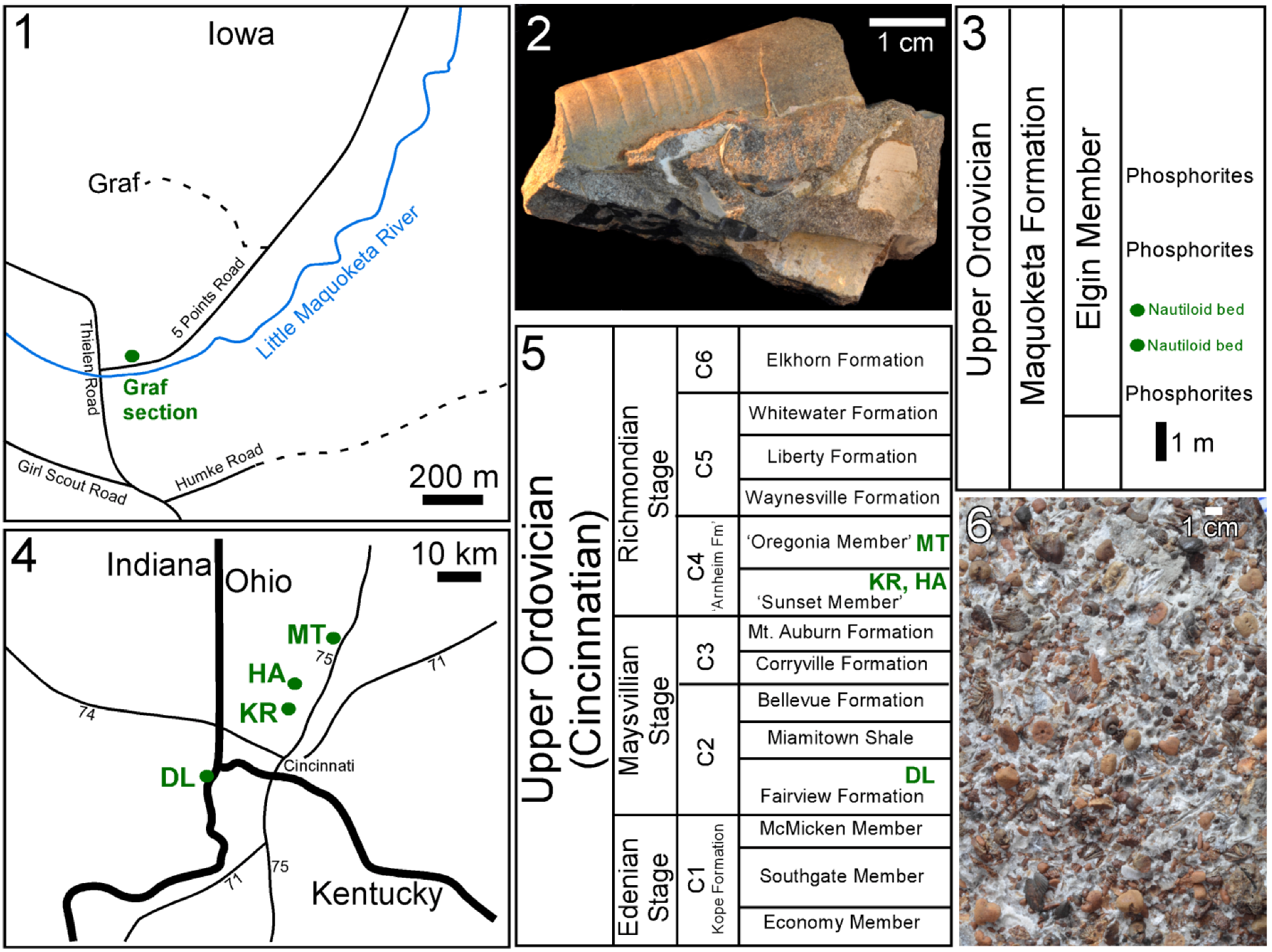
3.1. Isorthoceras sociale and Other Molluscs from the Ordovician of Iowa
Many fossils of the cephalopod
Isorthoceras sociale from the Graf section, Iowa have a pearly lustre on at least some part of the shell surface (
Figure 1.3 and
Figure 2.1). Horizontal and tangential sections are seen in SEM to consist of the texture that Mutvei [
11] described: somewhat regularly spaced vertical stacks of flat, sub-circular to polygonal units with a thin column in the centre of each stack. These units are referred to here as discs and their relationship to aragonite tablets of nacre is discussed below.
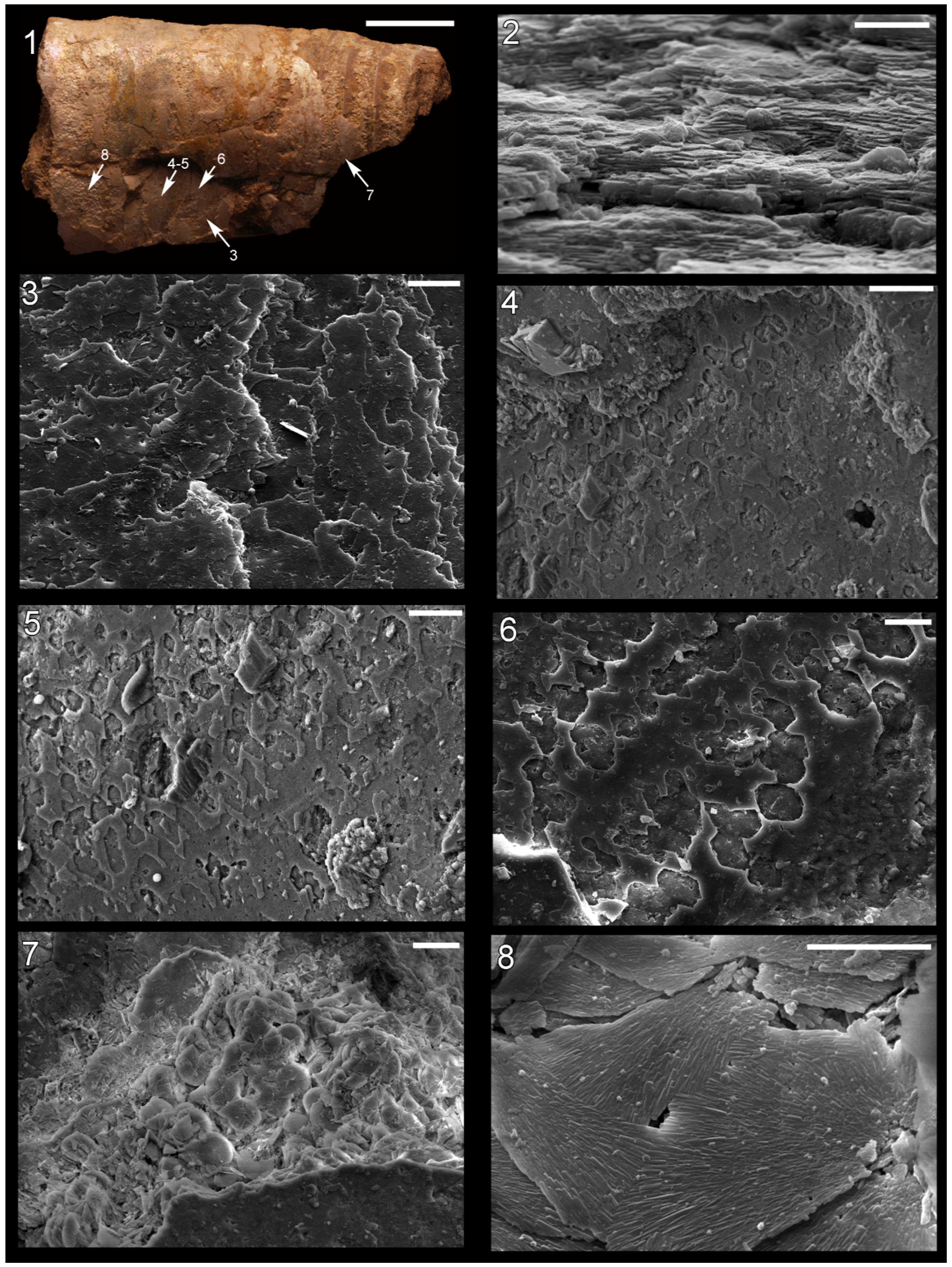
It is clear in our photos that the discs in adjacent stacks are laterally continuous with each other, forming broad laminae in tangential section (
Figure 2.2 and
Figure 3.2,
Figure 3.3,
Figure 3.4). Lamina thickness averages 218 nm (median 196 nm; standard deviation 57 nm; n = 73), and in
Figure 3.2 over 100 stacked layers can be seen. The space between laminae averages 410 nm (measured from midpoint of layer to that of overlying layer; median also 410 nm; standard deviation 62 nm; n = 74). This compares favourably with measurements in modern molluscs from our photographs and those from the literature: the mean of this dataset was 475 nm, median 429 nm, and standard deviation 17.4 nm (n = 76).
In horizontal sections merged discs can be seen (
Figure 2.7 and
Figure 2.8), although occasionally the discs in adjacent stacks are somewhat vertically offset from each other where they meet (
Figure 3.1,
Figure 4.2 and
Figure 4.3). In vertical (e.g., transverse) sections obliquely vertical columns can be seen (
Figure 3.3 and
Figure 3.4) that make up the centres of discs—these represent pores or canals that ran vertically through successive laminae.
The discs are sub-circular in horizontal section (
Figure 3.1), and typically range from about 10–34 µm in diameter (mean 21 µm; median 19.3 µm; standard deviation 5.9 µm; n = 40). These discs are larger than the nacreous tablets of modern molluscs: in 95 nacre tablets in modern molluscs, measured from our photographs and those in the literature, the mean diameter was 6.0 µm, median 5.5 µm, and standard deviation 2.9 µm. Open space between adjacent discs is common in these fossils, and in regions of some fossils, small discs occur at the top of a stack (
Figure 4.7). Open space between adjacent nacre tablet stacks with incipient tablets on top is common at the growth front of nacre, on the inner surface of the shell, of many modern gastropods (
Figure 6.2). However, in these fossils the voids are typically within the shell (
Figure 2.7 and
Figure 4.1), a location that in modern cephalopods and vetigastropods is characterized by fully formed nacre without inter-tablet space. The voids in the fossils were probably produced taphonomically, as described below.
Upon further magnification the discs are seen to consist of radiating crystals (
Figure 3.1 and
Figure 4.3). These crystals are polygonal in cross section (hexagonal whenever the outline can unequivocally be recognized;
Figure 3.5 and
Figure 3.6) and multiple spectra reveal that they have a composition of calcium phosphate (
Figure 4.8 and
Figure 4.9), as suggested by Grandjean
et al. [
28], who argued these crystals represent the dahllite form of apatite. There are typically 1–3 vertically stacked crystals in any one part of a disc (
Figure 3.6,
Figure 3.7 and
Figure 3.8). The central column is in some cases preserved as a cavity or mould (
Figure 3.2) and in other instances is a positive cast (
Figure 3.3 and
Figure 3.4) of the original structure, consisting of many individual apatite crystals that connect to the fossil laminae (
Figure 3.7 and
Figure 3.8).
Figure 2.
Shell microstructure in specimens of Isorthoceras from the Ordovician Maquoketa Formation. (1), (3)–(8) LACMIP 14434, arrows in (1) show locations of images in (3)–(8); (2) LACMIP 14435, view of semi-horizontal section of inner shell layer, top of photo is more internal. Scale bars: 10 µm for (1)–(3); 20 µm for (4)–(7); 5 µm for (8).
Figure 2.
Shell microstructure in specimens of Isorthoceras from the Ordovician Maquoketa Formation. (1), (3)–(8) LACMIP 14434, arrows in (1) show locations of images in (3)–(8); (2) LACMIP 14435, view of semi-horizontal section of inner shell layer, top of photo is more internal. Scale bars: 10 µm for (1)–(3); 20 µm for (4)–(7); 5 µm for (8).
Figure 3.
Shell microstructure in specimens of Isorthoceras from the Ordovician Maquoketa Formation. (1) LACMIP 14434, horizontal view of disc in middle shell layer, proximal region of the shell, topmost disc is the most external; (2)–(4), (7) and (8) LACMIP 14437; (5) and (6) LACMIP 14438. (2)–(8) Transverse sections of middle or inner shell layer where the top of the photograph is external. Scale bars: 5 µm for (1), (2) and (4); 1 µm for (3); 500 nm for (5) and (6); 200 nm for (7) and (8).
Figure 3.
Shell microstructure in specimens of Isorthoceras from the Ordovician Maquoketa Formation. (1) LACMIP 14434, horizontal view of disc in middle shell layer, proximal region of the shell, topmost disc is the most external; (2)–(4), (7) and (8) LACMIP 14437; (5) and (6) LACMIP 14438. (2)–(8) Transverse sections of middle or inner shell layer where the top of the photograph is external. Scale bars: 5 µm for (1), (2) and (4); 1 µm for (3); 500 nm for (5) and (6); 200 nm for (7) and (8).
Figure 4.
Shell microstructure in specimens of Isorthoceras from the Ordovician Maquoketa Formation. (1), (5) and (7) LACMIP 14439, horizontal sections of internal shell layer near internal surface, view is towards external surface; (2) and (4) LACMIP 14438, transverse sections of middle shell layer, top of photographs is external; (3) LACMIP 14440, horizontal section of inner or middle shell layer, top of discs is internal; and (6), (8) and (9) LACMIP 14437: (6) horizontal section of inner or middle shell layer, view towards external surface; (8) EDS analysis of crystal within the disc, revealing composition of calcium phosphate; (9) vertical section through shell showing location of EDS analysis depicted in (8). Scale bars: 100 µm for (1); 2 µm for (2)–(4); 10 µm for (5); 1 µm for (6) and (9); and 5 µm for (7).
Figure 4.
Shell microstructure in specimens of Isorthoceras from the Ordovician Maquoketa Formation. (1), (5) and (7) LACMIP 14439, horizontal sections of internal shell layer near internal surface, view is towards external surface; (2) and (4) LACMIP 14438, transverse sections of middle shell layer, top of photographs is external; (3) LACMIP 14440, horizontal section of inner or middle shell layer, top of discs is internal; and (6), (8) and (9) LACMIP 14437: (6) horizontal section of inner or middle shell layer, view towards external surface; (8) EDS analysis of crystal within the disc, revealing composition of calcium phosphate; (9) vertical section through shell showing location of EDS analysis depicted in (8). Scale bars: 100 µm for (1); 2 µm for (2)–(4); 10 µm for (5); 1 µm for (6) and (9); and 5 µm for (7).
In one specimen of
Isorthoceras that we examined, a few spots on the outer surface of the preserved shell contain polygonal imprints on a flat sheet (
Figure 2.4,
Figure 2.5 and
Figure 2.6). The polygonal outlines are consistent with imprints of incipient nacre tablets. However, the growth front of nacre in modern molluscs occurs at the innermost surface of the shell, whereas in this fossil the structure occurs at different levels
within the shell. These sheets are thin (less than 1 µm in thickness), and multiple lamellae can be seen even in dorsal view in places where the topmost layer is missing. The flat, polygonal and semi-polygonal imprints (moulds) show a broad range of shapes, but often with distinct straight sides. The angles where these straight sides meet fall into three rough groupings: 55°–64°, 105°–110°, and 119°–140°. This is similar to what is expected in nacre: 64°, 90°, 116° and 122° [
1], although measurements of these angles in tablets from our photographs and those in the literature show a more continuous distribution from 98° to 152° (n = 54). The measured angles in
Isorthoceras differ from that of calcite rhombohedra, 78° and 102°, expected if the shell microstructure were calcitic semi-nacre [
3]. The tablet moulds interconnect in places, but where separate the width of individual tablet moulds ranges from about 6 µm to 14 µm (mean 10 µm; median 9.1 µm; standard deviation 3.0 µm; n = 21). This compares favourably with measurements of tablets in modern nacre (mean = 6.0 µm; n = 95).
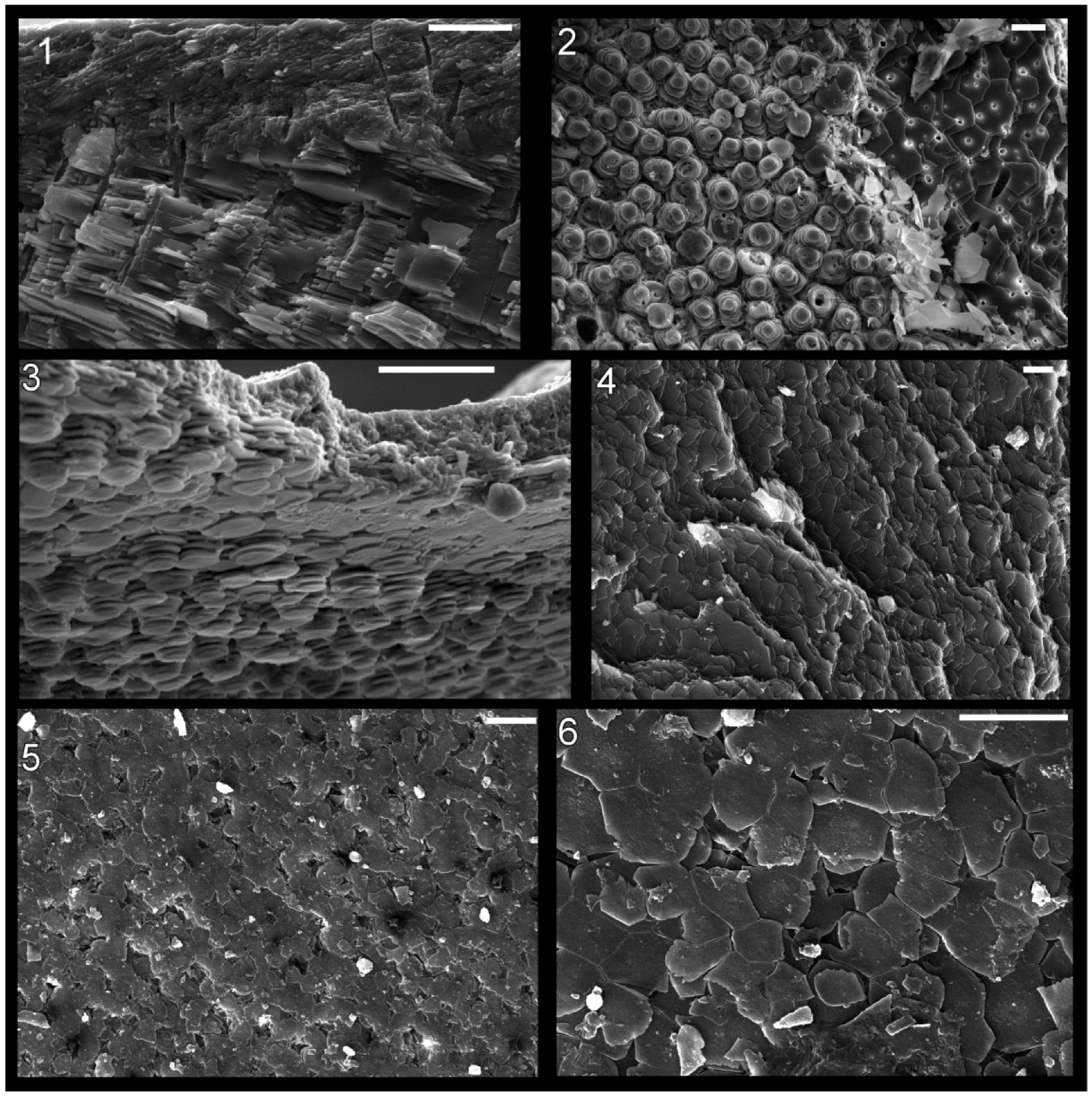
The shell microstructure in
Isorthoceras is laminar and consisted of roughly equidimensional units, as in modern nacre. Overall the textures in these fossils are highly reminiscent of modern nacre (e.g.,
Figure 6.1,
Figure 6.2 and
Figure 6.3), consisting of stacks of roughly equidimensional tablets, each with a central column. Key measurements such as laminae thickness and interfacial angles fall easily within the range of nacre in modern molluscs. The discs in
Isorthoceras are larger than expected if they represent nacre tablets, but the diameters of moulds of tablets in this fossil are within the expected range for tablets in modern nacre.
Isorthoceras discs contain a radial orientation of crystals instead of what is seen upon etching of modern nacre tablets: 2 and 3 sectors, each with a non-radial (parallel) orientation of “fibres”. The crystals in
Isorthoceras are often grouped into clumps (
Figure 3.1,
Figure 4.3 and
Figure 4.7), but this grouping is different from what is seen in partly dissolved nacre tablets of modern molluscs. Moreover, the disc constituents in
Isorthoceras have a polygonal cross-section not seen in the dissolution “fibres” of etched nacre tablets. Mutvei [
11] regarded the radiating “crystalline sectors” (p. 239) in
Isorthoceras to be similar to the dissolution “fibres” in modern
Nautilus Linnaeus, 1758 and
Haliotis Linnaeus, 1758, but we favour the hypothesis that diagenesis produced the radial elements in
Isorthoceras for reasons outlined below.
In general, Mutvei [
11] envisioned that the texture in
Isorthoceras represents a fairly accurate copy of the original nacre, but our observations indicate much of the structure is diagenetic. The detailed photographs of the central columns show that they consist of crystals of calcium phosphate that had grown upward or downward and that at regular vertical intervals extended radially outward to form the discs (
Figure 3.7). These observations suggest that phosphatization occurred from the external and/or internal shell surface into the shell. The diagenetic nature of the texture in the fossils is also revealed by: (1) the intermingling of calcium phosphate with disorganized, often large calcite rhombs (
Figure 4.5 and
Figure 4.6); (2) the cases where the phosphate crystals in adjacent discs are vertically offset from each other (
Figure 4.2); and (3) apatite crystals radiating outward in a less orderly manner, along a non-horizontal plane (
Figure 4.4).
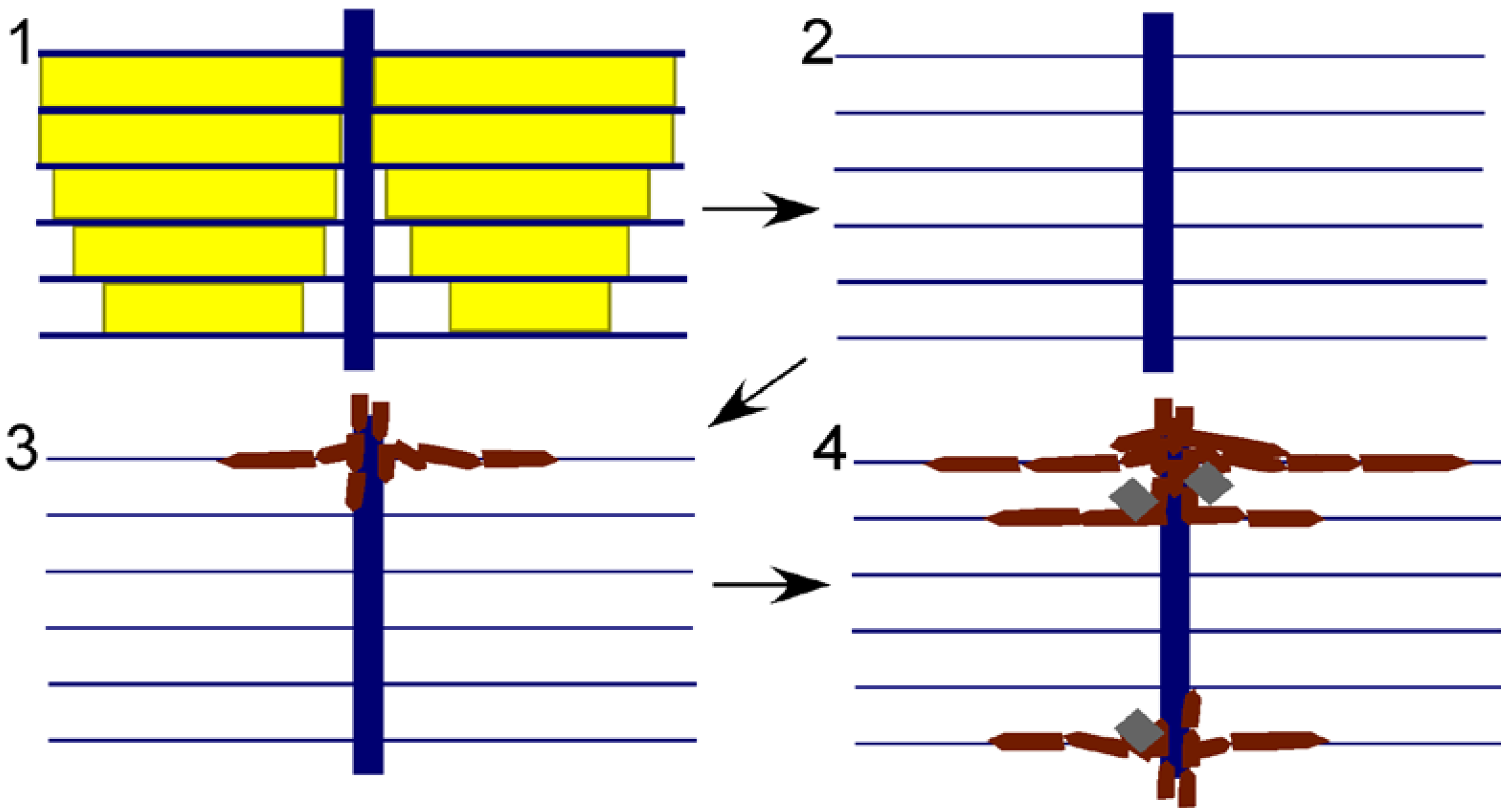
The apatite discs must have formed in the spaces left behind by the dissolution of nacre tablets. The original horizontal organic membranes must have been intact during phosphatization in order to produce the replica of nacre, and it is likely that bacterial decomposition of the organic matrices caused the precipitation of apatite therein. The organic-rich cores through the stacks of tablets would have allowed phosphatization to proceed from layer to layer, as revealed by the apatite replacement of the originally organic columns (
Figure 3.7). If this model (
Figure 5) is correct, then diagenesis in a way replayed the original mode of mineralization within the fossil shell, even replicating the pearly lustre of the original nacre. This hypothesis provides an alternative explanation for the high frequency of spaces between discs. Mutvei [
11] used this observation to infer that a large proportion of the shell was organic (hence his term “flexible nacre” for the fossil microstructure), but in our model the spaces represent regions where the phosphate infiltration had ceased. This model of phosphate mineralization can also explain the puzzling observation that, opposite to modern nacre, discs are often progressively smaller towards the external surface of the shell. This configuration would be expected where phosphate infiltration of the shell progressed from the shell chamber towards the external shell surface. The model can also explain the irregularity in spacing of the vertical columns that formed the centres of discs, and the large size of discs. Phosphatization might not have infiltrated each original column, allowing for some discs to grow larger before meeting a neighbour than did the original nacre tablets.
Figure 5.
Model for preservation of nacre in Isorthoceras from the Maquoketa Formation. External shell surface is above, internal surface below. Key: yellow shows original aragonitic nacre tablets; blue shows organic matrix; white is empty space; burgundy polygons represent secondary phosphate; and grey rhombohedra are secondary calcite. Steps: (1) original shell; (2) aragonite is dissolved, leaving behind the organic matrix; and (3) and (4) calcium phosphate infiltrates the organic matrix through the vertical tunnels, with calcite also occasionally precipitating within the empty spaces of the shell.
Figure 5.
Model for preservation of nacre in Isorthoceras from the Maquoketa Formation. External shell surface is above, internal surface below. Key: yellow shows original aragonitic nacre tablets; blue shows organic matrix; white is empty space; burgundy polygons represent secondary phosphate; and grey rhombohedra are secondary calcite. Steps: (1) original shell; (2) aragonite is dissolved, leaving behind the organic matrix; and (3) and (4) calcium phosphate infiltrates the organic matrix through the vertical tunnels, with calcite also occasionally precipitating within the empty spaces of the shell.
Replacement of calcium carbonate by apatite typically involves dissolution of the carbonate followed quickly by precipitation of apatite into the space left behind [
29]. Our model incorporates this component, but more specifically is similar to the model Balthasar [
30] described for the phosphatization of originally calcitic brachiopods from the Cambrian of Canada. He envisioned carbonate dissolution followed by apatite forming first on the external surface of the shell and then progressing inward through vertical canals. In this way the calcium phosphate penetrated all the shell layers down to the internal surface of the shell, similar to what occurred in
I. sociale.
In sum, the original shell microstructure of
I. sociale consisted of thin horizontal laminae penetrated by roughly equally-spaced organic columns. This form is seen only in nacre (see
Figure 6 for some modern examples of nacre). Nacre was defined by Carter
et al. [
31] as “polygonal to rounded aragonitic tablets arranged in broadly continuous, regular, mutually parallel laminae” (p. 611). Thus we agree with Grégoire [
10] and Mutvei [
11] that nacre is the type of shell microstructure replicated here.
Figure 6.
Shell microstructure in recent molluscs. (1) and (2) the gastropod Arene sp. SBMNH 230515, (1) semi-transverse section near aperture showing innermost about 1/4 thickness of shell with top of photograph internal, (2) horizontal section of inner shell layer near internal shell surface, view is towards external surface; (3) the gastropod Seguenzia sp.,MNHN 8528, semi-horizontal section of inner shell layer near internal surface, top of photograph is external; and (4)–(6) the cephalopod Nautilus pompilius, semi-horizontal and horizontal sections of the inner shell layer near internal shell surface at the aperture of the shell, view towards exterior. Scale bars all 10 µm. Image (3) was taken by Daniel Geiger.
Figure 6.
Shell microstructure in recent molluscs. (1) and (2) the gastropod Arene sp. SBMNH 230515, (1) semi-transverse section near aperture showing innermost about 1/4 thickness of shell with top of photograph internal, (2) horizontal section of inner shell layer near internal shell surface, view is towards external surface; (3) the gastropod Seguenzia sp.,MNHN 8528, semi-horizontal section of inner shell layer near internal surface, top of photograph is external; and (4)–(6) the cephalopod Nautilus pompilius, semi-horizontal and horizontal sections of the inner shell layer near internal shell surface at the aperture of the shell, view towards exterior. Scale bars all 10 µm. Image (3) was taken by Daniel Geiger.
3.2. Bivalves and Gastropods from the Ordovician Maquoketa Formation of Iowa
Mutvei [
11] mentioned that gastropods from the Maquoketa Formation show the same texture as what occurs in cephalopods from that rock unit; in another publication he [
12] provided one SEM photograph each of a bivalve and gastropod that show a similar structure. We examined 25 gastropods, 25 bivalves, and one bellerophont and found a few subtle traces of the
Isorthoceras-like shell microstructure in these other taxa.
Some patches on a gastropod,
Holopea Hall, 1847, contain adjoining discs that have a radiating ultrastructure (
Figure 7.5). Other gastropods show a distinct laminar texture but with the constituent crystallites unclear (
Figure 7.6). The merged tablets in the gastropod
Murchisonia d’Archiac and E.P. de Verneuil, 1841 [
12] (fig. 2) are clearly very similar to what occurs in
Isorthoceras. However Mutvei [
11,
12] did not include images of vertical sections and we were unable to find textures in the Maquoketa gastropods we examined that had the level of detail that characterizes the cephalopods from that unit.
The Maquoketa bivalve
Palaeoneilo Hall and Whitfield, 1869 appears to have had a laminar shell microstructure (
Figure 7.1), and many fossil bivalves (and gastropods) from these beds contain diagenetic rosettes (
Figure 7.3 and
Figure 7.4). The one image of shell microstructure in a bivalve from the Maquoketa Formation provided by Mutvei [
12] (fig. 3) records tablet-like groupings of elongate crystals that are similar to discs in
Isorthoceras. These bundles are not as well organized as is usual for specimens of
Isorthoceras, but they are consistent with what occurs in some regions of these fossils. Carboniferous specimens of
Palaeoneilo oweni (McChesney, 1859) show a prismatic outer shell layer, porcelaneous middle layer, and fine complex crossed lamellar (CCL), matted, and/or homogenous inner shell layer [
32]. The Ordovician specimens of
Palaeoneilo described herein show laminae but equivocal tablets and thus may have had a matted shell microstructure consisting of laminae without tablets, as in
P. oweni. Such a shell microstructure has been postulated to be an evolutionary transition between nacre and fine CCL [
32] and so it might be expected to comprise much of the middle and inner shell layers in the Ordovician representatives of this genus.
Mutvei [
12] described differences between shell microstructure in the bivalve
Palaeoconcha with what occurs in both
Isorthoceras and the gastropod
Murchisonia from the same formation.
Palaeoconcha, unlike the others, appeared to lack pores (or accumulations in the centre of discs) as well as vertical stacks of discs. This may be due to incomplete preservation, or perhaps another type of shell microstructure besides nacre is preserved here and similarities with textures in
Isorthoceras are due to a similar diagenetic history. Unfortunately, as for the gastropods, vertical sections confirming a nacre-like configuration of the discs are currently unknown. However, Carter [
33] described prismatic and nacreous shell layers in the Silurian
Praenucula faba Liljedahl, 1994, a taxon that, like
Palaeoconcha, is a member of the Praenuculidae. Carter’s [
33] observation corroborates the conclusion that nacre is present in
Palaeoconcha from the Ordovician. We likewise agree with Carter [
33] that
Palaeoconcha from the Maquoketa Formation shows nacre, and that phosphatization increased the size of discs from that of the original tablets.
Combining the data here with photographs in Mutvei [
12], we tentatively conclude that gastropods and bivalves from the Maquoketa Formation show a similar fossil microstructure to
Isorthoceras that represents phosphatized nacre. Our model that this texture is largely diagenetic, while still revealing nacre, helps explain why three different classes of mollusc have a similar, unusual fossil shell texture.
Figure 7.
Shell microstructure in specimens of the bivalve Palaeoneilo sp. (1)–(4) and the gastropod Holopea sp. (5) and (6) from the Ordovician Maquoketa Formation. (1) and (2) LACMIP 14441: (1) semi-transverse section of a portion of replaced shell near midpoint of valve with umbo towards top of photo; (2) horizontal section of replaced shell near aperture; (3) LACMIP 14442, horizontal section of replaced shell in region between valve midpoint and aperture; (4) LACMIP 14436, horizontal section of inferred inner shell layer, near midpoint of valve; (5) USNM 553688, horizontal section of shell replacement; (6) LACMIP 14443, semi-horizontal view of external shell surface (outer shell layer) along lateral region of shell. Scale bars: 20 µm for (1), (3)–(5); 5 µm for (2); 10 µm for (6).
Figure 7.
Shell microstructure in specimens of the bivalve Palaeoneilo sp. (1)–(4) and the gastropod Holopea sp. (5) and (6) from the Ordovician Maquoketa Formation. (1) and (2) LACMIP 14441: (1) semi-transverse section of a portion of replaced shell near midpoint of valve with umbo towards top of photo; (2) horizontal section of replaced shell near aperture; (3) LACMIP 14442, horizontal section of replaced shell in region between valve midpoint and aperture; (4) LACMIP 14436, horizontal section of inferred inner shell layer, near midpoint of valve; (5) USNM 553688, horizontal section of shell replacement; (6) LACMIP 14443, semi-horizontal view of external shell surface (outer shell layer) along lateral region of shell. Scale bars: 20 µm for (1), (3)–(5); 5 µm for (2); 10 µm for (6).
3.3. Molluscs from the Ordovician Cyclora beds of Indiana and Ohio
In mollusc fossils from the
Cyclora beds of the Kope, Fairview, and “Arnheim” Formations distinct shell microstructure was rarely observed. In contrast, imprints of fine microstructure are preserved in great detail in most internal moulds of the brachiopod
Zygospira (Hall, 1862) and in fossil trilobite spines (
Figure 8), and thus the fidelity of preservation in
Cyclora beds is sufficient to preserve shell microstructure imprints in molluscs. However, of the more than 400 mollusc fossils examined via SEM, only a few showed unambiguous textures. Nevertheless, the details on some of these specimens were distinct enough to allow inferences to be made about original shell microstructure.
Figure 8.
Imprints of shell microstructure in the brachiopod Zygospira (1) and (3) and a trilobite spine (2) and (4) from the “Arnheim Formation” Cyclora beds of Ohio. Images (1) and (2) are electronically inverted versions of the originals in (3) and (4), to show what the crystal structures imprinted in the fossils originally looked like. (1) and (3) Zygospira sp., LACMIP 14448, from the “Arnheim Formation” near Kemper Road, Cincinnati, Ohio, horizontal surface of internal mould; and (2) and (4) trilobite spine, LACMIP 14449, possibly from the “Arnheim Formation”, Hamilton, Ohio, horizontal surface of internal mould. Scale bars: 50 µm for (1) and (3); and 5 µm for (2) and (4).
Figure 8.
Imprints of shell microstructure in the brachiopod Zygospira (1) and (3) and a trilobite spine (2) and (4) from the “Arnheim Formation” Cyclora beds of Ohio. Images (1) and (2) are electronically inverted versions of the originals in (3) and (4), to show what the crystal structures imprinted in the fossils originally looked like. (1) and (3) Zygospira sp., LACMIP 14448, from the “Arnheim Formation” near Kemper Road, Cincinnati, Ohio, horizontal surface of internal mould; and (2) and (4) trilobite spine, LACMIP 14449, possibly from the “Arnheim Formation”, Hamilton, Ohio, horizontal surface of internal mould. Scale bars: 50 µm for (1) and (3); and 5 µm for (2) and (4).
Bivalves from the
Cyclora beds show the most distinct shell microstructure of any type of mollusc in the deposits. In some specimens of
Palaeoconcha, sub-circular and polygonal imprints occur throughout the surface of the internal mould (
Figure 9.3,
Figure 9.4,
Figure 9.5,
Figure 9.6 and
Figure 9.7). The range of shapes is consistent with growing nacre. Measurements of interfacial angles could be made in six cases, and these ranged from 113° to 132° (mean 123.2°), close to the 122° angle expected in nacre. Widths of the moulds of tablets ranged from 4 µm to 13.5 µm (mean 8.3 µm; median 8.6 µm; standard deviation 2.3 µm; n = 25). These measurements are similar to those of tablets in modern nacre (mean = 6.0 µm; median = 5.5 µm; standard deviation = 2.9 µm; n = 95). Moreover, in some regions of these fossils (
Figure 9.6), lines in high relief can be seen intersecting the polygonal depressions. These lines appear to correspond to the boundaries between merged tablets of the underlying lamella (more external in the original shell). These observations support the hypothesis that nacre was present in bivalves at this time, corroborated by Mutvei’s [
12] observations of this genus from the Maquoketa Formation, and Carter’s [
32] observations of Silurian representatives of the family.
Imprints of shell microstructure in internal moulds of bivalves from the Ordovician of the Midwest are distinct from those of the Cambrian bivalves
Pojetaia Jell, 1980 and
Fordilla Barrande, 1881. Namely the crystallites in
Pojetaia/
Fordilla are elongate blade-like laths, whereas the sub-units of the Ordovician bivalves are roughly equidimensional tablets. This microstructure of elongate blades of aragonite arranged in layers was termed “foliated aragonite” by Checa
et al. [
34,
35], although Carter
et al. [
36] suggested the proper term for this shell microstructure should be “semi-foliated aragonite”. In addition to the blade-like laths, in some regions of fossils of
Pojetaia, large, isolated, rounded tablets also occur [
9], a type of microstructure that Carter
et al. [
36] classified as “large tablet, imbricated nacre” (p. 88). Carter [
33] previously suggested that this type of microstructure was transitional to nacre. Thus it may be that the transition from foliated (or semi-foliated) aragonite to nacre began in
Pojetaia and its kin, and was finalized in the close ancestors of the
Cyclora-bed bivalves. This transition from foliated aragonite to nacre was hypothesized by Vendrasco
et al. [
9] and is illustrated in
Figure 10.
Internal moulds of gastropods from the
Cyclora beds show distinct layered textures, indicating that the innermost shell microstructure was laminar. However, details of the crystalline constituents of the lamellae have not yet been seen, and so it is unclear which type of laminar microstructure is preserved here. In general, coiled shells from Cambrian deposits tend to have lamello-fibrillar shell microstructure [
3,
37], where “the horizontal fibers in successive laminae differ in orientation by irregularly varying angles” (p. 611) [
31].
A small region near the aperture of one gastropod fossil contains a texture that appears to show nacre very clearly (
Figure 9.1 and
Figure 9.2). However, this texture seems to be below the surface of the internal mould. Although there are scenarios whereby part of the original shell might have been broken and collapsed inward, producing the pattern here, the lack of such textures elsewhere on the surface of the internal mould or on partial casts must call into question the classification of the texture as nacre. If it is not nacre, it will provide a cautionary tale about the over-interpretation of tantalizing but rare textures on fossils. This possibility highlights the need for finding the same textures in multiple specimens of the same species. An alternative interpretation is that this patch represents a fragment of a smaller shell lodged within the empty gastropod chamber prior to infill with phosphate. Such occurrences are common in other deposits of phosphatized molluscs [
38]. If this latter hypothesis is correct, then some unknown member of the
Cyclora fauna had nacreous shell microstructure.
Figure 9.
Shell microstructures in a specimen of the gastropod Cyclora (Cyclonema?) (1) and (2); the bivalves Palaeoconcha sp. (3)–(7); and the bellerophont Cyrtolites sp. (8) from Ordovician localities around Cincinatti, Ohio. (1) and (2) LACMIP 14444, from the Fairview Formation near Dearborn/Lawrenceburg, Indiana, arrow in (2) shows location of (1); (3)–(6) LACMIP 14445, from the Fairview Formation near Dearborn/Lawrenceburg, Indiana: (3) horizontal view of surface of internal mould near midpoint of valve; (4) horizontal view of surface of internal mould near aperture (broken aperture mould at top of photo); (5) horizontal view of surface of internal mould near umbo; (6) horizontal view of surface of internal mould near midpoint of valve; (7) LACMIP 14446, from the “Arnheim Formation” along Kemper Road, Cincinnati, Ohio , horizontal view of surface of internal mould in region between midpoint of valve and umbo; and (8) LACMIP 14447, from the “Arnheim Formation” in Middletown, Ohio, horizontal view of internal mould or thin shell replacement near columellar side of aperture, imaginary line normal to plane of aperture would extend diagonally down/leftward on the photo. Scale bars: 10 µm for (1), (6) and (7); 200 µm for (2); and 20 µm for (3)–(5) and (8).
Figure 9.
Shell microstructures in a specimen of the gastropod Cyclora (Cyclonema?) (1) and (2); the bivalves Palaeoconcha sp. (3)–(7); and the bellerophont Cyrtolites sp. (8) from Ordovician localities around Cincinatti, Ohio. (1) and (2) LACMIP 14444, from the Fairview Formation near Dearborn/Lawrenceburg, Indiana, arrow in (2) shows location of (1); (3)–(6) LACMIP 14445, from the Fairview Formation near Dearborn/Lawrenceburg, Indiana: (3) horizontal view of surface of internal mould near midpoint of valve; (4) horizontal view of surface of internal mould near aperture (broken aperture mould at top of photo); (5) horizontal view of surface of internal mould near umbo; (6) horizontal view of surface of internal mould near midpoint of valve; (7) LACMIP 14446, from the “Arnheim Formation” along Kemper Road, Cincinnati, Ohio , horizontal view of surface of internal mould in region between midpoint of valve and umbo; and (8) LACMIP 14447, from the “Arnheim Formation” in Middletown, Ohio, horizontal view of internal mould or thin shell replacement near columellar side of aperture, imaginary line normal to plane of aperture would extend diagonally down/leftward on the photo. Scale bars: 10 µm for (1), (6) and (7); 200 µm for (2); and 20 µm for (3)–(5) and (8).
![Geosciences 03 00001 g009]()
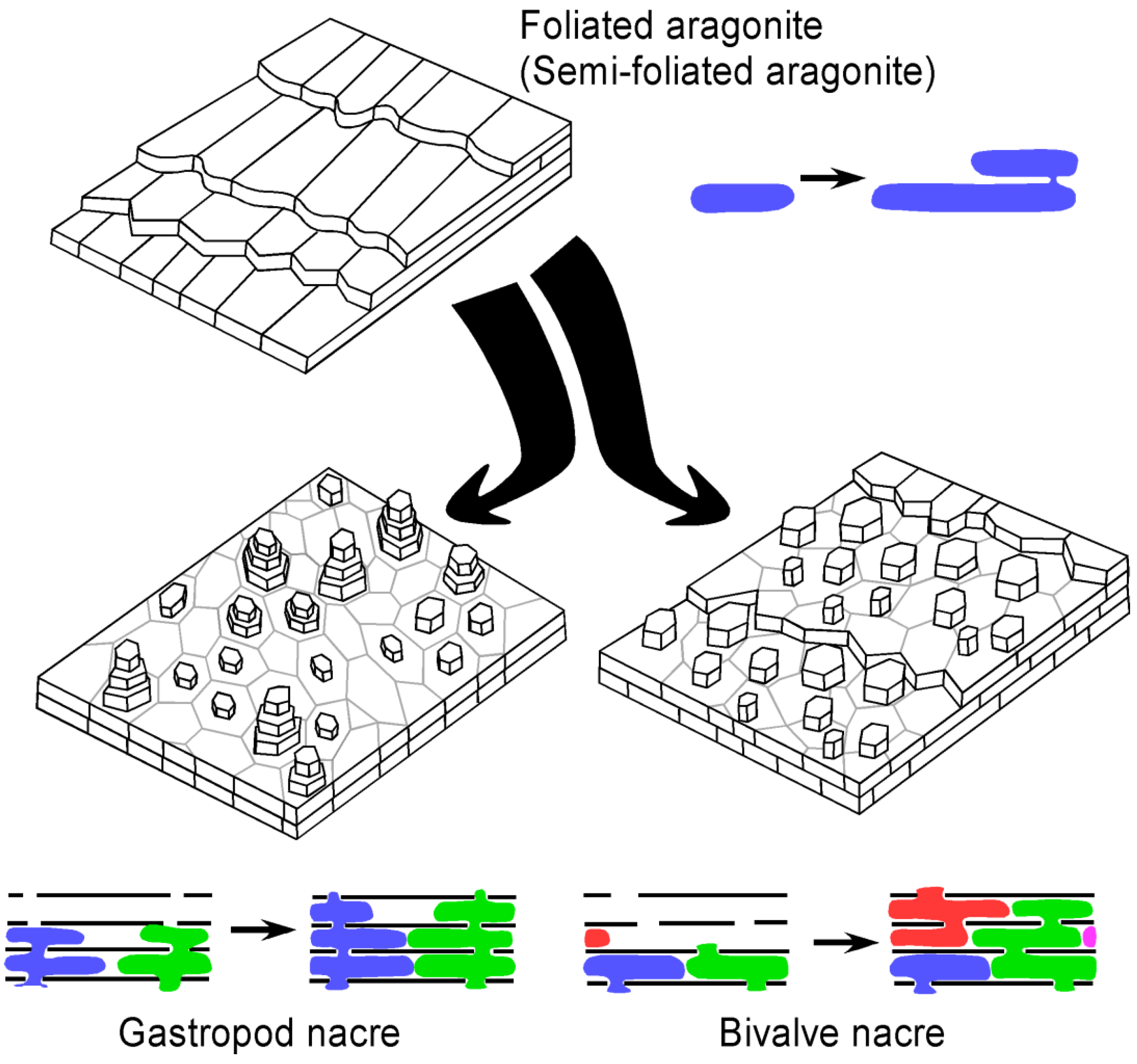
Figure 10.
3-D reconstruction block diagrams and inferred mode of growth of foliated (or semi-foliated) aragonite as well as bivalve and gastropod nacre. The colours represent different aragonite crystals growing through interlamellar membranes. The transformation of foliated (or semi-foliated) aragonite to nacre could have been achieved by addition of a porous interlamellar membrane.
Figure 10.
3-D reconstruction block diagrams and inferred mode of growth of foliated (or semi-foliated) aragonite as well as bivalve and gastropod nacre. The colours represent different aragonite crystals growing through interlamellar membranes. The transformation of foliated (or semi-foliated) aragonite to nacre could have been achieved by addition of a porous interlamellar membrane.
Some bellerophonts from the
Cyclora beds likewise reveal a laminar inner shell microstructure (
Figure 9.8). One specimen shows slightly raised polygonal sectors. This may represent a replication of nacre tablets, but it has thus far been seen in only one bellerophont, and the outlines of these sectors, while consistent with the range of nacre, show no characteristic that would allow the exclusion of the hypothesis that they are artefacts of diagenesis.
Such cases of equivocal shell microstructure in fossils suggest some principles in doing work on fossil shell microstructures: (1) the same texture should be seen on multiple specimens of the same species to provide certainty that the texture is characteristic of that species; and (2) if the same texture is seen in multiple types of distantly related fossils from the same bed, then some or all of the texture was likely produced by diagenesis.
3.4. Diversification of Shell Microstructures in Early Palaeozoic Molluscs
An overview of the distribution of shell microstructures in molluscs from the Cambrian and Ordovician reveal some basic patterns (
Table 1). First, shell microstructures rapidly diversified during the early stages of molluscan evolution, and early molluscs had a composite shell that was composed of multiple types of shell microstructure. In the early Cambrian alone, at least 7 types of shell microstructure occurred in molluscs.
Table 1.
Shell microstructure data for Cambrian molluscs.
Moreover, there is a clear correlation between shell microstructure configuration and phylogenetic relationships among Cambrian molluscs: for example, highly coiled molluscs tended to have lamello-fibrillar [
3]; stenothecids had an internal shell layer of calcitic semi-nacre [
4]; the
Anabarella Vostokova, 1962–
Watsonella Grabau 1900 lineage had the same complex three-component shell microstructure consisting of fibrous, foliated (or semi-foliated) aragonite, and prismatic textures [
39]; bivalves had foliated (or semi-foliated) aragonite [
9]; and
Pseudomyona Pojeta & Runnegar, 1976 and
Tuarangia Mackinnon, 1982 had foliated calcite [
40]. Shell microstructures appear to be fairly conserved in some lineages through the Cambrian Period, although it is also clear that modifications continued to take place in various mollusc lineages.
Prismatic was probably the first type of shell microstructure to originate in molluscs, although it may have been accompanied by some type of laminar inner shell microstructure. Prismatic shell microstructure is among the most common during the early Cambrian, and it is the typical type of outer shell microstructure in modern molluscs. Feng and Sun [
37] noted the commonality of lamello-fibrillar microstructure (a configuration similar to what occurs in plywood) in Cambrian molluscs, and this is seen in
Table 1 as well. This type of shell microstructure also occurs in the problematic mollusc
Ocruranus Liu, 1979 [
41], and it may represent the ancestral type of inner shell microstructure in molluscs. Thus the archetypal mollusc may have had a bi-layer shell with crystals in a prismatic configuration in the outer layer and lamello-fibrillar texture in the inner one.
Also, it appears that aragonitic shell mineralogy was common in newly originated metazoan taxa during the early Cambrian whereas calcitic microstructures became more common in those originating during the middle Cambrian [
42]. This pattern reflects the change from “aragonite seas” to “calcite seas” around this time [
42], although
within molluscs the record does not show a tight correlation with this aspect of seawater chemistry (
Table 1). Transitions from aragonite to calcite have been shown to occur in abundance during a switch to calcite seas [
43], but calcite can also be favoured for other reasons like temperature and in bivalves switches to calcite often occurred during times of aragonite seas [
44,
45]. Thus it is still unclear to what extent these changes in seawater chemistry controlled the transitions in the mineralogy of mollusc shells at this time.
Foliated (or semi-foliated) aragonite was recently discovered in modern monoplacophorans [
34,
35], and this type of microstructure characterized the earliest known bivalves [
9]. A similar shell microstructure can also be seen in other molluscs from the early and middle Cambrian [
9]. This shell microstructure appears to have been modified into nacre in bivalves, and possibly also other molluscs like monoplacophorans and cephalopods.
3.7. Evolution and Escalation during the Great Ordovician Biodiversification Event
Predation is a strong selective pressure. Evidence from the fossil record reveals that as predators became increasingly efficient at capturing prey through geologic time, ever-improving defensive co-adaptations evolved in their prey. This evolutionary escalation or arms race between predator and prey produced many of the notable characteristics of different groups of modern animals such as thick shells, mineral-reinforced jaws and teeth, powerful claws, sensory systems, and mobility.
Escalation is documented in molluscs through the Mesozoic and Cainozoic Eras (from 250 million years ago until now) [
52]. However, the earliest history of the arms race between predators and mollusc prey is poorly known, and most studies of escalation with molluscs focus only on macroscopic changes in the shell or on signs of predation like shell breakage. In addition, factors besides predation can also strongly influence mollusc shell mineralogy and microstructure, and the comparative importance of defence in controlling shell microstructure is still unclear. It is also uncertain to what extent nacre is ideal under intense predation, as many bivalves and gastropods appear to have lost it since the early Mesozoic while predation intensity increased [
46].
The arms race between predators and their molluscan prey began about 540 million years ago with the earliest diversification of animals during the “Cambrian Explosion” [
53]. This arms race appears to have accelerated during the continuation of this early major adaptive radiation: the Great Ordovician Biodiversification Event (GOBE, beginning about 490 million years ago). The compilation of key evolutionary events related to predation (
Table 2) shows a prominent increase in both predatory and defensive abilities during both the Cambrian and Ordovician. Specific evidence for predatory attacks on shelled prey from the Ordovician of the Midwestern United States include shell breakage and repair in brachiopod shells [
54], a fossil of an asteroid wrapped around a bivalve [
55], damage and repair to crinoid skeletons [
56,
57,
58], and damage to trilobite skeletons [
59]. Alexander [
54] hypothesized that nautiloids were the likely shell-breaking culprits in these cases; Babcock [
59] agreed and added eurypterids as an additional suspect.
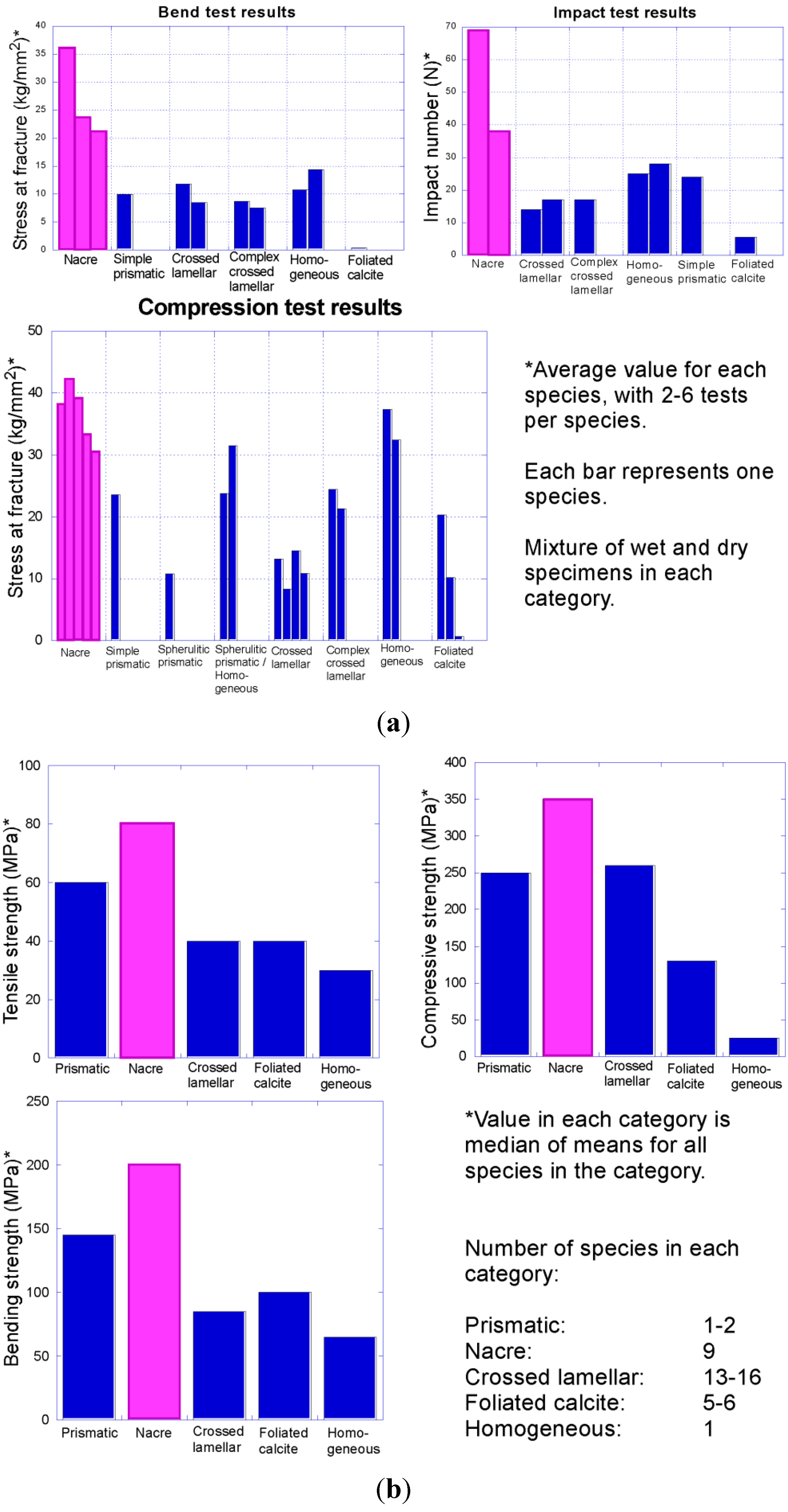
Increasing intensity of predation beginning in the early Palaeozoic put pressure on early molluscs to defend themselves. One of the primary ways that molluscs avoid being eaten is to strengthen the shell via thickening and through specific shell microstructures. Early tests indicated that nacre is the strongest shell microstructure (
Figure 11) [
5,
6]. Thus nacre might be expected to be selectively favoured under conditions of high shell-crushing intensity, when the benefits of forming a costly structure outweigh the disadvantages. However previous studies on strength of shell microstructure were not comprehensive in testing. Additional work is needed to test the strength of
all major types of shell microstructures in molluscs, to demonstrate whether nacre is in fact the ideal shell microstructure under most scenarios of high predation. Nacre is thought to possess its strength largely through the high abundance of organic matter within the shell, but not all other types of shell microstructure with high organic content have been tested for strength. For example, calcitic semi-nacre as well as calcitic prismatic and aragonitic granular prismatic shell microstructures have a high proportion of organic matrix in the shell but were never previously subjected to biomechanical tests against nacre. Nacre has been lost in many molluscan lineages, which might indicate it is not ideal under all types of predation pressure, or it may reflect the success of other predator-avoidance strategies.
One possible reason why nacre has been lost repeatedly in molluscs is that it is an energetically expensive type of shell microstructure to form [
7,
8]. This hypothesis is supported by the high proportion of organic matrix within nacre compared to other types of shell microstructure. The organic component should entail a greater cost than the mineral component of a shell, but the hypothesis that nacre is the most expensive type of shell microstructure has likewise not been rigorously tested. For example, the energetic cost of formation has not been estimated for the aforementioned calcitic semi-nacre as well as calcitic prismatic and aragonitic granular prismatic shell microstructures.
Table 2.
Estimated time of first appearance of key features in the early arms race between predators and their molluscan prey. Innovations listed in red are predatory, those in black are defence, and the one in blue indicates evidence for shell crushing predation. Data derived from [
1,
52,
54,
60,
61,
62,
63,
64,
65,
66] and herein.
Table 2.
Estimated time of first appearance of key features in the early arms race between predators and their molluscan prey. Innovations listed in red are predatory, those in black are defence, and the one in blue indicates evidence for shell crushing predation. Data derived from [1,52,54,60,61,62,63,64,65,66] and herein.
| Predator/prey innovation | Terreneuvian Cambrian | Series 2 Cambrian | Series 3 Cambrian | Furongian Cambrian | Lower Ordovician | Middle Ordovician | Upper Ordovician |
|---|
| 542 Ma | 521 Ma | 509 Ma | 497 Ma | 542 Ma | 542 Ma | 542 Ma |
|---|
| Shell drilling | ![Geosciences 03 00001 i010]() | | | | | | |
| Whole animal ingestion | | ![Geosciences 03 00001 i010]() | | | | | |
| Grasping appendages | | ![Geosciences 03 00001 i010]() | | | | | |
| >500 cm long predators | | ![Geosciences 03 00001 i010]() | | | | | |
| Arthropod predators | | ![Geosciences 03 00001 i010]() | | | | | |
| Durophagy | | ![Geosciences 03 00001 i010]() | | | | | |
| Crossed-lamellar shell | | ![Geosciences 03 00001 i011]() | | | | | |
| Type 1 driller (Brett) | | | ![Geosciences 03 00001 i010]() | | | | |
| Cephalopod predators | | | | ![Geosciences 03 00001 i010]() | | | |
| Large, thick mollusc shells | | | | ![Geosciences 03 00001 i011]() | | | |
| Diversification of cephalopods | | | | | ![Geosciences 03 00001 i010]() | | |
| Type 2 driller (Brett) | | | | | ![Geosciences 03 00001 i010]() | | |
| Very large (>1.5 m) predators | | | | | ![Geosciences 03 00001 i010]() | | |
| Echinoderm predators | | | | | ![Geosciences 03 00001 i010]() | | |
| Hard annelid jaws | | | | | ![Geosciences 03 00001 i010]() | | |
| Diversification of conodonts | | | | | ![Geosciences 03 00001 i010]() | | |
| Eurypterids | | | | | | ![Geosciences 03 00001 i010]() | |
| Shell lip peeling | | | | | | ![Geosciences 03 00001 i010]() | |
| Brachiopod spines | | | | | | ![Geosciences 03 00001 i011]() | |
| Large (>60 mm) shell scars | | | | | | ![Geosciences 03 00001 i012]() | |
| Durophagous eurypterids | | | | | | | ![Geosciences 03 00001 i011]() |
| Crinoid spines | | | | | | | ![Geosciences 03 00001 i011]() |
| Thick crinoid calyx | | | | | | | ![Geosciences 03 00001 i011]() |
| Brachiopod geniculation | | | | | | | ![Geosciences 03 00001 i011]() |
| Brachiopod commissure ridge | | | | | | | ![Geosciences 03 00001 i011]() |
| Nacre | | | | | | | ![Geosciences 03 00001 i011]() |
Figure 11.
Compilation of data on comparative strength of molluscan shell microstructures. Note that nacre (pink) consistently outperformed all other shell microstructures in tests of toughness in these two separate studies: (
a) Fracture resistance of different bivalve shell microstructures (data from Taylor and Layman [
5]); (
b) strength of different mollusc shell microstructures (data from Currey [
6]).
Figure 11.
Compilation of data on comparative strength of molluscan shell microstructures. Note that nacre (pink) consistently outperformed all other shell microstructures in tests of toughness in these two separate studies: (
a) Fracture resistance of different bivalve shell microstructures (data from Taylor and Layman [
5]); (
b) strength of different mollusc shell microstructures (data from Currey [
6]).













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
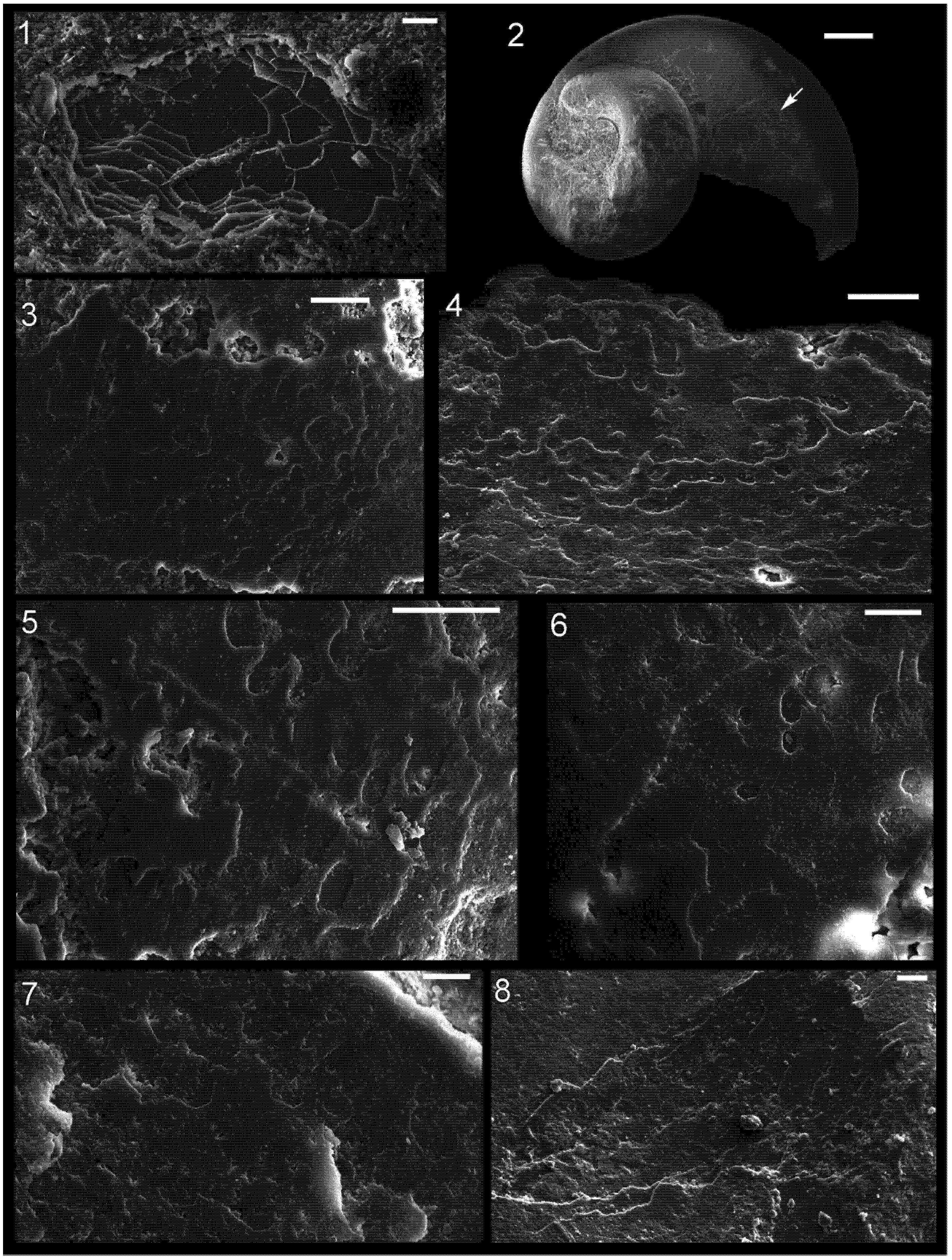{kind=link}
{kind=link}
{kind=link}












