Evolution of Endemic Species, Ecological Interactions and Geographical Changes in an Insular Environment: A Case Study of Quaternary Mammals of Sicily (Italy, EU)

Abstract
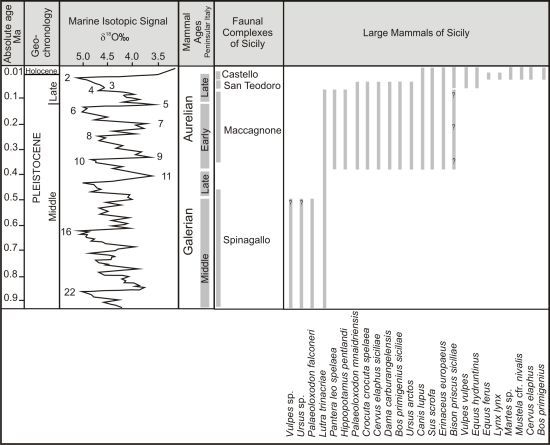
:
1. Introduction
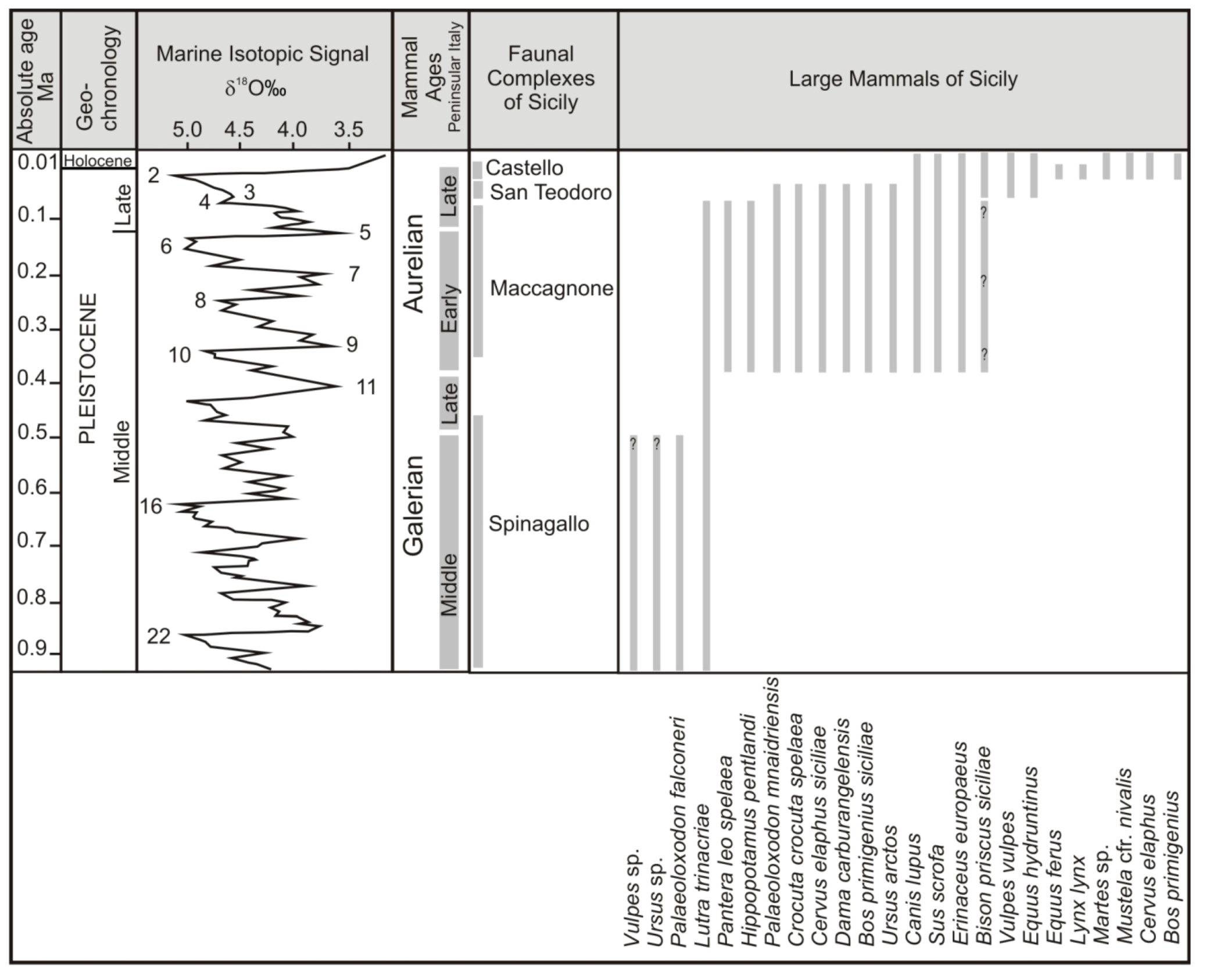
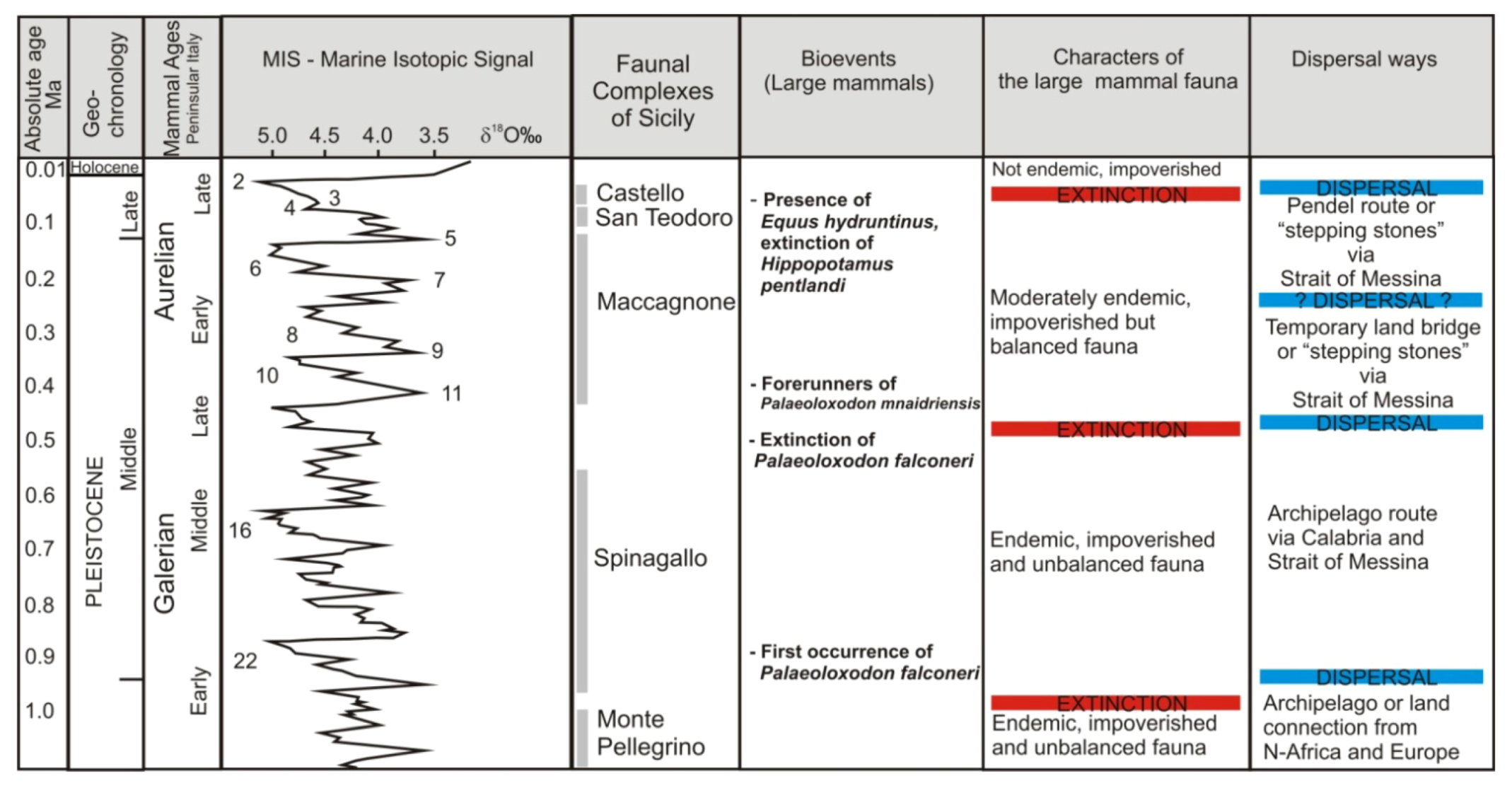
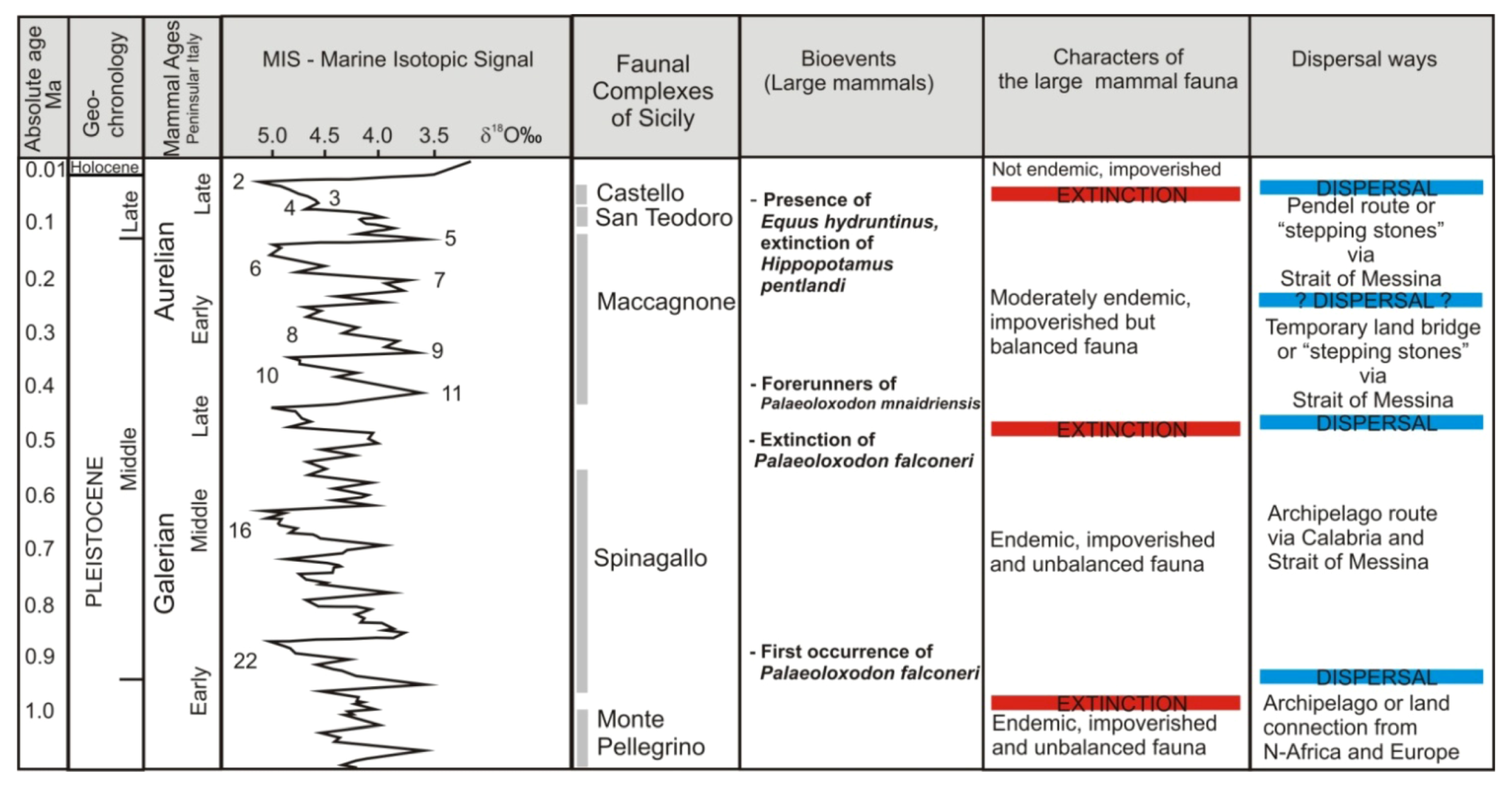
2. Review of Biochronology
- -
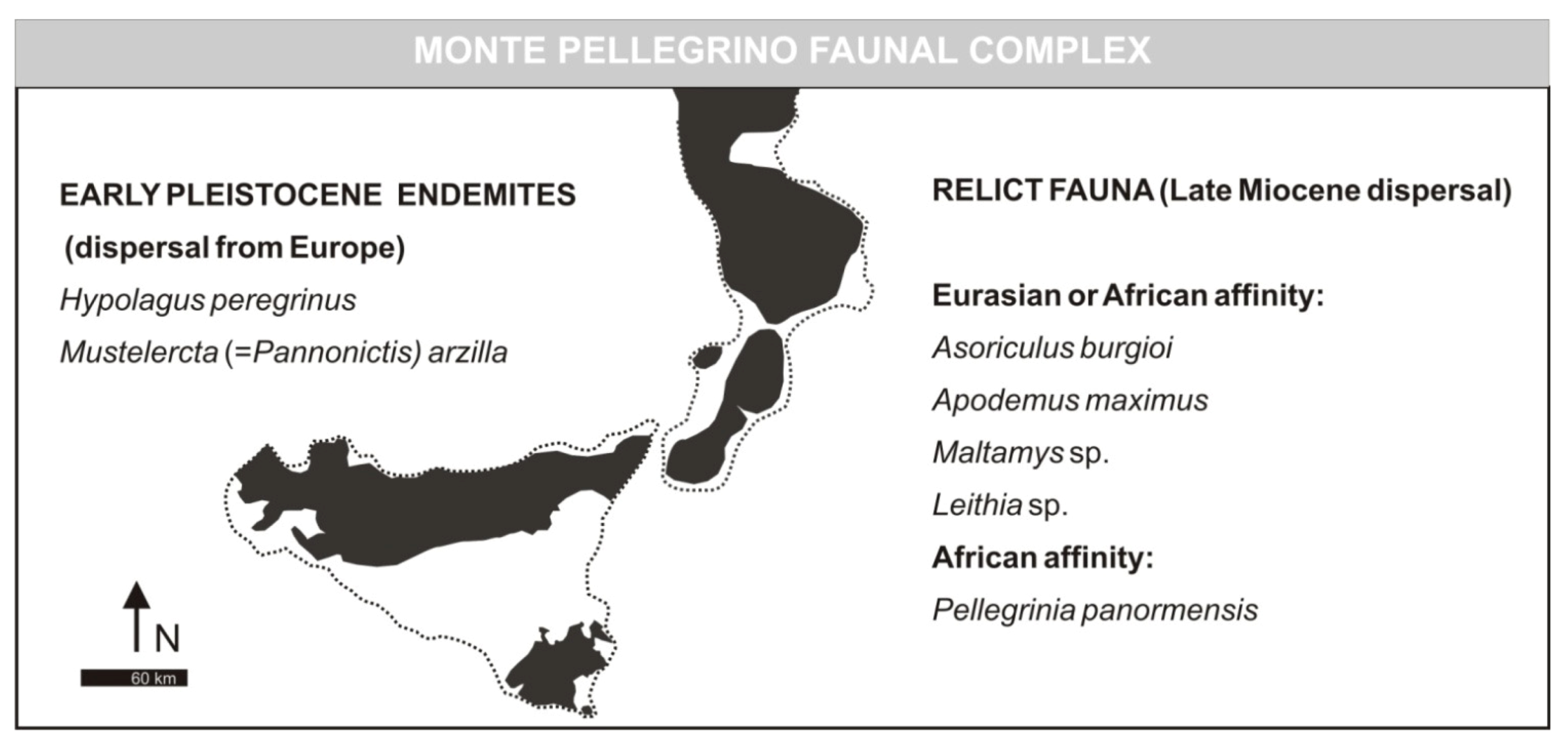
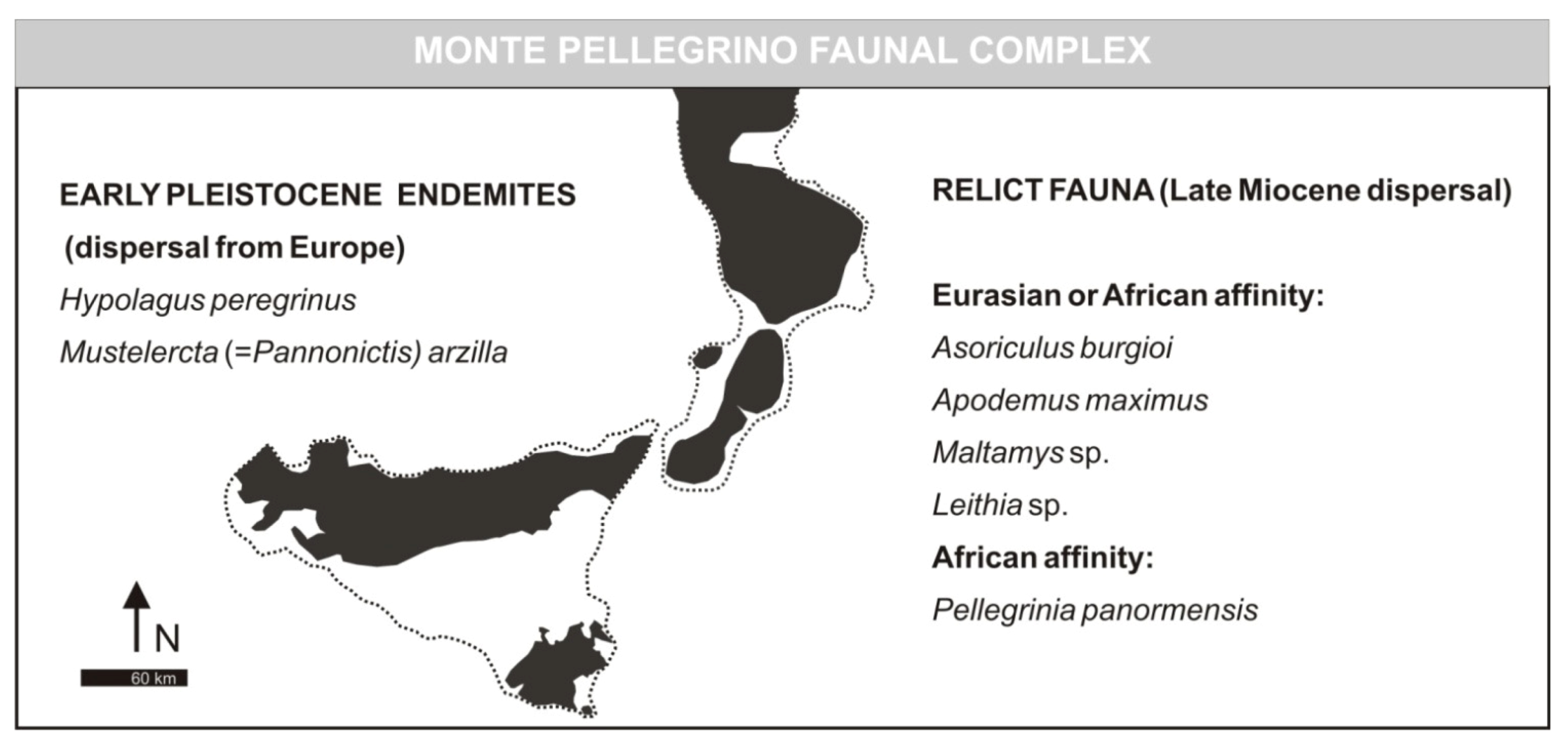
- Monte Pellegrino Faunal ComplexMammal association: Asoriculus burgioi (E), Mustelercta (=Pannonictis) arzilla (E), Apodemus maximus (E), Leithia sp. (E), Maltamys sp. (E), Pellegrinia panormensis (E), Hypolagus peregrinus (E). The Monte Pellegrino FC is based on a single faunal assemblage coming from a karst cavity with a problematic depositional setting.
- -
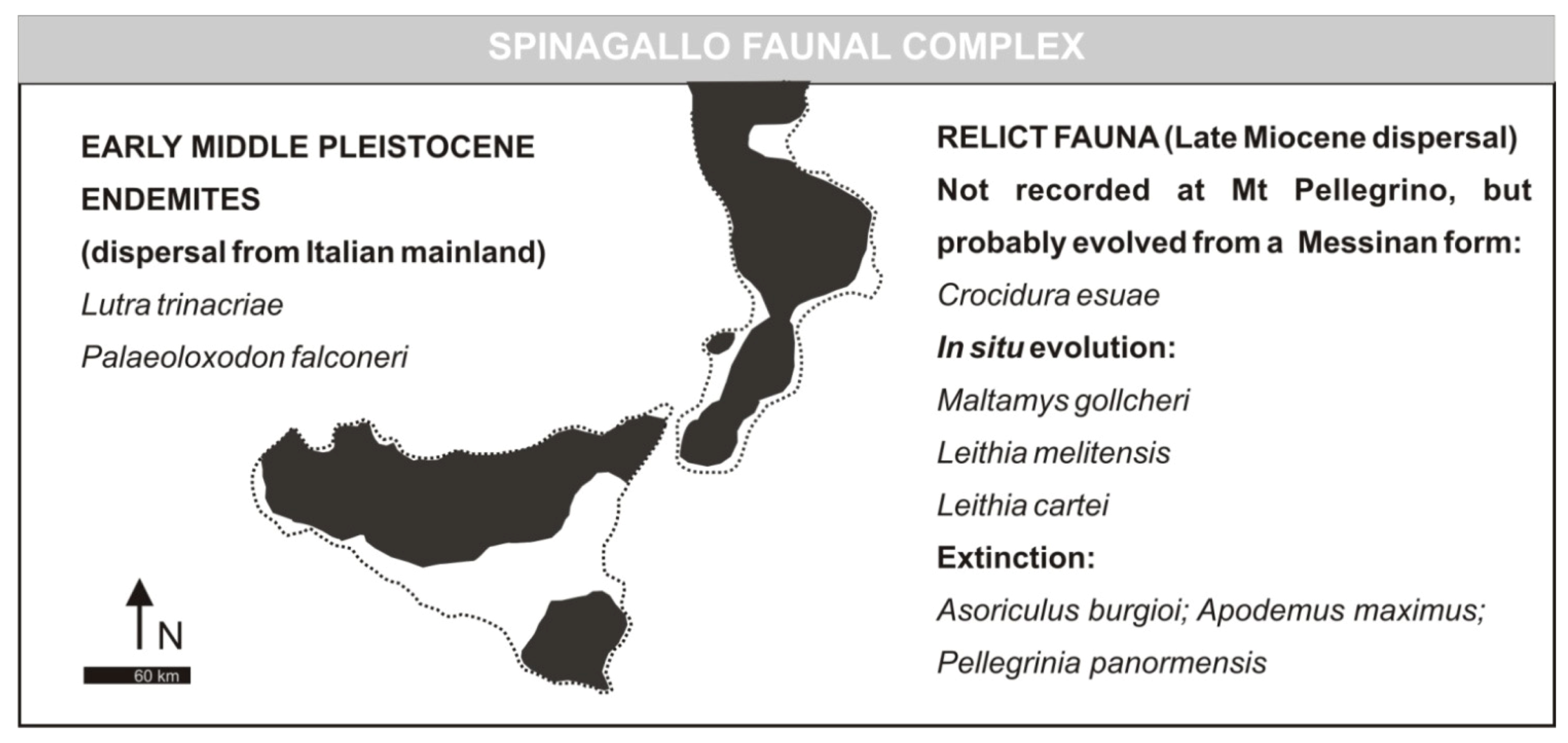
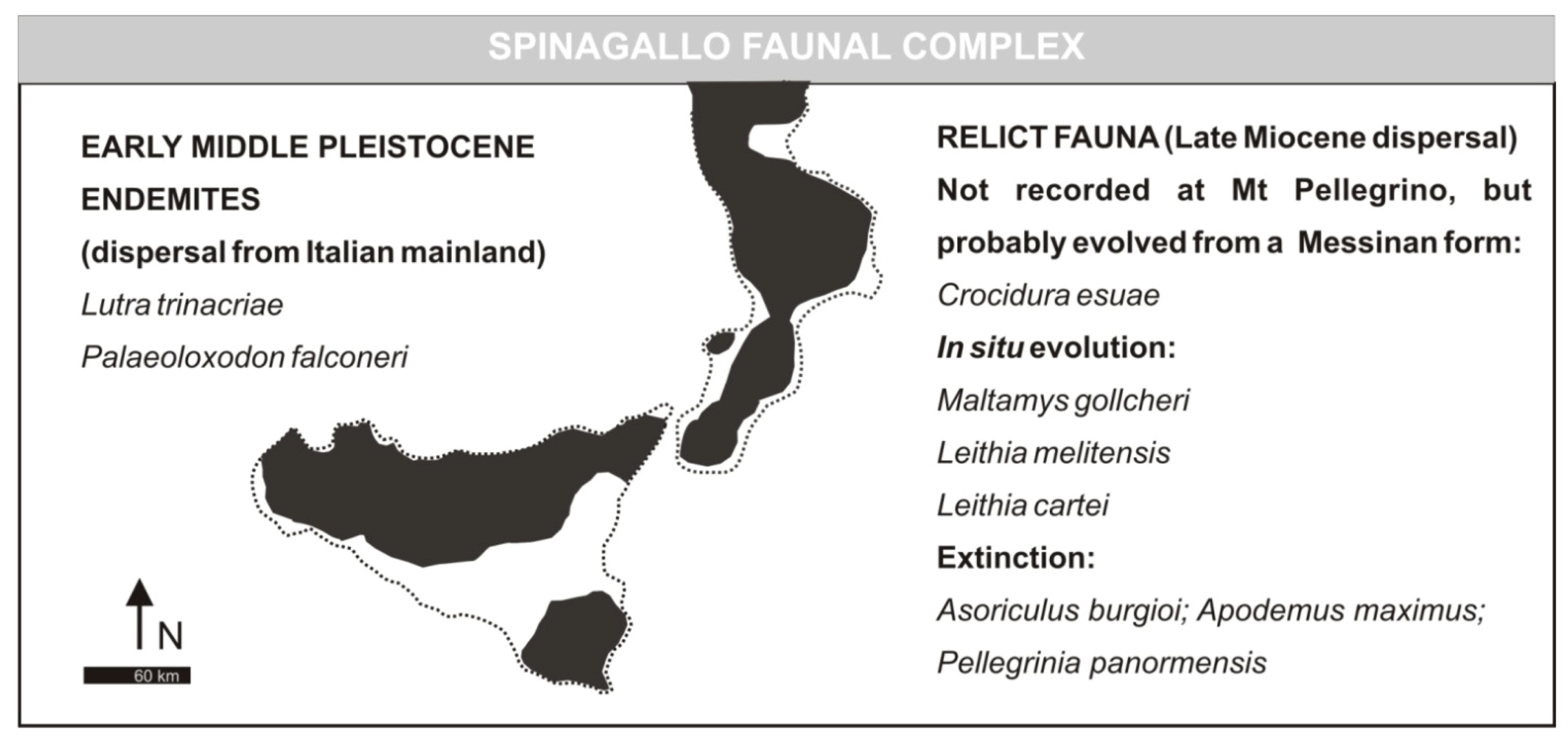
- Elephas falconeri Faunal ComplexMammal association: Crocidura esuae (E), Lutra trinacriae (E), Vulpes sp. (dubitative), Ursus sp. (“small bear,” dubitative), Elephas falconeri [recte Palaeoloxodon falconeri] (E), Leithia cartei (E), Leithia melitensis (E), Maltamys gollcheri (E), undetermined bats. The mammal fauna is associated with diversified herpetofauna and avifauna.Age: early Middle Pleistocene, probably MIS 22–11; Middle and Late Galerian of the biochronological chart of Italy.
- -
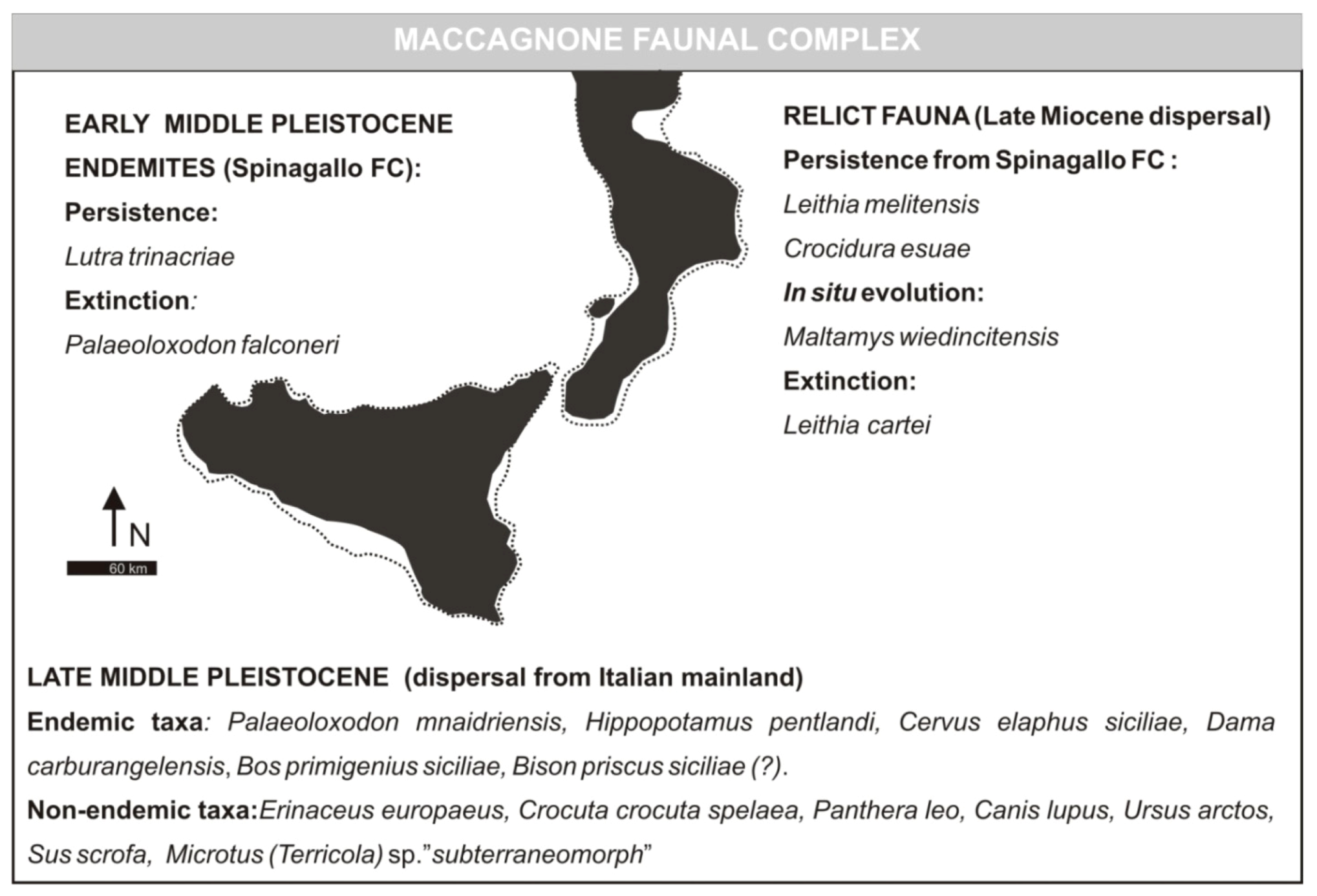
- Elephas mnaidriensis Faunal ComplexMammal association: Crocidura esuae (E), Erinaceus europaeus (E), Lutra trinacriae (E), Crocuta crocuta spelaea, Panthera leo, Canis lupus, Ursus arctos, Elephas mnaidriensis [recte Palaeoloxodon mnaidriensis] (E), Sus scrofa, Hippopotamus pentlandi (E), Cervus elaphus siciliae (E), Dama carburangelensis (E), Bos primigenius siciliae (E), Bison priscus siciliae (E), Leithia melitensis (E), Maltamys wiedincitensis (E), Microtus (Terricola) sp. In the opinion of Petruso et al. [11], Bison priscus siciliae is erroneously attributed to this faunal complex and must be considered present only in the “Grotta San Teodoro-Pianetti” FC.The mammal fauna is associated with diversified herpetofauna and avifauna.Age: late Middle Pleistocene, probably MIS 10–4; Aurelian Mammal Age of the biochronological chart of Italy.
- -
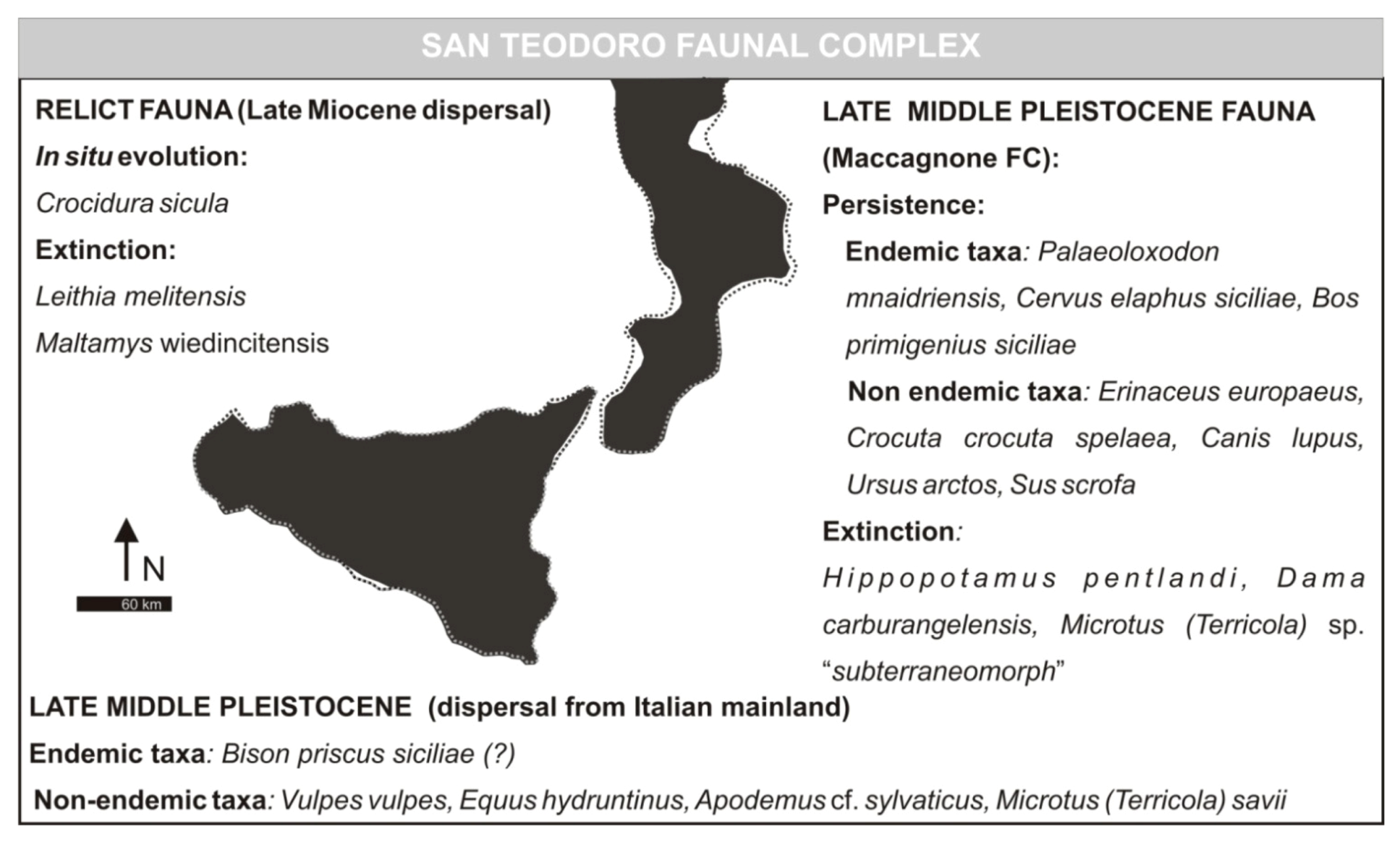
- Grotta San Teodoro—Pianetti Faunal ComplexMammal association: Erinaceus europaeus, Crocidura cf. sicula (E), Crocuta crocuta spelaea, Canis lupus, Vulpes vulpes, Ursus arctos, Elephas mnaidriensis [recte Palaeoloxodon mnaidriensis] (E), Sus scrofa, Cervus elaphus siciliae (E), Bos primigenius siciliae (E), Bison priscus siciliae (E), Equus hydruntinus, Apodemus cf. A. sylvaticus, Microtus (Terricola) savii (E).Mammal fauna is associated with herpetofauna and avifauna.Age: Late Pleistocene; Grotta San Teodoro: 32,000 ± 4000 yr BP (230Th/234U dating on a concretion, Bonfiglio et al. [16]; late Aurelian (Last Glacial cycle, more precisely) according to the biochronology of peninsular Italy.
- -
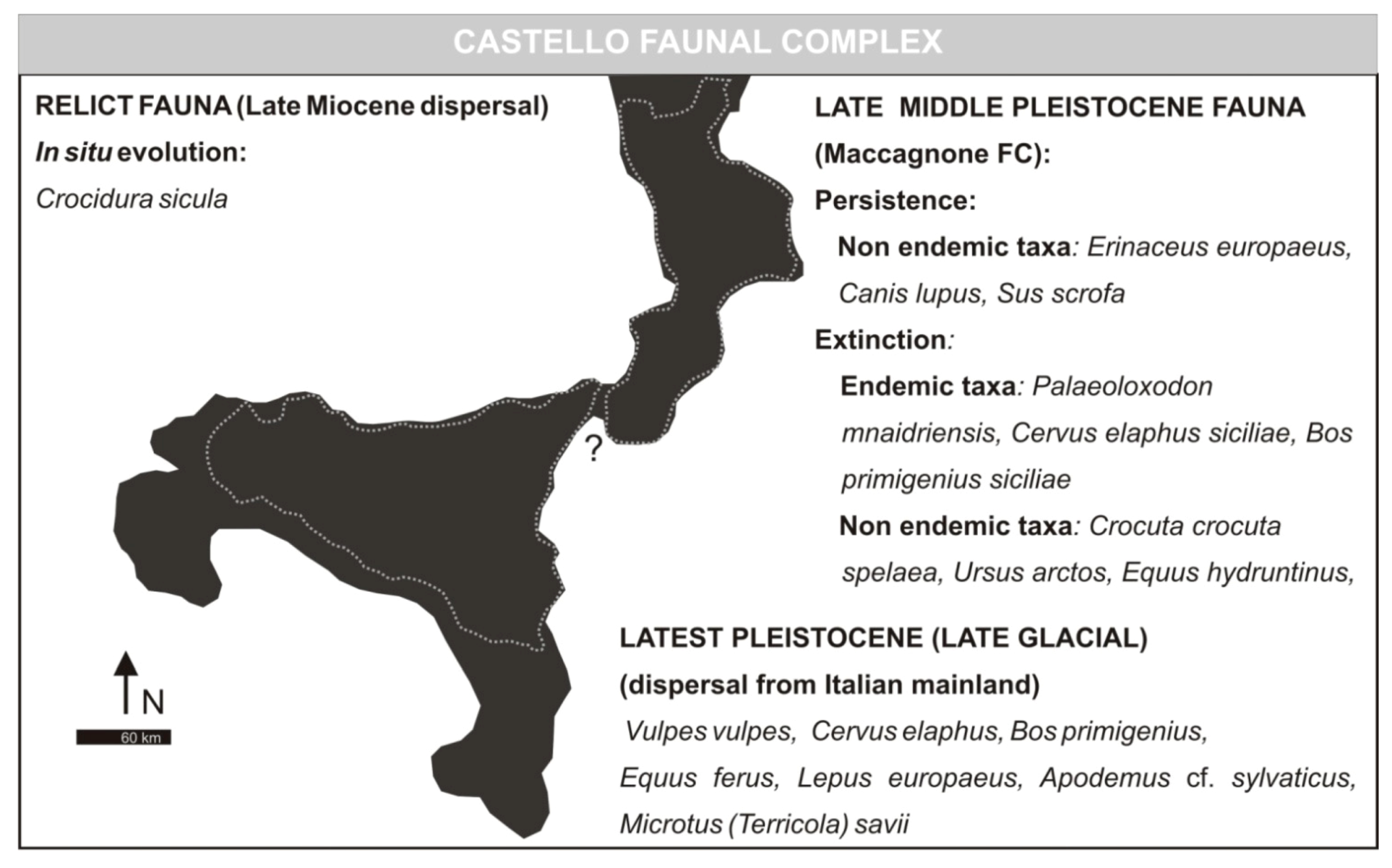
- Castello Faunal ComplexMammal association: Erinaceus europaeus, Crocidura cf. C. sicula (E), Canis lupus, Vulpes vulpes, Equus hydruntinus, Sus scrofa, Cervus elaphus, Bos primigenius, Equus ferus, Microtus (Terricola) savii, Apodemus cf. sylvaticus, Lepus europaeus.Age: late MIS 3—early MIS 2; late Aurelian (late Glacial) Mammal Age of the biochronological chart of Italy.
- -
- Monte Pellegrino Stage (fauna characterized by the presence of Pellegrinia panormensis associated with Leithia sp., a murid, a leporid, and a mustelid); type site: Monte Pellegrino (Palermo); Age: Ruscinian and/or Villafranchian and/or Galerian.
- -
- Capo Tindari Stage: fauna characterized by the presence of a large sized Hippopotamus sp. associated with Megaceros messinae (probably attributable to Dama carburangelensis) and Leithia; type site: Capo Tindari (Messina); Age: Galerian or Flaminian or Pariolin.
- -
- Maccagnone Stage (fauna characterized by the presence of Elephas mnaidriensis [recte Palaeoloxodon mnaidriensis] associated with Cervus siciliae [recte Cervus elaphus siciliae], Leithia melitensis, Leithia cartei), divided into two phases by the presence or absence of Hippopotamus pentlandi:(a) Hippopotamus pentlandi is present; type site: Maccagnone Cave (Palermo); Age: Pariolin, Rianin, Maspinian.(b)Hippopotamus pentlandi is absent; type site: Carburangeli Cave (Palermo); Age: end of Maspinian, Pontinian.
- -
- Spinagallo Stage (fauna characterized by the presence of Elephas falconeri [recte Palaeoloxodon falconeri] associated with Leithia melitensis and Leithia cartei); type site: Spinagallo cave (Syracuse); Age: Middle Pontinian.
- -
- Castello Stage (fauna characterized by the presence of Equus hydruntinus associated with Cervus elaphus and Homo sapiens; elephants are absent); type site: Castello cave (Palermo); Age: Upper Pontinain.
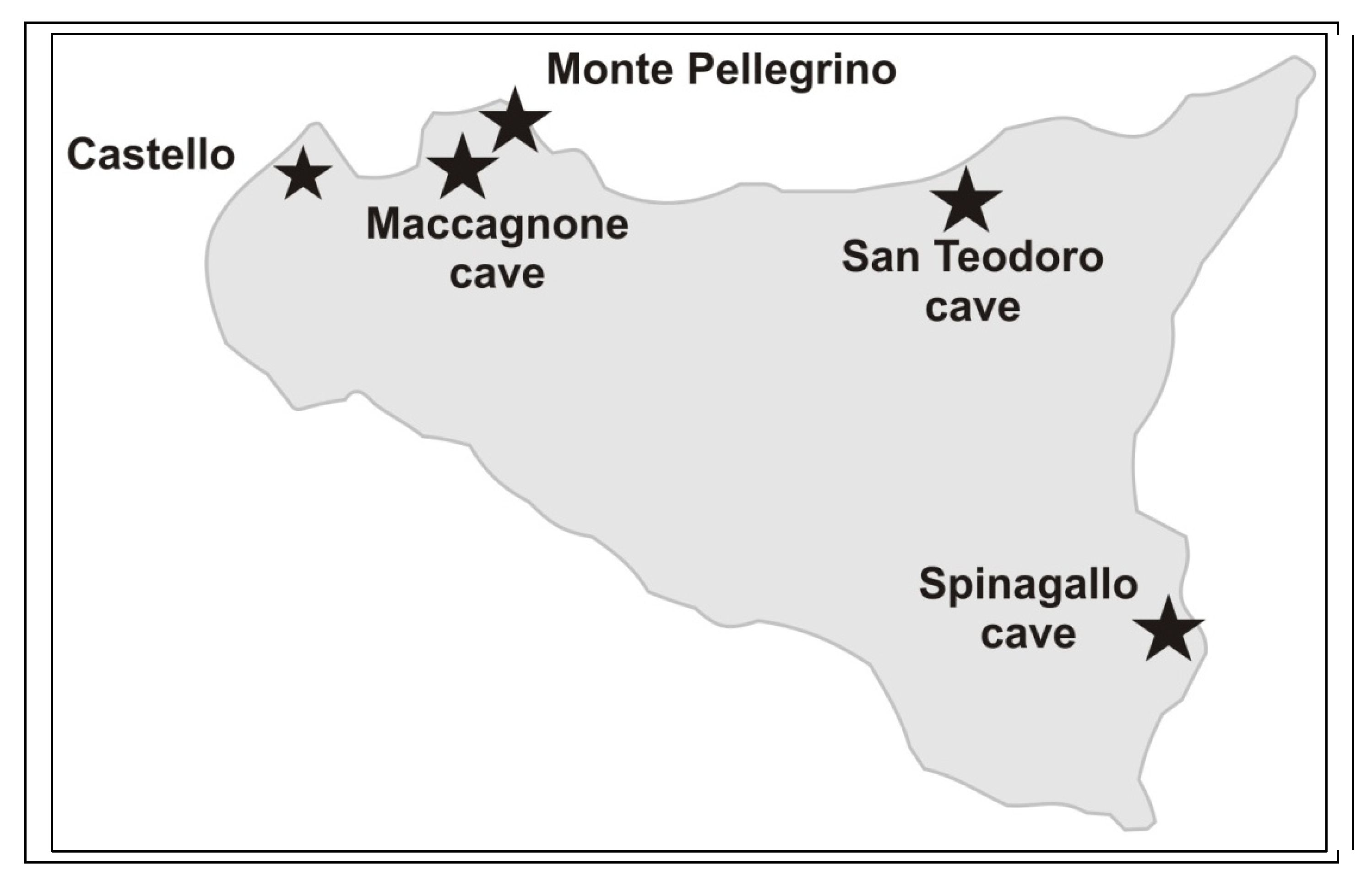
- -
- Monte Pellegrino Stage (fauna characterized by the presence of Pellegrinia panormensis). Faunal associations: Apodemus maximus, Maltamys cf. gollcheri, Pellegrinia panormensis, Hypolagus sp. [recte Hypolagus peregrines], Soricidae gen. et sp. indet., Mustelercta arzilla [recte Mustelercta (= Pannonictis) arzilla]
- -
- Spinagallo Stage (fauna characterized by the presence of Elephas falconeri [recte Palaeoloxodon falconeri]). Faunal association: Leithia melitensis, Leithia cartei, Crocidura esuae, Lutra trinacriae, Elephas falconeri [recte Palaeoloxodon falconeri]. Age: 500 ka by absolute dating [23]; Mindel-Riss.
- -
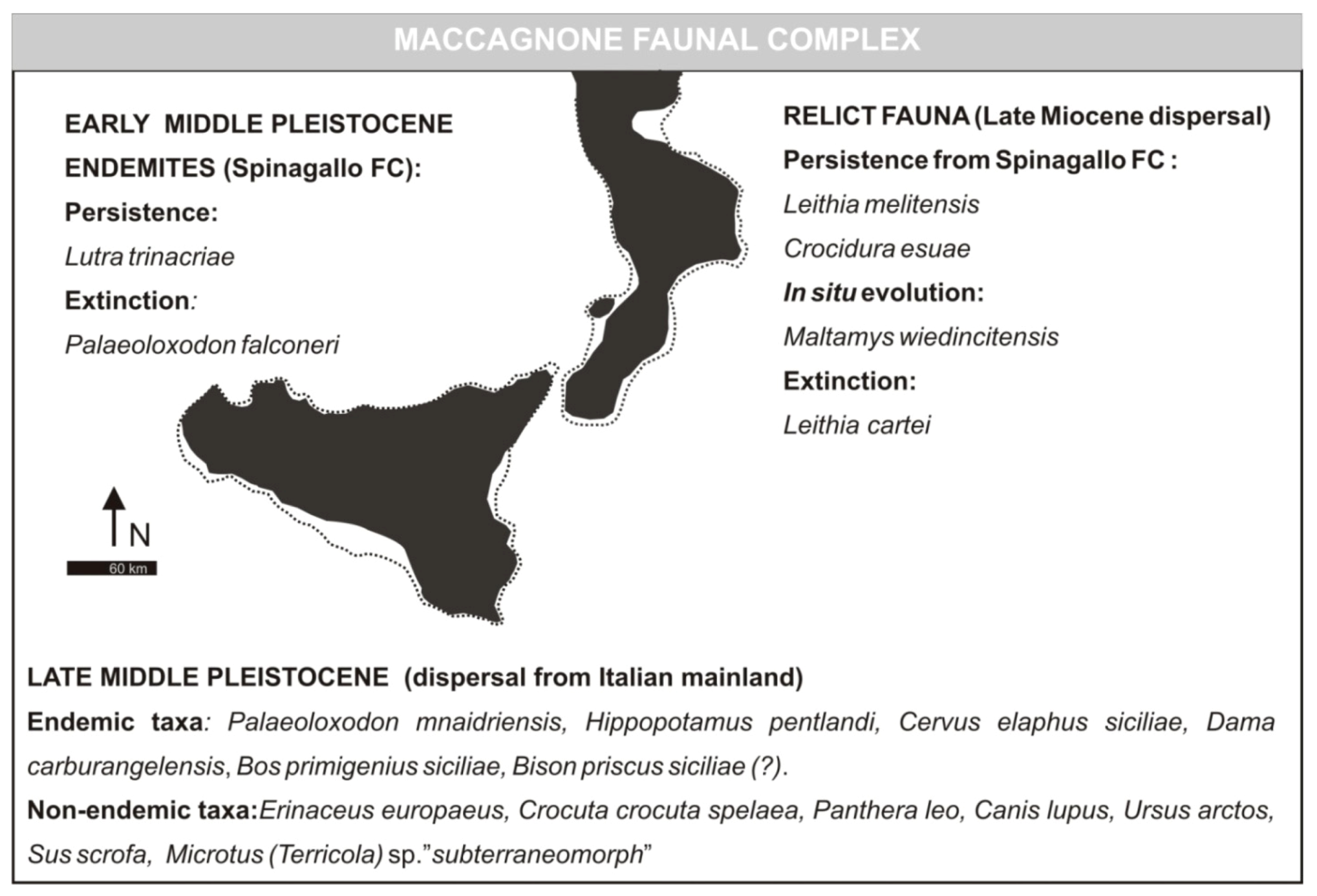
- Maccagnone Stage (fauna characterized by the presence of Elephas mnaidriensis [recte Palaeoloxodon mnaidriensis]). Faunal association: Leithia melitensis, Maltamys wiedinticitensis, Crocidura esuae, Crocuta crocuta spelaea, Panthera leo spelaea, Canis lupus, Ursus arctos, Elephas antiquus [recte Palaeoloxodon antiques], Elephas mnaidriensis [recte Palaeoloxodon mnaidriensis], Elephas sp., Sus scrofa, Hippopotamus pentlandi, Hippopotamus sp., Cervus elaphus siciliae, Dama dama [recte Dama carburangelensis], Megaceros (s.l.) carburangelensis [recte Dama carburangelensis], Bos primigenius siciliae, Bison priscus siciliae. Age: 200 ka by absolute dating [23]; attributable to MIS 5.
- -
- Castello Stage (fauna characterized by the presence of Equus hydruntinus). Faunal association: Hystrix cristata, Pytimis savii [recte Microtus (Terricola) savii], Lepus europaeus, Erinaceus europaeus, Crocidura sp., Lynx lynx, Felis silvestris, Canis lupus, Vulpes vulpes, Equus caballus [recte Equus ferus], Equus hydruntinus, Sus scrofa, Cervus elaphus elaphus, Bos primigenius primigenius, Homo sapiens sapiens. Age: some millennia before Holocene (10 ka), Upper Paleolithic.
3. Paleobiogeography, Endemism and Ecological Interactions
- -
- -
- denominations by species names are unusual; moreover, in this case, scientific names are obsolete, being Elephas falconeri and Elephas mnaidriensis currently named Palaeoloxodon falconeri and Palaeoloxodon mnaidriensis, respectively [25];
- -
- -
- the names indicated by Burgio [19] are maintained in the other FCs: Monte Pellegrino FC and Castello FC;
- -
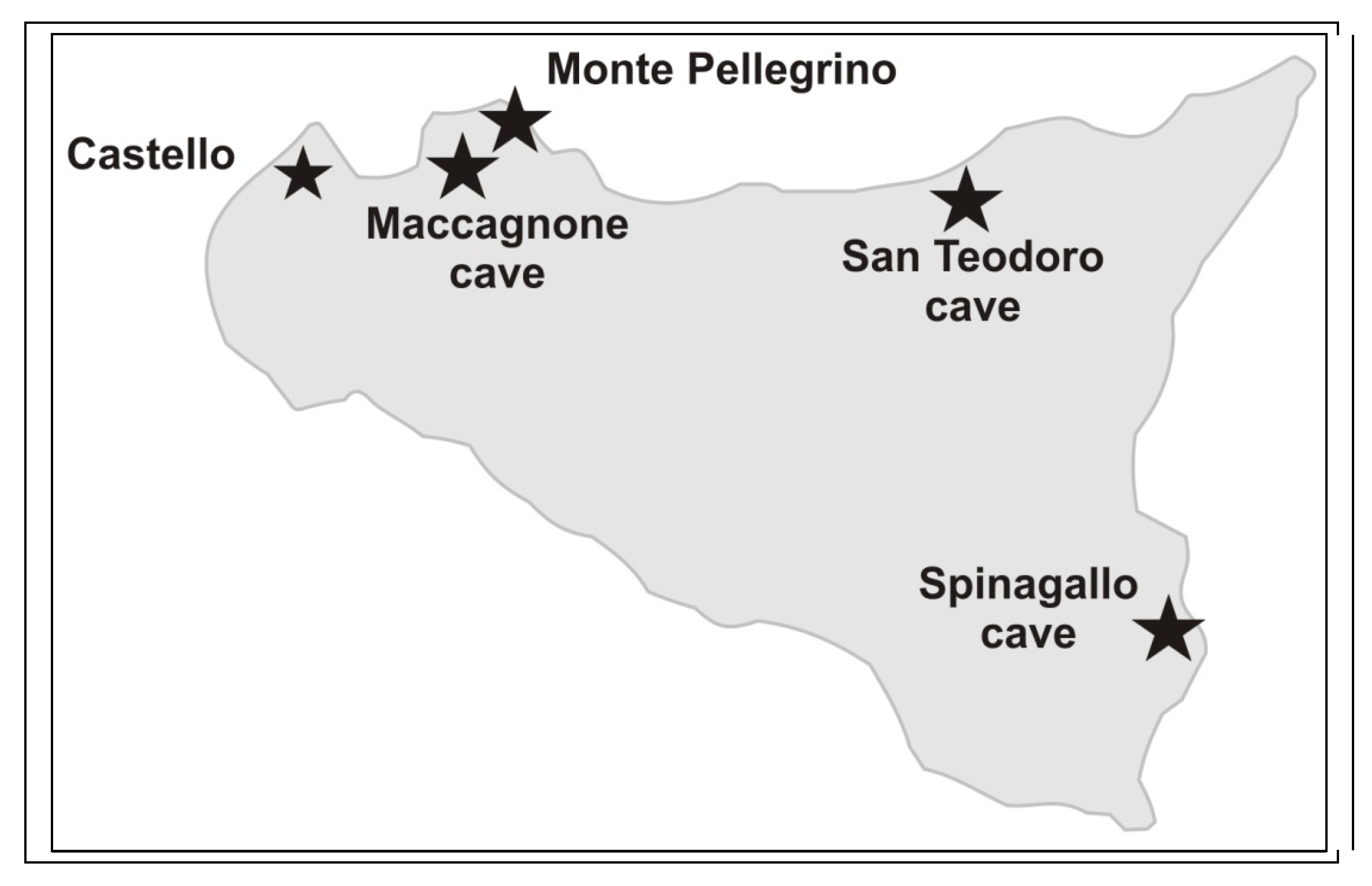
- Spinagallo and Maccagnone caves are particularly significant in the fossil record of mammals of Sicily.
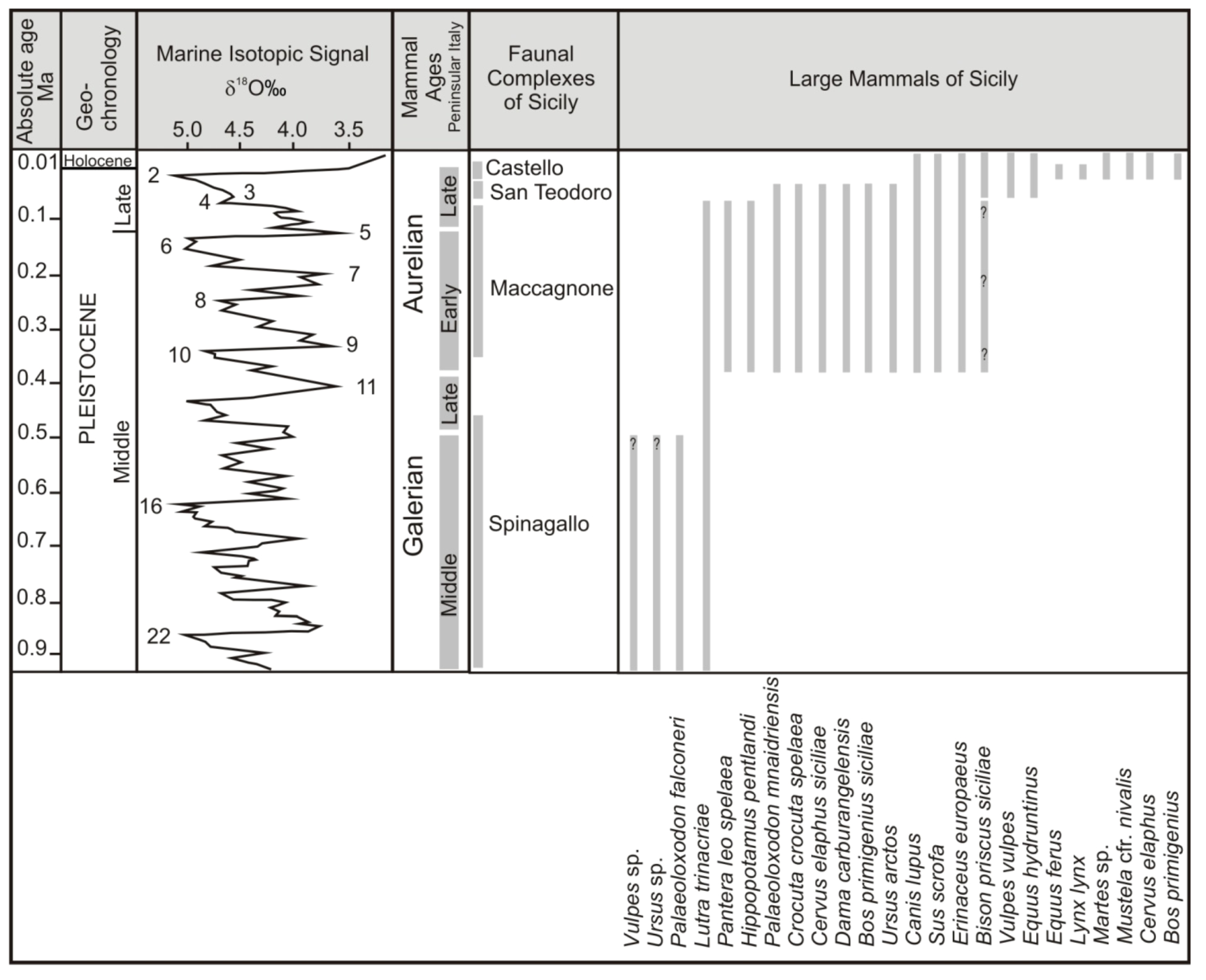
3.1. Monte Pellegrino Faunal Complex
3.2. Spinagallo Faunal Complex (Formerly Elephas falconeri FC)
3.3. Maccagnone Faunal Complex (Formerly Elephas mnaidriensis FC)

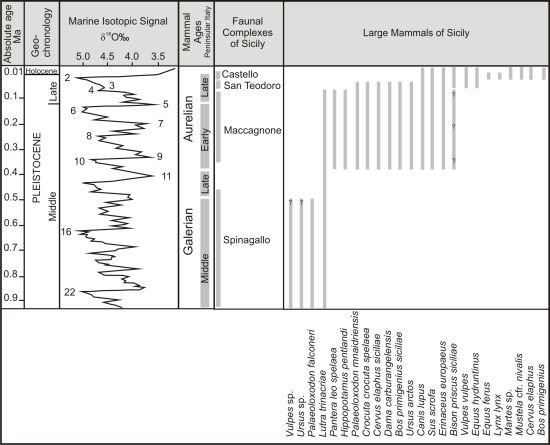{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Large mammal | Habitat | Feeding behavior | Body mass | Faunal complexes | |||
|---|---|---|---|---|---|---|---|
| Spinagallo | Maccagnone | S. Teodoro | Castello | ||||
| Palaeoloxodon falconeri | O-Wc | Br | V | X | |||
| Lutra trinacriae | F | sP | II | X | X | ||
| Panthera leo spelaea | O | SP | V | X | |||
| Hippopotamus pentlandi | O-F | Gr | VII | X | |||
| Dama carburangelensis | O-Wc | MF | IV | X | |||
| Crocuta crocuta spelaea | O-Wc | Sc-SP | III/IV | X | X | ||
| Cervus elaphus siciliae | Wc | MF | V | X | X | ||
| Bison priscus siciliae | O | Gr | VII | X | X | ||
| Bos primimigenius siciliae | O-Wc | Gr | VII | X | X | ||
| Palaeoloxodon mnaidriensis | O-Wc | Br | VII | X | X | ||
| Ursus arctos | Wc | Om | IV | X | X | ||
| Canis lupus | O-W-Wc | P | III | X | X | X | |
| Vulpes vulpes | O-Wc | sP | II | X | X | X | |
| Sus scrofa | Wc | Om | IV | X | X | X | |
| Equus hydruntinus | O | Gr | V | X | X | ||
| Equus ferus | O | Gr | VI/VII | X | |||
| Lynx lynx | W | P | III | X | |||
| Martes sp. | O-Wc | sP | II | X | |||
| Mustela cf. nivalis | Wc | sP | I | X | |||
| Cervus elaphus | Wc | MF | V | X | |||
| Bos primigenius | O-Wc | Gr | VII | X | |||
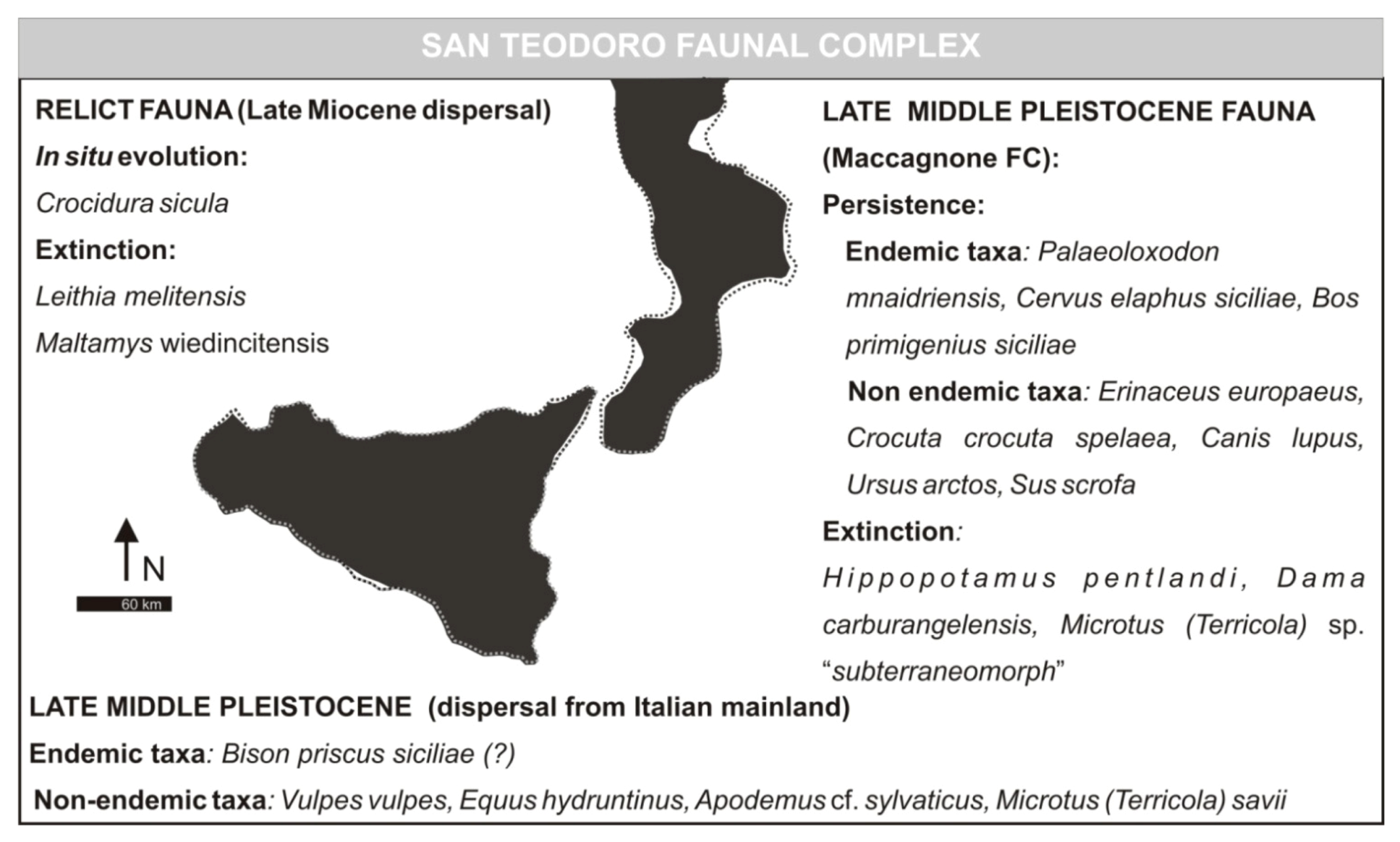
3.4. San Teodoro Faunal Complex (Formerly Grotta S. Teodoro—Pianetti Faunal Complex)
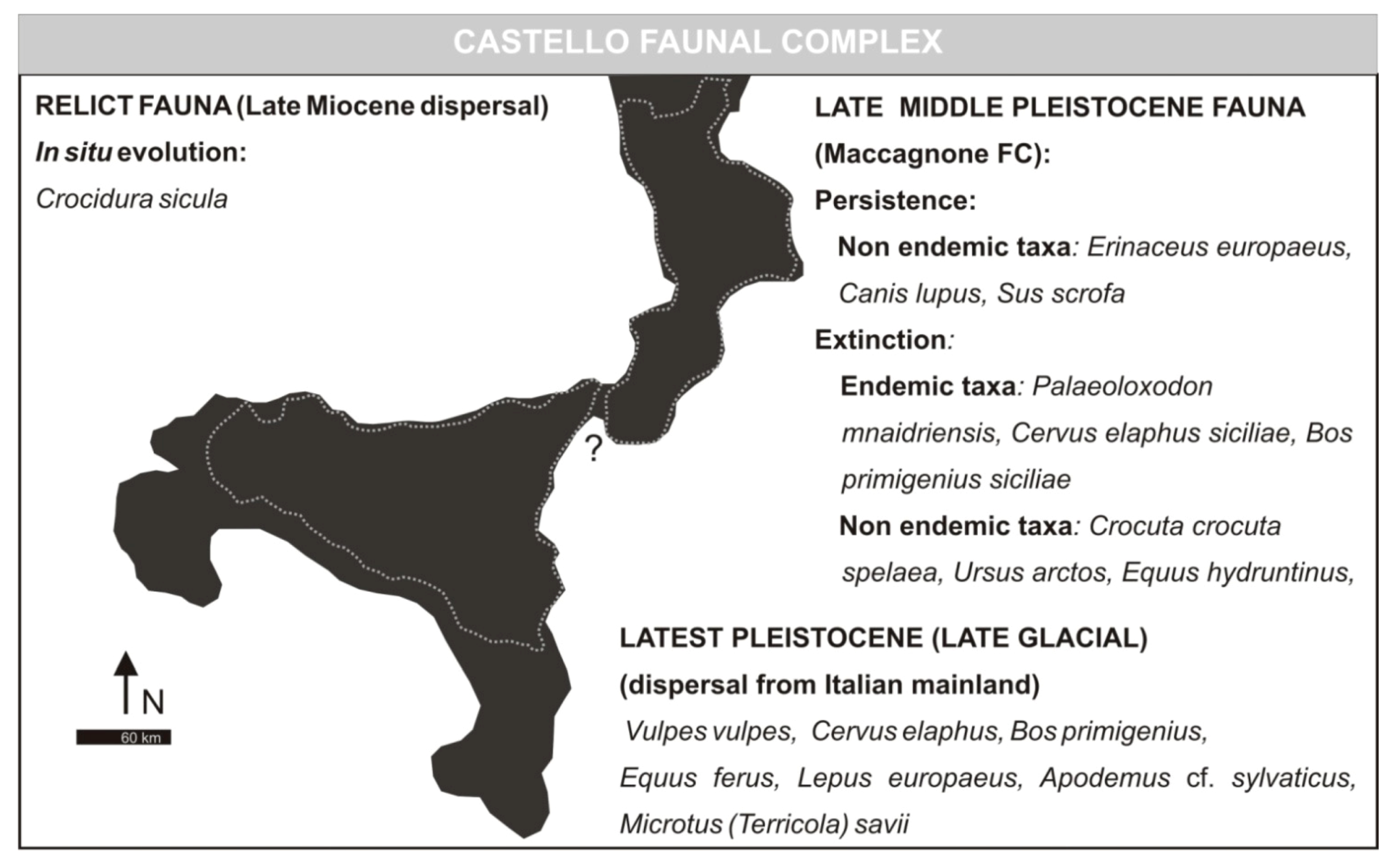
3.5. Castello Faunal Complex
4. Discussion
5. Conclusions
Acknowledgments
References
- Bonfiglio, L.; Marra, A.C.; Masini, F. The contribution of Quaternary vertebrates to the paleoenvironmental and paleoclimatic reconstructions in Sicily. Spec. Pubbl. Geol. Soc. London 2000, 181, 169–182. [Google Scholar]
- Bonfiglio, L.; Mangano, G.; Marra, A.C.; Masini, F.; Pavia, M.; Petruso, D. Pleistocene Calabrian and Sicilian bioprovinces. Geobios. Spec. Mem. 2002, 24, 29–39. [Google Scholar]
- Bonfiglio, L.; di Maggio, C.; Marra, A.C.; Masini, F.; Petruso, D. Biochronology of Pleistocene vertebrate faunas of Sicily and correlation of verterate bearing deposits with marine deposits. Ital. J. Quat. Sci. 2003, 16, 107–114. [Google Scholar]
- Masini, F.; Petruso, D.; Bonfiglio, L.; Mangano, G. Origination and extinction patterns of mammals in three central western Mediterranean islands from the Late Miocene to Quaternary. Quat. Int. 2008, 182, 63–79. [Google Scholar] [CrossRef] [Green Version]
- Marra, A.C. Pleistocene Mammals of Mediterranean Islands. Quat. Int. 2005, 129, 5–14. [Google Scholar] [CrossRef]
- Marra, A.C. Pleistocene mammal faunas of Calabria. Boll. Soc. Paleontol. Ital. 2009, 48, 113–122. [Google Scholar]
- Simpson, G.G. The Geography of Evolution; Chilton Books: New York, NY, USA, 1965. [Google Scholar]
- Dermitzakis, M.D.; Sondaar, P.Y. The importance of fossil mammals in reconstructing palaeogeography wight special reference to the Pleistocene Aegean Archipelago. Ann. Geolog. Pays Ellen. 1978, 29, 808–840. [Google Scholar]
- Sondaar, P.Y.; Dermitzakis, M.D.; de Vos, J. The paleogeography and faunal evolution of the land mammals of Crete. In Pleistocene and Holocene Fauna of Crete and Its First Settlers; Reese, D.S., Ed.; Prehistory Press: Madison, WI, USA, 1996; pp. 61–67. [Google Scholar]
- Barrier, P. Stratigraphie des dépôts pliocènes et quaternaires du Dètroit de Messine [in French]. In Le Détroit de Messine (Italie) Évolution Tectono-Sédimentaire Récente (Pliocène et Quaternaire et Environmnt Actuel); Barrier, P., di Geronimo, I., Montenat, C., Eds.; Institut Géologique Albert de Lapparent (IGAL): Paris, France, 1987; pp. 59–81. [Google Scholar]
- Petruso, D.; Sarà, M.; Surdi, G.; Masini, F. Le faune a mammiferi della Sicilia tra il tardo-glaciale e l’Olocene [in French]. Biogeographia 2011, 30, 27–39. [Google Scholar]
- Gliozzi, E.; Abbazzi, L.; Ambrosetti, P.; Argenti, P.; Azzaroli, A.; Caloi, L.; Capasso Barbato, L.; di Stefano, G.; Esu, D.; Ficcarelli, G.; et al. Biochronology of selected Mammals, Molluscs, Ostracods from the Middle Pliocene to the Late Pleistocene in Italy. The state of the art. Riv. Ital. Palentol. Strat. 1997, 103, 369–388. [Google Scholar]
- Palombo, M.R. Biochronology of Plio-Pleistocene mammalian faunas of the Italian peninsula: Knowledge, problems and perspectives. Ital. J. Quat. Sci. 2004, 17, 565–582. [Google Scholar]
- Thaler, L. Les rongeurs (Rodentia et Lagomorpha) du Monte Pellegrino et la question des anciens isthmus de la Sicilie [in French]. Comptes Rendues de l'Académie de Sciences 1972, 274, 188–190. [Google Scholar]
- Burgio, E.; Fiore, M. Pannonictis arzilla (DE GREGORIO, 1886) a “Villafranchian” element in the fauna of Monte Pellegrino (Palermo, Sicily). Ital. J. Quat. Sci. 1997, 10, 65–74. [Google Scholar]
- Bonfiglio, L.; Esu, D.; Mangano, G.; Masini, F.; Petruso, D.; Soligo, M.; Tuccimei, P. Late Pleistocene vertebrate-bearing deposits at San Teodoro Cave (Northe-Eastern Sicily): Preliminary data on faunal diversification and chronology. Quat. Int. 2008, 190, 26–37. [Google Scholar] [CrossRef]
- Vaufrey, R. Les Elephants nains des iles meditérrannéennes et la question des Isthmes Pléistocène [in French]. Archives de l'Institut de Paléontologie Humaine 1929, 6, 1–220. [Google Scholar]
- Kotsakis, T. Sulle mammalofaune quaternarie siciliane. Boll. Serv. Geol. Ital. 1979, 99, 263–276. [Google Scholar]
- Burgio, E. Le attuali conoscenze sui Mammiferi terrestri quaternary della Sicilia. [in Italian]; In Prima Sicilia:Alle Origini Della Società Siciliana; Tusa, S., Ed.; Edilprint: Palermo, Italy, 1997; pp. 55–72. [Google Scholar]
- Ambrosetti, P.; Azzaroli, A.; Bonadonna, F.P.; Follieri, M. A scheme of Pleistocene chronology for the Thyrrenian side of centra Italy. Boll. Soc. Geol. Ital. 1972, 91, 169–184. [Google Scholar]
- Burgio, E.; Cani, M. Sul ritrovamento di elefanti fossili ad Alcamo (Trapani, Sicilia). Il Naturalista Sicil. 1988, 12, 87–97. [Google Scholar]
- Bonfiglio, L.; Insacco, G. Palaeoenvironmental, paleontologic and stratigraphic significante of vertebrate remains in Pleistocene limnic and alluvial deposits from South Eastern Sicily. Palaeogeogr. Palaeoclimatol. Paleoecol. 1992, 95, 195–208. [Google Scholar] [CrossRef]
- Bada, J.L.; Belluomini, G.; Bonfiglio, L.; Branca, M.; Burgio, E.; Delitala, L. Isoleucine Epimerization ages of Quaternary Mammals of Sicily. Ital. J. Quat. Sci. 1991, 4, 5–11. [Google Scholar]
- Tedford, R.H. Principles and Practices of Mammalian Geochronology in North America. In Proceedings of the North American Paleontological Convention, Chicago, IL, USA, 5–7 September 1969; pp. 666–703, Part F.
- Ferretti, M.P. he dwarf elephant Palaeoloxodon mnaidriensis from Puntali cave, Carini (Sicily; late Middle Pleistocene): Anatomy, systematics and phylogenetic relationships. Quat. Int. 2008, 182, 90–108. [Google Scholar] [CrossRef]
- Bonfiglio, L.; Mangano, G.; Marra, A.C.; Masini, F. A new Late Pleistocene vertebrate faunal complex from Sicily (S. Teodoro cave. North-eastern Sicily, Italy). Boll. Soc. Paleontol. Ital. 2001, 40, 149–1587. [Google Scholar]
- Van der Geer, A.; Lyras, G.; de Vos, J.; Dermitzakis, M. Evolution of Island Mammals Adaptation and Extinction of Placental Mammals on Islands; Wiley-Blackwell: Hoboken, NJ, USA, 2010. [Google Scholar]
- Masini, F.; Sarà, M. Asoricolus burgioi sp. nov. (Soricidae, Mammalia) from the Monte Pellegrino faunal complex (Sicily). Acta Zool. Crac. 1998, 41, 111–124. [Google Scholar]
- Petruso, D. Il Contributo dei Micro Mammiferi Alla Stratigrafia e Paleogeografia del Quaternario Continentale Siciliano. Ph.D. Thesis, Universities of Naples and Palermo, Palermo, Italy, 2003. [Google Scholar]
- Daams, R.; de Bruijn, H. A classification of the Gliridae (Rodentia) on the basis of dental morphology. Hystrix 1995, 6, 3–50. [Google Scholar]
- Fladerer, F.A.; Fiore, M. The Early Pleistocene insular hare Hypolagus pellegrinus sp. nov. from Northern Sicily. Palaeontogr. Ital. 2002, 89, 37–63. [Google Scholar]
- Agnesi, V.; Macaluso, T.; Masini, F. L’ambiente e il clima della Sicilia nell’ultimo milione di anni. [in Italian]; In Prima Sicilia:Alle Origini Della Società Siciliana; Tusa, S., Ed.; Edilprint: Palermo, Italy, 1997; pp. 31–53. [Google Scholar]
- Di Geronimo, I. Bionomie des peuplements benthiques des sustrats meubles et rocheux plio-quaternaires du Dètroit de Messine [in French]. In Le Détroit de Messine (Italie) Évolution Tectono-Sédimentaire Récente (Pliocène et Quaternaire et Environmnt Actuel); [in Italian]; Barrier, P., di Geronimo, I., Montenat, C., Eds.; Institut Géologique Albert de Lapparent (IGAL): Paris, France, 1987; pp. 153–169. [Google Scholar]
- Palombo, M.R. Climate Change versus biotic interaction: A case study of large mammal faunal complexes on the Italian Peninsula from the Pliocene to the Late Pleistocene. New methodological approaches. Cour. Forsch. Senckenberg 2007, 259, 13–46. [Google Scholar]
- Cavinato, G.P.; Petronio, C.; Sardella, R. The Mercure River Basin (Southern Italy): Quaternary stratigraphy and large mammal biochronology. In The World of Elephants, Proceedinggs of the 1st International Congress, Rome, 16–20 October 2001; Cavaretta, G., Gioia, P., Mussi, M., Palombo, M.R., Eds.; Consiglio Nazionale delle Ricerche: Rome, Italy, 2001; pp. 187–190. [Google Scholar]
- Zammit Maempel, G.; de Bruijn, H. The Plio-Pleistocene Gliridae grom the Mediterranean Islands reconsidered. Proc. K. Ned. Akad. Ser. B Phys. 1982, 85, 113–128. [Google Scholar]
- Filippucci, M.G.; Kotkakis, T. Biochemical systematic and evolution of Myoxidae. Hystrix 1994, 6, 77–97. [Google Scholar]
- Kotsakis, T. Crocidura esui n. sp. (Soricidae, Insectivora) du Plèistocène de Spinagallo (Sicile orientale, Italie) [In French]. Geol. Rom. 1986, 23, 51–64. [Google Scholar]
- Esu, D.; Kotsakis, T.; Burgio, E. Vertebrati e Molluschi continentali fossili del Pleistocene di Poggio Schinaldo (Palermo, Sicilia). [in Italian]. Boll. Soc. Geol. Ital. 1986, 105, 233–241. [Google Scholar]
- Sarà, M. The Sicilian (Crocidura sicula) and the Canary (C.canariensis) shrew (Mammalia, Soricidae): Peripheral isolate formation and geographic variation. Boll. Zool. 1995, 62, 173–182. [Google Scholar] [CrossRef]
- Maddalena, T.; Ruedi, M. Chromosomal evolution in the genus Crocidura. In Advances in the Biology of Shrews; Merritt, F.J., Kirkland, G.L., Rose, R.K., Eds.; Carnegie Museum of Natural History Special Publication: Pittsburgh, PA, USA, 1994; Volume 18, pp. 334–344. [Google Scholar]
- Vogel, P.; Cosson, J.P.; Lòpez Jurado, L.F. Taxonomic status and origin of the shrews (Soricidae) from the Canary islands inferred from mtDNA comparison with the European Crocidura species. Mol. Phyl. Evol. 2003, 27, 271–282. [Google Scholar] [CrossRef]
- Dubey, S.; Koyasua, K.; Parapanova, R.; Ribic, M.; Hutterere, R.; Vogela, P. Molecular phylogenetics reveals Messinian, Pliocene, and Pleistocene colonizations of islands by North African shrews. Mol. Phylogenet. Evol. 2007, 47, 877–882. [Google Scholar]
- Johnson, D.L. Problems in the Land Vertebrate Zoogeography of certain islands and the swimming powers of elephants. J. Biogeog. 1980, 7, 383–398. [Google Scholar]
- Quammen, D. The Song of the Dodo. Island Biogeography in an Age of Extinctions; Pimlico: London, UK, 1996; pp. 1–702. [Google Scholar]
- Alcover, J.A.; McMinn, M. Predators of vertebrates on islands. BioScience 1994, 44, 12–18. [Google Scholar] [CrossRef]
- Burgio, E.; Fiore, M. Nesolutra trinacriae n. sp. lontra quaternaria della Sicilia. Boll. Soc. Paleontol. Ital. 1988, 23, 259–275. [Google Scholar]
- Palombo, M.R. How can endemic proboscideans help us to understand the “island rule”? A case study of Mediterranean islands. Quat. Int. 2007, 169–170, 105–124. [Google Scholar] [CrossRef]
- Pavia, M. The evolution dynamics of the Strigiformes in the mediterranean islands with the description of Aegolius martae n. sp. (Aves, Strigidae). Quat. Int. 2008, 182, 80–89. [Google Scholar] [CrossRef]
- Guglielmo, M.; Marra, A.C. Le due Sicilie del Pleistocene Medio: osservazioni paleogeografiche. [in Italian]. Biogeographia 2011, 30, 11–25. [Google Scholar]
- Pavia, M. A new large Barn Owl (Aves, strigiformes, Tytonidae) from the Middle Pleistocene of Sicily, Italy, and its taphonomical significance. Geobios 2004, 37, 631–641. [Google Scholar] [CrossRef]
- Raia, P.; Barbera, C.; Conte, M. The fast life of a dwarfed elephant. Evol. Morph. 2003, 17, 293–312. [Google Scholar]
- Petruso, D.; Locatelli, E.; Dalla Valle, C.; Masini, F.; Sala, B. Phylogeny and biogeography of fossil and extant Microtus (Terricola) (Mammalia, Rodentia) of Sicily and the southern Italian peninsula based on current. Quat. Int. 2011, 243, 192–203. [Google Scholar] [CrossRef]
- Colantoni, P. Marine geology of the Strait of Messina. In Le Détroit de Messine (Italie) Évolution Tectono-Sédimentaire Récente (Pliocène et Quaternaire et Environmnt Actuel); Barrier, P., Di Geronimo, I., Montenat, C., Eds.; Institut Géologique Albert de Lapparent (IGAL): Paris, France, 1987; pp. 191–210. [Google Scholar]
- Abbazzi, L.; Bonfiglio, L.; Marra, A.C.; Masini, F. A revision of medium and small sized deer from the Middle and Late Pleistocene of Calabria and Sicily. Boll. Soc. Paleontol. Ital. 2001, 40, 115–126. [Google Scholar]
- Brugal, J.P. Cas de ”Nanisme” Insulaire dans L’Auroche [in Franch]. In Proceedings of 112th Congrès National des Sociétés Savantes, Lyon, France, 1987; pp. 53–66.
- Marra, A.C. Pleistocene Hippopotamuses of Mediterranean Islands: Looking for Ancestors. In Proceedings of the International Symposium Insular Vertebrate Evolution: The Palaeontological Approach, Mollorca, Spain, 16–19 September 2003.
- Marra, A.C. Ursus arctos from selected Pleistocene sites of eastern Sicily. Boll. Soc. Paleontol. Ital. 2003, 42, 145–150. [Google Scholar]
- Yll, R.; Carriòn, J.S.; Marra, A.C.; Bonfiglio, L. Vegetation reconstruction on the basis of pollen in late Pleistocene hyena coprolites from San Teodoro Cave (Sicily, Italy). Palaeogeogr. Palaeoclim. Palaeoecol. 2006, 237, 32–39. [Google Scholar] [CrossRef]
- Allen, J.R.M.; Watts, W.A.; Huntley, B. Weichselian palynostratigraphy, palaeovegetation and palaeoenvironment: The record from Lago Grande di Monticchio, southern Italy. Quat. Int. 2000, 74, 91–110. [Google Scholar] [CrossRef]
- Pavia, M. The Middle Pleistocene Fossil Avifauna from the “Elephas mnaidriensis Faunal Complex” of Sicily (Italy). Preliminary Results. In The World of Elephants—International Congress, Proceedings of the First International Congress, Rome, Italy, 16–20 October 2001; pp. 497–501.
- Lambeck, K.; Antonioli, F.; Purcell, A.; Silenzi, S. Sea-level change along the Italian coasts for the past 10,000 years. Quat. Sci. Rev. 2004, 23, 1567–1598. [Google Scholar] [CrossRef]
- Masini, F.; Sala, B. Large- and small-mammal distribution patterns and chronostratigraphic boundaries from the Late Pliocene to the Middle Pleistocene of the Italian peninsula. Quat. Int. 2007, 160, 43–56. [Google Scholar] [CrossRef] [Green Version]
- Seguenza, G. I vertebrati fossili della provincia di Messina: Parte II, Mammiferi e Geologia del piano Pontico. [in Italian]. Boll. Soc. Geol. Ital. 1902, 21, 115–175. [Google Scholar]
- Seguenza, G. Nuovi resti di mammiferi pontici di Gravitelli presso Messina. [in Italian]. Boll. Soc. Geol. Ital. 1907, 26, 7–122. [Google Scholar]
- Rook, L.; Mazza, P.; Rustioni, M.; Torre, D. Lands and Endemic Mammals in the Late Miocene of Italy: Paleogeographic Outlines of Tyrrhenian and Adriatic Areas between 11–9 and 7–4 Ma. In Proceedings of Environments and Ecosystem Dynamics of the Eurasian Neogene (EEDEN), State of the Art Workshop, Lyon, France, 16–18 November 2000; pp. 56–58.
- Herridge, V.L. Dwarf Elephants on Mediterranean Islands: A Natural Experiment in Parallel Evolution. Ph.D. Thesis, University College London, London, UK, 2010. [Google Scholar]
- Alcover, J.A.; Sans, A.; Palmer, M. The extent of extinction of mammals on islands. J. Biogeogr. 1998, 25, 913–918. [Google Scholar]
- Louchart, A. Integrating the Fossil Record in the Study of Insular Body Size Evolution: Examples in Owls (Aves: Strigiformes). In Proceedings of the Internation Symposyum Insular Vertebrate Evolution: the Palaeontological Approach, Mollorca, Spain, 16–19 September 2003; pp. 155–174.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Marra, A.C. Evolution of Endemic Species, Ecological Interactions and Geographical Changes in an Insular Environment: A Case Study of Quaternary Mammals of Sicily (Italy, EU). Geosciences 2013, 3, 114-139. https://doi.org/10.3390/geosciences3010114
Marra AC. Evolution of Endemic Species, Ecological Interactions and Geographical Changes in an Insular Environment: A Case Study of Quaternary Mammals of Sicily (Italy, EU). Geosciences. 2013; 3(1):114-139. https://doi.org/10.3390/geosciences3010114
Chicago/Turabian StyleMarra, Antonella Cinzia. 2013. "Evolution of Endemic Species, Ecological Interactions and Geographical Changes in an Insular Environment: A Case Study of Quaternary Mammals of Sicily (Italy, EU)" Geosciences 3, no. 1: 114-139. https://doi.org/10.3390/geosciences3010114