
Life History Traits and Longevity of the Invasive Asian Common Toad Duttaphrynus melanostictus (Schneider, 1799) in Madagascar

 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
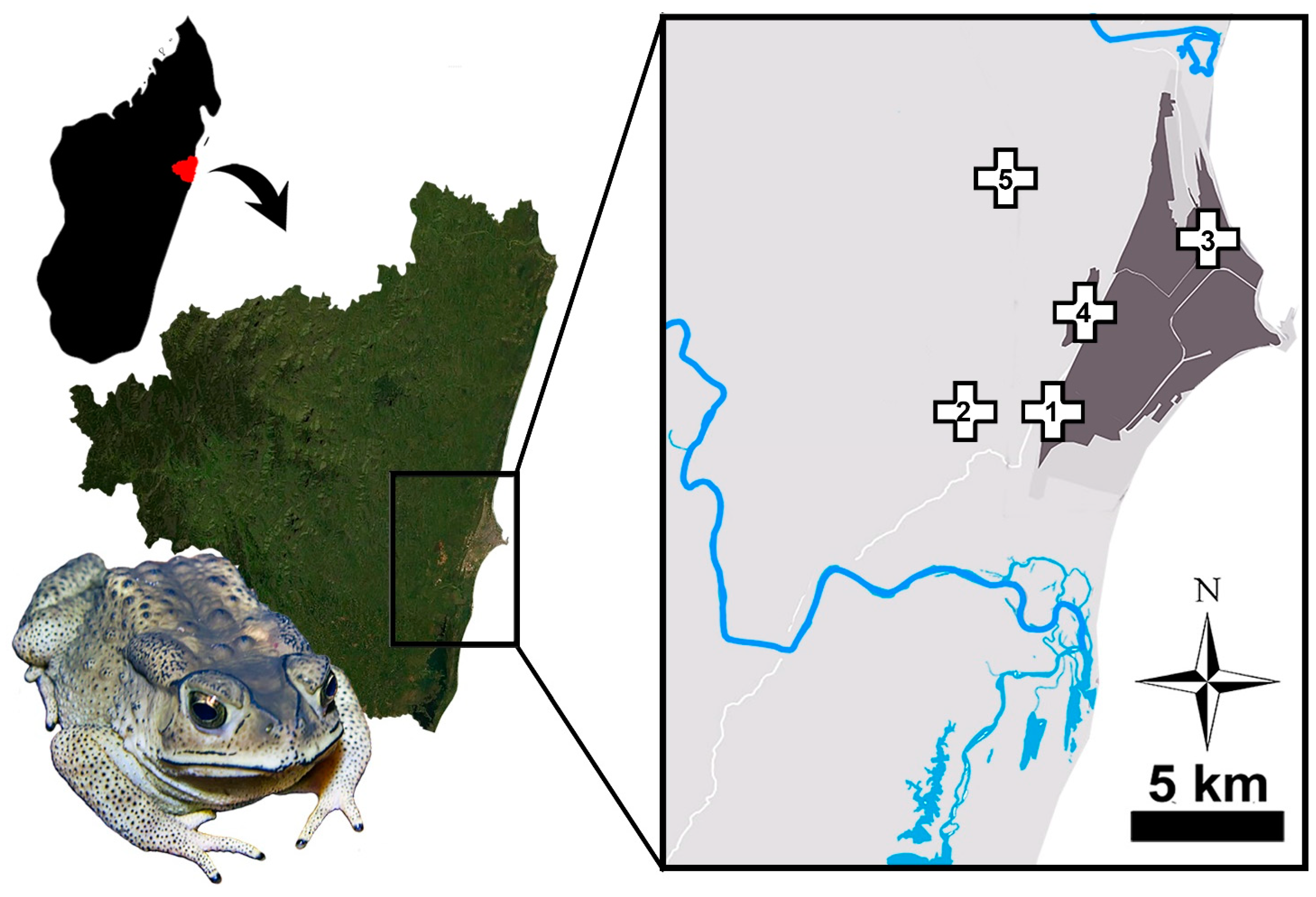
2.1. Sampling
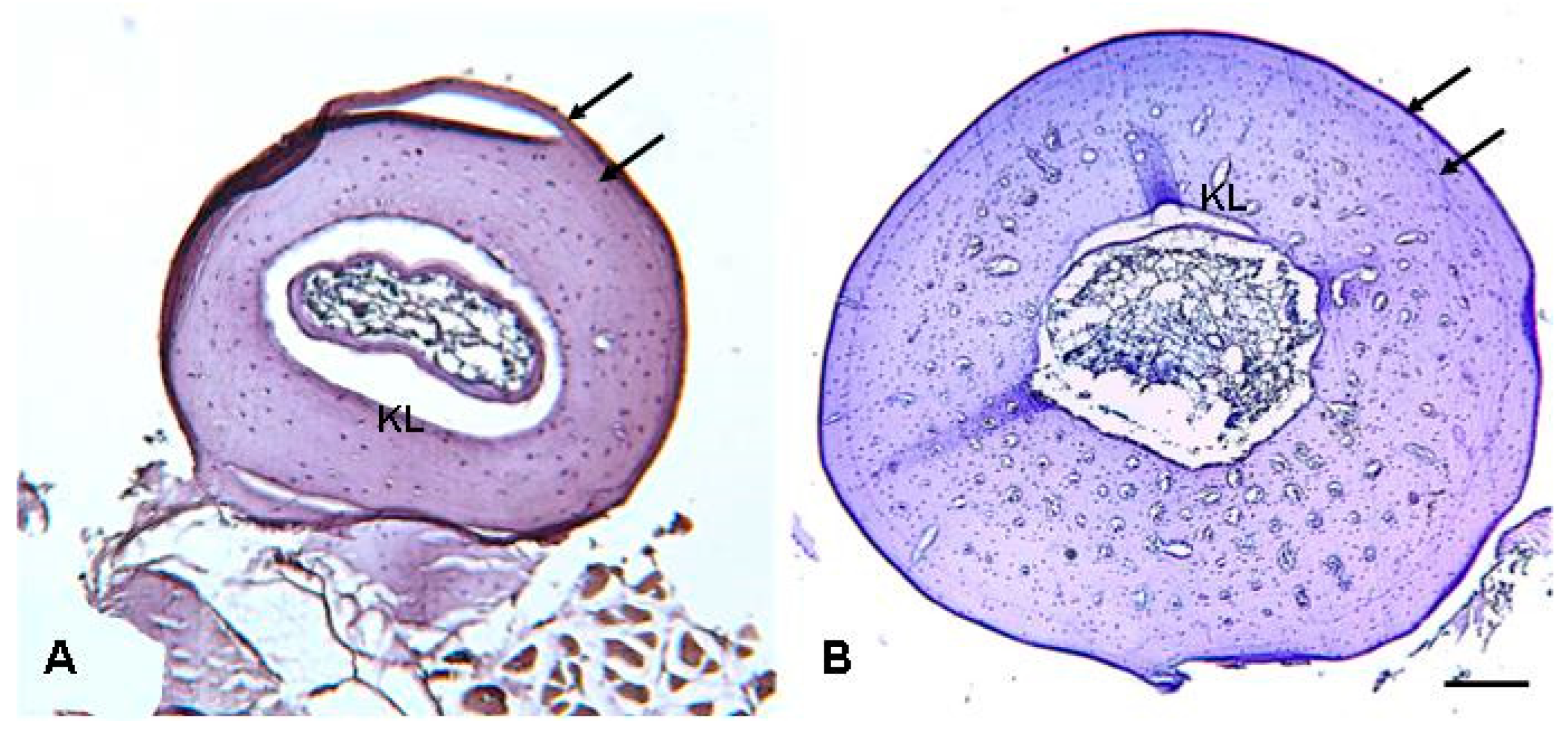
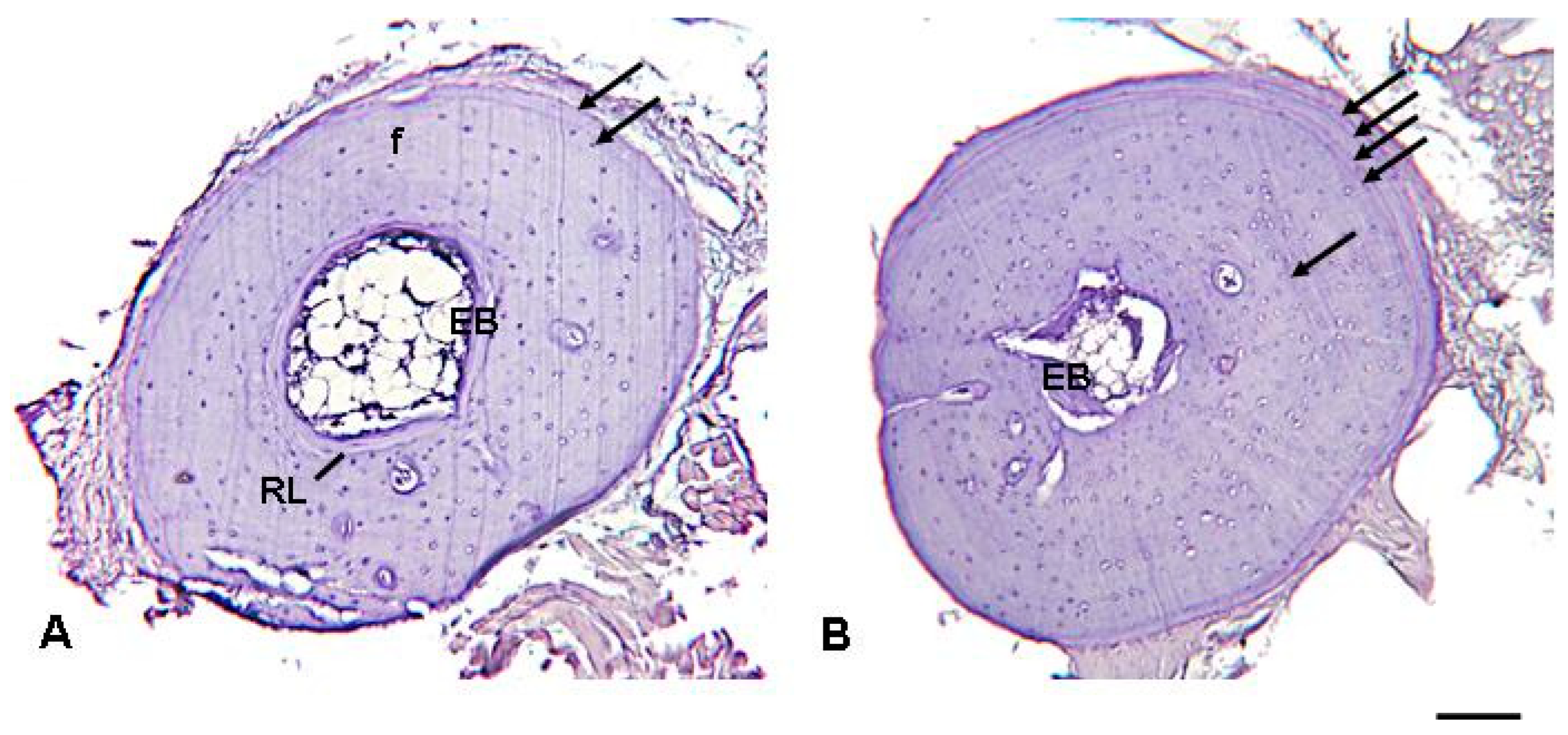
2.2. Skeletochronology
2.3. Sexual Maturity
2.4. Statistical Analysis
3. Results
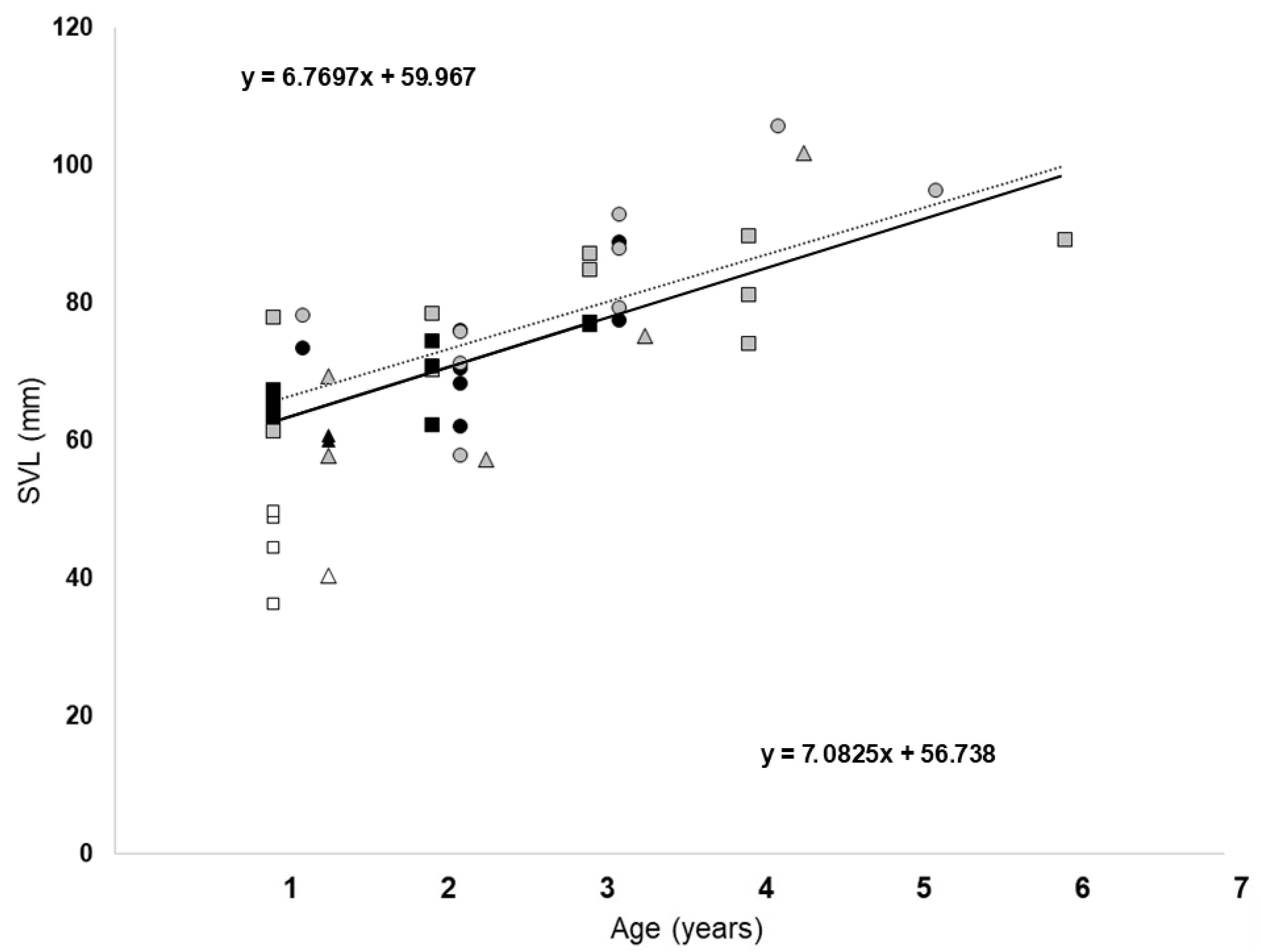
3.1. Body Length
3.2. Skeletochronological Observations
3.3. Age
3.4. Assessment of Gonad Status and Individual Age
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Measey, A.; Vimercati, G.; de Villiers, F.A.; Mokhatla, M.; Davies, S.J.; Thorp, G.J.; Rebelo, A.D.; Kumschick, S. Global assessment of alien amphibian impacts in a formal framework. Divers. Distrib. 2016, 22, 970–981. [Google Scholar] [CrossRef]
- Mezzasalma, M.; Visone, V.; Petraccioli, A.; Odierna, G.; Capriglione, T.; Guarino, F.M. Non-random accumulation of LINE1-like sequences on differentiated snake W chromosomes. J. Zool. 2016, 300, 67–75. [Google Scholar] [CrossRef]
- Mezzasalma, M.; Andreone, F.; Odierna, G.; Guarino, F.M.; Crottini, A. Comparative cytogenetics on eight Malagasy Mantellinae (Anura, Mantellidae) and a synthesis of the karyological data on the subfamily. Comp. Cytogen. 2022, 16, 11–17. [Google Scholar] [CrossRef]
- Sidhom, M.; Said, K.; Chatti, N.; Guarino, F.M.; Odierna, G.; Petraccioli, A.; Picariello, O.; Mezzasalma, M. Karyological characterization of the common chameleon (Chamaeleo chamaeleon) provides insights on the evolution and diversification of sex chromosomes in Chamaeleonidae. Zoology 2020, 141, 125738. [Google Scholar] [CrossRef]
- Andreone, F.; Carpenter, A.; Crottini, A.; D’Cruze, N.; Dubos, N.; Edmonds, D.; Garcia, G.; Luedtke, J.; Megson, S.; Rabemananjara, F.; et al. Chapter 4. Amphibian conservation in Madagascar: Old and novel threats for a peculiar fauna. In Status and Threats of Afrotropical Amphibians. Sub-Saharian Africa, Madagascar, Western Indian Ocean Islands; Heatwole, H., Roedel, M., Eds.; Chimaira: Frankfurt, Germany, 2021; Volume II, pp. 147–186. [Google Scholar]
- Andreone, F.; Rabibisoa, N.; Randrianantoandro, C.; Crottini, A.; Edmonds, D.; Kraus, F.; Lewis, J.P.; Moore, M.; Rabemananjara, F.C.E.; Rabemanantoa, J.C.; et al. Risk review is under way for invasive toad. Nature 2014, 512, 253. [Google Scholar] [CrossRef] [Green Version]
- Crottini, A.; Andreone, F.; Edmonds, D.; Hansen, C.M.; Kraus, F.; Lewis, J.P.; Moore, M.; Rabemananjara, F.; Rabemanantsoa, J.C.; Randrianantoandro, C.; et al. A new challenge for amphibian conservation in Madagascar: The invasion of Duttaphrynus melanostictus in Toamasina province. FrogLog 2014, 22, 46–47. [Google Scholar]
- Kolby, J.E.; Kraus, F.; Rabemananjara, F.; Rabesihanaka, S.; Rabibisoa, N.; Rafanomezantsoa, J. Stop Madagascar’s toad invasion now. Nature 2014, 509, 563. [Google Scholar] [CrossRef] [Green Version]
- Moore, M.; Solofo Niaina Fidy, J.F.; Edmonds, D. The new toad in town: Distribution of the Asian toad, Duttaphrynus melanostictus, in the Toamasina area of eastern Madagascar. Trop. Conserv. Sci. 2015, 8, 440–455. [Google Scholar] [CrossRef]
- Freeman, K.L.M.; Andreone, F.; Randriamoria, T.M.; Randrianizahana, H.; Raselimanana, A.P.; Raxworthy, C.J.; Robsomanitrandrasana, E.; Crottini, A. Bufonidae: Duttaphrynus melanostictus, Black-spined toad. In The new natural history of Madagascar; Goodman, S.M., Ed.; Princeton University Press: Princeton, NJ, USA, 2022; pp. 1404–1410. [Google Scholar] [CrossRef]
- Licata, F.; Andreone, F.; Crottini, A.; Harison, R.F.; Ficetola, G.F. Does spatial sorting occur in the invasive Asian toad in Madagascar? Insights into the invasion unveiled by morphological analyses. J. Zool. Syst. Evol. Res. 2021, 59, 2161–2169. [Google Scholar] [CrossRef]
- Reilly, S.B.; Wogan, G.O.; Stubbs, A.L.; Arida, E.; Iskandar, D.T.; McGuire, J.A. Toxic toad invasion of Wallacea: A biodiversity hotspot characterized by extraordinary endemism. Glob. Change Biol. 2017, 23, 5029–5031. [Google Scholar] [CrossRef] [Green Version]
- Soorae, P.T.; Frankham, G.J.; Ali Mohamed, A. The first record of the Asian common toad Duttaphrynus melanostictus Schneider, 1799 in Abu Dhabi, United Arab Emirates. Bioinvasions Rec. 2020, 9, 434–443. [Google Scholar] [CrossRef]
- Licata, F.; Harison, F.F.; Ficetola, F.G.; Freeman, K.; Muller, B.J.; Rodriguez Ponga, V.; Andreone, F.; Crottini, A. Toad invasion of Malagasy forests triggers severe mortality of a predatory snake. Biol. Invasions 2022, 24, 1189–1198. [Google Scholar] [CrossRef]
- Vences, M.; Brown, J.L.; Lathrop, A.; Rosa, G.M.; Cameron, A.; Crottini, A.; Dolch, R.; Edmonds, D.; Freeman, K.L.M.; Glaw, F. Tracing a toad invasion: Lack of mitochondrial DNA variation, haplotype origins, and potential distribution of introduced Duttaphrynus melanostictus in Madagascar. Amphib-reptil 2017, 38, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Santos, B.; Bletz, M.C.; Sabino-Pinto, J.; Cocca, W.; Fidy, J.F.; Freeman, K.L.; Kuenzel, S.; Ndriantsoa, S.; Noel, J.; Rakotonanahary, T.; et al. Characterization of the microbiome of the invasive Asian toad in Madagascar across the expansion range and comparison with a native co-occurring species. PeerJ 2021, 9, e11532. [Google Scholar] [CrossRef] [PubMed]
- Licata, F.; Mohanty, N.P.; Crottini, A.; Andreone, F.; Harison, R.F.; Randriamoria, T.M.; Freeman, K.; Muller, B.; Birkinshaw, C.; Tilahimena, A.; et al. Using public surveys to rapidly profile biological invasions in hard-to monitor areas. Anim. Cons., online version; 2023. [Google Scholar] [CrossRef]
- Merkel, A. Toamasina Climate (Madagascar). 2023. Available online: https://en.climate-data.org/africa/madagascar/toamasina/toamasina-4029/ (accessed on 18 April 2023).
- Gordon, A. Secondary sexual characters of Bufo melanostictus Schneider. Copeia 1933, 1933, 204–207. [Google Scholar] [CrossRef]
- Licata, F.; Andreone, F.; Freeman, K.; Rabesihanaka, S.; Robsomanitrandrasana, E.; Reardon, J.T.; Crottini, A. The Asian toad (Duttaphrynus melanostictus) in Madagascar: A report of an ongoing invasion. In Problematic Wildlife II: New Conservation and Management Challenges in the Human-Wildlife Interactions, 1st ed.; Angelici, F.M., Rossi, L., Eds.; Springer: Berlin, Germany, 2020; pp. 617–638. [Google Scholar]
- Erismis, U.C.; Arikan, U.; Konuk, M.; Guarino, F.M. Age structure and growth in caucasian parsley frog Pelodytes caucasicus (Boulenger, 1896) from Turkey. Russ. J. Herpetol. 2009, 16, 19–26. [Google Scholar]
- Guarino, F.M.; Crottini, A.; Mezzasalma, M.; Randrianirina, J.E.; Andreone, F. A skeletochronological estimate of age and growth in a large riparian frog from Madagascar (Anura, Mantellidae, Mantidactylus). Herpetozoa 2019, 32, 39–44. [Google Scholar] [CrossRef]
- Guarino, F.M.; Di Nocera, F.; Pollaro, G.; Galiero, G.; Iaccarino, D.; Iovino, D. Skeletochronology, age at maturity and cause of mortality of loggerhead sea turtles Caretta caretta stranded along the beaches of Campania (south-western Italy, western Mediterranean Sea). Herpetozoa 2020, 33, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Driscoll, D.A. Skeletochronological assessment of age structure and population stability for two threatened frog species. Austral. J. Ecol. 1999, 24, 182–189. [Google Scholar] [CrossRef]
- Sinsch, U.; Dehling, J.M. Tropical anurans mature early and die young: Evidence from eight Afromontane Hyperolius species and a meta-analysis. PLoS ONE 2017, 12, e0171666. [Google Scholar] [CrossRef] [Green Version]
- Miaud, C.; Guyetant, R.; Elmberg, J. Variations in life-history traits in the common frog Rana temporaria (Amphibia: Anura): A literature review and new data from the French Alps. J. Zool. 1999, 249, 61–73. [Google Scholar] [CrossRef]
- Guarino, F.M.; De Pous, P.; Crottini, A.; Mezzasalma, M.; Andreone, F. Age structure and growth in a population of Pelobates varaldii (Anura, Pelobatidae) from northwestern Morocco. Amphib-reptil. 2011, 32, 550–556. [Google Scholar] [CrossRef]
- Bionda, C.L.; Kost, S.; Salas, N.E.; Lajmanovich Sinsch, U.; Martino, A.L. Age structure, growth and longevity in the common toad, Rhinella arenarum, from Argentina. Acta Herpetol. 2015, 10, 55–62. [Google Scholar]
- Mogali, S.; Saidapur, S.; Shanbhag, B. Influence of desiccation threat on the metamorphic traits of the Asian common toad, Duttaphrynus melanostictus (Anura). Acta Herpetol. 2017, 12, 75–80. [Google Scholar]
- Henderson, P.A.; Seaby, R.M. Growth II. Pisces; Conservation Ltd.: Lymington, UK, 2001. [Google Scholar]
- Ngo, B.V.; Ngo, C.D. Reproductive activity and advertisement calls of the Asian common toad Duttaphrynus melanostictus (Amphibia, Anura, Bufonidae) from Bach Ma National Park, Vietnam. Zool. St. 2013, 52, 12. [Google Scholar] [CrossRef] [Green Version]
- Lofts, B. Amphibians. In Marshall’s Physiology of Reproduction. I. Reproductive Cycles of Vertebrates; Lamming, G.E., Ed.; Churchill Livingstone: Edinburgh, Scotland, 1984; Volume 1, pp. 127–205. [Google Scholar]
- Jørgensen, C.; Barker, K.S.; Vijayakumar, S. Body size, reproduction and growth in a tropical toad, Bufo melanostictus, with a comparison of ovarian cycles in tropical and temperate zone anurans. Oikos 1986, 46, 379–389. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Rozenblut, B.; Ogielska, M. Development and growth of long bones in european water frogs (Amphibia: Anura: Ranidae), with remarks on age determination. J. Morphol. 2005, 265, 304–317. [Google Scholar] [CrossRef]
- Rahman, M.M.; Lee, Y.Y.; Park, S.M.; Ham, C.H.; Sung, H.C. Confirmation of the applicability of skeletochronology and estimating the age Structure of Kaloula borealis (Microhylidae: Anura) at Lake Sihwa, South Korea. Biology 2022, 11, 898. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Bonin, A.; Miaud, C. Population genetics reveals origin and number of founders in a biological invasion. Mol. Ecol. 2008, 17, 773–782. [Google Scholar] [CrossRef]
- Tessa, G.; Del Forno, C.; Govindarajulu, P.; Tissot, N.; Miaud, C.; Andreone, F. Age and body size in four introduced populations of the American bullfrog, Lithobates catesbeianus (Ranidae). Eur. Zool. J. 2016, 83, 497–502. [Google Scholar] [CrossRef]
- Hemelaar, A. Age, growth and other population characteristics of Bufo bufo from different latitudes and altitudes. J. Herpetol. 1988, 22, 369–388. [Google Scholar] [CrossRef]
- Cherry, M.I.; Francillon-Vieillot, H. Body size, age and reproduction in the leopard toad, Bufo pardalis. J. Zool. 1992, 228, 41–45. [Google Scholar] [CrossRef]
- Licata, F.; Ficetola, G.F.; Falaschi, M.; Muller, B.J.; Andreone, F.; Harison, R.F. Spatial ecology of the invasive Asian common toad in Madagascar and its implications for invasion dynamics. Sci. Rep. 2023, 13, 3526. [Google Scholar] [CrossRef] [PubMed]
- Castanet, J. Age estimation and longevity in reptiles. Gerontology 1994, 40, 174–192. [Google Scholar] [CrossRef] [PubMed]
- Castanet, J.; Francillon-Vieillot, H.; Meunier, F.J.; de Ricqles, A. Bone and individual aging. In Bone; Hall, B.B.K., Ed.; CRC Press: Boca Raton, FL, USA, 1993; Volume 7, pp. 245–283. [Google Scholar]
- Smirina, E.M. Age determination and longevity in amphibians. Gerontology 1994, 40, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Zhang, L.; Lu, X. Global gaps in age data based on skeletochronology for amphibians. Integr. Zool. 2022, 17, 752–763. [Google Scholar] [CrossRef]
- Nayak, S.; Mahapatra, P.K.; Mishra, S.; Dutta, S.K. Age determination by skeletochronology in the common Indian toad Bufo melanostictus Schneider, 1799 (Anura: Bufonidae). Herpetozoa 2007, 19, 111–119. [Google Scholar]
- Andia, B.N.; Dixit, P.K.; Behera, S.; Behera, H.N. Age determination in common Indian Toad, Bufo melanostictus, by skeletochronology. J. Sci. Technol. 2010, 6, 42–56. [Google Scholar]
- Kumbar, S.M.; Lad, S. Determination of age and longevity of road mortal indian common toad Duttaphrynus melanostictus by skeletochronology. Rus. J. Herpetol. 2017, 24, 217–222. [Google Scholar] [CrossRef]
- Sahoo, D.D.; Kara, T.C. Determination of age, longevity and age at sexual maturity in common Asian Toad (Duttaphrynus Melanostictus) by skeletochronology. Octa J. Biosci. 2017, 5, 5–8. [Google Scholar]
- Leclair, M.H.; Leclair, R., Jr.; Gallant, J. Application of skeletochronology to a population of Pelobates cultripes (Anura: Pelobatidae) from Portugal. J. Herpetol. 2005, 39, 199–207. [Google Scholar] [CrossRef]
- Tessa, G.; Guarino, F.M.; Giacoma, C.; Mattioli, F.; Andreone, F. Longevity and body size in three populations of Dyscophus antongilii (Microhylidae, Dyscophinae), the tomato frog from north-eastern Madagascar. Acta Herpetol. 2007, 2, 139–147. [Google Scholar]
- Altunisik, A.; Ozdemir, N. Life history traits in Bufotes variabilis (Pallas, 1769) from 2 different altitudes in Turkey. Turkish J. Zool. 2015, 39, 153–159. [Google Scholar] [CrossRef]
- Guarino, F.M.; Tessa, G.; Mercurio, V.; Andreone, F. Rapid sexual maturity and short life span in the blue-legged frog and the rainbow frog from the arid Isalo Massif, southern-central Madagascar. Zoology 2010, 113, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Sinsch, U. Review: Skeletochronological assessment of demographic life-history traits in amphibians. Herpetol. J. 2015, 25, 5–13. [Google Scholar]
- Tessa, G.; Crottini, A.; Giacoma, C.; Guarino, F.M.; Randrianirina, J.E.; Andreone, F. Comparative longevity and age at sexual maturity in twelve rainforest frogs of the genera Boophis, Gephyromantis, and Mantidactylus (Anura: Mantellidae) from Madagascar. Phyllomedusa 2017, 16, 13–21. [Google Scholar] [CrossRef]
- Licata, F.; Ficetola, G.F.; Freeman, K.; Mahasoa, H.R.; Ravololonarivo, V.; Fidy, F.J.S.N. Abundance, distribution and spread of the invasive Asian toad Duttaphrynus melanostictus in eastern Madagascar. Biol. Invasions 2019, 21, 1615–1626. [Google Scholar] [CrossRef]
- Halliday, T.; Verrell, P. Body size and age in amphibians and reptiles. J. Herpetol. 1988, 22, 253–265. [Google Scholar] [CrossRef]
- Andreone, F.; Dawson, J.S.; Rabemananjara, F.C.E.; Rabibisoa, N.H.C.; Rakotonanahary, T.S. New Sahonagasy Action Plan 2016–2020; Museo Regionale di Scienze Naturali and Amphibian Survival Alliance: Turin, Italy, 2016; pp. 1–46. [Google Scholar]
- Rakotoarison, A.; Ndriantsoa, S.H.; Rabemananjara, F.C.E.; Rabibisoa, N.H.C.; Rakotonanahary, T.F.; Randriamahazo, J.A.R.H.; Andreone, F. More than 15 years of amphibian conservation in Madagascar under the flag of IUCN SSC Amphibian Specialist Group. Naturalista sicil. 2022, 46, 305–312. [Google Scholar]
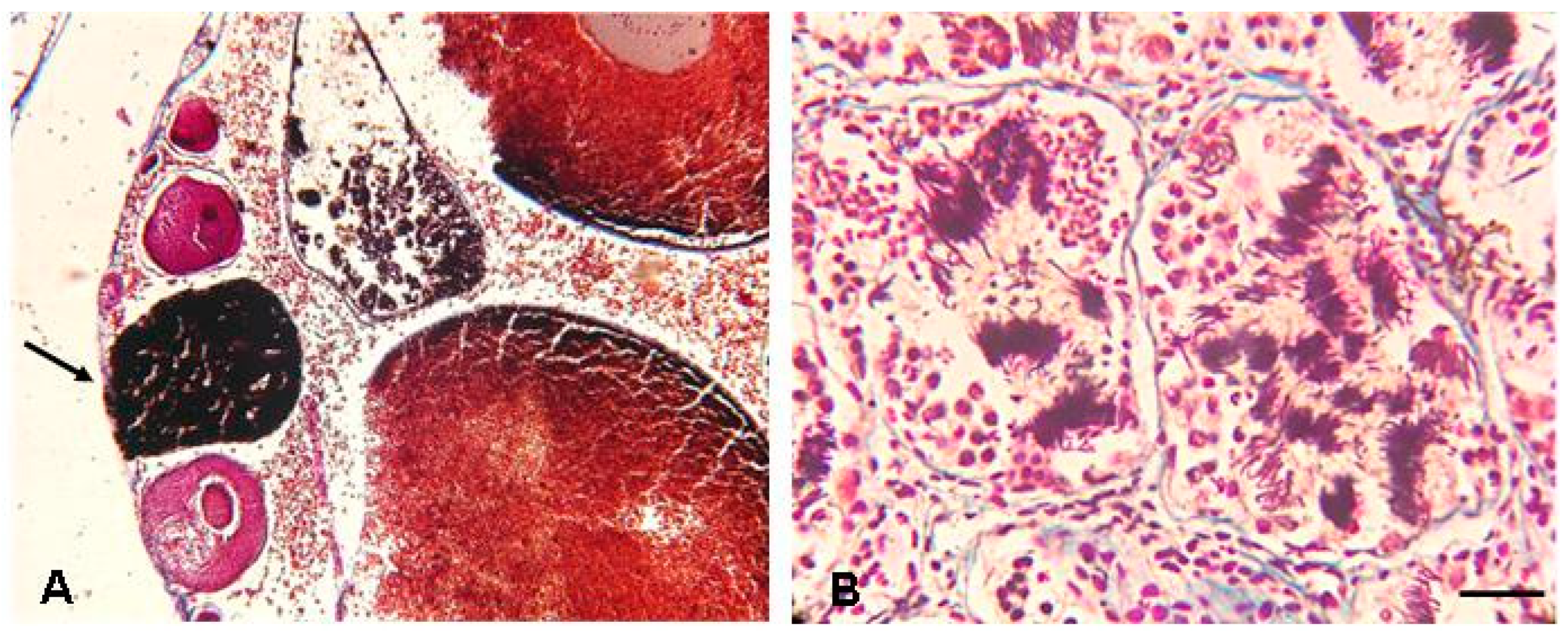

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site 1 | Site 2 | Site 3 | Pooled Sites | |
|---|---|---|---|---|
| Males | 73.4 ± 2.1 | 70.9 ± 2.3 | 67.5 ± 8.4 | 71.4 ± 1.6 |
| 62.2–89.8 | 60.0–88.2 | 58.8–88.4 | 59.8–89.8 | |
| n = 15 | n = 11 | n = 3 | n = 29 | |
| Females | 81.5 ± 2.2 | 78.7 ± 3.8 | 71.0 ± 5.6 | 78.2 ± 1.9 |
| 61.4–89.7 | 57.5–104.8 | 57.6–109.0 | 57.5–104.8 | |
| n = 14 | n = 13 | n = 7 | n = 34 | |
| Juveniles | 45.1 ± 2.0 | 45.7 ± 4.2 | 38.5 ± 3.0 | 41.5 ± 1.9 |
| (ND) | 36.2–49.7 | 41.5–49.9 | 25.4–49.2 | 25.4–49.0 |
| n = 6 | n = 2 | n = 10 | n = 18 |
| Site 1 | Site 2 | Site 3 | Pooled Sites | |||||
|---|---|---|---|---|---|---|---|---|
| Mean | Mode | Mean | Mode | Mean | Mode | Mean | Mode | |
| Males | 1.7 ± 0.8 1–3 n = 9 | 3 | 2.1 ± 0.6 1–3 n = 8 | 2 | 1.0 ± 0 1 n = 2 | - | 1.8 ± 0.7 1–3 n = 19 | 2 (36.8%) |
| Females | 2.9 ± 1.4 1–6 n = 12 | 2, 3, 4 | 2.7 ± 1.2 1–5 n = 9 | 2, 3 | 2.2 ± 1.3 1–3 n = 5 | 1 | 2.7 ± 1.3 1–6 n = 26 | 2, 3 (53.8%) |
| Juveniles ND | 1.0 ± 0 1 n = 4 | 1 | - - n = 0 | - | 1 1 n = 1 | - | 1 1 n = 5 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guarino, F.M.; Andreone, F.; Mezzasalma, M.; Licata, F.; Puoti, S.; Santos, B.; Cocca, W.; Solofoniaina Fidy, J.F.; Ndriantsoa, S.H.; Noel, J.; et al. Life History Traits and Longevity of the Invasive Asian Common Toad Duttaphrynus melanostictus (Schneider, 1799) in Madagascar. Animals 2023, 13, 2099. https://doi.org/10.3390/ani13132099
Guarino FM, Andreone F, Mezzasalma M, Licata F, Puoti S, Santos B, Cocca W, Solofoniaina Fidy JF, Ndriantsoa SH, Noel J, et al. Life History Traits and Longevity of the Invasive Asian Common Toad Duttaphrynus melanostictus (Schneider, 1799) in Madagascar. Animals. 2023; 13(13):2099. https://doi.org/10.3390/ani13132099
Chicago/Turabian StyleGuarino, Fabio Maria, Franco Andreone, Marcello Mezzasalma, Fulvio Licata, Simona Puoti, Bárbara Santos, Walter Cocca, Jean Francois Solofoniaina Fidy, Serge Herilala Ndriantsoa, Jean Noel, and et al. 2023. "Life History Traits and Longevity of the Invasive Asian Common Toad Duttaphrynus melanostictus (Schneider, 1799) in Madagascar" Animals 13, no. 13: 2099. https://doi.org/10.3390/ani13132099