Early Silurian (Aeronian) East Point Coral Patch Reefs of Anticosti Island, Eastern Canada: First Reef Recovery from the Ordovician/Silurian Mass Extinction in Eastern Laurentia

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
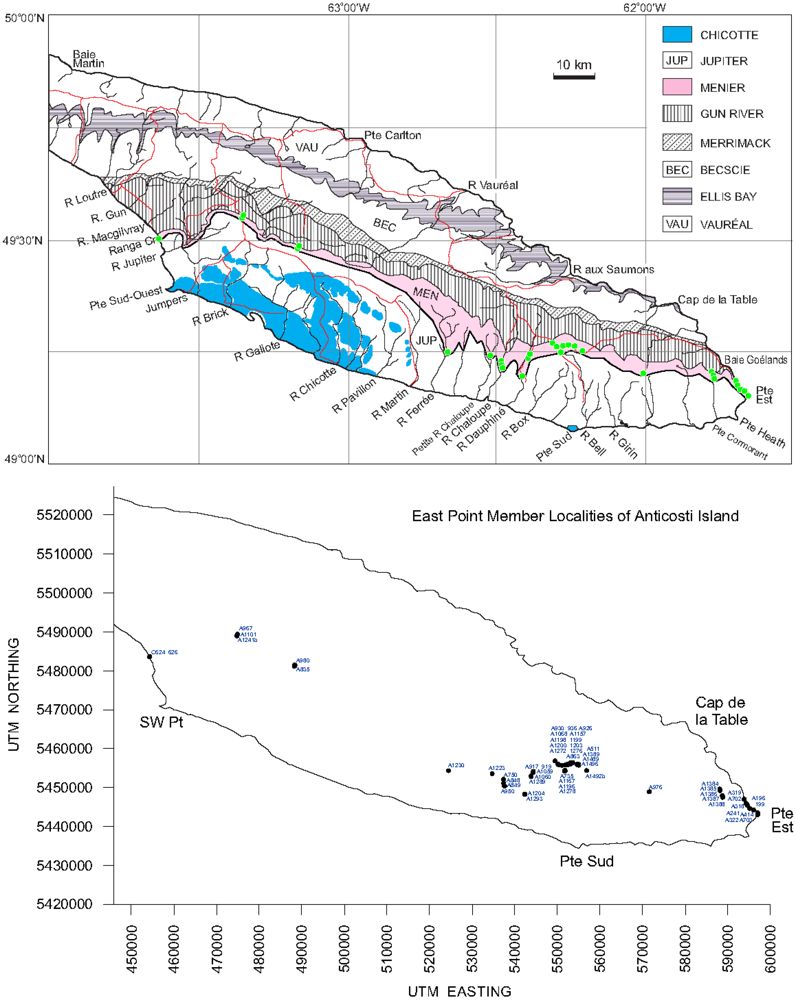
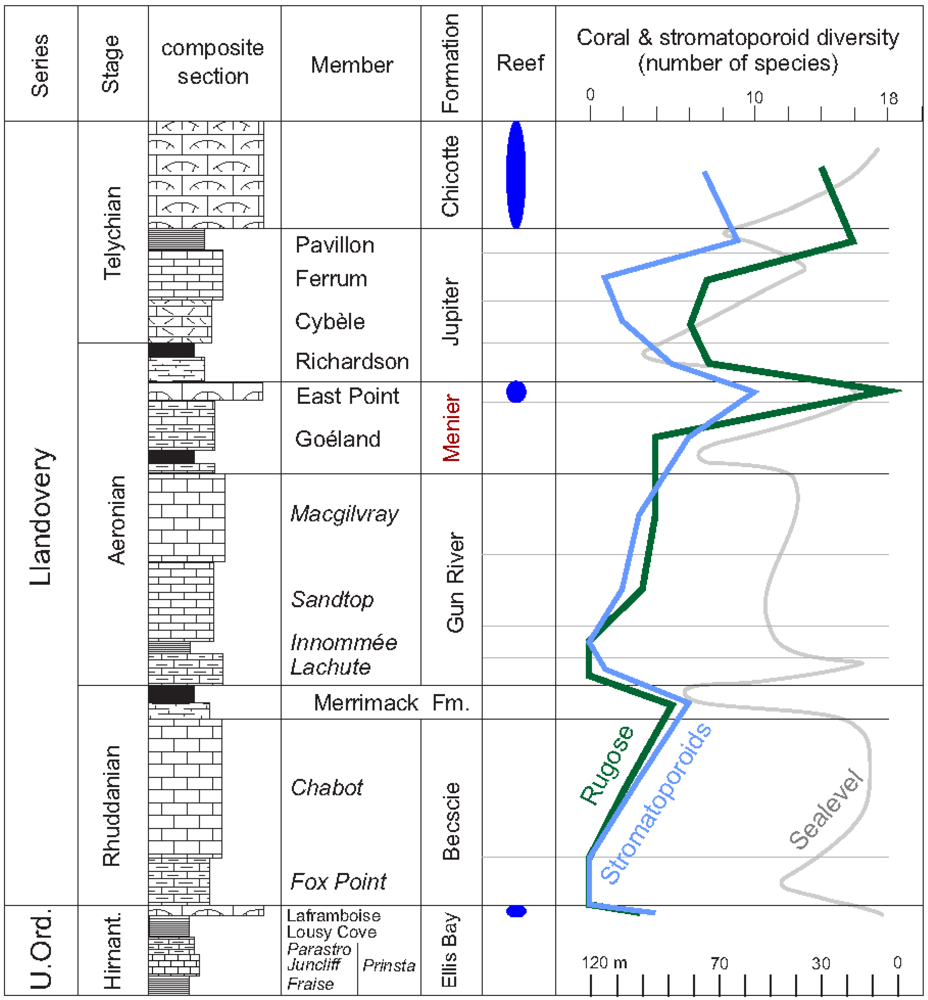
2. The Anticosti Section
- (1) The intent to designate a formal unit. The Menier Formation is proposed herein as a formal stratigraphic unit.
- (2) Category and rank of unit. Menier Formation (new), consists of the two previously established Goéland and East Point members.
- (3) Selection and derivation of the name. The new formation is named after the Menier family from Chateau Chenonceau, France, the owners of Anticosti Island after Louis Jolliet.
- (4) Stratotype of the two members. See Copper and Long [26].
- (5) Description of unit. See formal descriptions of the Goéland and East Point members by Copper and Long [26].
- (6) Boundaries. The lower boundary of the Menier Formation is the base of the Goéland Member, and the upper boundary the top of the East Point Member, as defined in Copper and Long [26].
- (7) Historical background. See Copper and Long [26].
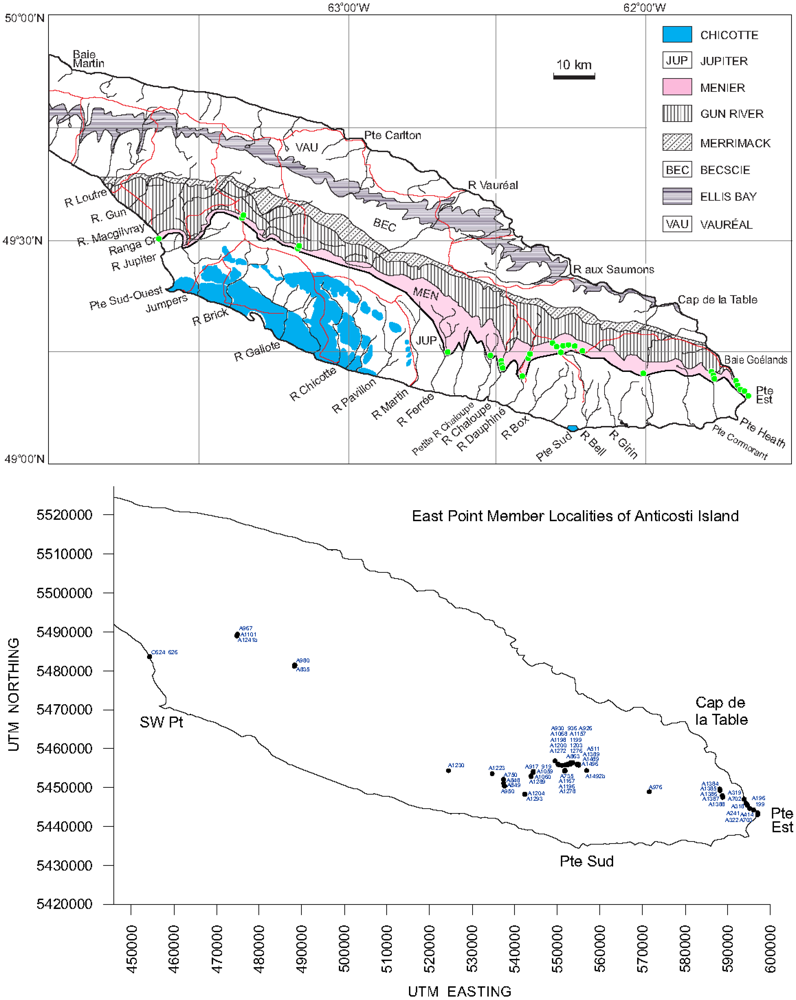
- (8) Dimensions of the unit. The Menier Formation has a total thickness of 65 m, and extends from the eastern end to the southwestern coast of Anticosti Island as a continuous belt (Figure 1).
- (9) Age and correlations. The first appearance datum (FAD) of Kulumbella and Pentamerus in the uppermost Macgilvray Member of the Gun River Formation, which directly underlies the Menier Formation, indicates a mid-Aeronian age. Corals and brachiopods within the new formation carry elements readily differentiated from those of the overlying Jupiter and Chicotte formations. The Richardson and lower Cybèle members of the Jupiter Formation above carry a Stimulograptus sedgwicki fauna, denoting a latest Aeronian age. The base of the East Point Member in the Box River region contains common graptolites alongside abundant Stegerhynchus, although the graptolites remain to be identified. The faunal data, therefore, suggest a mid- to late Aeronian age for the Menier Formation.

3. Distribution and Geometry of the East Point Reefs



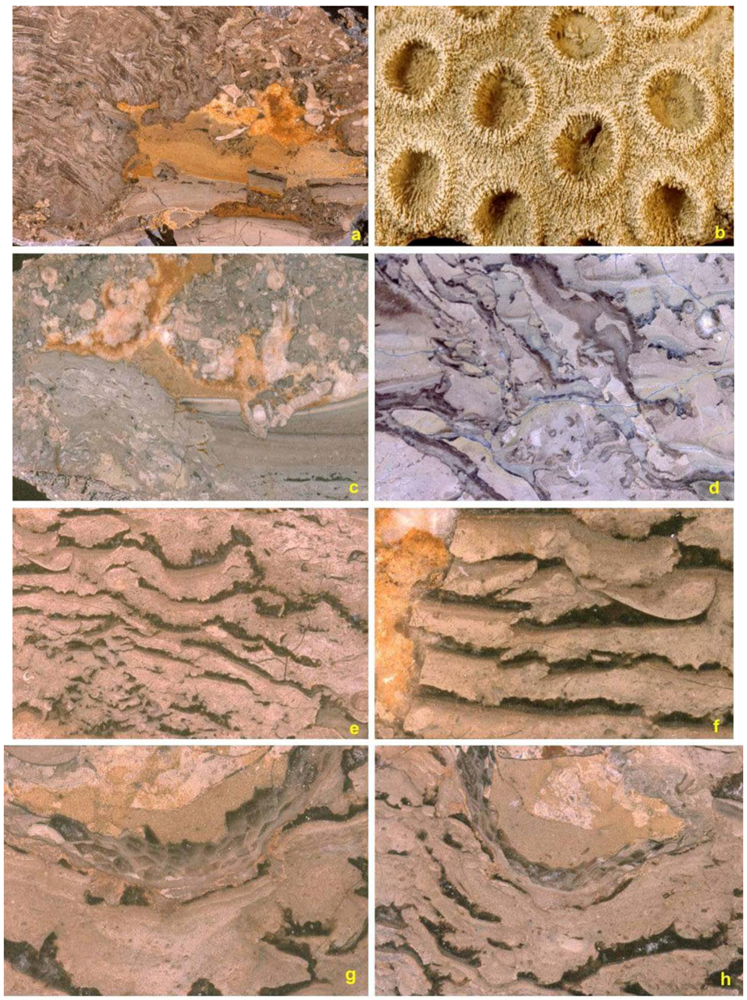
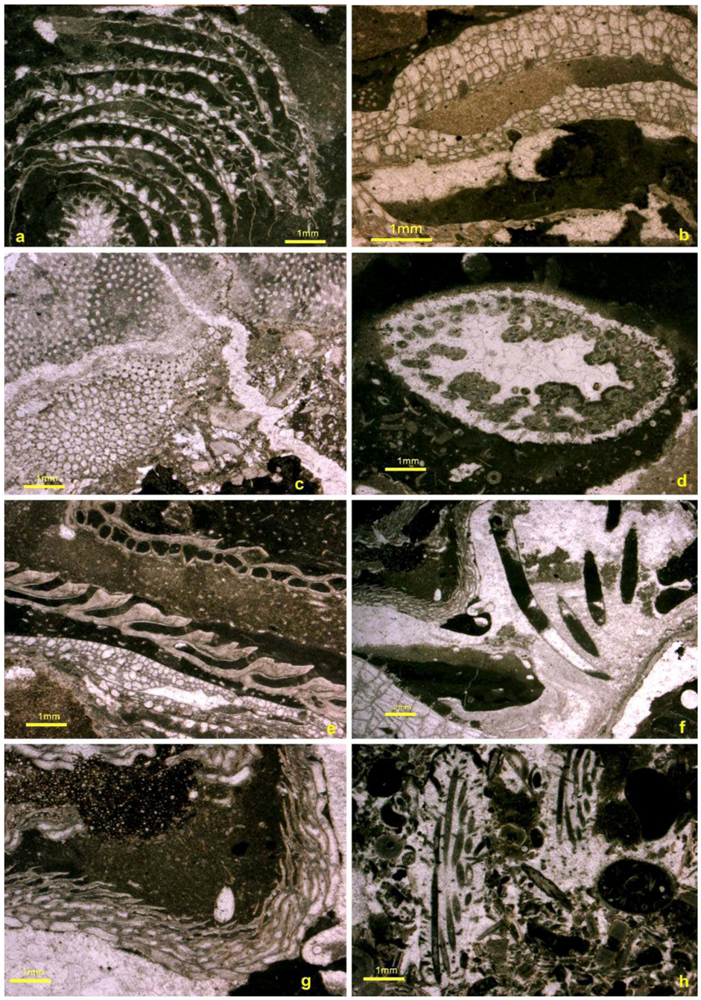
4. Reef Builders and Carbonate Re-Cyclers
5. Reef Constructors
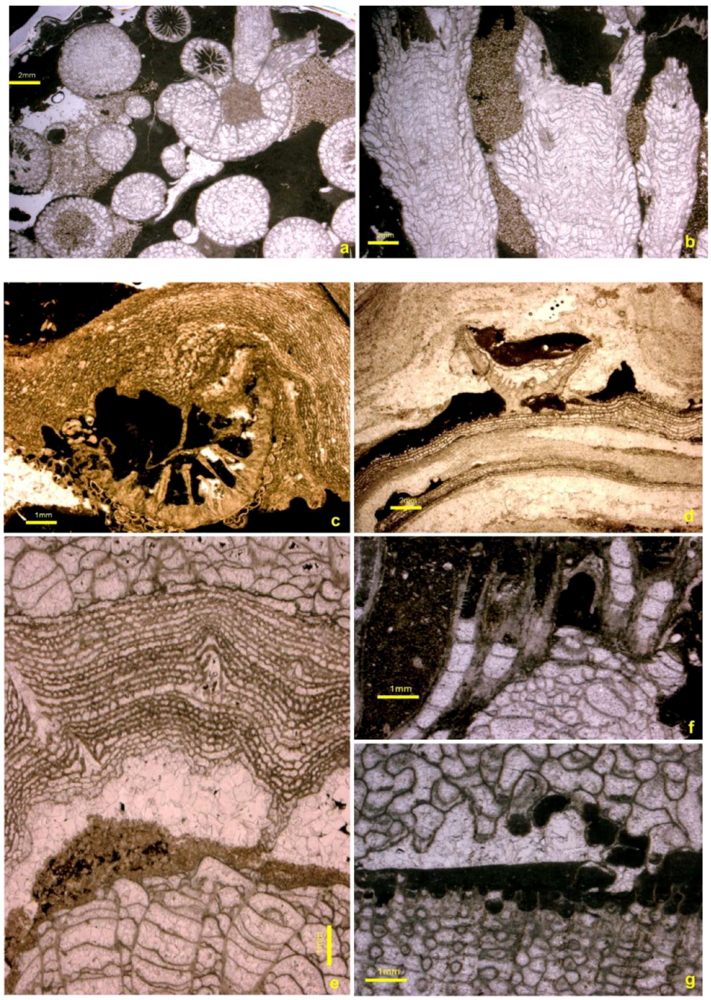
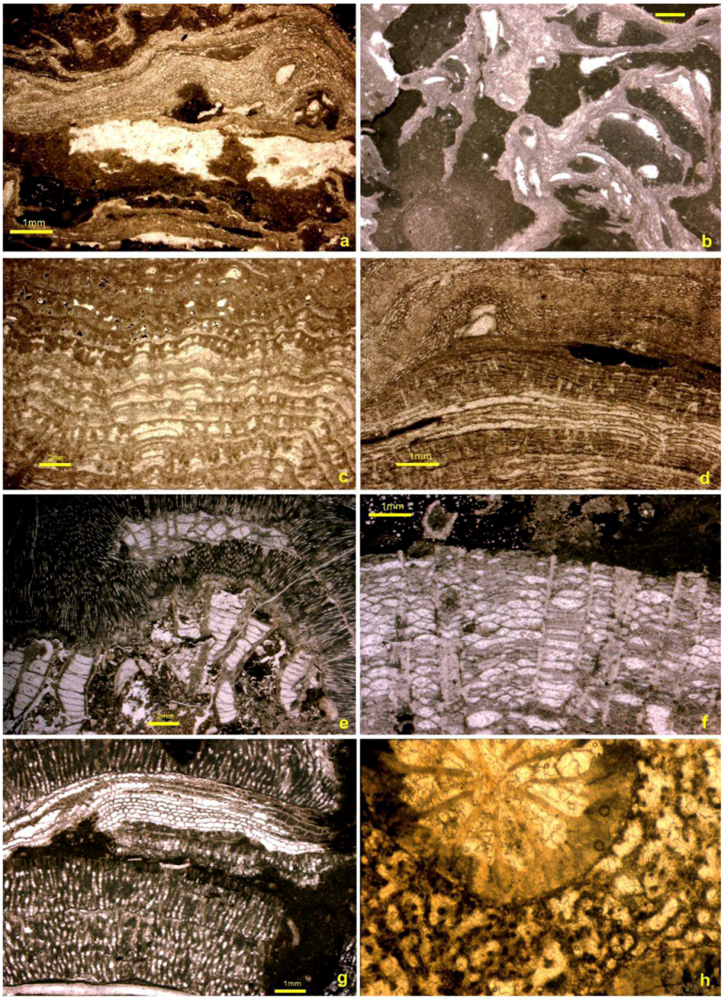
5.1. Corals

5.2. Stromatoporoids
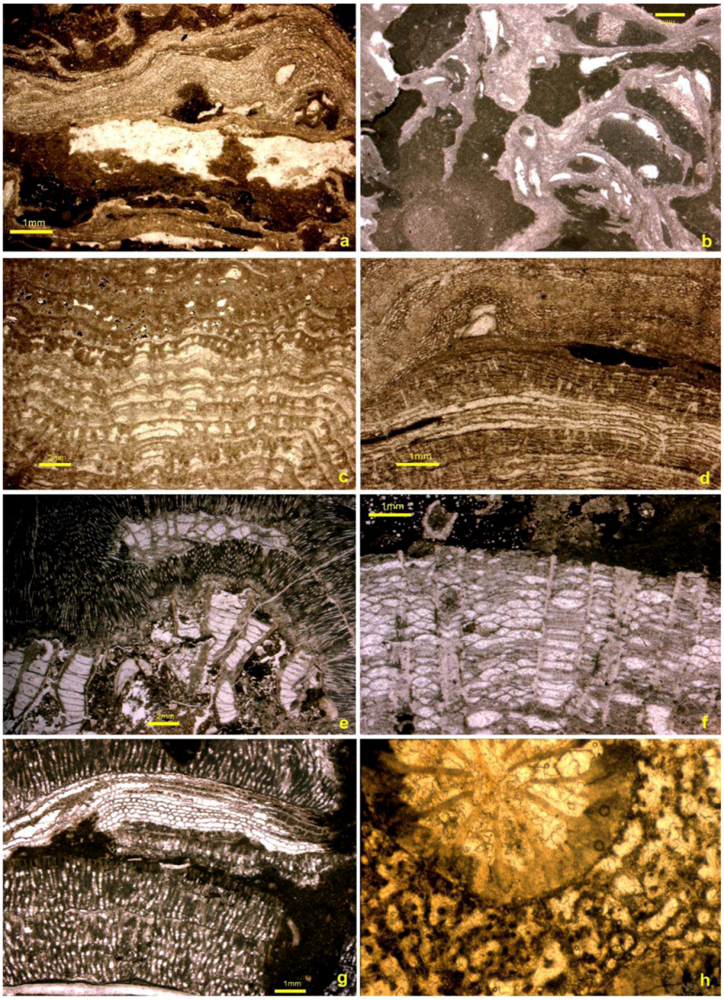
5.3. Cavity Structures and “Stromatactis”
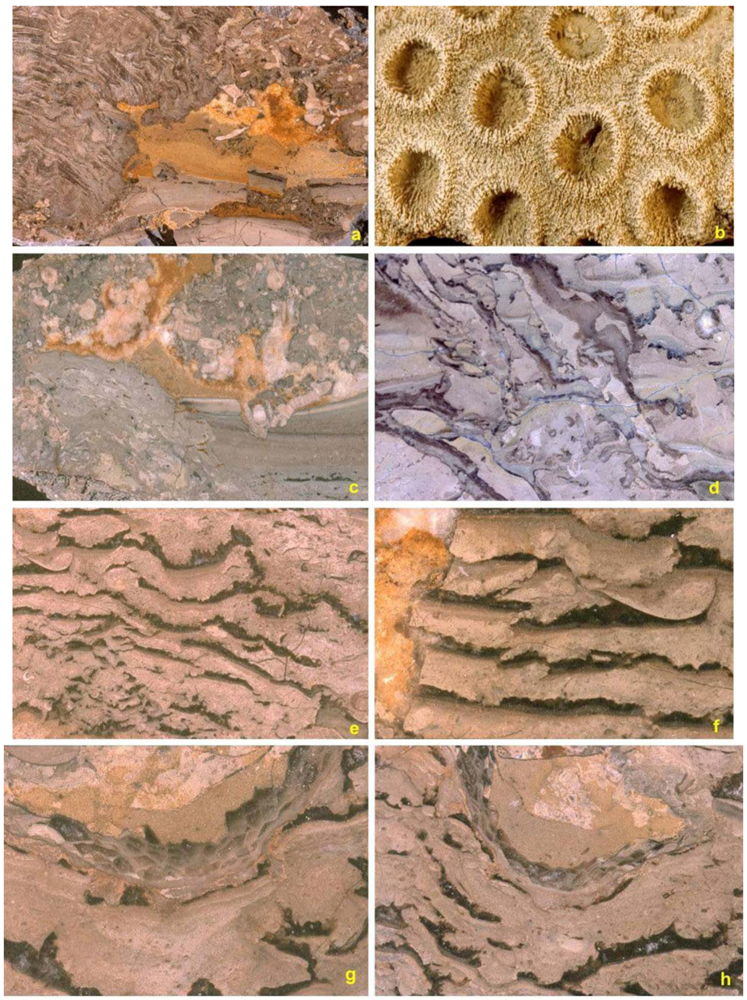
5.4. Binding and Encrusting Microbial Biota
5.5. Bafflers

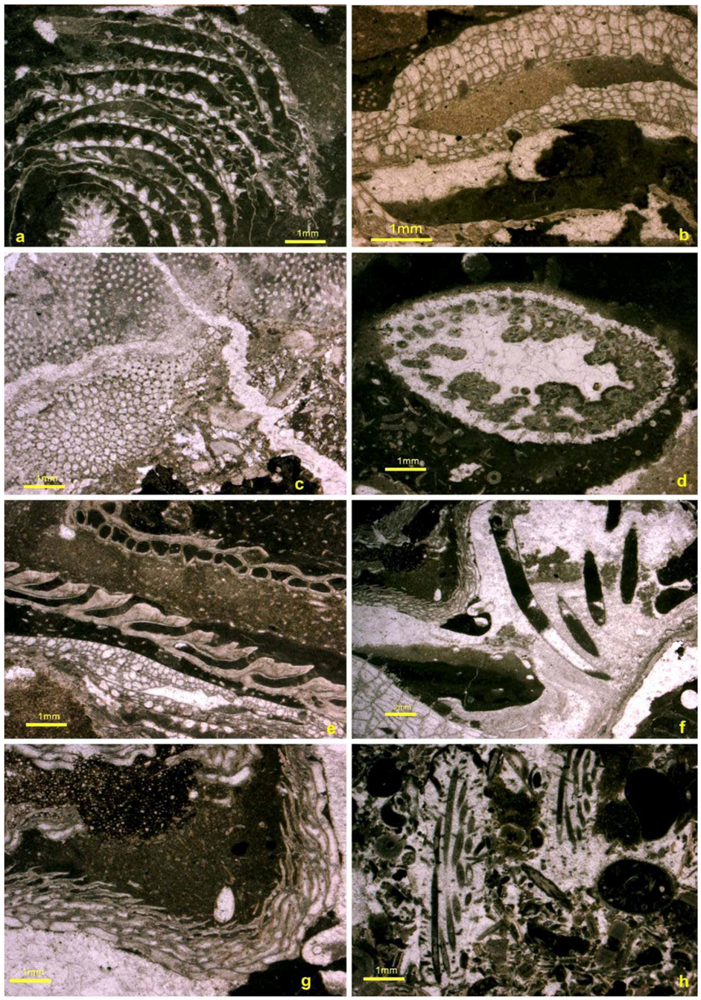
5.6. Reef Dwellers
6. Conclusions: What Triggered Reef Growth?
Acknowledgments
Appendix: Outcrops of East Point Member, Menier Formation
References
- Ghienne, J.F. The Late Ordovician glacial record: State of the art. In Ordovician of the World; Gutiérrez-Marco, J.C., Rábano, I., García-Bellido, D., Eds.; Cuadernos del Museo Geominero, Instituto Geológico y Minero de España: Madrid, Spain, 2011; Volume 14, pp. 13–19. [Google Scholar]
- Caputo, M.V. Ordovician-Silurian glaciations and global sea-level changes. N.Y. State Mus. Bull. 1998, 491, 15–25. [Google Scholar]
- Diaz-Martinez, E.; Vavrdova, M.; Isaacson, E.; Grahn, C.Y. Early Silurian vs. Late Ordovician glaciation in South America. In Ordovician of the World; Gutiérrez-Marco, J.C., Rábano, I., García-Bellido, D., Eds.; Cuadernos del Museo Geominero, Instituto Geológico y Minero de España: Madrid, Spain, 2011; Volume 14, pp. 127–134. [Google Scholar]
- Copper, P. 100 million years of reef prosperity and collapse: Ordovician to Devonian interval. In Corals and Reefs: Crises, Collapse and Change; Stanley, G.D., Ed.; Volume 17, pp. 15–32, The Paleontological Society Papers.
- Copper, P. Upper Ordovician and Lower Silurian reefs of Anticosti Island, Quebec. In Reefs, Canada and Adjacent Areas; Geldsetzer, H.H.J., James, N.P., Tebbutt, G.E., Eds.; Memoirs Canadian Society of Petroleum Geologists: Alberta, Canada, 1989; Volume 13, pp. 271–276. [Google Scholar]
- Copper, P. Reefs during the multiple crises towards the Ordovician-Silurian boundary: Anticosti Island, eastern Canada, and worldwide. Can. J. Earth Sci. 2001, 38, 153–171. [Google Scholar] [CrossRef]
- Nestor, H.; Copper, P.; Stock, C. Late Ordovician and Early Silurian Stromatoporoid Sponges from Anticosti Island, Eastern Canada: Crossing the O/S Mass Extinction Boundary; NRC Research Press: Ottawa, Canada, 2010; pp. 1–163. [Google Scholar]
- Copper, P.; Brunton, F.R. A global review of Silurian reefs. Spec. Pap. Palaeont. 1991, 44, 225–259. [Google Scholar]
- Brunton, F.R.; Copper, P.; Dixon, O.A. Silurian reef-building episodes. In Proceedings of the 8th International Coral Reef Symposium, Panama City, Panama, June 1996; 2, pp. 1643–1650.
- Copper, P. Silurian and Devonian reefs: 80 million years of global greenhouse between two ice ages. In Phanerozoic Reef Patterns; Kiessling, W., Flügel, E., Golonka, J., Eds.; SEPM: Tulsa, OK, USA, 2002; Volume 72, pp. 181–238. [Google Scholar]
- Copper, P.; Fay, I. An Early Silurian reef complex, Manitoulin Island, northern Ontario. Mem. Can. Soc. Petrol. Geol. 1989, 13, 277–282. [Google Scholar]
- Bergström, S.M.; Kleffner, M.; Schmitz, B.; Cramer, B.D. Revision of the position of the Ordovician-Silurian boundary in southern Ontario: Regional chronostratigraphic implications of δ13C chemostratigraphy of the Manitoulin Formation and associated strata. Can. J. Earth Sci. 2011, 48, 1447–1470. [Google Scholar] [CrossRef]
- Aaloe, A.O.; Nestor, H.E. Biogermnaya fatsiya v Yuruskom Gorizonte (nizhnii Llandoveri) v severo-zapadnoi Estonii [Biohermal facies in the Juuru Stage (lower Llandovery) in northwest Estonia]. In Facies and Fauna of the Baltic Silurian; Kaljo, D., Ed.; Akademiya Nauk Estonskoi SSR, Institut Geologii: Tallin, Estonia, 1977; pp. 71–88. [Google Scholar]
- Nestor, H. Silurian. In Geology and Mineral Resources of Estonia; Raukas, A., Teedumäe, A., Eds.; Estonian Academy Publishers: Tallinn, Estonia, 1997; pp. 89–106. [Google Scholar]
- Klaamann, E.R. Inkommunikatnye tabulyaty Estonii [The incommunicate Tabulata of Estonia]; Eesti NSV Teaduste Akadeemia Geoloogia Instituut: Tallinn, Estonia, 1966; p. 97. [Google Scholar]
- Kaljo, D. Silur Estonii [Silurian of Estonia]; Valgus: Tallinn, Estonia, 1970; p. 343. [Google Scholar]
- Bolton, T.E. Geological map and notes on the Ordovician and Silurian litho- and biostratigraphy, Anticosti Island, Quebec. Geol. Surv. Can. 1972, 71(19), 1–45. [Google Scholar]
- Bolton, T.E. Late Ordovician and early Silurian Anthozoa of Anticosti Island, Québec. In Proceedings of IUGS Subcommission on Silurian Stratigraphy, Ordovician-Silurian Boundary Working Group, Anticosti-Gaspé, Québec, Canada, February 1981. In Stratigraphy and Paleontology; Lespérance, P.J. (Ed.) Université de Montréal: Montréal, Canada, 1981; pp. 107–135.
- Kaljo, D. Fatsii I fauna Silura Pribaltiki [Facies and fauna of the Silurian of the Prebaltic]; Akademiya Nauk Estonskoi SSR, Institut Geologii: Tallinn, Estonia, 1977; p. 286. [Google Scholar]
- Richardson, J. Report for the year 1856, of Mr. James Richardson, explorer, addressed to Sir William E. Logan, Provincial Geologist. Report of Progress for the Years 1853-54-55-56, John Lovell: Toronto, Canada, 1857; 191–245. [Google Scholar]
- Schuchert, C.; Twenhofel, W.H. Ordovicic-Siluric section of the Mingan and Anticosti islands, Gulf of Saint Lawrence. Bull. Geol. Soc. Am. 1910, 21, 677–716. [Google Scholar]
- Twenhofel, W.H. Faunal and sediment variation in the Anticosti sequence. Geol. Surv. Can. Bull. 1921, 33, 1–14. [Google Scholar]
- Twenhofel, W.H. Geology of Anticosti Island. Geol. Surv. Can. Mem. 1928, 154, 1–481. [Google Scholar]
- Petryk, A.A. Ile d’Anticosti, Geologic Map, Ministère de l’Energie et des Ressources, Québec, DPV 823 (three map sheets 1:100,000); Ministère de l'Energie et des Ressources: Québec, Canada, 1979. [Google Scholar]
- Petryk, A.A. Stratigraphy, sedimentology and paleogeography of the Upper Ordovician–Lower Silurian of Anticosti Island, Québec. In. In Proceedings of IUGS Subcommission on Silurian Stratigraphy, Ordovician-Silurian Boundary Working Group, Anticosti-Gaspé, Québec, Canada, February 1981. In Stratigraphy and Paleontology; Lespérance, P.J. (Ed.) Université de Montréal: Montréal, Canada, 1981; pp. 11–39.
- Copper, P.; Long, D.G.F. Stratigraphic revision of the Jupiter Formation, Anticosti Island, Canada: A major reference section above the Ordovician-Silurian boundary. Newsl. Stratigr. 1990, 23, 11–36. [Google Scholar]
- Copper, P. Articulate brachiopod shellbeds: Silurian examples from Anticosti, eastern Canada. Geobios Mém. Spéc. 1997, 20, 133–148. [Google Scholar] [CrossRef]
- Jin, J. Environmental control on temporal and spatial differentiation of Early Silurian pentameride brachiopod communities, Anticosti Island, eastern Canada. Can. J. Earth Sci. 2008, 45, 159–187. [Google Scholar] [CrossRef]
- McLean, R.A.; Copper, P. The early Silurian (late Rhuddanian-Telychian) rugose coral fauna of Anticosti Island, eastern Canada: Diversity during the post O/S mass extinction recovery interval. Palaeontogr. Can. 2012, in press. [Google Scholar]
- Long, D.G.F.; Copper, P. The Late Ordovician-Early Silurian Carbonate Tract of Anticosti Island, Gulf of St. Lawrence, Eastern Canada; Field Trip B4 Guidebook; Geological Association of Canada: Newfoundland, Canada, 1994; pp. 1–69. [Google Scholar]
- Copper, P.; Long, D.G.F. Sedimentology and paleontology of the Late Ordovician through Early Silurian shallow water carbonates and reefs of the Anticosti Island, Québec. In Sedimentology and Paleontology of the Late Ordovician through Early Silurian Shallow Water Carbonates of the Mingan Islands National Park and Anticosti Island, Québec; Field Trip B8 Guidebook; Desrochers, A., Copper, P., Long, D.G.F., Eds.; Geological Association of Canada-Mineralogical Association of Canada: Quebec, Canada, 1998; pp. 55–97. [Google Scholar]
- Spalding, M.D.; Ravilious, C.; Green, E.P. World Atlas of Coral Reefs; University of California Press: Berkeley, CA, USA, 2001; p. 424. [Google Scholar]
- Desrochers, A. Rocky shoreline deposits in the Lower Silurian (upper Llandovery) Chicotte Formation, Anticosti Island, Québec. Can. J. Earth Sci. 2006, 43, 205–1214. [Google Scholar] [CrossRef]
- Desrochers, A.; Bourque, P.A.; Neuweiler, F. Diagenetic versus biotic accretionary mechanisms of bryozoan-sponge buildups (Lower Silurian, Anticosti Island, Canada). J. Sediment. Res. 2007, 77, 564–571. [Google Scholar] [CrossRef]
- Billings, E. New species of fossils from different parts of the Lower, Middle and Upper Silurian rocks of Canada. Geol. Surv. Can. 1862, 1(4), 96–168. [Google Scholar]
- Lindström, G. Remarks on the Heliolitidae. Handl. Köngl. Svensk. Vetenskaps-akad. 1899, 32, 1–140. [Google Scholar]
- Bourque, A.; Boulvain, F. A model for the origin and petrogenesis of the red stromatactis limestone of Paleozoic carbonate mounds. J. Sediment. Petrol. 1993, 63, 607–619. [Google Scholar]
- Ausich, W.I.; Copper, P. The Crinoidea of Anticosti Island, Québec (Late Ordovician to Early Silurian). Paleontogr. Can. 2010, 29, 157. [Google Scholar]
- Gauthier-Coulloudon, L.; Mamet, B.L. Algues et sedimentation carbonatés dans l’Ordovicien et la Silurien de l’île d’Anticosti. In. In Proceedings of IUGS Subcommission on Silurian Stratigraphy, Ordovician-Silurian Boundary Working Group, Anticosti-Gaspé, Québec, Canada, February 1981. In Stratigraphy and Paleontology; Lespérance, P.J. (Ed.) Université de Montréal: Montréal, Canada, 1981; pp. 87–90.
- Azmy, K.; Veizer, J.; Bassett, M.G.; Copper, P. Oxygen and carbon isotopic composition of Silurian brachiopods: Implications for coeval seawater and glaciations. Geol. Soc. Am. Bull. 1998, 110, 1499–1512. [Google Scholar]
- Munnecke, A.; Samtleben, C.; Bickert, T. The Ireviken Event in the lower Silurian of Gotland, Sweden—Relation to similar Palaeozoic and Proterozoic events. Palaeogeogr. Palaeocl. Palaeoecol. 2003, 195, 99–124. [Google Scholar] [CrossRef]
- Cramer, B.D.; Loydell, D.K.; Samtleben, C.; Munnecke, A.; Kaljo, D.; Männik, P.; Martma, T.; Jeppsson, L.; Kleffner, M.A.; Barrick, J.E.; et al. Testing the limits of Paleozoic chronostratigraphic correlation via high-resolution (<500 kyr) integrated conodont, graptolite, and carbon isotope (δ13C-carb) biochemostratigraphy across the Llandovery-Wenlock (Silurian) boundary: Is a unified Phanerozoic timescale achievable? Geol. Soc. Am. Bull. 2010, 122, 1700–1716. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Copper, P.; Jin, J. Early Silurian (Aeronian) East Point Coral Patch Reefs of Anticosti Island, Eastern Canada: First Reef Recovery from the Ordovician/Silurian Mass Extinction in Eastern Laurentia. Geosciences 2012, 2, 64-89. https://doi.org/10.3390/geosciences2020064
Copper P, Jin J. Early Silurian (Aeronian) East Point Coral Patch Reefs of Anticosti Island, Eastern Canada: First Reef Recovery from the Ordovician/Silurian Mass Extinction in Eastern Laurentia. Geosciences. 2012; 2(2):64-89. https://doi.org/10.3390/geosciences2020064
Chicago/Turabian StyleCopper, Paul, and Jisuo Jin. 2012. "Early Silurian (Aeronian) East Point Coral Patch Reefs of Anticosti Island, Eastern Canada: First Reef Recovery from the Ordovician/Silurian Mass Extinction in Eastern Laurentia" Geosciences 2, no. 2: 64-89. https://doi.org/10.3390/geosciences2020064