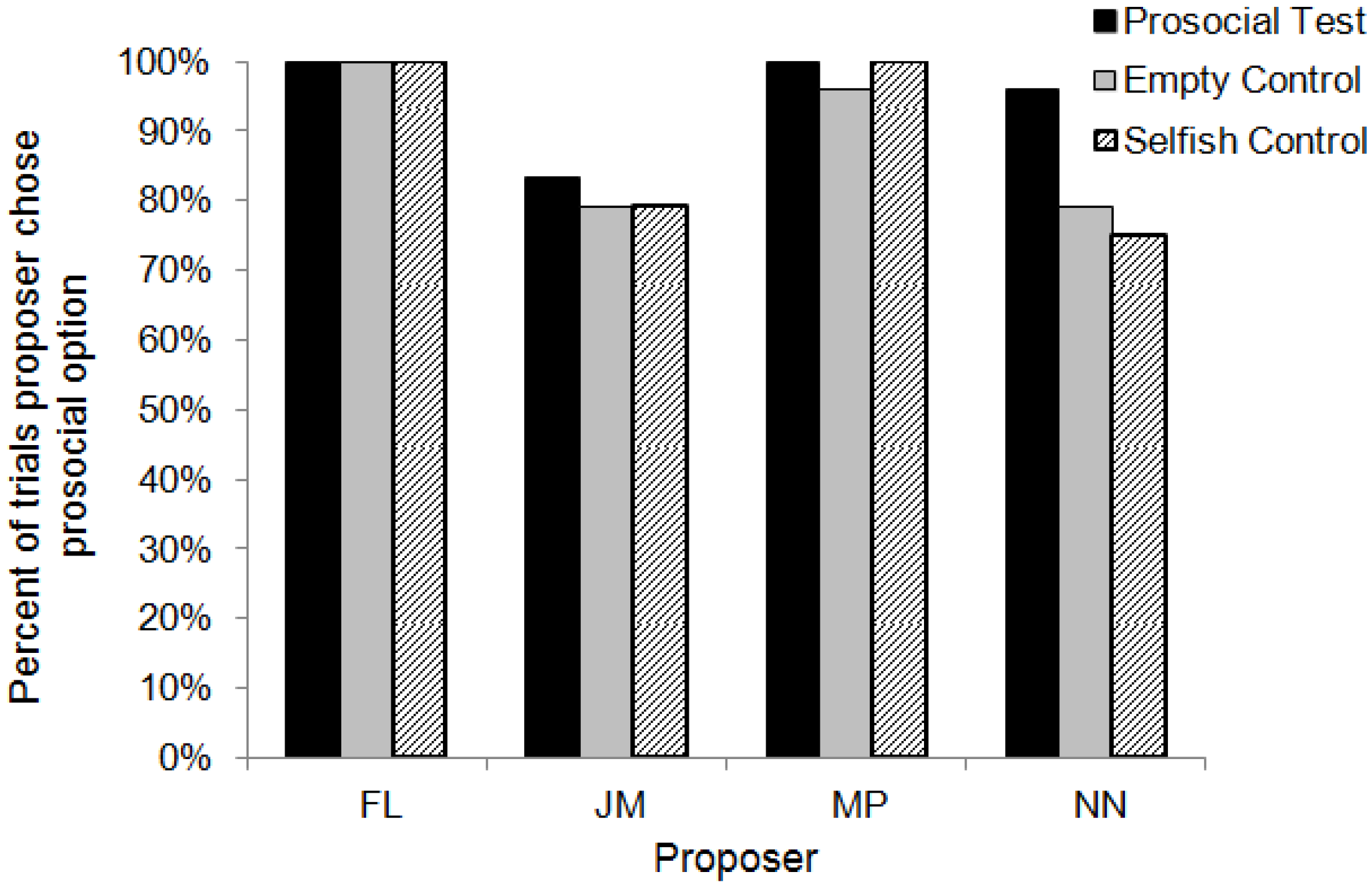
3. Results
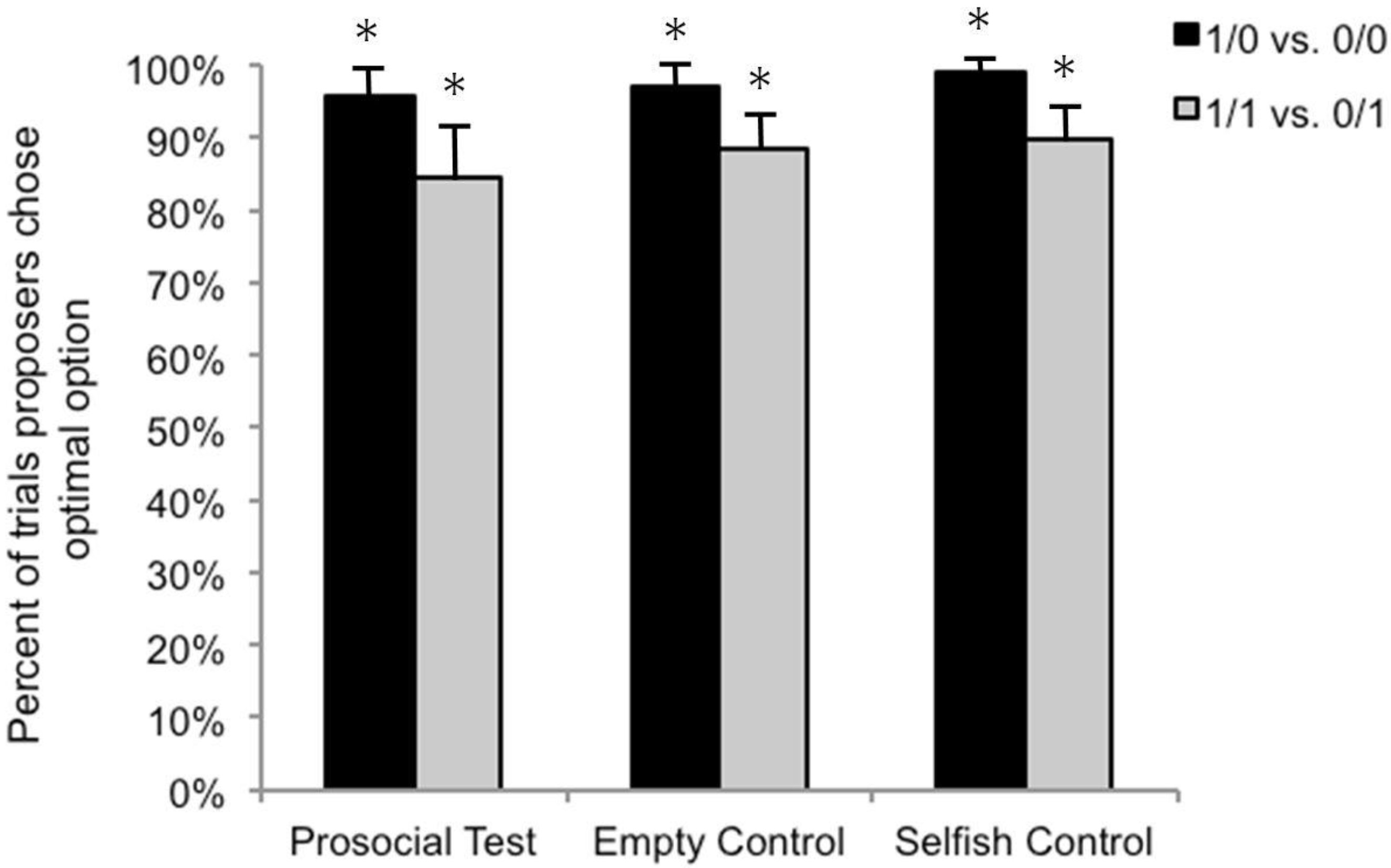
3.1. Stimulus Preference Control Sessions
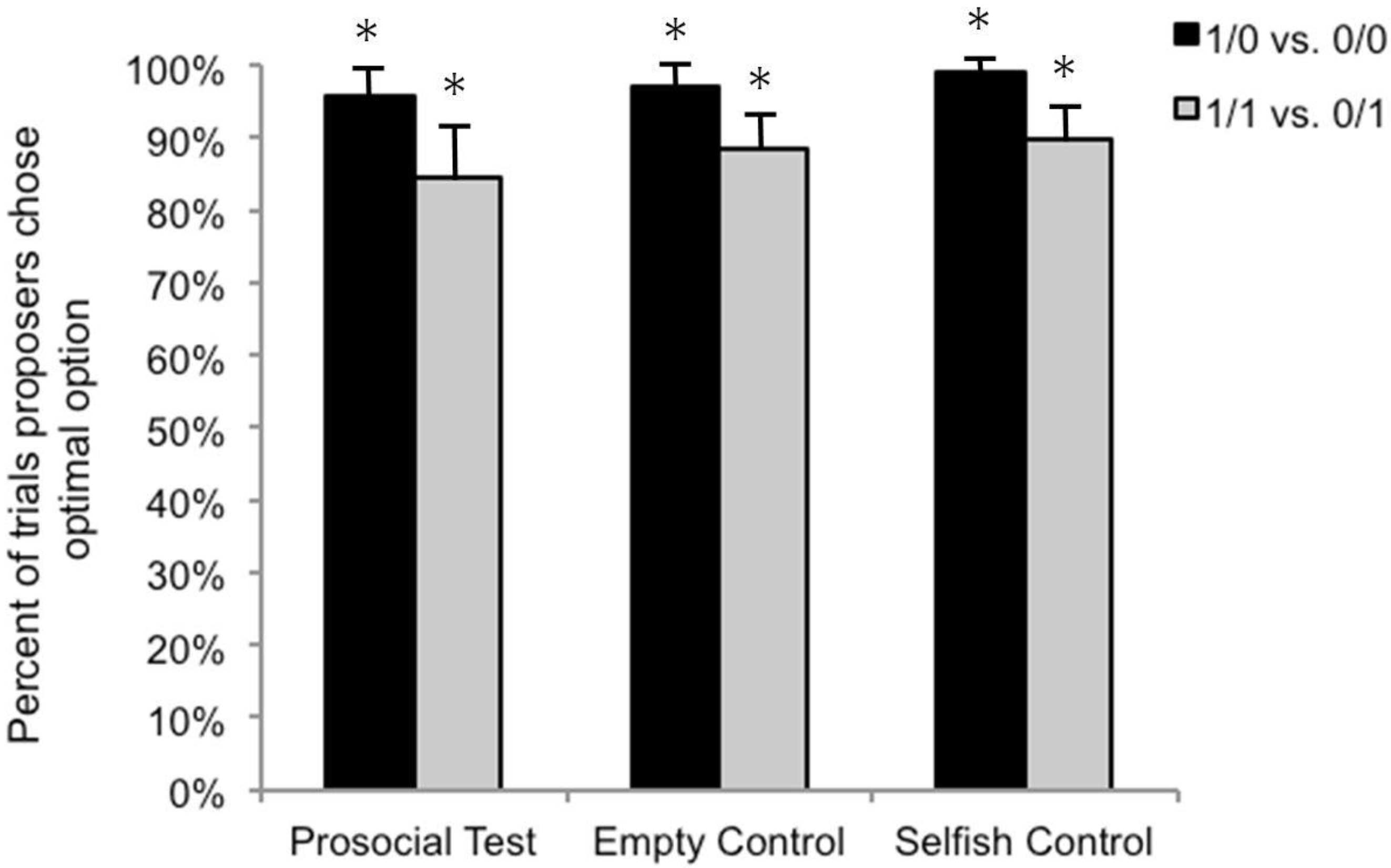
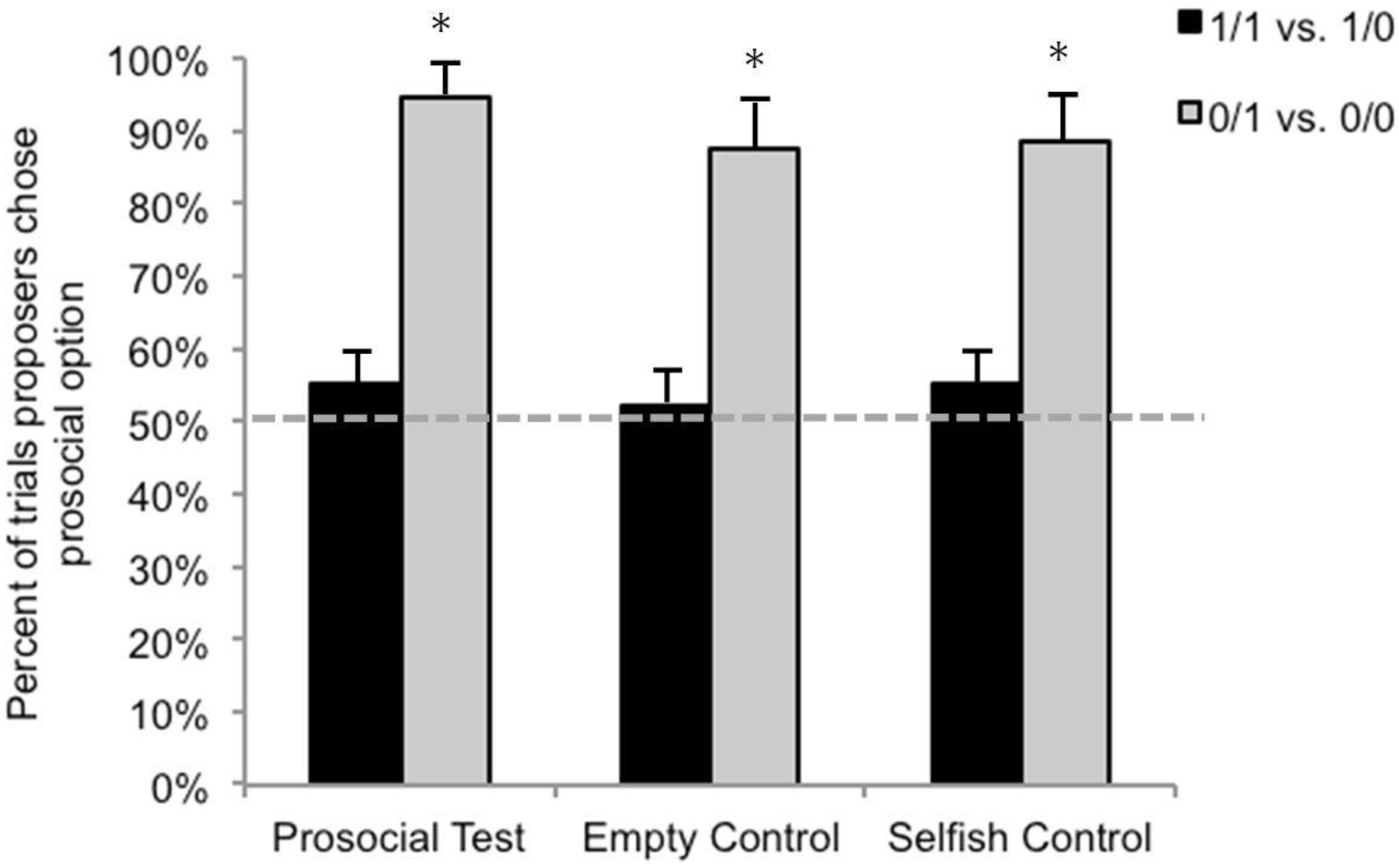
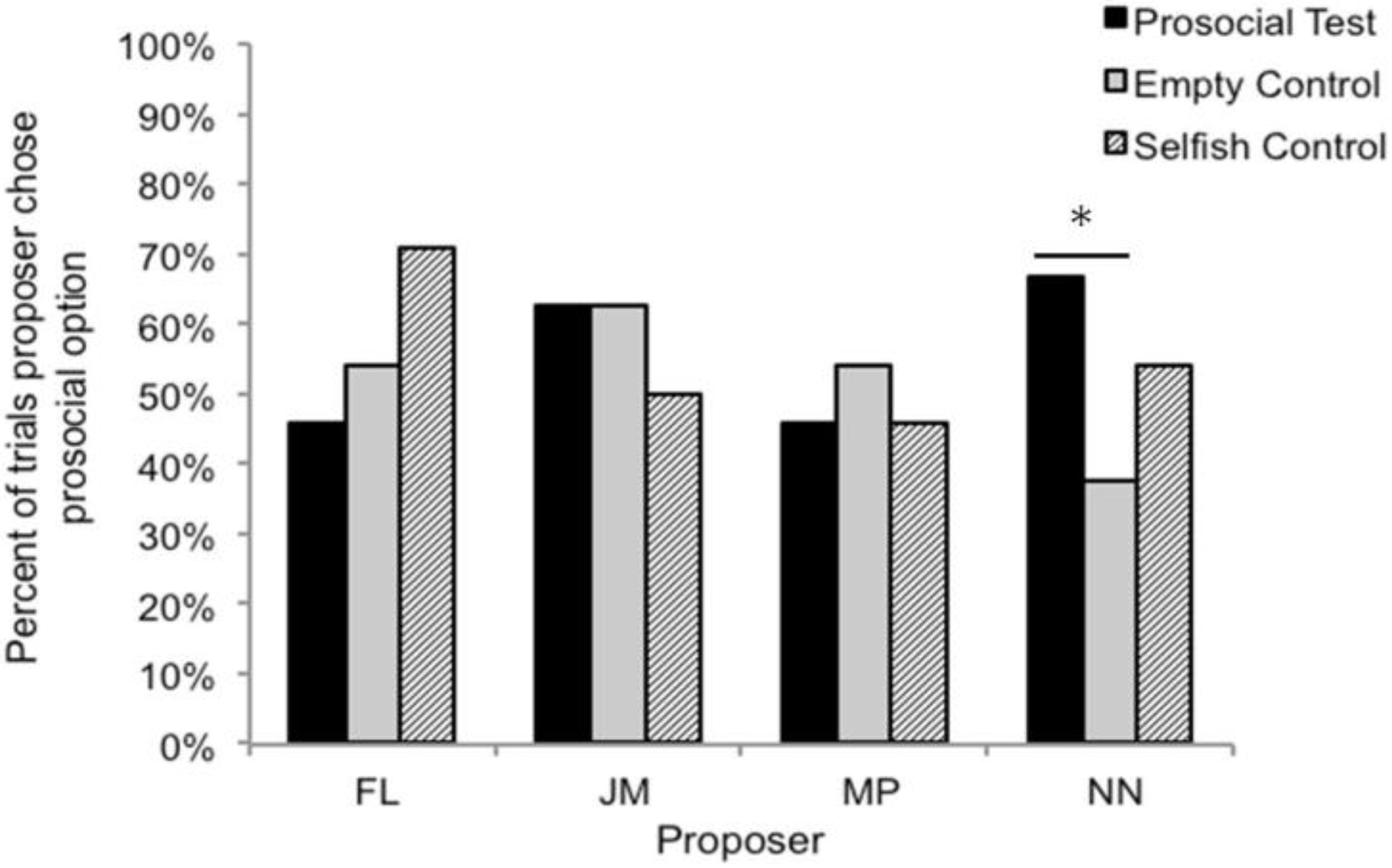
3.2. Test Sessions

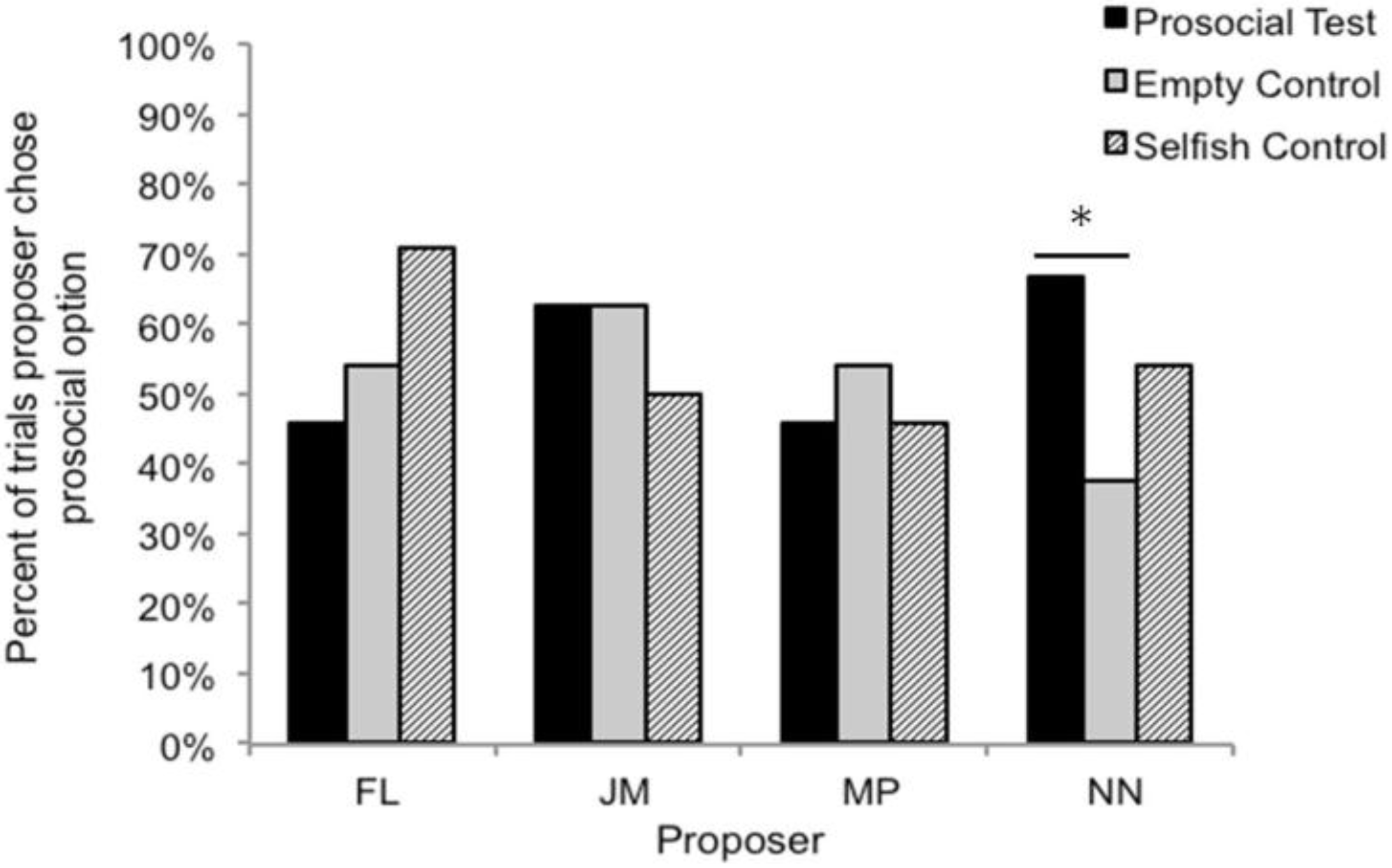
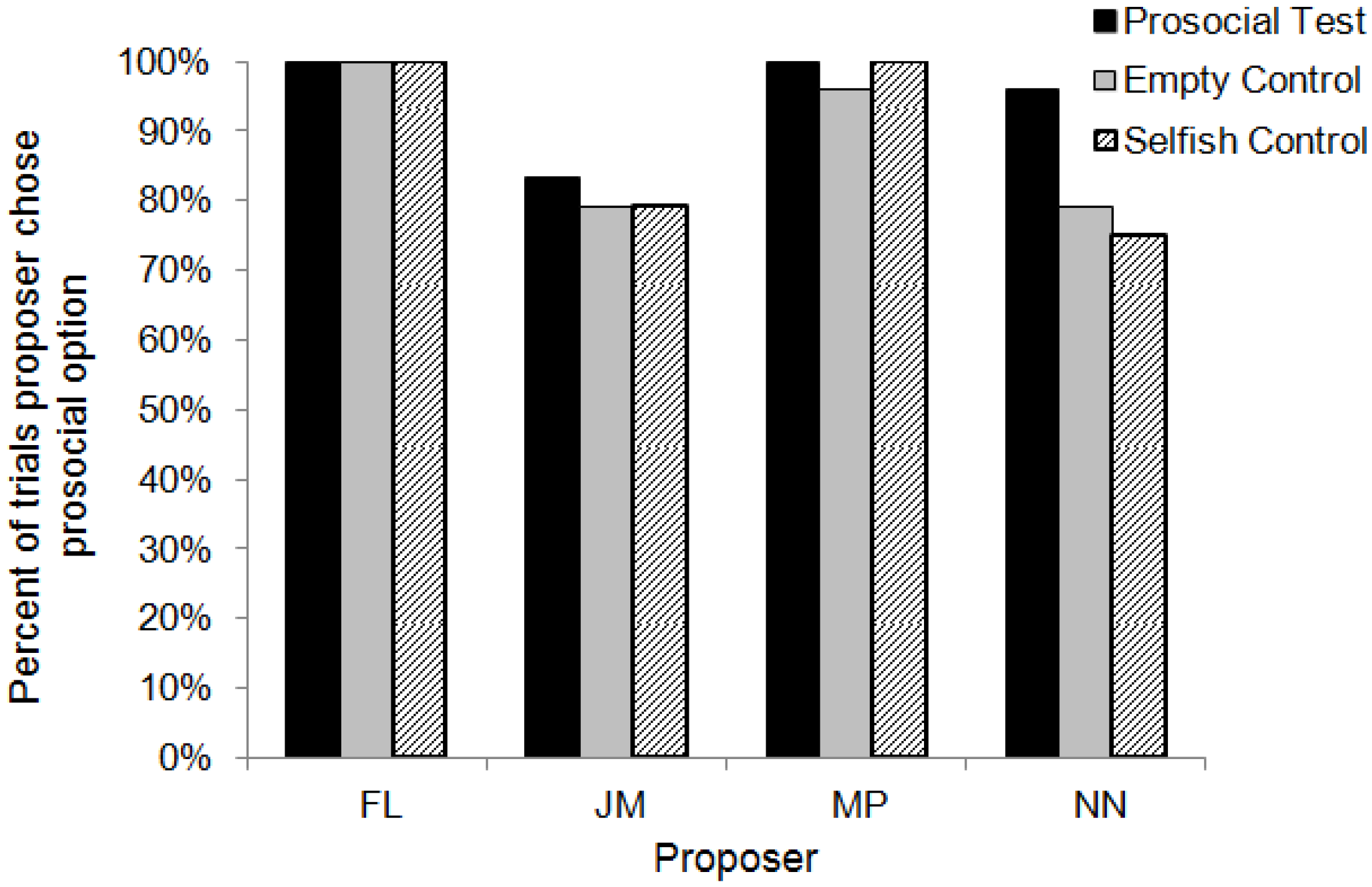
3.3. Comparison with Lakshminarayanan and Santos (2008)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Actor’s payoff regardless of choice | FL | JM | NN |
|---|---|---|---|---|
| Current study | Food reward | ns | ns | * |
| No food reward | ns | ns | + | |
| Lakshminarayanan and Santos (2008) | High-value food | ns | ns | ns |
| Low-value food | ns | * | ns |
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Brown, G.R.; Almond, R.E.A.; Bergen, Y. Begging, stealing, and offering: Food transfer in nonhuman primates. Adv. Stud. Behav. 2004, 34, 265–295. [Google Scholar] [CrossRef]
- Barnes, J.L.; Hill, T.; Langer, M.; Martinez, M.; Santos, L.R. Helping behaviour and regard for others in capuchin monkeys (Cebus apella). Biol. Letters. 2008, 4, 638–640. [Google Scholar] [CrossRef]
- Warneken, F.; Tomasello, M. Altruistic helping in human infants and young chimpanzees. Science 2006, 311, 1301–1303. [Google Scholar] [CrossRef]
- Warneken, F.; Hare, B.; Melis, A.P.; Hanus, D.; Tomasello, M. Spontaneous altruism by chimpanzees and young children. PloS Biol. 2007, 5, e184. [Google Scholar] [CrossRef]
- Yamamoto, S.; Humle, T.; Tanaka, M. Chimpanzees help each other upon request. PloS One 2009, 4, e7416. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, S.; Humle, T.; Tanaka, M. Chimpanzees’ flexible targeted helping based on an understanding of conspecifics’ goals. Proc. Natl. Acad. Sci. USA 2012, 108, 3588–3592. [Google Scholar] [CrossRef] [Green Version]
- De Waal, F.B.M.; Aureli, F. Consolation, reconciliation, and a possible cognitive difference between macaque and chimpanzee. In Reaching into Thought: The Minds of the Great Apes; Russon, A.E., Bard, K.A., Parker, S.T., Eds.; Cambridge University Press: Cambridge, UK, 1996; pp. 80–110. [Google Scholar]
- Silk, J.B.; House, B.R. Evolutionary foundations of human prosocial sentiments. Proc. Natl. Acad. Sci. 2011, 108, 10910–10917. [Google Scholar] [CrossRef]
- Brosnan, S.F.; Silk, S.B.; Henrich, J.; Mareno, M.C.; Lambeth, S.P.; Schapiro, S.J. Chimpanzees (Pan troglodytes) do not develop contingent reciprocity in an experimental task. Anim. Cogn. 2009, 12, 587–597. [Google Scholar] [CrossRef]
- Cronin, K.A.; Schroeder, K.K.; Rothwell, E.S.; Silk, J.B.; Snowdon, C.T. Cooperatively breeding cottontop tamarins (Saguinus oedipus) do not donate rewards to their long-term mates. J. Comp. Psychol. 2009, 123, 231–241. [Google Scholar] [CrossRef]
- Cronin, K.A.; Schroeder, K.K.; Snowdon, C.T. Prosocial behavior emerges independent of reciprocity in cottontop tamarins. Proc. Biol. Sci. 2010, 277, 3845–3851. [Google Scholar] [CrossRef]
- De Waal, F.B.M.; Leimgruber, K.; Greenberg, A.R. Giving is self-rewarding for monkeys. Proc. Natl. Acad. Sci. USA 2008, 105, 13685–13689. [Google Scholar] [CrossRef]
- Horner, V.; Carter, J.D.; Suchak, M.; de Waal, F.B.M. Spontaneous prosocial choice by chimpanzees. Proc. Natl. Acad. Sci. USA 2011, 108, 13847–13851. [Google Scholar]
- Jensen, K.; Hare, B.; Tomasello, M. What’s in it for me? Self-regard precludes altruism and spite in chimpanzees. Proc. Biol. Sci. 2006, 273, 1013–1021. [Google Scholar] [CrossRef]
- Lakshminarayanan, V.R.; Santos, L.R. Capuchin monkeys are sensitive to others’ welfare. Curr. Biol. 2008, 18, R999–R1000. [Google Scholar]
- Massen, J.J.M.; van den Berg, L.M.; Spruijt, B.M.; Sterck, E.H.M. Generous leaders and selfish underdogs: Prosociality in despotic macaques. PloS One 2010, 5, e9734. [Google Scholar]
- Massen, J.J.M.; Luyten, I.J.A.F.; Spruijt, B.M.; Sterck, E.H.M. Benefiting friends or dominants: prosocial choices mainly depend on rank position in long-tailed macaques (Macaca fascicularis). Primates 2011, 52, 237–247. [Google Scholar] [CrossRef]
- Silk, J.B.; Brosnan, S.F.; Vonk, J.; Henrich, J.; Povinelli, D.J.; Richardson, A.S.; Lambeth, S.P.; Mascaro, J.; Schapiro, S.J. Chimpanzees are indifferent to the welfare of unrelated group members. Nature 2005, 437, 1357–1359. [Google Scholar] [CrossRef]
- Stevens, J.R. Donor payoffs and other regarding preferences in cotton-top tamarins (Saguinus oedipus). Anim. Cogn. 2010, 13, 663–670. [Google Scholar] [CrossRef]
- Takimoto, A.; Kuroshima, H.; Fujta, K. Capuchin monkeys (Cebus apella) are sensitive to others’ reward: An experimental analysis of food-choice for conspecifics. Anim. Cogn. 2010, 13, 249–261. [Google Scholar] [CrossRef] [Green Version]
- Vonk, J.; Brosnan, S.F.; Silk, J.B.; Henrich, J.; Richardson, A.S.; Lambeth, S.P.; Schapiro, S.J.; Povinelli, D.J. Chimpanzees do not take advantage of very low cost opportunities to deliver food to unrelated group members. Anim. Behav. 2008, 75, 1757–1770. [Google Scholar] [CrossRef]
- Burkart, J.M.; Fehr, E.; Efferson, C.; van Schaik, C.P. Other-regarding preferences in a non-human primate: Common marmosets provision food altruistically. Proc. Natl. Acad. Sci. USA 2007, 104, 19762–19766. [Google Scholar] [CrossRef]
- Fragaszy, D.M.; Feuerstein, J.M.; Mitra, D. Transfers of food from adults to infants in tufted capuchins (Cebus apella). J. Comp. Psychol. 1997, 111, 194–200. [Google Scholar] [CrossRef]
- Hattori, Y.; Kuroshima, H.; Fujita, K. Cooperative problem solving by tufted capuchin monkeys (Cebus apella): Spontaneous division of labor, communication, and reciprocal altruism. J. Comp. Psychol. 2005, 119, 335–342. [Google Scholar] [CrossRef]
- Mendres, K.A.; de Waal, F.B.M. Capuchins do cooperate: The advantage of an intuitive task. Anim. Behav. 2000, 60, 523–529. [Google Scholar] [CrossRef]
- Basile, B.M.; Hampton, R.R. Monkeys recall and reproduce simple shapes from memory. Curr. Biol. 2011, 21, 775–778. [Google Scholar] [CrossRef]
- Smith, J.D.; Beran, M.J.; Redford, J.S.; Washburn, D.A. Dissociating uncertainty responses and reinforcement signals in the comparative study of uncertainty monitoring. J. Exp. Psychol. Gen. 2006, 135, 282–297. [Google Scholar] [CrossRef]
- Shepherd, S.V.; Deaner, R.O.; Platt, M.L. Social status gates social attention in monkeys. Curr. Biol. 2006, 16, 119–120. [Google Scholar] [CrossRef]
- Brosnan, S.F.; Wilson, B.; Beran, M.J. Old World monkeys are more similar to humans than New World monkeys when playing a coordination game. Proc. Biol. Sci. 2012, 279, 1522–1530. [Google Scholar] [CrossRef]
- Furlong, E.E.; Santos, L.R. “The onus of the bonus”: Large incentives impair capuchin monkey performance. Unpublished work. 2014. [Google Scholar]
- Cronin, K.A. Prosocial behavior in animals: The influence of social relationships, communication, and rewards. Animal Behav. 2012, 84, 1085–1093. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Drayton, L.A.; Santos, L.R. Insights into Intraspecies Variation in Primate Prosocial Behavior: Capuchins (Cebus apella) Fail to Show Prosociality on a Touchscreen Task. Behav. Sci. 2014, 4, 87-101. https://doi.org/10.3390/bs4020087
Drayton LA, Santos LR. Insights into Intraspecies Variation in Primate Prosocial Behavior: Capuchins (Cebus apella) Fail to Show Prosociality on a Touchscreen Task. Behavioral Sciences. 2014; 4(2):87-101. https://doi.org/10.3390/bs4020087
Chicago/Turabian StyleDrayton, Lindsey A., and Laurie R. Santos. 2014. "Insights into Intraspecies Variation in Primate Prosocial Behavior: Capuchins (Cebus apella) Fail to Show Prosociality on a Touchscreen Task" Behavioral Sciences 4, no. 2: 87-101. https://doi.org/10.3390/bs4020087