Cretaceous-Paleogene Dinoflagellate Biostratigraphy and the Age of the Clayton Formation, Southeastern Missouri, USA

Abstract
:1. Introduction
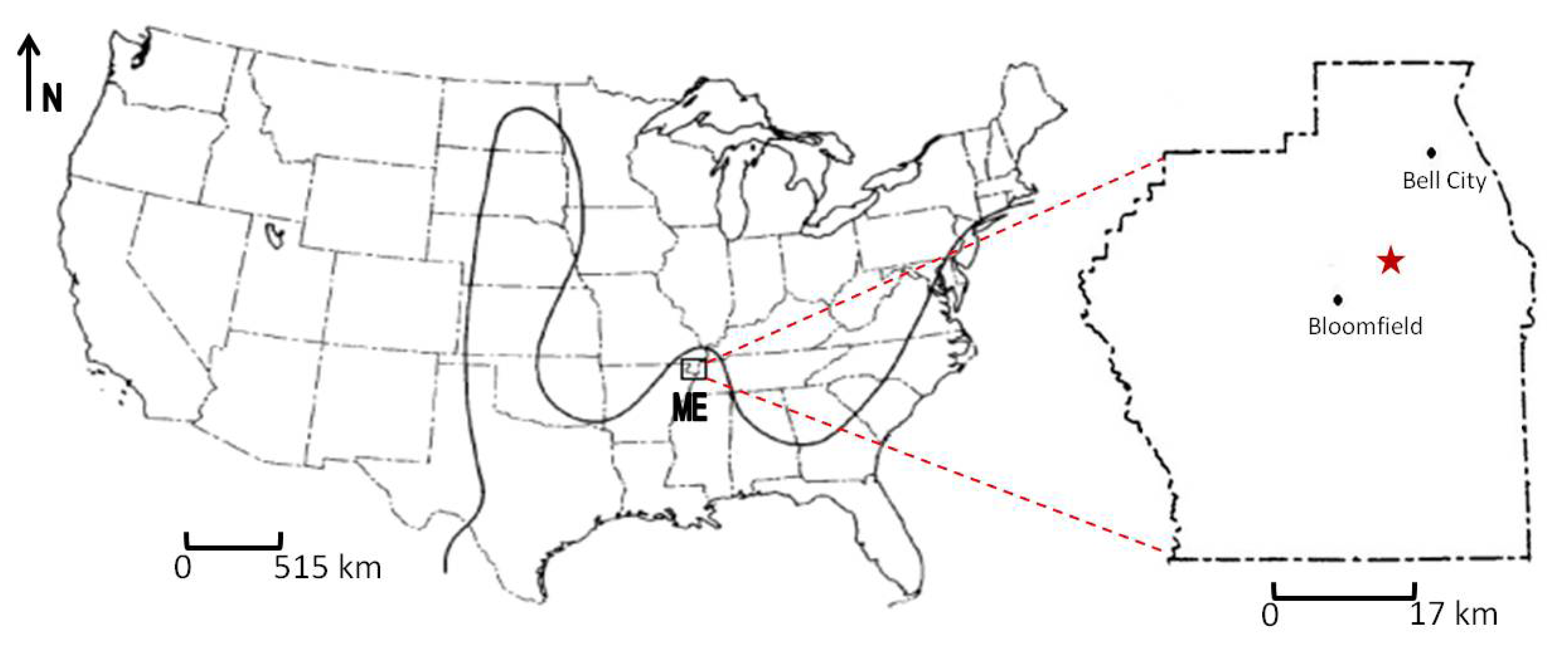
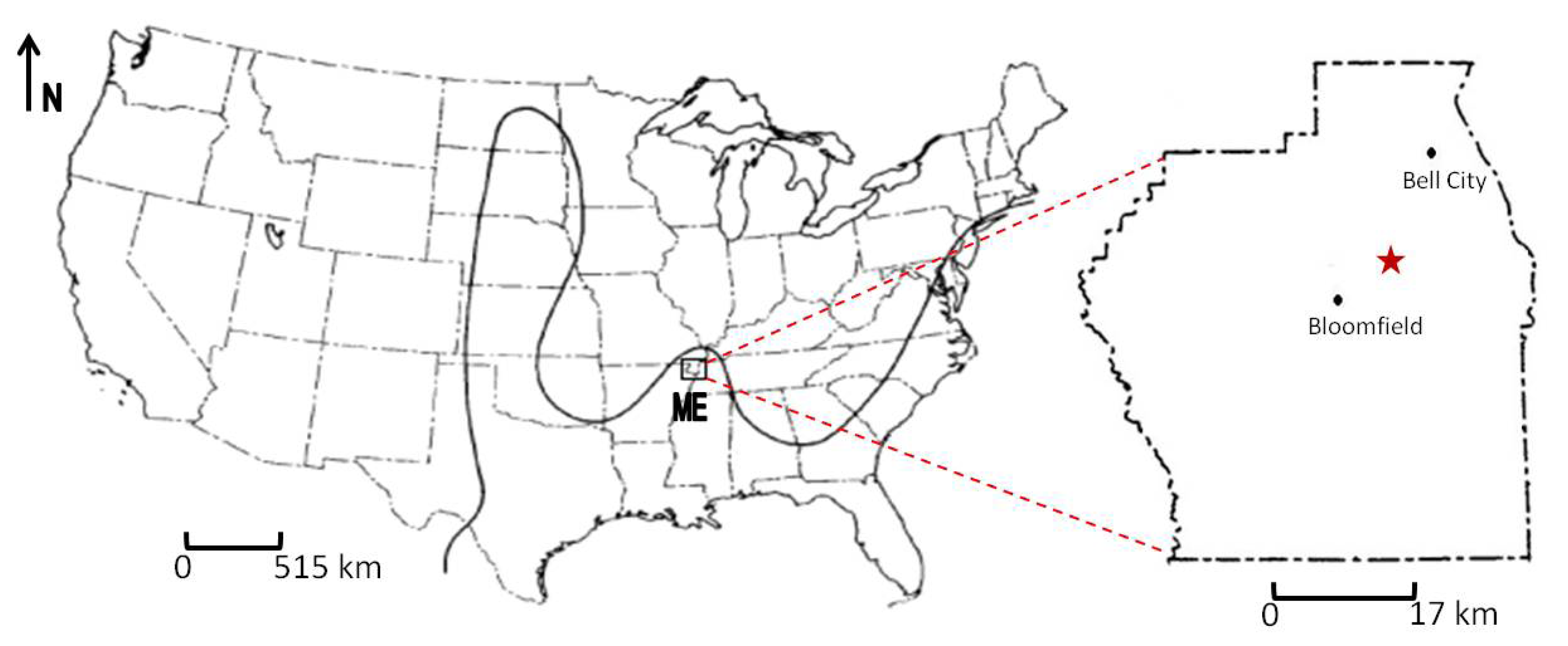
2. Geologic Setting
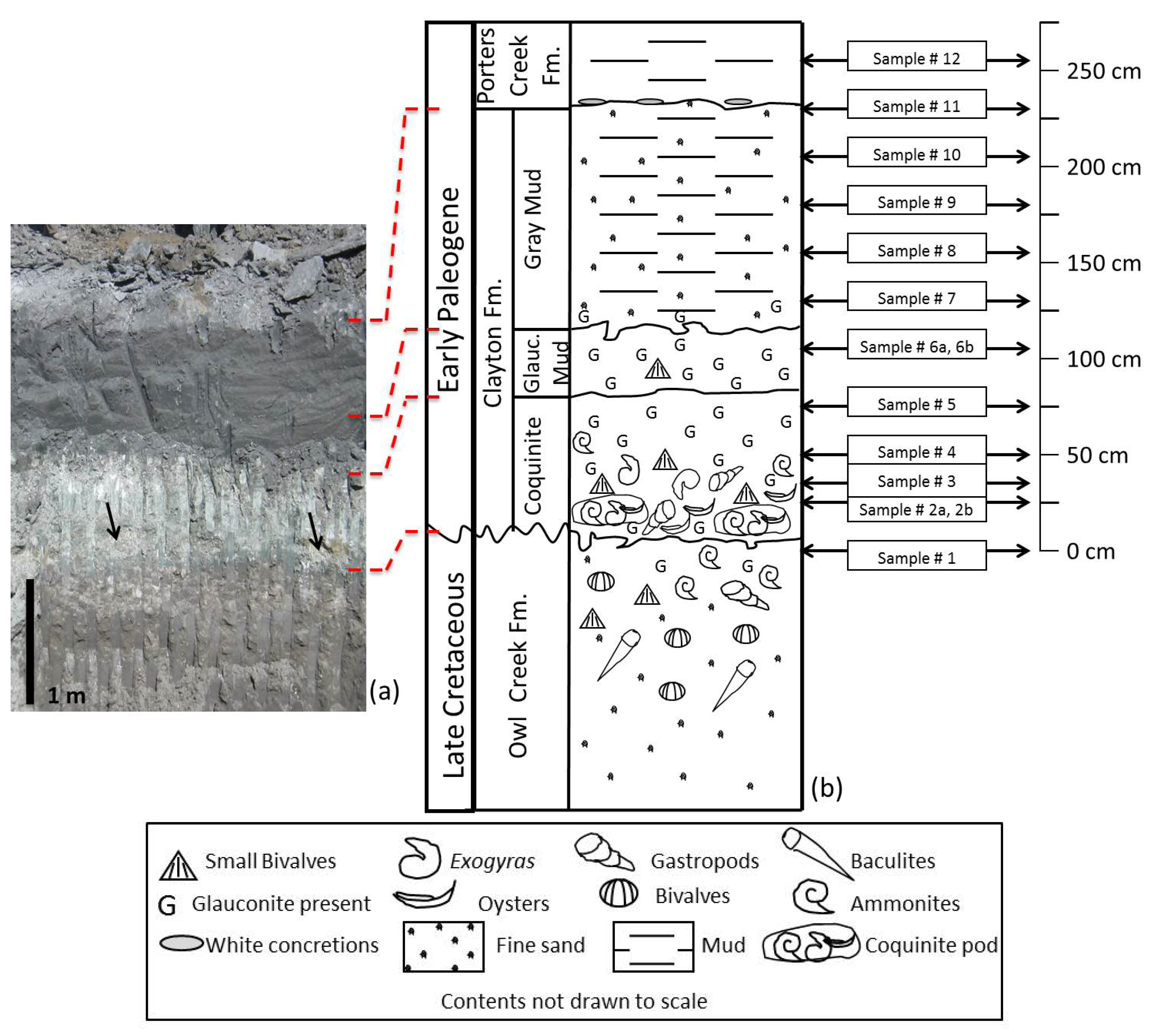
3. K-Pg Boundary Sequence at Bloomfield, Missouri
3.1. Owl Creek Formation
3.2. Clayton Formation
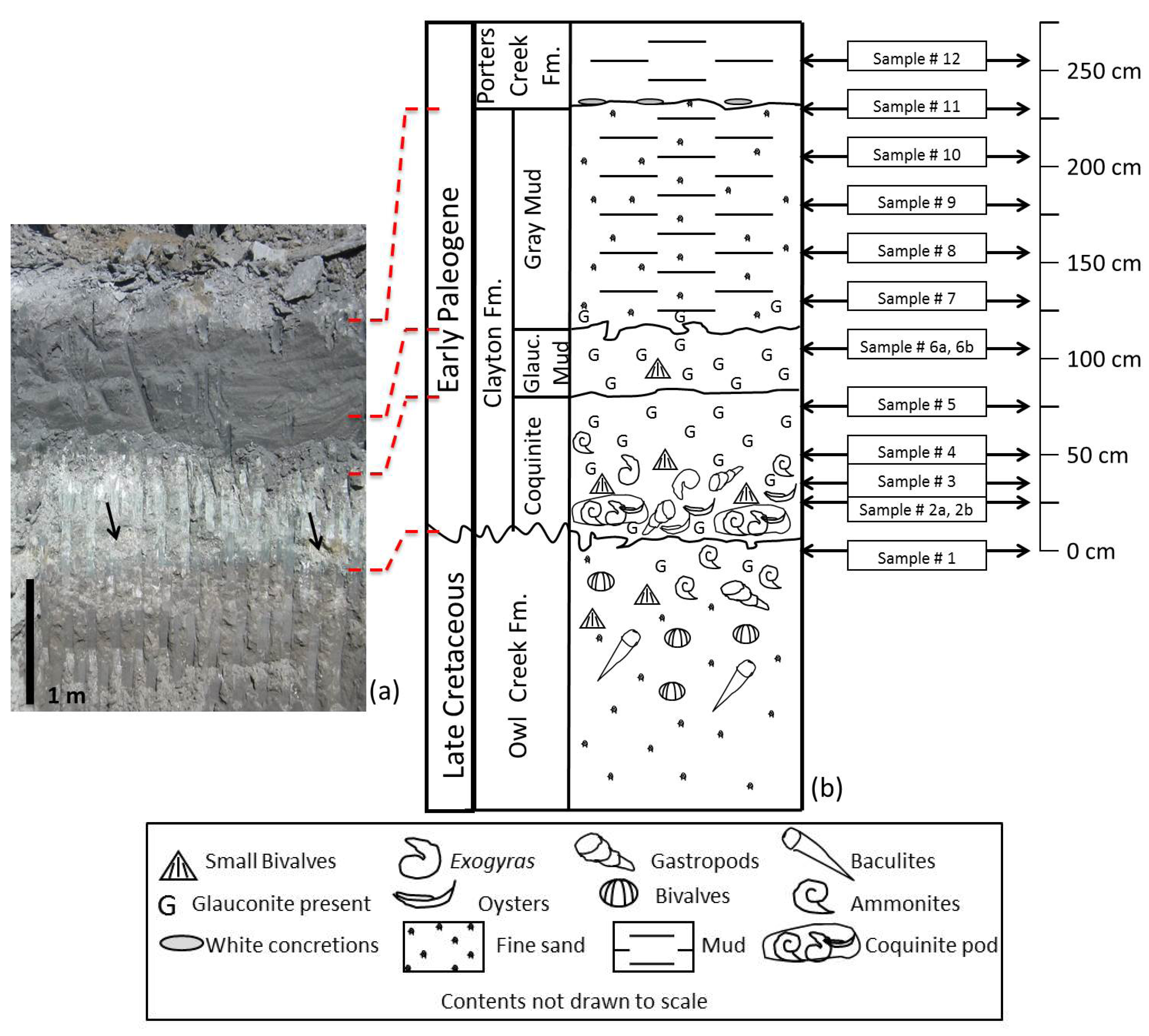

3.3. Porters Creek Formation
4. Methods
4.1. Collecting Procedures

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
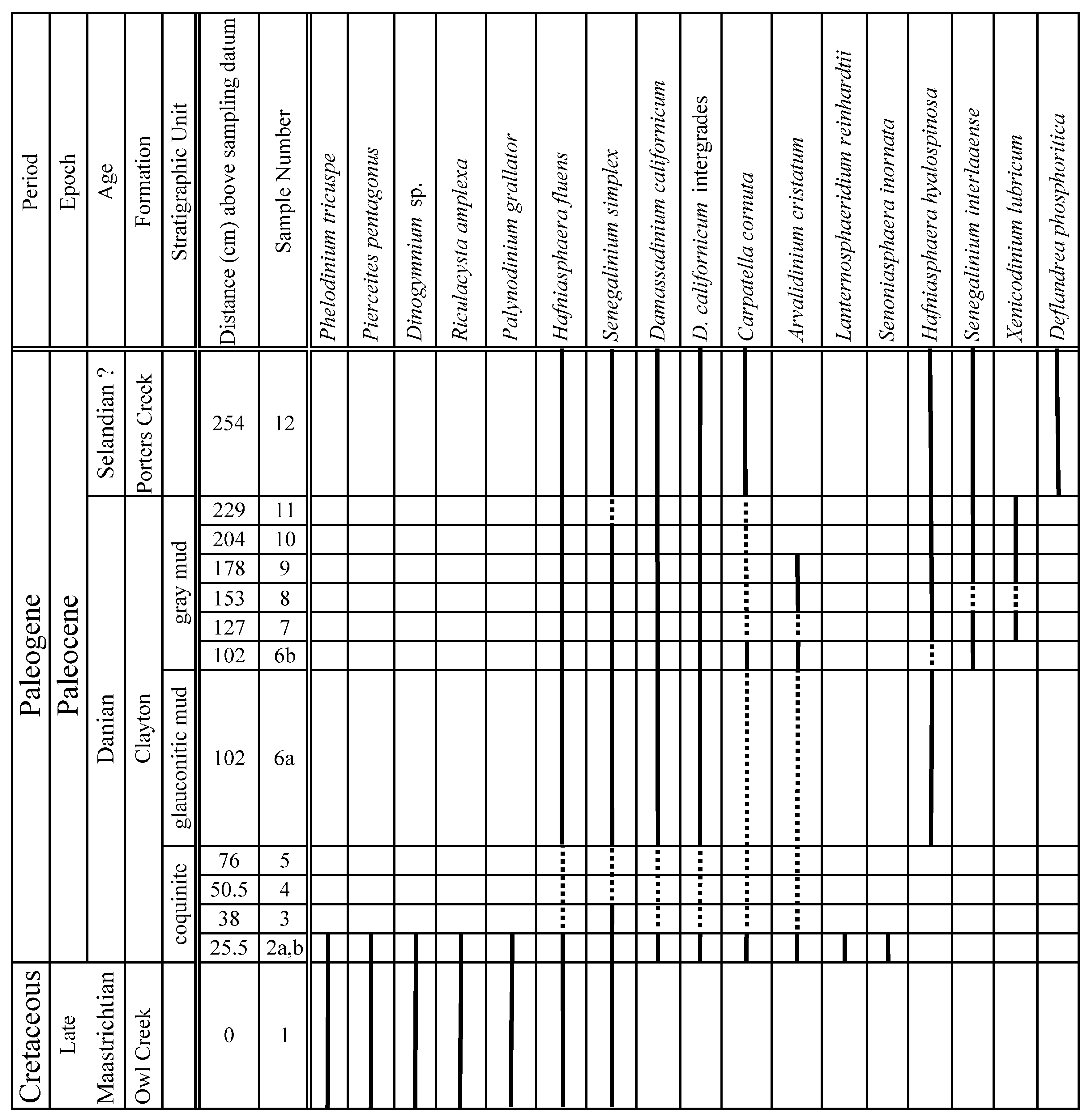
| Sample # | 1 | 2a | 2b | 3 | 4 | 5 | 6a | 6b | 7 | 8 | 9 | 10 | 11 | 12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Horizon sampled above 0 cm datum | 0 | 25.5 | 25.5 | 38 | 50.5 | 76 | 101.5 | 101.5 | 127 | 152.5 | 178 | 203.5 | 229 | 254 |
| # Of slides analyzed | 2 | 3 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Microfossil recovery | High | Low to High | Low to High | Low | Low | Low | High | High | High | High | High | High | High | High |
4.2. Palynological Preparation Procedures
4.3. Microscope Procedures
5. Results: Marine Palynomorph Occurrences
5.1. Late Cretaceous Taxa
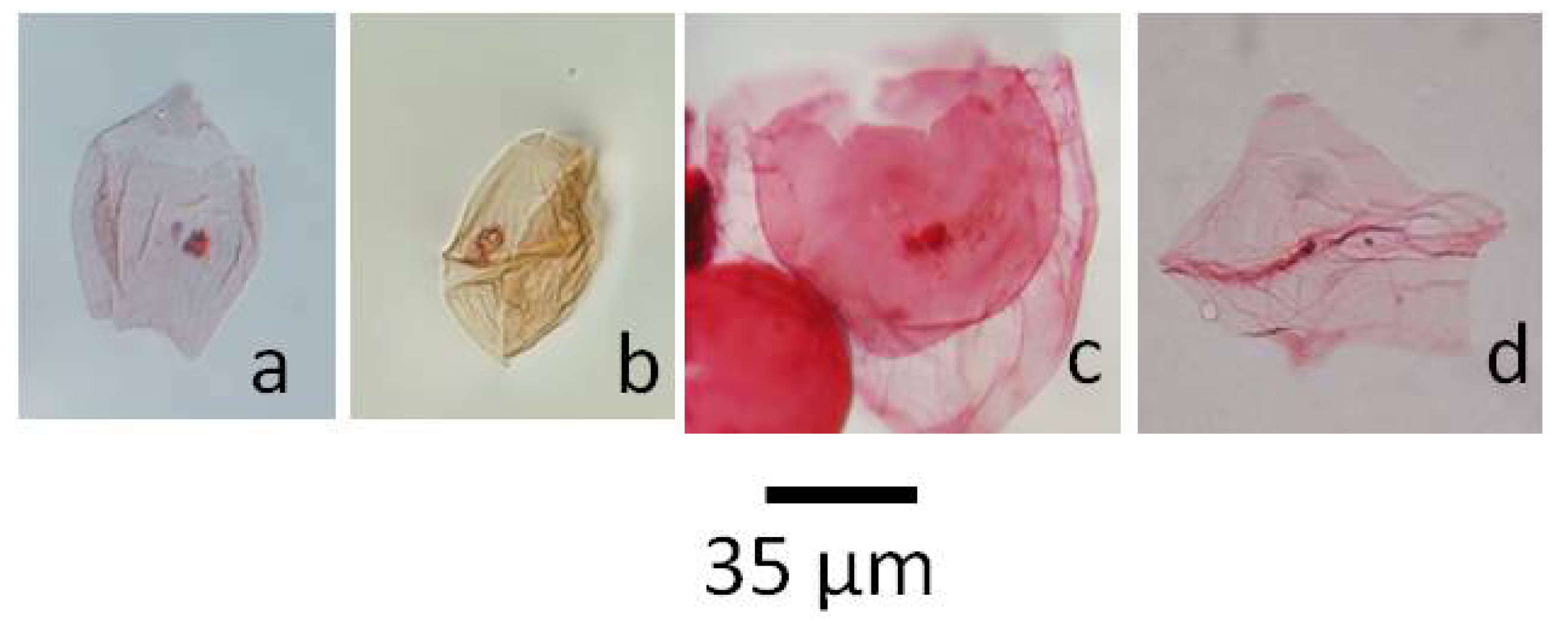
5.1.1. Pierceites pentagonus, Figure 4a
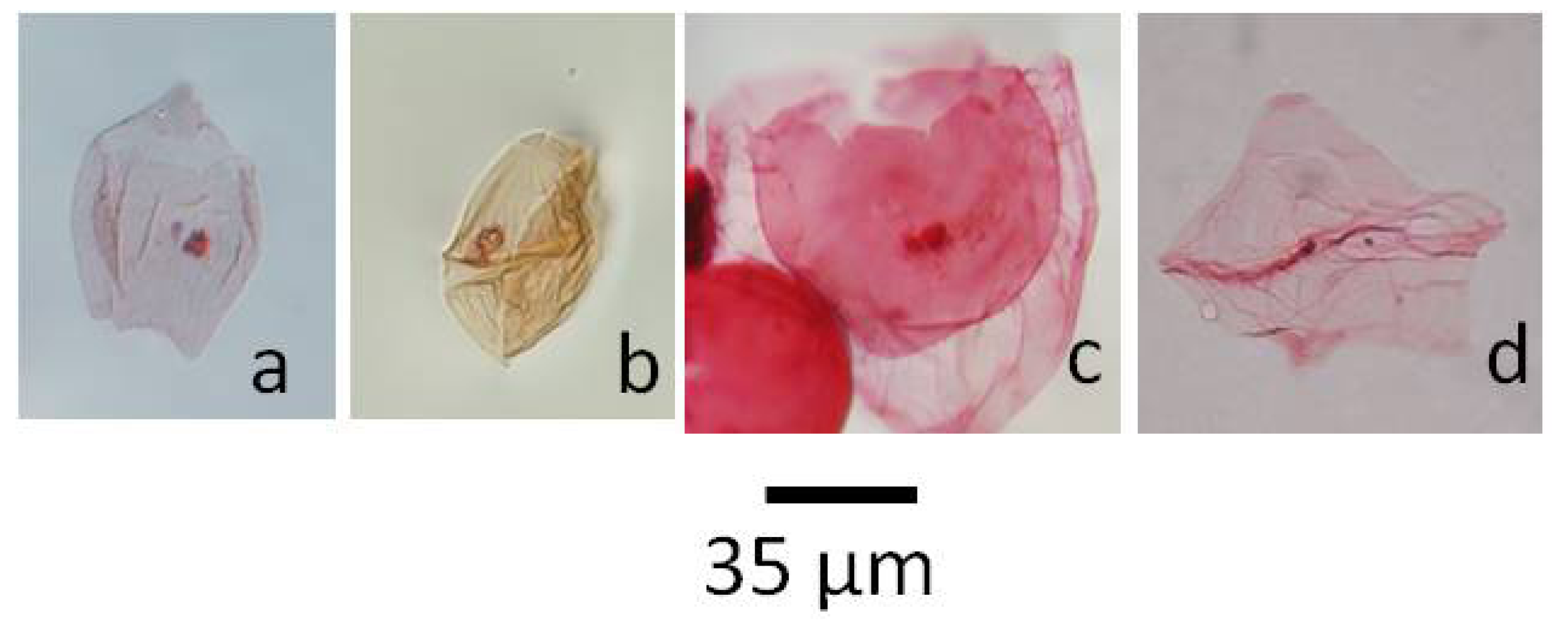
5.1.2. Dinogymnium sp., Figure 4b
5.1.3. Riculacysta amplexa, Figure 4c
5.1.4. Phelodinium tricuspe, Figure 4d
5.2. Paleocene Taxa
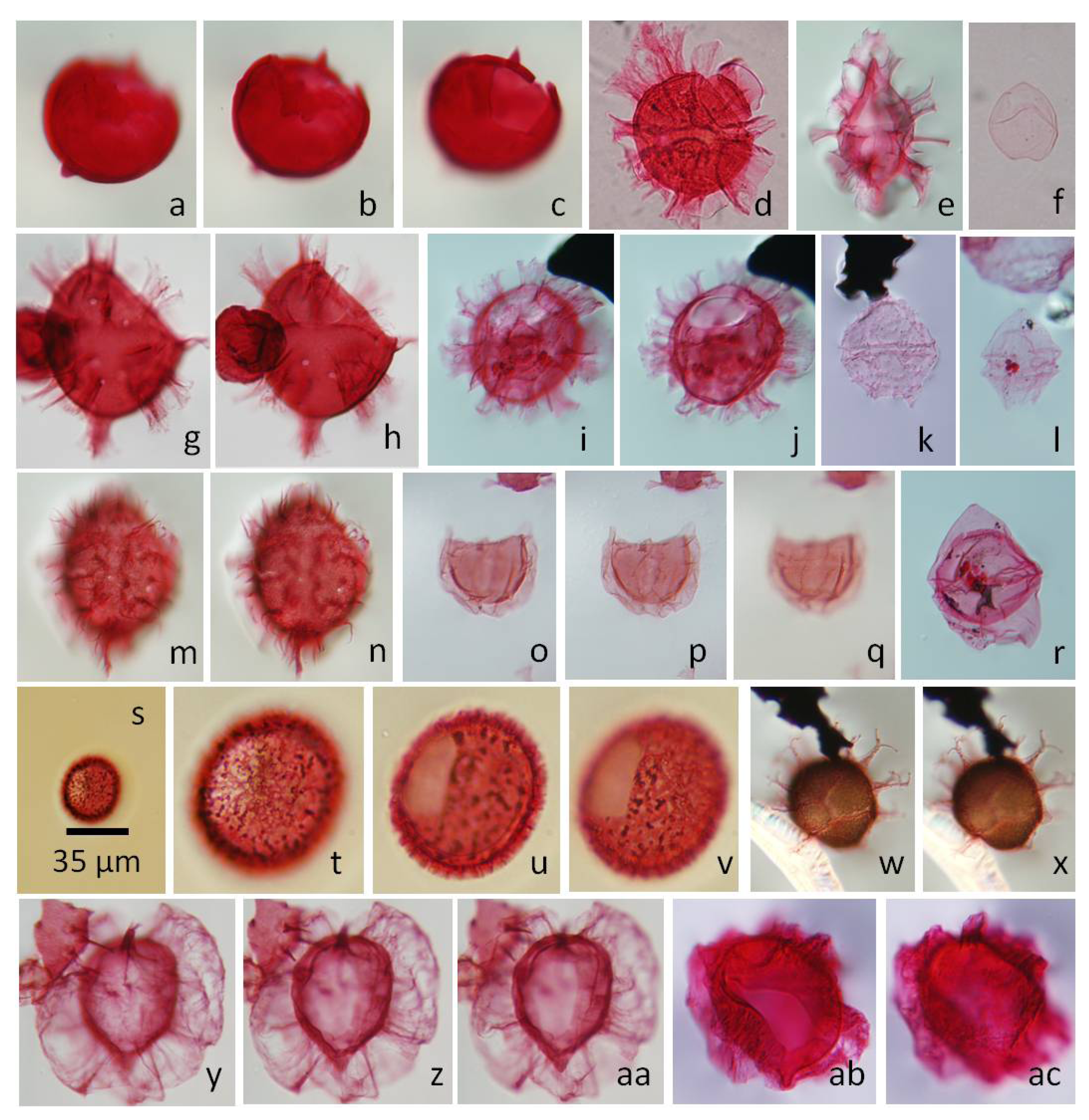
5.2.1. Carpatella cornuta, Figure 5a–c
5.2.2. Damassadinium californicum, Figure 5d,e,g,h
5.2.3. Senoniasphaera inornata, Figure 5o–q
5.2.4. Xenocodinium lubricum, Figure 5s–v
5.2.5. Lanternosphaeridium reinhardtii, Figure 5y,z,aa
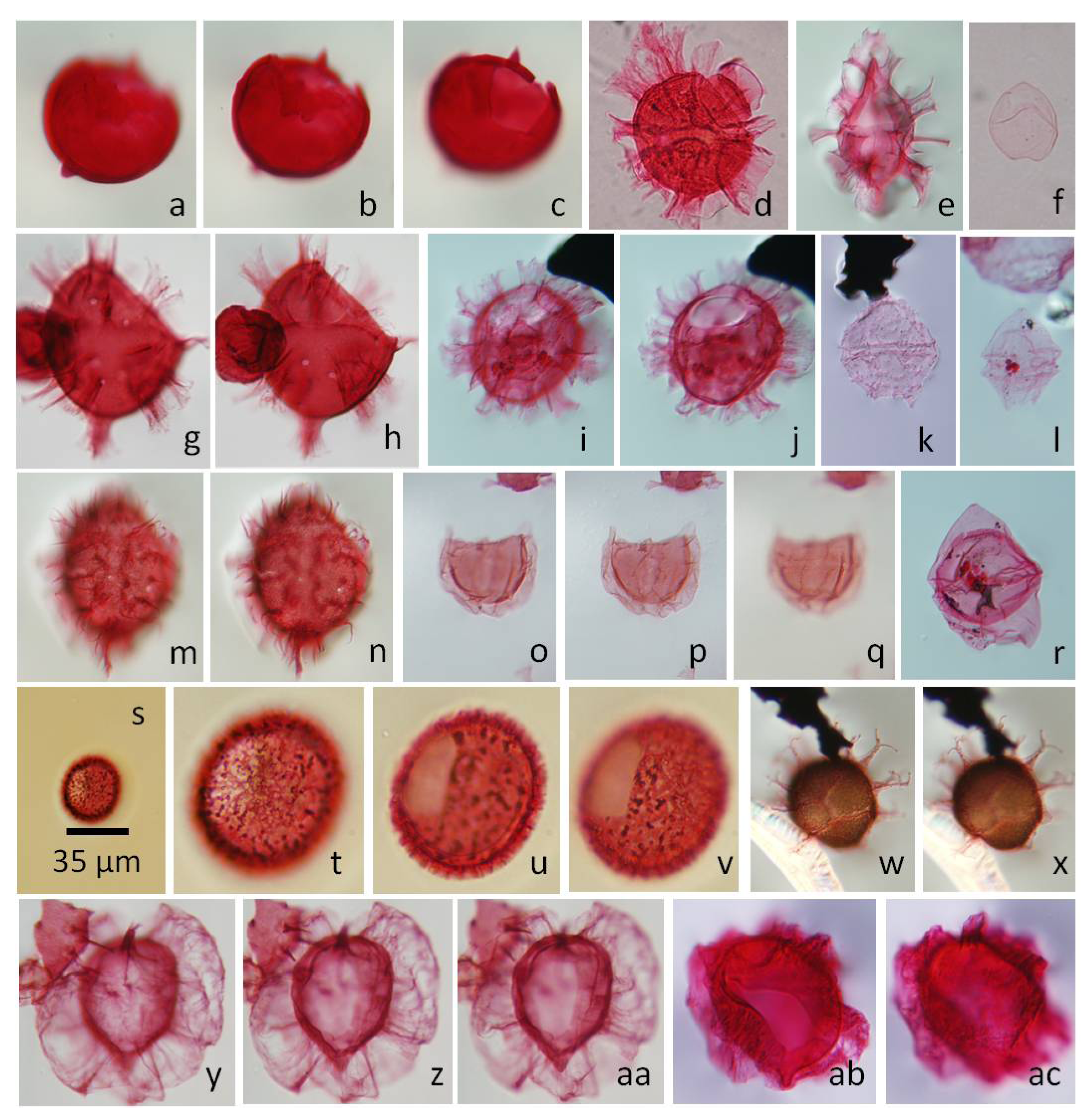
5.2.6. Senegalinium simplex, Figure 5f
5.2.7. Hafniasphaera hyalospinosa, Figure 5w,x
5.2.8. Arvalidinium cristatum, Figure 5k
5.2.9. Senegalinium interlaaense, Figure 5l
5.2.10. Deflandrea phosphoritica, Figure 5r
5.3. Index Species for the Early Danian and the K-Pg Boundary
5.4. K-Pg Boundary-Crossing Taxa
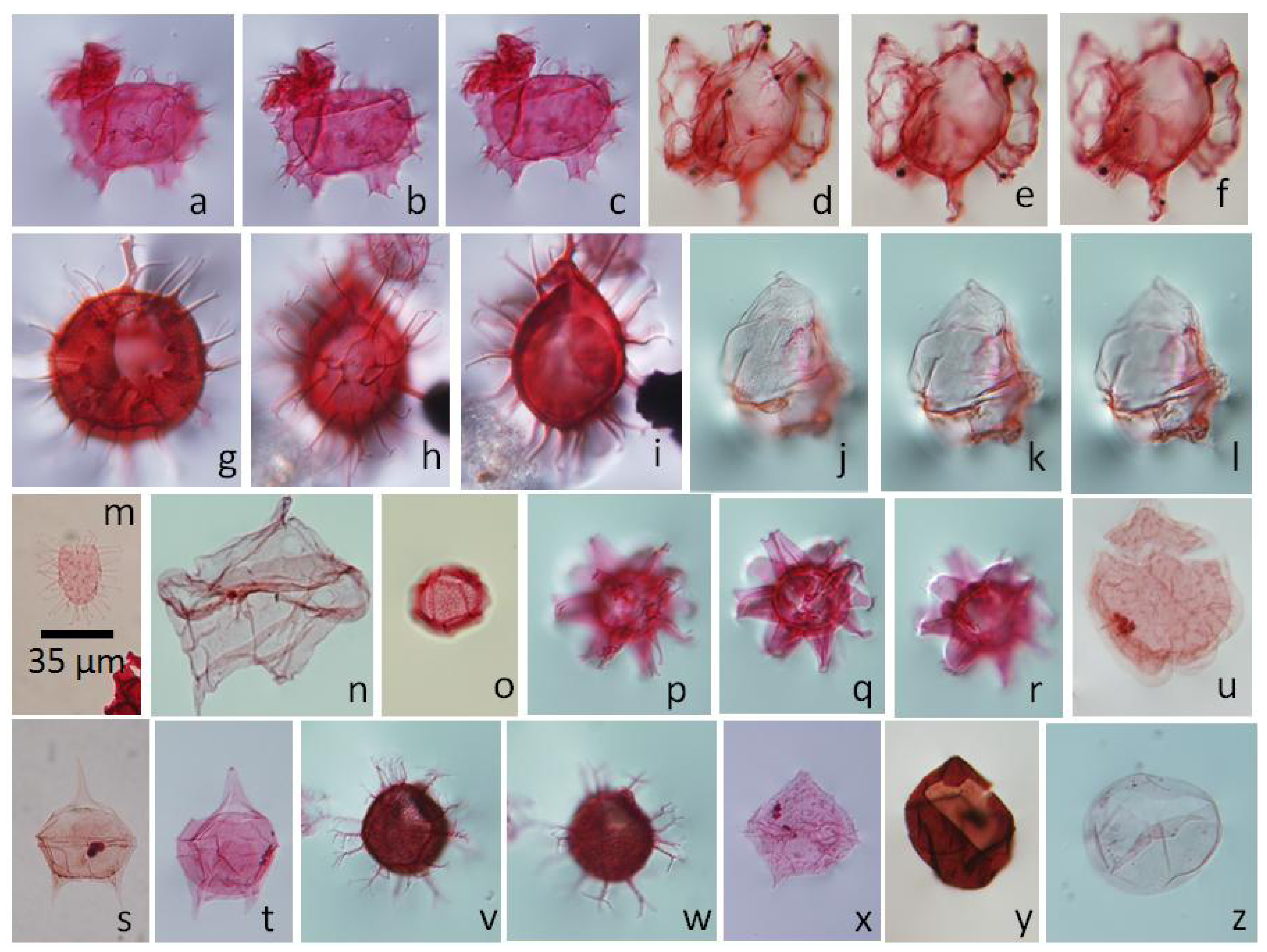
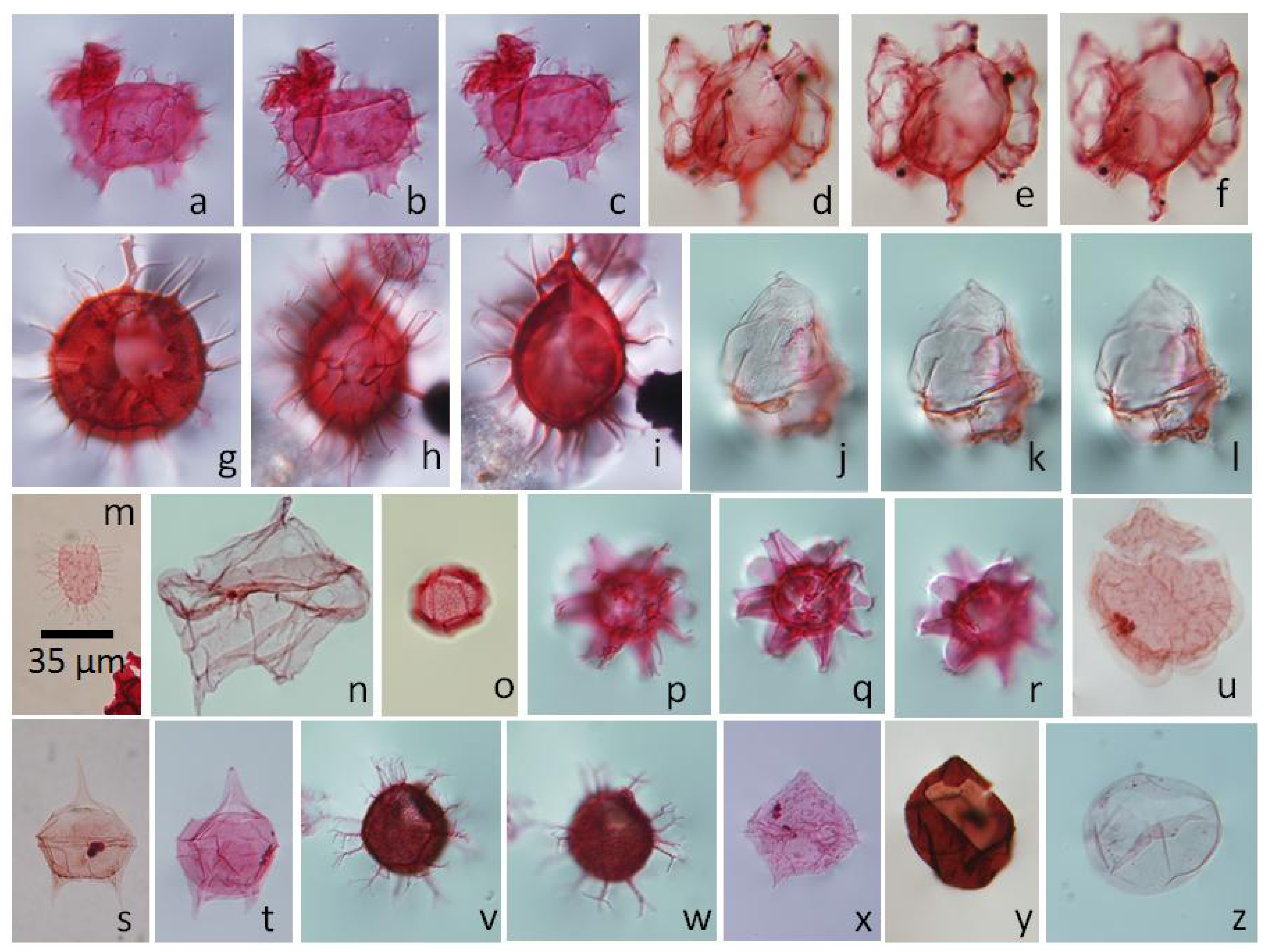
5.4.1. Palynodinium grallator, Figure 6a–c
5.4.2. Hafniasphaera fluens, Figure 6v,w
6. Discussion
6.1. Age of the Cretaceous-Paleogene Sequence at Bloomfield, Missouri

6.1.1. Age of the Owl Creek Formation
6.1.2. Age of the Clayton Coquinite
6.1.3. Age of the Glauconitic and Gray Muds of the Clayton Formation
6.1.4. Age of Basal Porters Creek Formation
6.2. Shell Concentrations and the Clayton Coquinite
6.3. The Chicxulub Impact and the Clayton Coquinite
6.4. The Origin of the Clayton Coquinite
Sea Level Change
7. Conclusions
Acknowledgments
Conflicts of Interest
References
- Pryor, W.A. Mesozoic-Cenozoic History of the Mississippi Embayment. In Guidebook to the Geology in the Vicinity of Cape Girardeau, Missouri Including Crowleys Ridge; Johnson, C.H., Ed.; Association of Missouri Geologists: Cape Girardeau, MO, USA, 1962; pp. 42–48. [Google Scholar]
- Pryor, W.A.; Glass, H.D. Cretaceous-Tertiary clay mineralogy of the upper Mississippi Embayment. J. Sediment. Petrol. 1961, 31, 38–51. [Google Scholar]
- Harrison, R.W.; Litwin, R.J. Campanian coastal plain sediments in southeastern Missouri and southern Illinois—Significance to the early geologic history of the northern Mississippi Embayment. Cretac. Res. 1997, 18, 687–696. [Google Scholar] [CrossRef]
- Cushing, E.M.; Boswell, E.H.; Hosman, R.L. General Geology of the Mississippi Embayment; Geological Survey Professional Paper 448-B; United States Government Printing Office: Washington, DC, USA, 1964. [Google Scholar]
- Tschudy, R.H. Palynology of the Cretaceous-Tertiary Boundary in the Northern Rocky Mountain and Mississippi Embayment Regions. In Symposium on Palynology of the Late Cretaceous and Early Tertiary; Kosanke, R.M., Cross, A.T., Eds.; Special Papers Volume 127; Geological Society of America: Boulder, CO, USA, 1971; pp. 65–111. [Google Scholar]
- Moshkovitz, S.; Habib, D. Calcareous nannofossil and dinoflagellate stratigraphy of the Cretaceous-Tertiary boundary, Alabama and Georgia. Micropaleontology 1993, 39, 167–191. [Google Scholar] [CrossRef]
- Campbell, C.E.; Oboh-Ikuenobe, F.E.; Eifert, T.L. Megatsunami Deposit in Cretaceous-Paleogene Boundary Interval of Southeastern Missouri; Special Papers Volume 437; Geological Society of America: Boulder, CO, USA, 2008; pp. 189–198. [Google Scholar]
- Kennedy, W.J.; Landman, N.H.; Christensen, W.K.; Cobban, W.A.; Hancock, J.M. Marine connections in North America during the late Maastrichtian: Palaeogeographic and palaeobiogeographic significance of Jeletzkytes nebrascense Zone cephalopod fauna from the Elk Butte Member of the Pierre Shale, SE South Dakota and NE Nebraska. Cretac. Res. 1998, 19, 745–775. [Google Scholar] [CrossRef]
- Grohskopf, J.G. Subsurface Geology of Mississippi Embayment of Southeast Missouri; Reports 37; Missouri Geological Survey and Water Resources: Rolla, MO, USA, 1955. [Google Scholar]
- Stephenson, L.W. Owl Creek (Upper Cretaceous) Fossils from Crowley’s Ridge, Southeastern Missouri; Geological Survey Professional Paper 274-E; United States Government Printing Office: Washington, DC, USA, 1955; pp. 97–137. [Google Scholar]
- Matthes, F.E. Cretaceous sediments in Crowleys Ridge, southeastern Missouri. AAPG Bull. 1933, 17, 1003–1015. [Google Scholar]
- Stephenson, L.W.; Monroe, W.H. Prairie Bluff chalk and Owl Creek formation of eastern Gulf region. AAPG Bull. 1937, 21, 806–809. [Google Scholar]
- Jones, E.L. Palynology of the Midway-Wilcox boundary in south-central Arkansas. GCAGS Trans. 1962, 12, 285–294. [Google Scholar]
- Frederiksen, N.O.; Bybell, L.M.; Christopher, R.A.; Crone, A.J.; Edwards, L.E.; Gibson, T.G.; Hazel, J.E.; Repetski, J.E.; Russ, D.P.; Smith, C.C.; et al. Biostratigraphy and paleoecology of lower Paleozoic, Upper Cretaceous, and lower Tertiary rocks in U.S. Geological Survey New Madrid test wells, southeastern Missouri. Tulane Stud. Geol. Paleontol. 1982, 17, 23–45. [Google Scholar]
- Harrison, R.W.; Litwin, R.J.; Repetski, J.E.; Mason, D.; Schultz, A. Results of Drilling in the English Hill Area, Benton Hills, Scott County, Missouri; Open-File Report 96-44; U.S. Geological Survey: Reston, VA, USA, 1996. [Google Scholar]
- Eifert, T.L. The Cretaceous-Paleogene Transition in the Northern Mississippi Embayment, S.E. Missouri: Palynology, Micropaleontology, and Evidence of a Mega-Tsunamite Deposit. Ph.D. Thesis, Missouri University of Science and Technology, Rolla, MO, USA, 2009. [Google Scholar]
- Oboh-Ikuenobe, F.E.; Spencer, M.K.; Campbell, C.E.; Haselwander, R.D. A portrait of Late Maastrichtian and Paleocene palynoflora and paleoenvironment in the northern Mississippi Embayment, southeastern Missouri. Palynology 2012, 36, 63–79. [Google Scholar] [CrossRef]
- Wade, B. The Fauna of the Ripley Formation on Coon Creek, Tennessee; Geological Survey Professional Paper 137; United States Government Printing Office: Washington, DC, USA, 1926. [Google Scholar]
- Farrar, W. The Cretaceous and Tertiary Geology (of Southeast Missouri); 58th General Assembly Biennial Report; Missouri Geological Survey and Water Resources: Rolla, MO, USA, 1935. [Google Scholar]
- Farrar, W.; McManamy, L. The Geology of Stoddard County, Missouri; 59th General Assembly Biennial Report; Missouri Geological Survey and Water Resources: Rolla, MO, USA, 1937. [Google Scholar]
- Cooper, C.L. Smaller Foraminifera from the Porters Creek formation (Paleocene) of Illinois. J. Paleontol. 1944, 18, 343–354. [Google Scholar]
- Dockery, D.T., III. Punctuated succession of Paleogene mollusks in the northern Gulf Coastal Plain. Palaios 1986, 1, 582–589. [Google Scholar] [CrossRef]
- Cope, K.H.; Utgaard, J.E.; Masters, J.M.; Feldmann, R.M. The fauna of the Clayton Formation (Paleocene, Danian) of southern Illinois: A case of K/P survivorship and Danian recovery. Bull. Mizunami Foss. Mus. 2005, 32, 97–108. [Google Scholar]
- Becker, M.; Chamberlain, J.A., Jr.; Wolf, G. Chondrichthyans from the Arkadelphia Formation (Upper Cretaceous: Upper Maastrichtian) of Hot Spring County, Arkansas. J. Paleontol. 2006, 80, 700–716. [Google Scholar] [CrossRef]
- Traverse, A. Paleopalynology, 2nd ed.; Topics in Geobiology Series Volume 28; Springer: Dordrecht, The Netherlands, 2007. [Google Scholar]
- Fensome, R.A.; Williams, G.L. The Lentin and Williams Index of Fossil Dinoflagellates; AASP Contributions Series 42; American Association of Stratigraphic Palynologists Foundation: Salt Lake City, UT, USA, 2004. [Google Scholar]
- Fensome, R.A.; MacRae, R.A.; Williams, G.L. DINOFLAJ2, Version 1; Data Series No. 1; American Association of Stratigraphic Palynologists: Reston, VA, USA, 2008. [Google Scholar]
- Lucas-Clark, J. Small peridinioid dinoflagellate cysts from the Paleocene of South Carolina, U.S.A. Palynology 2006, 30, 183–210. [Google Scholar]
- Lentin, J.K.; Williams, G.L. A Monograph of Fossil Peridinioid Dinoflagellate Cysts; Bedford Institute of Oceanography Report Series B-R-75-16; Bedford Institute of Oceanography: Dartmouth, NS, Canada, 1976. [Google Scholar]
- Stover, L.E.; Evitt, W.R. Analyses of Pre-Pleistocene Organic-Walled Dinoflagellates; Stanford University Publications, Geological Sciences; Stanford University: Stanford, CA, USA, 1978. [Google Scholar]
- Palynodata Inc.; White, J.M. Palynodata Datafile, 2006 version [CD-ROM], Open File 5793, Geological Survey of Canada: Ottawa, ON, Canada, 2008.
- Benson, D.G. Dinoflagellate taxonomy and biostratigraphy at the Cretaceous-Tertiary boundary, Round Bay, Maryland. Tulane Stud. Geol. Paleontol. 1976, 12, 169–233. [Google Scholar]
- Brinkhuis, H.; Leereveld, H. Dinoflagellate cysts from the Cretaceous/Tertiary boundary sequence of El Kef, northwest Tunisia. Rev. Palaeobot. Palynol. 1988, 56, 5–19. [Google Scholar] [CrossRef]
- Brinkhuis, H.; Zachariasse, W.J. Dinoflagellate cysts, sea level changes and planktonic foraminifers across the Cretaceous/Tertiary boundary at El Haria, northwest, Tunisia. Mar. Micropaleontol. 1988, 13, 153–191. [Google Scholar] [CrossRef]
- Brinkhuis, H.; Bujak, J.P.; Smit, J.; Versteegh, G.J.M.; Visscher, H. Dinoflagellate based sea surface temperature reconstructions across the Cretaceous-Tertiary boundary. Palaeogeogr. Palaeoclim. Palaeoecol. 1998, 141, 67–83. [Google Scholar] [CrossRef]
- Brinkhuis, H.; Sengers, S.; Sluijs, A.; Warnaar, J.; Williams, G.L. Latest Cretaceous earliest Oligocene and Quaternary dinoflagellate cysts from ODP Site 1172, East Tasman Plateau. Proc. Ocean Drill. Program Sci. Results 2003, 189, 1–36. [Google Scholar]
- Damassa, S.P. Carpatella cornuta Grigorovich 1969 (Dinophyceae): A member of the Aptiana-Ventriosum complex. Palynology 1988, 12, 167–177. [Google Scholar] [CrossRef]
- Damassa, S.P.; Goodman, D.K.; Kidson, E.J.; Williams, G.L. Correlation of Paleogene dinoflagellate assemblages to standard nannofossil zonation in North Atlantic DSDP sites. Rev. Palaeobot. Palynol. 1990, 65, 331–339. [Google Scholar] [CrossRef]
- Edwards, L.E.; Frederiksen, N.O.; Bybell, L.M.; Gibson, T.G.; Gohn, G.S.; Bukry, D.; Self-Trail, J.M.; Litwin, R.J. Overview of the Biostratigraphy and Paleoecology of Sediments from Five Course from Screven and Burke Counties, Georgia. In Geology and Paleontology of Five Cores from Screven and Burke Counties, Eastern Georgia; Edwards, L.E., Ed.; U.S. Geological Survey: Reston, VA, USA, 2001; pp. 1–19. [Google Scholar]
- Edwards, L.E.; Goodman, D.K.; Witmer, R.J. Lower Tertiary (Pamunkey Group) Dinoflagellate Biostratigraphy, Potomac River Area, Virginia and Maryland. In Cretaceous and Tertiary Stratigraphy, Paleontology, and Structure, Southwestern Maryland, and Northeastern Virginia: American Association of Stratigraphic Palynologists Field Trip Volume Guidebook; Frederiksen, N.O., Krafft, K., Eds.; American Association of Stratigraphic Palynologists: Reston, VA, USA, 1984; pp. 137–152. [Google Scholar]
- Firth, J.V. Dinoflagellate biostratigraphy of the Maastrichtian to Danian interval in the U.S. Geological Survey Albany Core, Georgia, U.S.A. Palynology 1987, 11, 199–216. [Google Scholar] [CrossRef]
- Firth, J.V. Dinoflagellate assemblages and sea-level fluctuations in the Maastrichtian of southwest Georgia. Rev. Palaeobot. Palynol. 1993, 79, 179–204. [Google Scholar] [CrossRef]
- Firth, J.V. Upper Middle Eocene to Oligocene dinoflagellate biostratigraphy and assemblage variations in Hole 913B, Greenland Sea. Proc. Ocean Drill. Program Sci. Results 1996, 151, 203–242. [Google Scholar]
- Gedl, P. Dinoflagellate Cyst Record of the Deep-Sea Cretaceous-Tertiary Boundary at Uzgru, Carpathian Mountains, Czech Republic. In The Palynology and Micropalaeontology of Boundaries; Special Publications Volume 230; The Geological Society of London: London, UK, 2004; pp. 257–273. [Google Scholar]
- Habib, D.; Drugg, W.S. Palynology of sites 603 and 605, Leg 93, Deep Sea Drilling Project. Initial Rep. Deep Sea Drill. Proj. 1987, 93, 751–775. [Google Scholar]
- Habib, D.; Olsson, R.K.; Liu, C.; Moshkovitz, S. High-resolution biostratigraphy of sea-level low, biotic extinction, and chaotic sedimentation at the Cretaceous-Tertiary boundary in Alabama, north of the Chicxulub crater. GSA Spec. Pap. 1996, 307, 243–252. [Google Scholar]
- Hansen, J.M. Dinoflagellate stratigraphy and echinoid distribution in Upper Maastrichtian and Danian deposits from Denmark. Bull. Geol. Soc. Den. 1977, 26, 1–26. [Google Scholar]
- Hansen, J.M. A new dinoflagellate zone at the Maastrichtian/Danian boundary in Denmark. Dan. Geol. Unders. Arbog 1979, 1979, 131–140. [Google Scholar]
- Heilmann-Clausen, C. Dinoflagellate Stratigraphy of the Uppermost Danian to Ypresian in the Viborg 1 Borehole, Central Jylland, Denmark; Series A; Volume 7, Danmarks Geologiske Undersøgelse: Copenhagen, Danmark, 1985. [Google Scholar]
- Hultberg, S.U.; Malmgren, B.A. Dinoflagellate and planktonic foraminiferal paleobathymetrical indices in the boreal uppermost Cretaceous. Micropaleontology 1985, 32, 316–323. [Google Scholar] [CrossRef]
- Hultberg, S.U. Danian dinoflagellate zonation, the C-T boundary and the stratigraphical position of the fish clay in southern Scandinavia. J. Micropalaeontol. 1986, 5, 37–47. [Google Scholar] [CrossRef]
- Koch, R.C.; Olsson, R.K. Dinoflagellate and planktonic foraminiferal biostratigraphy of the uppermost Cretaceous of New Jersey. J. Paleontol. 1977, 51, 480–491. [Google Scholar]
- Kurita, H.; McIntyre, D.J. Paleocene dinoflagellates from the Turtle Mountain Formation, southwestern Manitoba, Canada. Palynology 1995, 19, 119–136. [Google Scholar] [CrossRef]
- Landman, N.H.; Johnson, R.O.; Edwards, L.E. Cephalopods from the Cretaceous/Tertiary Boundary Interval on the Atlantic Coastal Plain, with a Description of the Highest Ammonite Zones in North America. Part 1. Maryland and North Carolina; American Museum Novitates No. 3454; American Museum of Natural History: New York, NY, USA, 2004. [Google Scholar]
- Landman, N.H.; Johnson, R.O.; Edwards, L.E. Cephalopods from the Cretaceous/Tertiary Boundary Interval on the Atlantic Coastal Plain, with a Description of the Highest Ammonite Zones in North America. Part 2. Northeastern Monmouth County, New Jersey; Bulletin of The American Museum of Natural History No. 287; American Museum of Natural History: New York, NY, USA, 2004. [Google Scholar]
- Landman, N.H.; Johnson, R.O.; Garb, M.; Edwards, L.E.; Kyte, F.T. Cephalopods from the Cretaceous/Tertiary Boundary Interval on the Atlantic Coastal Plain, with a Description of the Highest Ammonite Zones in North America. Part 3. Manasquan River Basin, Monmouth County, New Jersey; Bulletin of The American Museum of Natural History No. 303; American Museum of Natural History: New York, NY, USA, 2007. [Google Scholar]
- Morgenroth, P. Zur Kenntnis der Dinoflagellaten und Hystrichosphaeridien des Danien. Geologisches Jarhbuch 1968, 86, 533–578. (In German) [Google Scholar]
- Mudge, D.C.; Bujak, J.P. Palaeocene biostratigraphy and sequence stratigraphy of the UK central North Sea. Mar. Petrol. Geol. 1996, 13, 295–312. [Google Scholar] [CrossRef]
- Nøhr-Hansen, H.; Dam, G. Palynology and sedimentology across a new marine Cretaceous-Tertiary boundary section on Nuussuaq, West Greenland. Geology 1997, 25, 851–854. [Google Scholar] [CrossRef]
- Nøhr-Hansen, H.; Dam, G. Emendation of Trithyrodinium evittii Drug 1967 and Trithyrodinium fragile (Davey 1969) an artificial split of one dinoflagellate cyst species-Stratigraphic and paleaoenvironmental importance. Grana 1999, 138, 125–133. [Google Scholar]
- Nøhr-Hansen, H.; Heilmann-Clausen, C. Cerodinium kangiliense n. sp. and Senegalinium iterlaaense n. sp.—Two new stratigraphically important Paleocene species from West Greenland and Denmark. Neues Jahrb. Geol. Paläontol. Abh. 2001, 219, 153–170. [Google Scholar]
- Powell, A.J. A Stratigraphic Index of Dinoflagellate Cysts; British Micropaleontological Society Publication Series; Springer: Berlin, Germany, 1992. [Google Scholar]
- Pross, J.; Schmiedl, G. Early Oligocene dinoflagellate cysts from the Upper Rhine Graben (SW Germany): Paleoenvironmental and paleoclimatic implications. Mar. Micropaleontol. 2002, 45, 1–24. [Google Scholar] [CrossRef]
- Prowell, D.C.; Edwards, L.E.; Frederiksen, N.O. The Ellenton Formation in South Carolina—A Revised Age Designation from Cretaceous to Paleocene; U.S. Geological Survey Bulletin 1605-A; U.S. Geological Survey: Reston, VA, USA, 1985; pp. A63–A69. [Google Scholar]
- Schiøler, P.; Wilson, G.J. Maastrichtian dinoflagellate zonation in the Dan Field, Danish North Sea. Rev. Palaeobot. Palynol. 1993, 78, 321–351. [Google Scholar] [CrossRef]
- Stanley, E.A. Upper Cretaceous and Paleocene plant microfossils and Paleocene dinoflagellates and hystrichosphaerids from northwestern South Dakota. Bull. Am. Paleontol. 1965, 49, 179–384. [Google Scholar]
- Stover, L.E.; Brinkhuis, H.; Damassa, S.P.; de Verteuil, L.; Helby, R.J.; Monteil, E.; Partridge, A.D.; Powell, A.J.; Riding, J.B.; Smelror, M.; Williams, G.L. Mesozoic-Tertiary Dinoflagellates, Acritarchs and Prasinophytes. In Palynology: Principles and Applications; Jansonius, J., McGregor, D.C., Eds.; American Association of Stratigraphic Palynologists Foundation: College Station, TX, USA, 1996; Volume 2, pp. 641–750. [Google Scholar]
- Williams, G.L.; Brinkhuis, H.; Pearce, M.A.; Fensome, R.A.; Weegink, J.W. Southern ocean and global dinoflagellate cyst events compared: Index events for the Late Cretaceous-Neogene. Proc. Ocean Drill. Program Sci. Results 2004, 189, 1–98. [Google Scholar]
- Willumsen, P.S. Palynodinium minus sp. nov., a new dinoflagellate cyst from the Cretaceous Paleogene transition in New Zealand; Its significance and palaeoecology. Cretac. Res. 2006, 27, 954–963. [Google Scholar] [CrossRef]
- Wilson, G.J. Upper Campanian and Maastrichtian Dinoflagellate Cysts from the Maastricht Region and Denmark. Ph.D. Thesis, University of Nottingham, Nottingham, UK, 1974. [Google Scholar]
- Habib, D.; Miller, J.A. Dinoflagellate species and organic facies evidence of marine transgression and regression in the Atlantic Coastal Plain. Palaeogeogr. Palaeoclim. Palaeoecol. 1989, 74, 23–47. [Google Scholar] [CrossRef]
- Moullade, M.; Watkins, D.K.; Oboh-Ikuenobe, F.E.; Bellier, J.-P.; Masure, E.; Holbourn, A.E.L.; Erbacher, J.; Kuhnt, W.; Pletsch, T.; Kaminski, T.A.; et al. Mesozoic biostratigraphic, paleaoenvironmental, and paleobiogeographic synthesis, equatorial Atlantic. Proc. Ocean Drill. Program Sci. Results 1998, 159, 481–490. [Google Scholar]
- Brinkhuis, H.; Klinkenberg, E.; Williams, G.L.; Fensome, R.A. Two unusual new dinoflagellate cyst genera from the Bunde Borehole, Maastrichtian type area, southern Netherlands. Rev. Palaeobot. Palynol. 2000, 93–110. [Google Scholar]
- Crouch, E.M.; Willumsen, P.S.; Kulhanek, D.; Gibbs, S. A revised Paleocene (Tuerian) dinoflagellate cyst zonation from eastern New Zealand. Rev. Palaeobot. Palynol. 2014, in press. [Google Scholar]
- Renne, P.R.; Deino, A.L.; Hilgen, F.J.; Kuiper, K.F.; Mark, D.F.; Mitchell, W.S., III; Morgan, L.E.; Mundil, R.; Smit, J. Time scales of critical events around the Cretaceous-Paleogene boundary. Science 2013, 339, 684–687. [Google Scholar] [CrossRef]
- Beckvar, N.; Kidwell, S.M. Hiatal shell concentrations, sequence analysis and sea level history of a Pleistocene coastal alluvial fan, Punta Chueca, Sonora. Lethaia 1988, 21, 257–270. [Google Scholar] [CrossRef]
- Banerjee, I.; Kidwell, S.M. Significance of molluscan shell beds in sequence stratigraphy: An example from the Lower Cretaceous Mannville Group of Canada. Sedimentology 1991, 38, 913–934. [Google Scholar] [CrossRef]
- Kidwell, S.M. Palaeobiological and sedimentological implications of fossil concentrations. Nature 1985, 318, 457–460. [Google Scholar] [CrossRef]
- Kidwell, S.M. Patterns of Time Averaging in the Shallow Marine Fossil Record. In Taphonomic Approaches to Time Resolution in Fossil Assemblages; Kidwell, S.M., Behrensmeyer, A.K., Eds.; The Paleontological Society: Knoxville, TN, USA, 1993; pp. 275–300. [Google Scholar]
- Kidwell, S.M. Condensed Deposits in Siliciclastic Sequences: Expected and Observed Features. In Cycles and Events in Stratigraphy; Einsele, G., Ricken, W., Seilacher, A., Eds.; Springer-Verlag: Berlin, Germany, 1991; pp. 683–695. [Google Scholar]
- Kidwell, S.M. The Stratigraphy of Shell Concentrations. In Taphonomy: Releasing the Data Locked in the Fossil Record; Allison, P.A., Briggs, E.G., Eds.; Plenum Press: New York, NY, USA, 1991; Volume 9, pp. 211–279. [Google Scholar]
- Larina, E.; Landman, N.H.; Garb, M.P.; Meyers, C.E.; Rovelli, R. Upper Maastrichtian Ammonite Biostratigraphy of the Northeastern Gulf Coastal Plain. In Proceedings of 125th Anniversary Annual Meeting and Exposition of Geological Society of America, Denver, CO, USA, 27–30 October 2013. Paper No. 380-8.
- Smith, C.C. The Cretaceous-Tertiary boundary at Moscow Landing, west-central Alabama. GCAGS Trans. 1997, 47, 533–540. [Google Scholar]
- Hart, M.B.; Harries, P.J.; Cárdenas, A.L. The Cretaceous/Paleogene boundary events in the Gulf Coast: Comparisons between Alabama and Texas. GCAGS Trans. 2013, 63, 235–255. [Google Scholar]
- Boas, C.; Garb, M.P.; Landman, N.H.; Rovelli, R.; Larina, E. The Significance of a Fossiliferous Spherule Bed at the K/Pg Boundary in Northern Mississippi. In Proceedings of 62nd Annual Meeting of Geological Society of America Southeastern Section, San Juan, Puerto Rico, 20–21 March 2013.
- Boas, C.; Garb, M.P.; Rovelli, R.; Larina, E.; Myers, C.E.; Naujokaityte, J.; Landman, N.H.; Phillips, G.E. New K/Pg Localities along the Eastern Gulf Coastal Plain: More Evidence of Impact and Tsunamis. In Proceedings of 125th Anniversary Annual Meeting and Exposition of Geological Society of America, Denver, CO, USA, 27–30 October 2013.
- Landman, N.H.; Garb, M.P.; Rovelli, R.; Ebel, D.S.; Edwards, L.E. Short-term survival of ammonites in New Jersey after the End-Cretaceous Bolide impact. Acta Palaeontol. Pol. 2012, 57, 703–715. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Dastas, N.R.; Chamberlain, J.A., Jr.; Garb, M.P. Cretaceous-Paleogene Dinoflagellate Biostratigraphy and the Age of the Clayton Formation, Southeastern Missouri, USA. Geosciences 2014, 4, 1-29. https://doi.org/10.3390/geosciences4010001
Dastas NR, Chamberlain JA Jr., Garb MP. Cretaceous-Paleogene Dinoflagellate Biostratigraphy and the Age of the Clayton Formation, Southeastern Missouri, USA. Geosciences. 2014; 4(1):1-29. https://doi.org/10.3390/geosciences4010001
Chicago/Turabian StyleDastas, Natalie R., John A. Chamberlain, Jr., and Matthew P. Garb. 2014. "Cretaceous-Paleogene Dinoflagellate Biostratigraphy and the Age of the Clayton Formation, Southeastern Missouri, USA" Geosciences 4, no. 1: 1-29. https://doi.org/10.3390/geosciences4010001