The Extinction of the Conulariids


Abstract
:1. Introduction
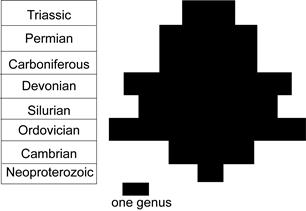
2. Conulariid Anatomy, Affinities, Fossil Record and Paleoecology
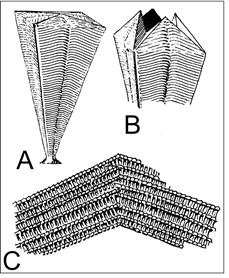
3. Cretaceous Conulariid

4. Triassic Conulariids

{kind=link}
{kind=link}
{kind=link}
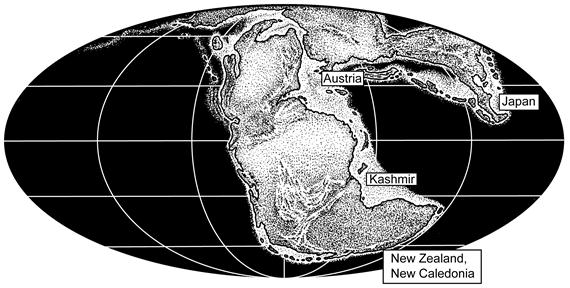{kind=link}
| Taxon | Location | Age | Publication | Reference |
|---|---|---|---|---|
| Paraconularia matauraensis | New Zealand | Rhaetian | Waterhouse (1979) | [18] |
| Paraconularia matauraensis | New Caledonia | Rhaetian | Hagdorn & Campbell (1993) | [37] |
| Conularia stromeri | Austria | Rhaetian | Osswald (1918) | [30] |
| Flectoconularia abapertura | New Zealand | Carnian | Waterhouse (1979) | [18] |
| Indeterminate conulariid | New Zealand | Carnian | Waterhouse (1979) | [18] |
| Conularia triadica | Austria | Carnian | Bittner (1890) | [29] |
| Conularia sp. | Kashmir | Anisian-Ladinian | Diener (1913) | [33] |
| Conulariopsis quadrata | Japan | Induan | Sugiyama (1942) | [36] |

5. Conulariid Extinction
Acknowledgements
References
- Marques, A.C.; van Iten, H.; Rodrigues, S.C.; Leme, J.M.; de Simões, M.G. Major developments in conulariid research: Problems of interpretation and future perspectives. Ameghiniana 2008, 45, 407–420. [Google Scholar]
- Chapman, M.G.; Lucas, S.G.; Tanner, L.H. Assessing the record and causes of Late Triassic extinctions. Earth-Sci. Rev. 2004, 65, 103–139. [Google Scholar] [CrossRef]
- Sepkoski, J.J., Jr.; Nudds, J.R. Coelenterata. In The Fossil Record; Benton, M.J. Ed., Ed.; 1993; Chapman and Hall: London,UK. [Google Scholar]
- Willard, B. The Harvey Bassler Collection of Peruvian Fossils; 1966; p. 255. Lehigh University: Bethlehem,PA,USA. [Google Scholar]
- Willard, B.A. Cretaceous conularid from Peru. Penn. Acad. Sci. 1967, 40, 87–89. [Google Scholar]
- Moore, R.C., Lalicker; Fischer, A.G., Lalicker. Invertebrate Fossils; 1952; p. 766. McGraw-Hill Book Company,Inc.: New York,NY,USA. [Google Scholar]
- Collins, A.G.; Marques, A.C.; Simõess, M.G.; Leme, J.; Van Iten, H. Reassessment of the phylogenetic position of conulariids (?Ediacaran-Triassic) within the subphylum Medusozoa (phylum Cnidaria). J. Syst. Palaeontol. 2006, 4, 109–118. [Google Scholar] [CrossRef]
- Simões, M.G.; de Rodrigues, S.C.; Leme, J.M.; Van Iten, H. Reinterpretation of a conulariid-like fossil from the Vendian of Russia. Palaeont. 2005, 48, 619–622. [Google Scholar] [CrossRef]
- Vyrhlasová, Z.; Van Iten, H. Conulariids. In The Great Ordovician Biodiversification Event; Webby, B.D., Paris, Ed.; 2004; Columbia Universitry Press: New York,NY,USA. [Google Scholar]
- Van Iten, H. Written communication., Hanover, IN, USA, 2012.
- Harrington, H.J.; Moore, R.C. Conulata. In Treatise on Invertebrate paleontology Part F Coelenterata; Ed. Moore, R.C., Ed. Geological Society of America: Boulder, CO, Eds.; 1956; pp. 0–0. University of Kansas Press: Lawrence,KS,USA. [Google Scholar]
- Feldman, R.M.; Babcock, L.E. Mysterious fossils. Earth Sci. 1984, 37, 16–17. [Google Scholar]
- Finks, R.M. J. Paleont ol. 1955, 29, 831–836.
- Pickerill, R.K.; Harland, T.L. Epizoic Schizocrania sp. from the Ordovician Trenton Group of Quebec, with comments on mode of life of conulariids. J. Paleontol. 1987, 61, 844–849. [Google Scholar]
- Van Iten, H. J. Paleontol. 1991, 65, 335–337.
- Brood, K. Morphology, structure, and systematics of the conulariids. GFF 1995, 117, 121–137. [Google Scholar] [CrossRef]
- de M Leme, J.; Marques, A.C.; de Rodrigues, S.C.; Mello, L.H.C.; Simões, M.G. Conulariid taphonomy as a tool in paleoenvironmental analysis. Rev. Bras. Geocienc. 2000, 30, 757–762. [Google Scholar]
- Waterhouse, J.B. Permian and Triassic conulariid species from New Zealand. J. Roy. Soc. New. Zeal. 1979, 9, 475–489. [Google Scholar] [CrossRef]
- Babcock, L.E. A new Ordovician conulariid from Oklahoma? Okla. Geol. Notes 1985, 45, 66–70. [Google Scholar]
- Siegele, P.K.; Himes, G.T.; Boucot, A.J.; Gray, J.; Babcock, L.E. First Silurian conulariids from Paraguay. J. Paleont. 1990, 64, 897–902. [Google Scholar]
- Benavides-Cáceres, V.E. Cretaceous system in Northern Peru. B. Am. Mus. Nat.Hist. 1956, 108, 353–494. [Google Scholar]
- Jaillard, E.; Dhondt, A.V. Cretaceous bivalves from Ecuador and northern Peru. J. South Amer. Earth Sci. 2005, 19, 325–342. [Google Scholar] [CrossRef]
- Dhondt, A.V.; Bengtson, P.; Jaillard, E. Late Cretaceous marine transgressions in Ecuador and northern Peru: A preliminary stratigraphic framework. J. S. Am. Earth Sci. 2005, 19, 307–323. [Google Scholar] [CrossRef]
- Hertlein, L.G.; Haas, F.; Fleming, C.A.; Dechaseaux, C.; Coogan, A.H.; Chavan, A.; Casey, R.; Branson, C.C.; Newell, N.D.; Cox, L.R.; et al. Systematic descriptions. In Treatise on Invertebrate Paleontology; Moore, R.C., Ed.; 1969; Volume 1, pp. 0–0. University of Kansas: Lawrence,KS,USA. [Google Scholar]
- Durney, K.; Spielmann, J.A.; Krainer, K.; Lucas, S.G. Cretaceous stratigraphy, paleontology, petrography, depositional environments, and cycle stratigraphy at Cerro de Cristo Rey, Doña Ana County, New Mexico. New Mex. Geol. 2010, 32, 103–130. [Google Scholar]
- Thomas, G.A. Notoconularia, a new conularid genus from the Permian of Eastern Australia. J. Paleontol. 1969, 43, 1283–1290. [Google Scholar]
- Argéliez, M. Letter to Élie de Beaumont. Soc. Géol. Fr. Bull. 1856, 13, 186–188. [Google Scholar]
- Sandberger, G. Die Flossenfüsser der Pteropoda der ersten Erdbildungs-Epoche. ConulariaColeoprion. Neues Jahrb. Mineral Jahrg 1847, 8–25. [Google Scholar]
- Bittner, A. Eine Triadische Conularia. Kaiserl.-königl. Geol. Reichanstalt Verhand. 1890, 9, 177–178. [Google Scholar]
- Osswald, K. Mesozoische Conulariiden. Cent. Mineral. 1981, 21, 337–344. [Google Scholar]
- Richardson, E.S.; Sinclair, G.W. A bibliography of the Conularida. B. Am. Paleontol. 1954, 34, 1–143. [Google Scholar]
- Bittner, A. Conularia in der Trias. Kaiserl.-königl. Geol. Reichanstalt Verhand. 1878, 12, 281–282. [Google Scholar]
- Diener, C. Triassic faunas of Kashmir. Geol. Surv. India Mem. Palaeontol. Indic. New Ser. 1913, 5, 1–133. [Google Scholar]
- Nicora, A.; Balini, M.; Krystyn, L. Lower and Middle Triassic stage and substage boundaries in Spiti. Albertiana 2004, 30, 40–53. [Google Scholar]
- Trechmann, C.T. The Trias of New Zealand. Q. J. Geol. Soc. 1918, 73, 165–246. [Google Scholar]
- Sugiyama, T. Studies on the Japanese Conularida. Geol. Soc. Jpn. 1942, 49, 390–399. [Google Scholar] [CrossRef]
- Campbell, H.J.; Hagdorn, H. Paracomatula triadica sp. nov.—An early comatulid crinoid from the Otapirian (Late Triassic) of New Caledonia. Alcheringa 1990, 17, 1–17. [Google Scholar]
- Kozur, H. Written communication. Budapest, Hungary, 2012. [Google Scholar]
- Shi, G.R.; Weldon, E.A. Global distribution of Permian conulariids and palaeobiogeographical implications. Gondwana Res. 2003, 6, 791–802. [Google Scholar] [CrossRef]
- Vermeij, G.J. The Mesozoic marine revolution: Evidence from snails, predators and grazers. Paleobiology 1977, 3, 245–258. [Google Scholar]
- Vermeij, G.J. Evolution and Escalation; 1983; p. 527. Princeton University Press: Princeton,NJ,USA. [Google Scholar]
- Jablonski, D.; Fürsich, F.T. Late Triassic naticid drillholes: Carnivorous gastropods gain a major adaptation but fail to radiate. Science 1984, 224, 78–80. [Google Scholar] [CrossRef] [PubMed]
- Skelton, P.W.; Harper, E.M. The Mesozoic marine revolution and epifaunal bivalves; 1993; In Proceedings of Symposium ‘Molluscan Palaeontology’,11th International Malacological Congress,Siena,Italy,30 August–5 September 1992; National Museum of Natural History: Leiden,The Netherlands. [Google Scholar]
- Palmer, T.; Forsythe, T.W.; Harper, E.M. Taphonomy and the Mesozoic marine revolution: Preservation state masks the importance of boring predators. Palaios 1998, 13, 352–360. [Google Scholar] [CrossRef]
- Nützel, A. The Late Triassic species Cryptaulax? bittneri (Mollucsa: Gastropoda: Procerithiidae) and remarks on some aspects of the early Mesozoic marine revolution. Paleontol. 2002, 76, 57–63. [Google Scholar]
- Hautmann, M. Early Mesozoic evolution of articular bivalve ligaments and its implications for the timing of the 'Mesozoic marine revolution'. Lethaia 2004, 37, 165–172. [Google Scholar] [CrossRef]
- Tanner, L.H.; Lucas, S.G. Reexamination of the end-Triassic mass extinction. In Mass Extinction; Elewa, A.M., Ed., Ed.; 2008; Springer-Verlag: Berlin,Germany. [Google Scholar]
- Wilson, M.T.; Feldmann, R.M.; Babcock, L.E. Teratology and pathology of some Paleozoic conulariids. Lethaia 1987, 20, 93–105. [Google Scholar] [CrossRef]
- Babcock, L.E.; Fahrer, T.R.; Mapes, R.H. Sublethal and lethal injuries of Pennsylvanian conulariids from Oklahoma. J. Paleontol. 1989, 63, 34–37. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lucas, S.G. The Extinction of the Conulariids. Geosciences 2012, 2, 1-10. https://doi.org/10.3390/geosciences2010001
Lucas SG. The Extinction of the Conulariids. Geosciences. 2012; 2(1):1-10. https://doi.org/10.3390/geosciences2010001
Chicago/Turabian StyleLucas, Spencer G. 2012. "The Extinction of the Conulariids" Geosciences 2, no. 1: 1-10. https://doi.org/10.3390/geosciences2010001
APA StyleLucas, S. G. (2012). The Extinction of the Conulariids. Geosciences, 2(1), 1-10. https://doi.org/10.3390/geosciences2010001