
Fin Whale (Balaenoptera physalus) Mortality along the Italian Coast between 1624 and 2021
 , ,
, ,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of Mortality Data
2.2. Statistical Analysis
2.2.1. Temporal Analysis
2.2.2. Spatial Analysis
2.2.3. Spatial–Temporal Analysis
2.2.4. Correlation Analysis
3. Results
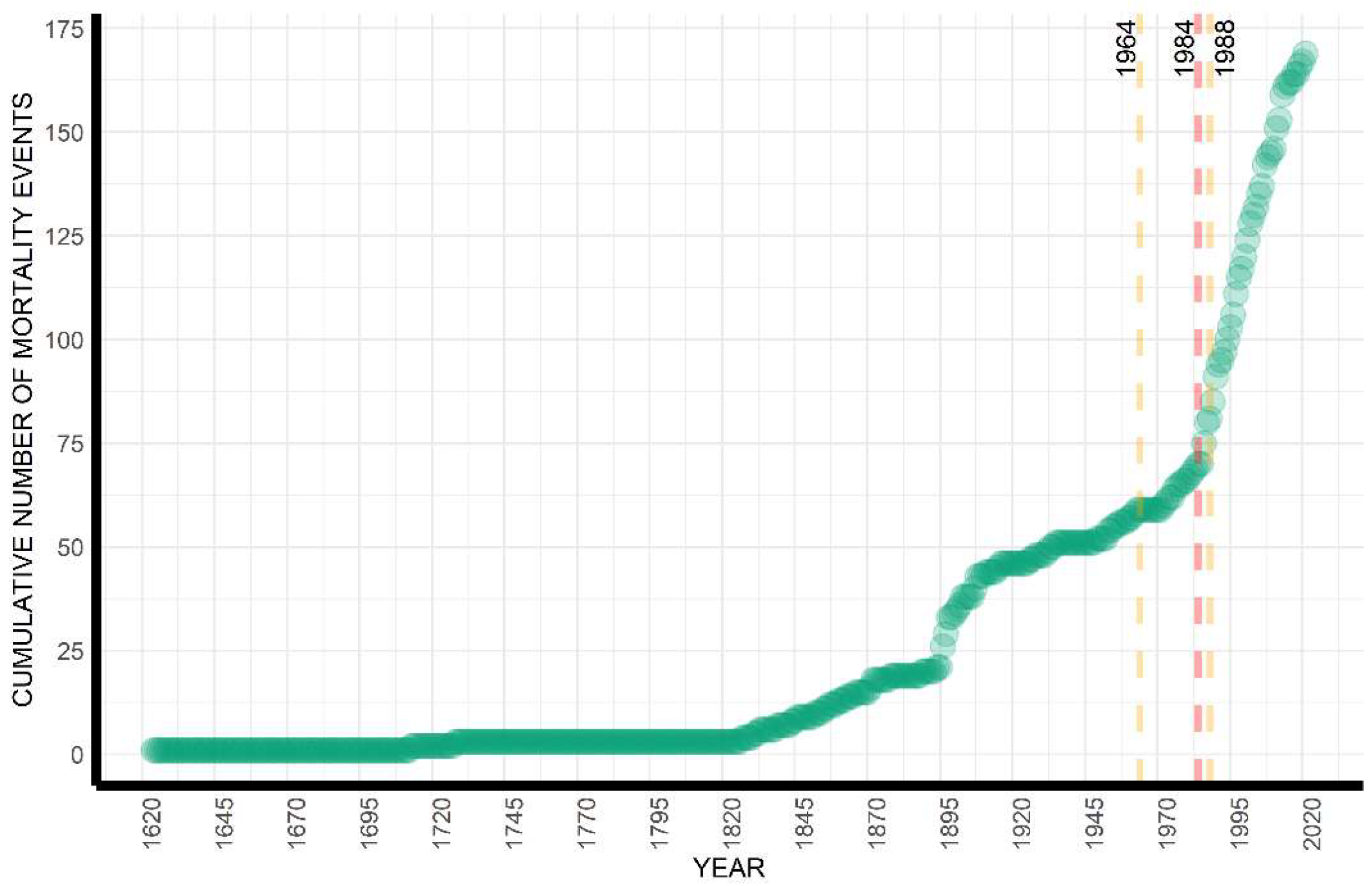
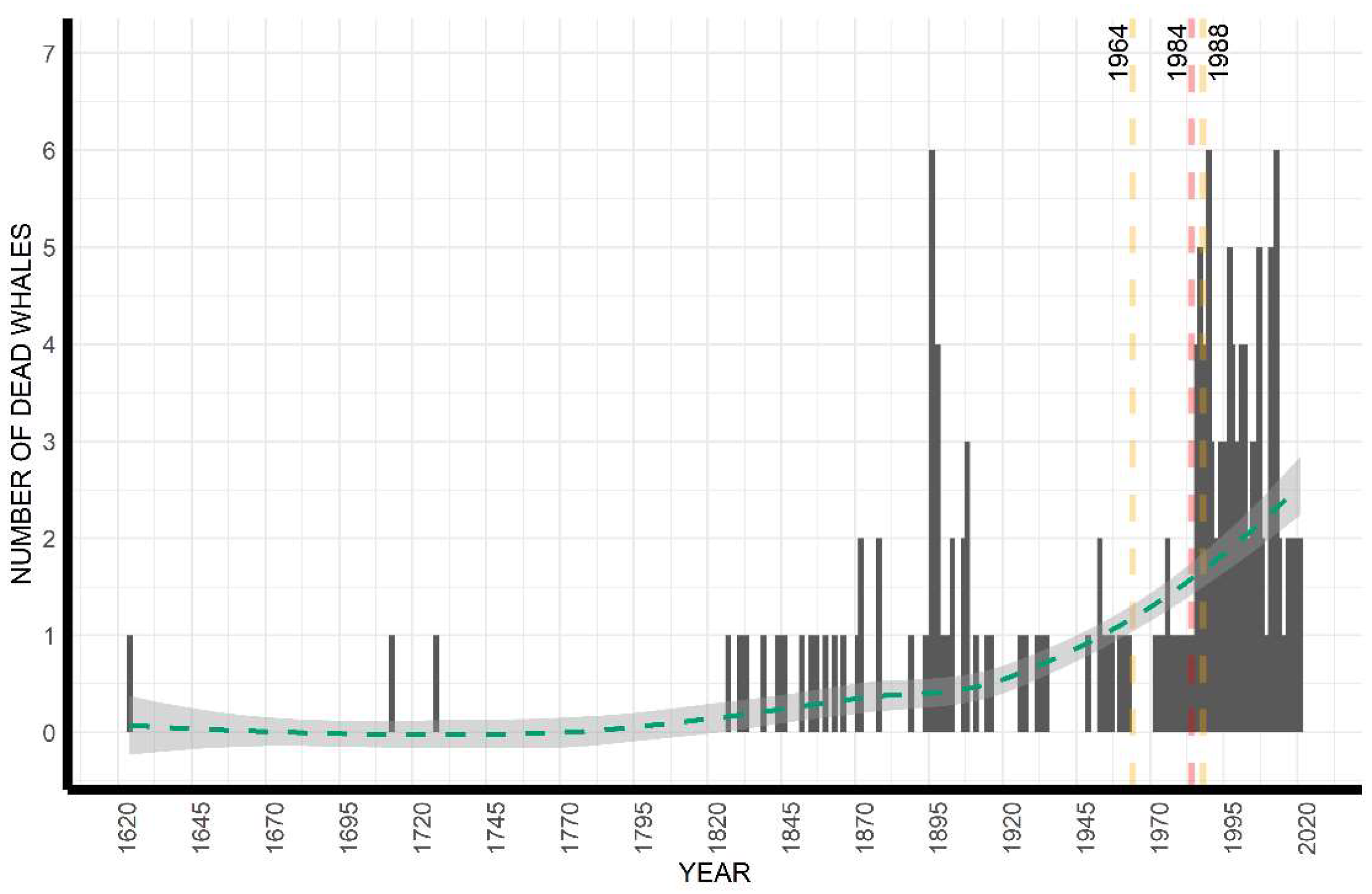
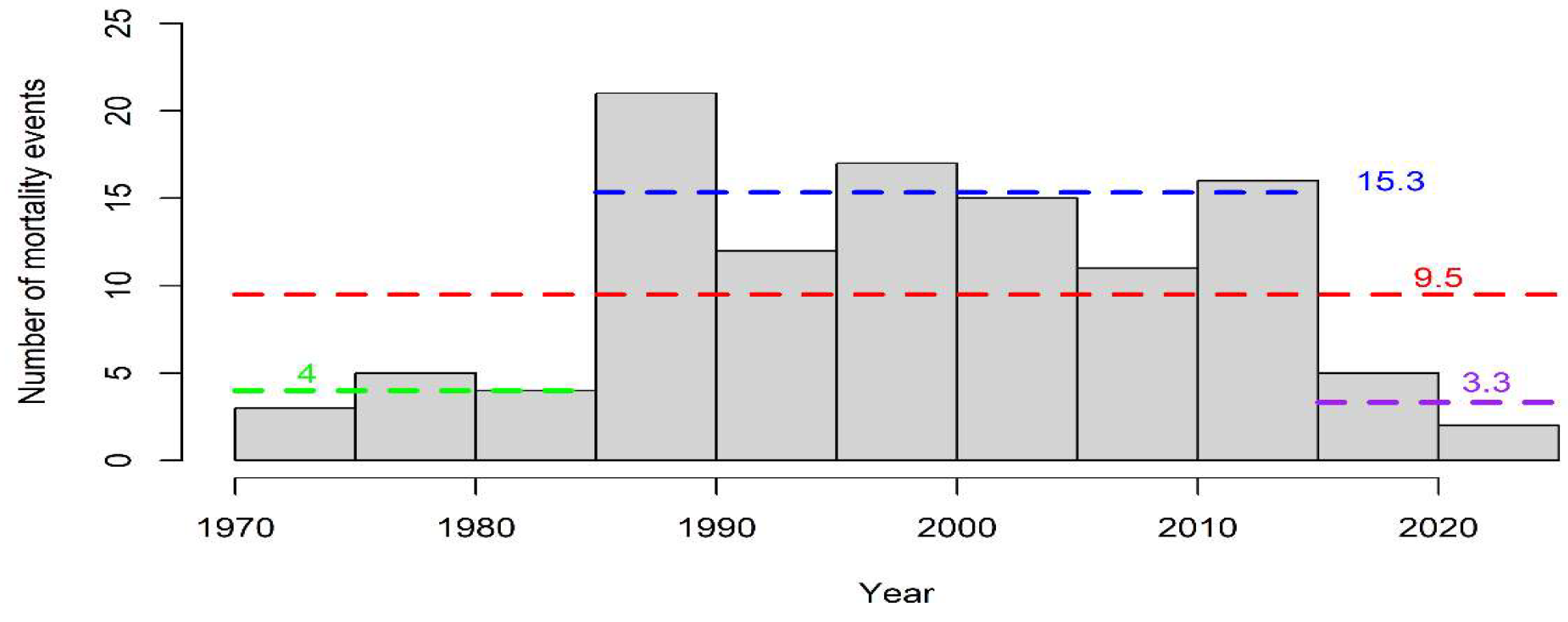
3.1. Temporal Analysis
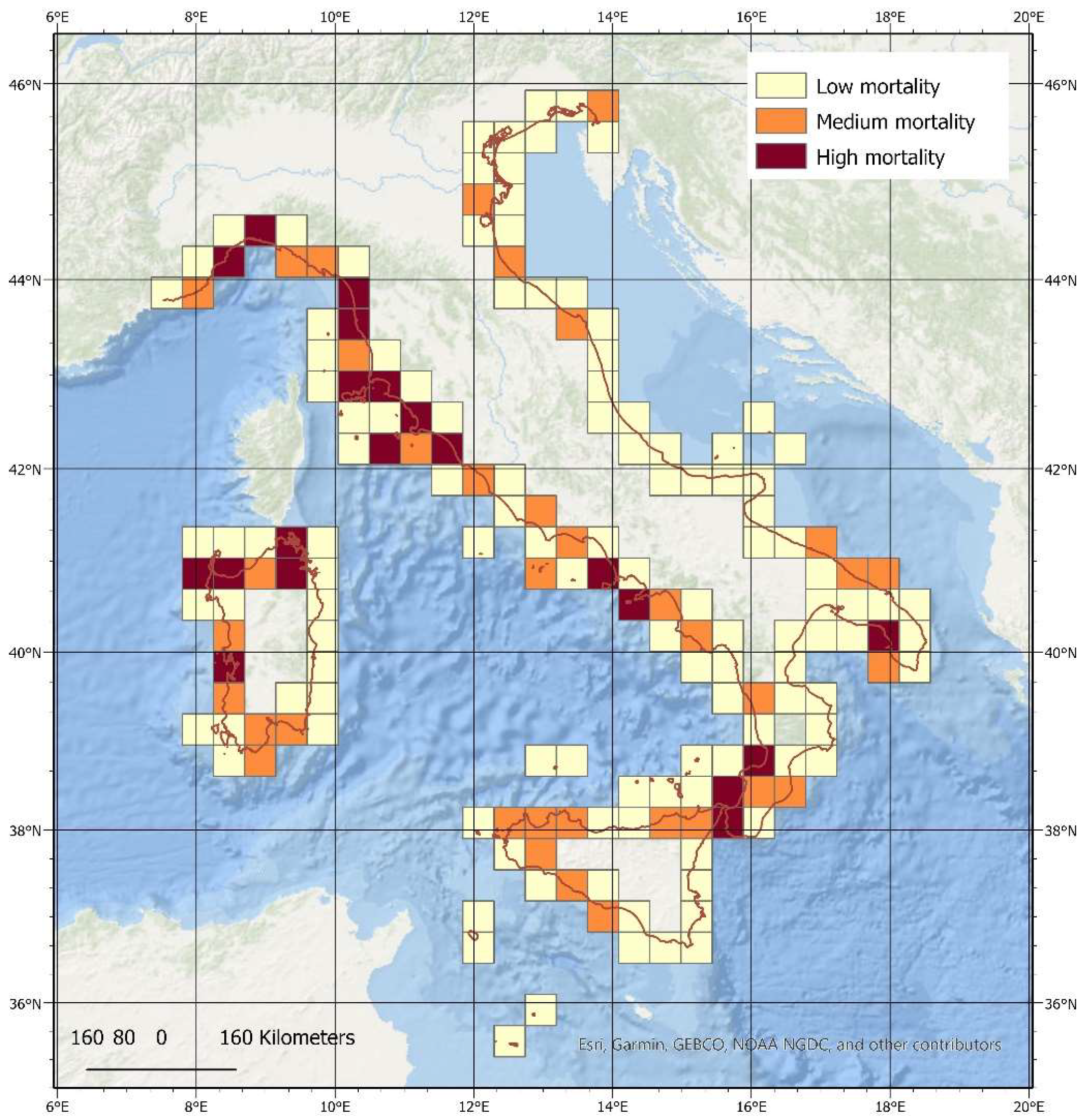
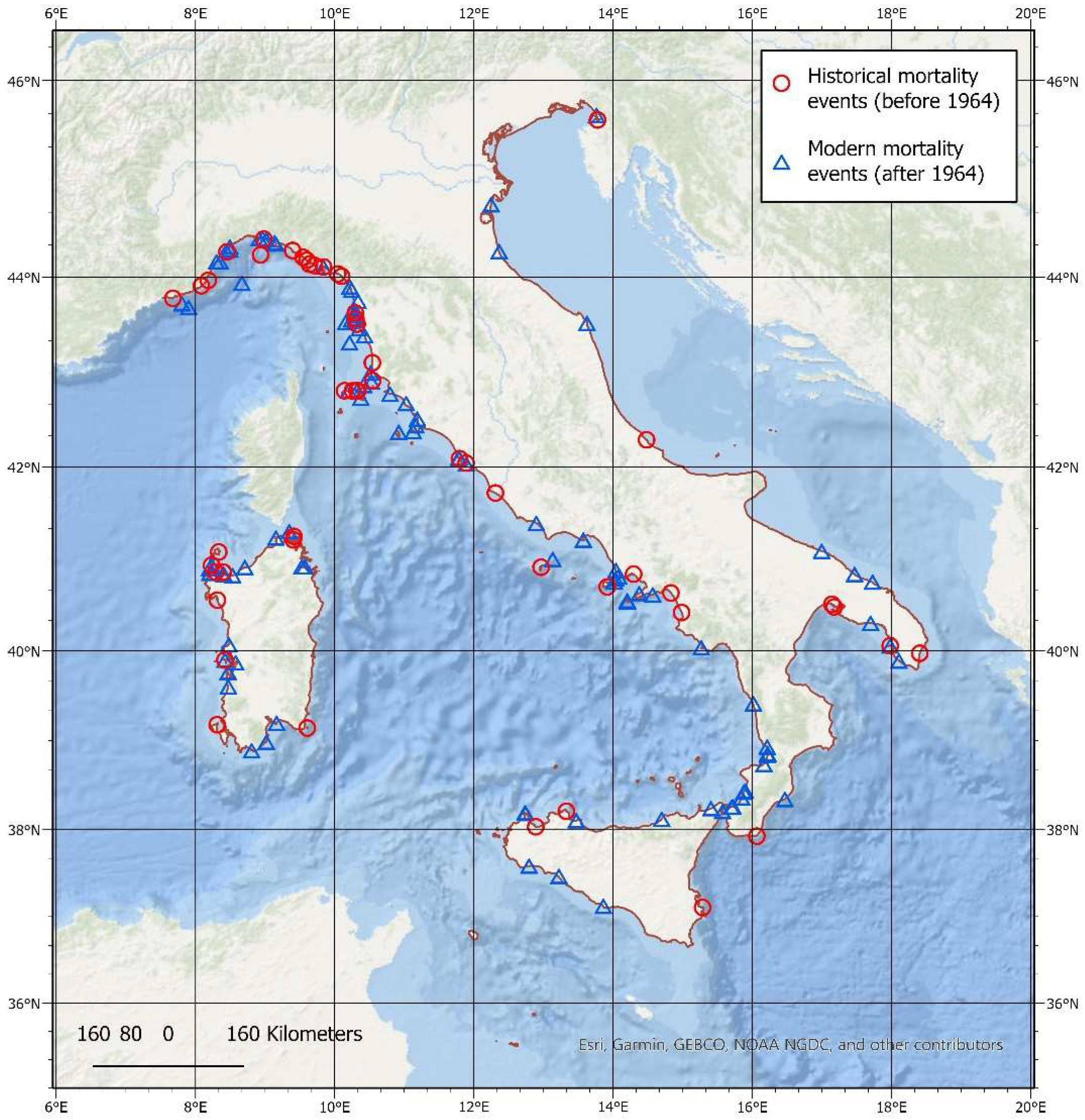
3.2. Spatial Analysis
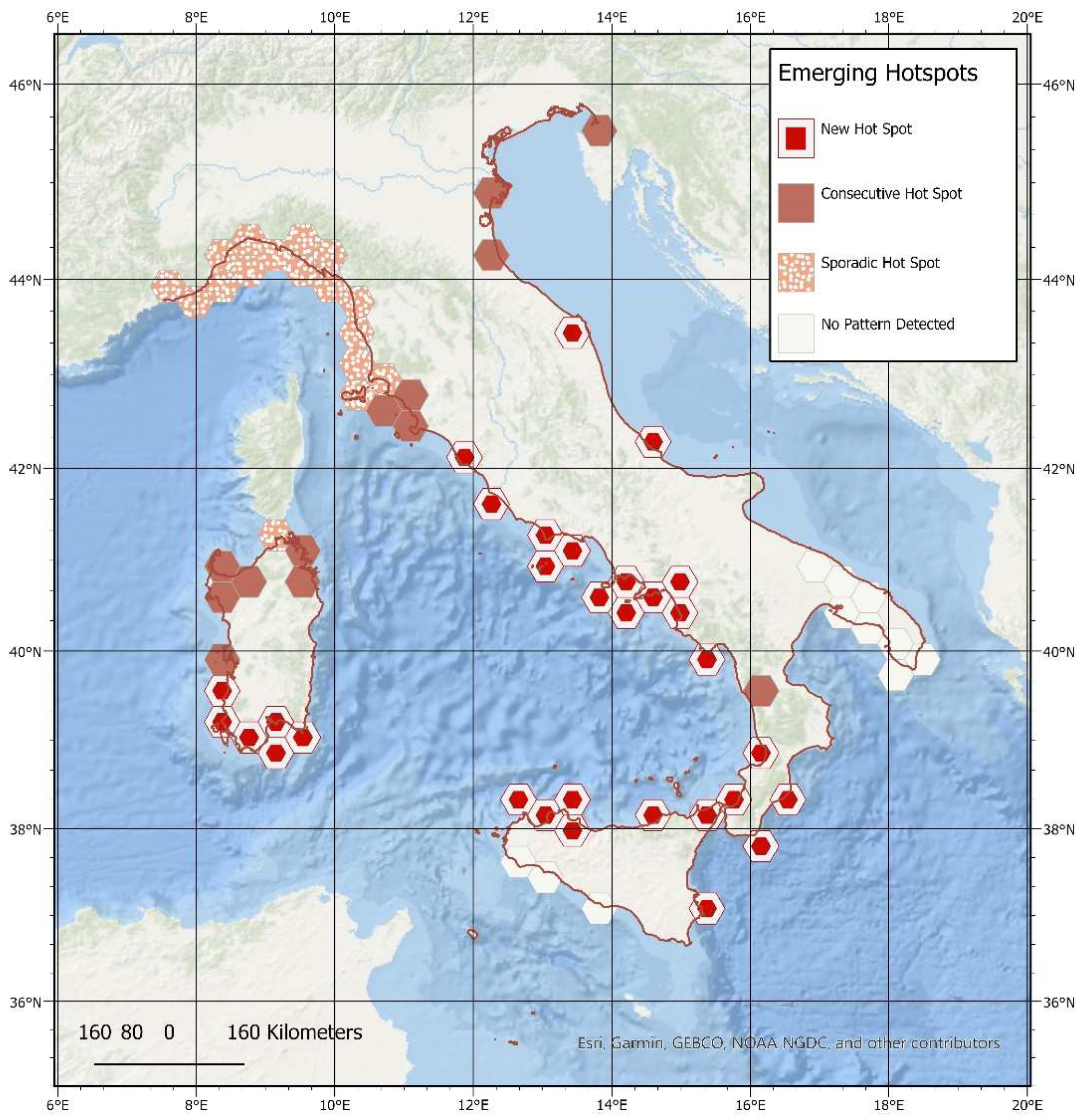
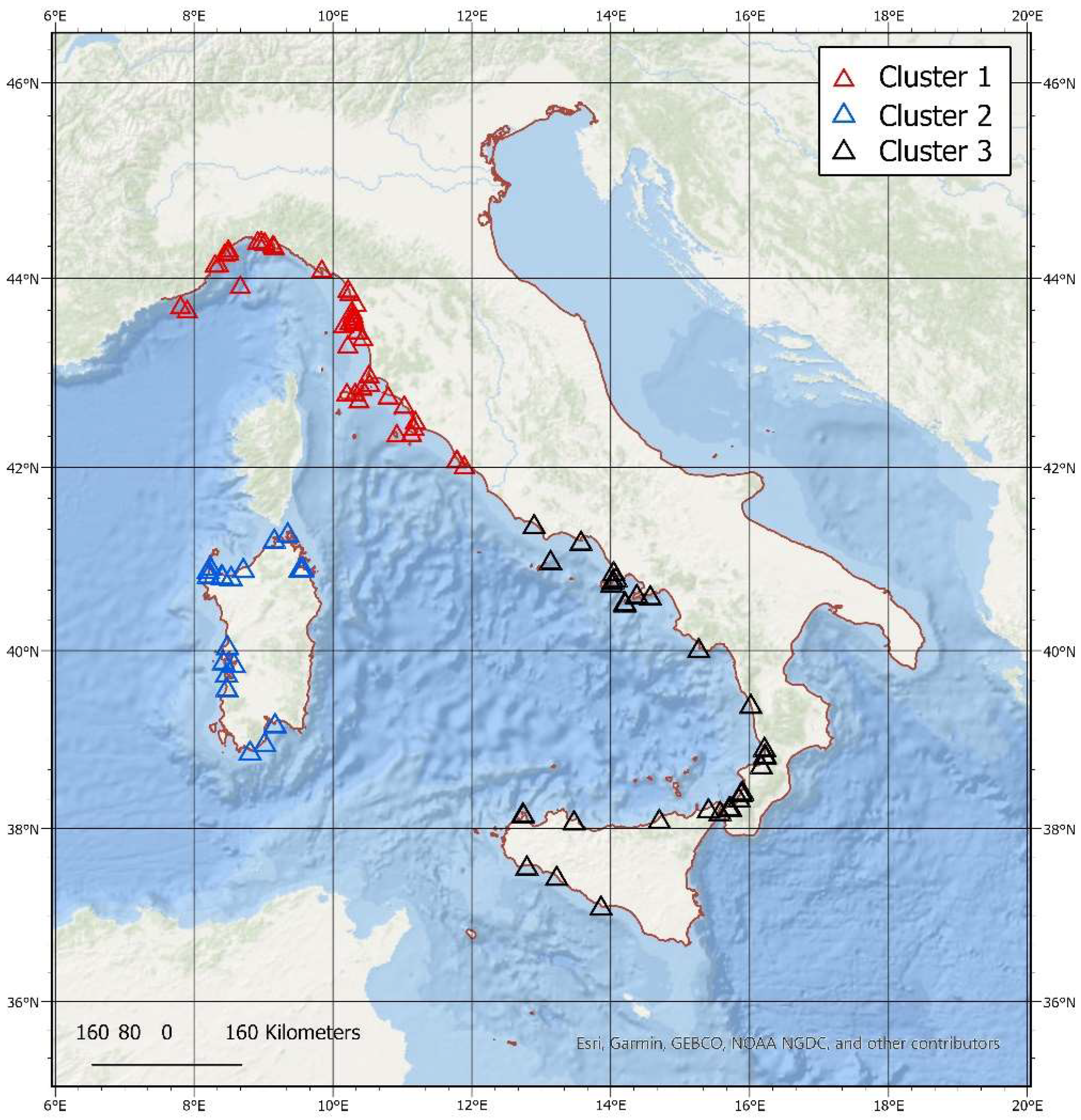
3.3. Spatial–Temporal Analysis
3.4. Correlation Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Ben Rais Lasram, F.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The Biodiversity of the Mediterranean Sea: Estimates, Patterns, and Threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pyenson, N.D. Carcasses on the coastline: Measuring the ecological fidelity of the cetacean stranding record in the eastern North Pacific Ocean. Paleobiology 2010, 36, 453–480. [Google Scholar] [CrossRef]
- Pyenson, N.D. The high fidelity of the cetacean stranding record: Insights into measuring diversity by integrating taphonomy and macroecology. Proc. R. Soc. B 2011, 278, 3608–3616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coombs, E.J.; Deaville, R.; Sabin, R.C.; Allan, L.; O’Connell, M.; Berrow, S.; Smith, B.; Brownlow, A.; Doeschate, M.T.; Penrose, R.; et al. What can cetacean stranding records tell us? A study of UK and Irish cetacean diversity over the past 100 years. Mar. Mam. Sci. 2019, 35, 1527–1555. [Google Scholar] [CrossRef]
- Mazzariol, S.; Siebert, U.; Scheinin, A.; Deaville, R.; Brownlow, A.; Uhart, M.; Marcondes, M.; Hernandez, G.; Stimmelmayr, R.; Rowles, T.; et al. Summary of Unusual Cetaceans Strandings Events Worldwide (2018–2020); SC-68B/E/09 Rev1; 2020. [Google Scholar]
- IJsseldijka, L.L.; ten Doeschate, M.T.I.; Brownlow, A.; Davison, N.J.; Deaville, R.; Galatius, A.; Gilles, A.; Haelters, J.; Jepson, P.D.; Keijl, G.O.; et al. Spatiotemporal mortality and demographic trends in a small cetacean: Strandings to inform conservation management. Biol. Cons. 2020, 249, 108733. [Google Scholar] [CrossRef]
- Li, W.T.; Chou, L.S.; Chiou, H.Y.; Chen, I.H.; Yang, W.C. Analyzing 13 Years of Cetacean Strandings: Multiple Stressors to Cetaceans in Taiwanese Waters and Their Implications for Conservation and Future Research. Front. Mar. Sci. 2021, 8, 606722. [Google Scholar] [CrossRef]
- Betty, E.L.; Stockin, K.A.; Hinton, B.; Bollard, B.A.; Smith, A.N.H.; Orams, M.B.; Murphy, S. Age, growth, and sexual dimorphism of the Southern Hemisphere long-finned pilot whale (Globicephala melas edwardii). J. Mammal. 2022, 103, 560–575. [Google Scholar] [CrossRef]
- Reddy, M.L.; Dierauf, L.A.; Gulland, F.M.D. Marine Mammals as Sentinels of Ocean Health in CRC Handbook of Marine Mammal Medicine, 2nd ed.; Dierauf, L.A., Gulland, F.M.D., Eds.; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Cornaglia, E.; Rebora, L.; Gili, C.; Di Guardo, G. Histopathological and immunohistochemical studies on cetaceans found stranded on the coast of Italy between 1990 and 1997. J. Vet. Med. A Physiol. Pathol. Clin. Med. 2000, 47, 129–142. [Google Scholar] [CrossRef]
- Bonar, C.J.; Boede, E.O.; Hartmann, M.G.; Lowenstein-Whaley, J.; Mujica-Jorquera, E.; Parish, S.V.; Parish, J.V.; Garner, M.M.; Stadler, C.K. A retrospective study of pathologic findings in the Amazon and Orinoco river dolphin (Inia geoffrensis) in captivity. J. Zoo Wildl. Med. 2007, 38, 177–191. [Google Scholar] [CrossRef]
- Diaz-Delgado, J.; Fernandez, A.; Sierra, E.; Sacchini, S.; Andrada, M.; Vela, A.I.; Quesada-Canales, Ó.; Paz, Y.; Zucca, D.; Groch, K.; et al. Pathologic findings and causes of death of stranded cetaceans in the Canary Islands (2006–2012). PLoS ONE 2018, 13, e0204444. [Google Scholar] [CrossRef]
- Puig-Lozano, R.; Bernaldo, R.; De Quirós, Y.; Díaz-Delgado, J.; García-Ñlvarez, N.; Sierra, E.; De la Fuente, J.; Sacchini, S.; Suárez-Santana, C.M.; Zucca, D.; et al. Retrospective study of foreign body-associated pathology in stranded cetaceans, Canary Islands (2000–2015). Environ. Pollut. 2018, 43, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Klinowska, M. Cetacean live strandings relate to geomagnetic topography. Aquat. Mamm. 1985, 1, 27–32. [Google Scholar]
- Brabyn, M.W.; McLean, I.G. Oceanography and coastal topography of herd-stranding sites for whales in New Zealand. J. Mamm. 1992, 73, 469–476. [Google Scholar] [CrossRef]
- Evans, K.; Thresher, R.; Warneke, R.M.; Bradshaw, C.J.A.; Pook, M.; Thiele, D.; Hindell, M.A. Periodic variability in cetacean strandings: Links to large-scale climate events. Biol. Lett. 2005, 1, 147–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quaggiotto, M.M.; Sánchez-Zapata, J.A.; Bailey, D.M.; Payo-Payo, A.; Navarro, J.; Brownlow, A.; Deaville, R.; Lambertucci, S.A.; Selva, N.; Cortés-Avizanda, A.; et al. Past, present and future of the ecosystem services provided by cetacean carcasses. Ecosyst. Serv. 2022, 54, 101406. [Google Scholar] [CrossRef]
- Bérubé, M.; Aguilar, A.; Dendanto, D.; Larsen, F.; Notarbartolo di Sciara, G.; Sears, R.; Sigurjónsson, J.; Urban-R, J.; Palsbøll, P.J. Population genetic structure of North Atlantic, Mediterranean Sea and Sea of Cortez fin whales, Balaenoptera physalus (Linnaeus 1758): Analysis of mitochondrial and nuclear loci. Mol. Ecol. 1998, 7, 585–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palsbøll, P.J.; Bérubé, M.; Aguilar, A.; Notarbartolo Di Sciara, G.; Nielsen, R. Discerning between recurrent gene flow and recent divergence under a finite-site mutation model applied to North Atlantic and Mediterranean Sea fin whale (Balaenoptera physalus) populations. Evolution 2004, 58, 670–675. [Google Scholar] [CrossRef]
- Notarbartolo di Sciara, G.; Agardy, T.; Hyrenbach, D.; Scovazzi, T.; Van Klaveren, P. The Pelagos sanctuary for 570 Mediterranean marine mammals. Aquat. Conserv. Mar. Freshw. Ecosyst. 2008, 18, 367–391. [Google Scholar] [CrossRef]
- Canese, S.; Cardinali, A.; Fortuna, C.; Giusti, M.; Lauriano, G.; Salvati, E.; Greco, S. The first identified winter feeding ground of fin whales (Balaenoptera physalus) in the Mediterranean Sea. J. Mar. Biol. Assoc. United Kingd. 2006, 86, 903–907. [Google Scholar] [CrossRef]
- Panigada, S.; Donovan, G.P.; Druon, J.-N.; Lauriano, G.; Pierantonio, N.; Pirotta, E.; Zanardelli, M.; Zerbini, A.N.; Notarbartolo di Sciara, G.N. Satellite Tagging of Mediterranean Fin Whales: Working towards the Identification of Critical Habitats and the Focussing of Mitigation Measures. Sci. Rep. 2017, 7, 3365. [Google Scholar] [CrossRef] [Green Version]
- Pintore, L.; Sciacca, V.; Viola, S.; Giacoma, C.; Papale, E.; Giorli, G. Fin Whale (Balaenoptera physalus) in the Ligurian Sea: Preliminary Study on Acoustics Demonstrates Their Regular Occurrence in Autumn. J. Mar. Sci. Eng. 2021, 9, 966. [Google Scholar] [CrossRef]
- ACCOBAMS. Estimates of Abundance and Distribution of Cetaceans, Marine Mega-Fauna and Marine Litter in the Mediterranean Sea from 2018–2019 Surveys; Panigada, S., Boisseau, O., Canadas, A., Lambert, C., Laran, S., McLanaghan, R., Moscrop, A., Eds.; ACCOBAMS—ACCOBAMS Survey Initiative Project: Monaco City, Monaco, 2021; 177p. [Google Scholar]
- Forcada, J.; Aguilar, A.; Hammond, P.; Pastor, X.; Aguilar, R. Distribution abundance of fin whales (Balaenoptera physalus) in the western Mediterranean Sea during the summer. J. Zool. 1996, 238, 23–24. [Google Scholar] [CrossRef]
- Forcada, J.; Notarbartolo di Sciara, G.; Fabbri, F. Abundance of fin whales and striped dolphins summering in the Corso Ligurian basin. Mammalia 1995, 59, 127–140. [Google Scholar] [CrossRef]
- Panigada, S.; Gauffier, P.; Notarbartolo di Sciara, G. Balaenoptera physalus (Mediterranean subpopulation). In The IUCN Red List of Threatened Species; 2021; p. e.T16208224A50387979. [Google Scholar] [CrossRef]
- Mazzariol, S.; Marcer, F.; Mignone, W.; Serracca, L.; Goria, M.; Marsili, L.; Di Guardo, G.; Casalone, C. Dolphin morbillivirus and Toxoplasma gondii coinfection in a Mediterranean fin whale (Balaenoptera physalus). BMC Vet. Res. 2012, 8, 20. [Google Scholar] [CrossRef] [Green Version]
- Casalone, C.; Mazzariol, S.; Pautasso, A.; Di Guardo, G.; Di Nocera, F.; Lucifora, G.; Ligios, C.; Franco, A.; Fichi, G.; Cocumelli, C.; et al. Cetacean strandings in Italy: An unusual mortality event along the Tyrrhenian Sea coast in 2013. Dis. Aquat. Organ. 2014, 109, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Mazzariol, S.; Centelleghe, C.; Beffagna, G.; Povinelli, M.; Terracciano, G.; Cocumelli, C.; Pintore, A.; Denurra, D.; Casalone, C.; Pautasso, A.; et al. Mediterranean Fin Whales (Balaenoptera physalus) Threatened by Dolphin MorbilliVirus. Emerg. Infect. Dis. 2016, 22, 302–305. [Google Scholar] [CrossRef]
- Mira, F.; Rubio-Guerri, C.; Purpari, G.; Puleio, R.; Caracappa, G.; Gucciardi, F.; Russotto, L.; Loria, G.R.; Guercio, A. Circulation of a novel strain of dolphin morbillivirus (DMV) in stranded cetaceans in the Mediterranean Sea. Sci. Rep. 2019, 9, 9792. [Google Scholar] [CrossRef] [Green Version]
- Panigada, S.; Lauriano, G.; Donovan, G.; Pierantonio, N.; Cañadas, A.; Vázquez, J.A.; Burt, L. Estimating Cetacean Density and Abundance in the Central and Western Mediterranean Sea through Aerial Surveys: Implications for Management. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2017, 141, 41–58. [Google Scholar] [CrossRef] [Green Version]
- Panigada, S.; Pesante, G.; Zanardelli, M.; Capoulade, F.; Gannier, A.; Weinrich, M.T. Mediterranean fin whales at risk from fatal ship strikes. Mar. Pollut. Bull. 2006, 52, 1287–1298. [Google Scholar] [CrossRef]
- Peltier, H.; Beaufils, A.; Cesarini, C.; Dabin, W.; Dars, C.; Demaret, F.; Dhermain, F.; Doremus, G.; Labach, H.; Van Canneyt, O.; et al. Monitoring of Marine Mammal Strandings Along French Coasts Reveals the Importance of Ship Strikes on Large Cetaceans: A Challenge for the European Marine Strategy Framework Directive. Front. Mar. Sci. 2019, 6, 486. [Google Scholar] [CrossRef]
- Ritter, F.; Panigada, S. Collisions of Vessels with Cetaceans—The Underestimated Threat World Seas: An Environmental Evaluation Volume III: Ecological Issues and Environmental Impacts; Elsevier: Amsterdam, The Netherlands, 2019; pp. 531–547. ISBN 978-0-12-805052-1. [Google Scholar]
- Panigada, S.; Azzellino, A.; Cubaynes, H.; Folegot, T.; Fretwell, P.; Jacob, T.; Lanfredi, C.; Leaper, R.; Ody, D.; Ratel, M. Proposal to Develop and Evaluate Mitigation Strategies to Reduce the Risk of Ship Strikes to Fin and Sperm Whales in the Pelagos Sanctuary—Final Report. In Pelagos Secretariat—Convention; Tethys Research Institute: Milan, Italy, 2020; 67p. [Google Scholar]
- Winkler, C.; Panigada, S.; Murphy, S.; Ritter, F. Global Numbers of Ship Strikes: An Assessment of Collisions Between Vessels and Cetaceans Using Available Data in the IWC Ship Strike Database; Report to the International Whaling Commission, IWC/68B/SC HIM09 Rev1; International Whaling Commission: Cambridge, UK, 2020; 33p. [Google Scholar]
- Sèbe, M.; Kontovas, C.A.; Pendleton, L.; Gourguet, S. Cost-effectiveness of measures to reduce ship strikes: A case study on protecting the Mediterranean fin whale. Sci. Total Environ. 2022, 827, 154236. [Google Scholar] [CrossRef] [PubMed]
- Fossi, M.C.; Marsili, L.; Neri, G.; Natoli, A.; Politi, E.; Panigada, S. The use of a non-lethal tool for evaluating toxicological hazard of organochlorine contaminants in Mediterranean cetaceans: New data 10 years after the first paper published in MPB. Mar. Pollut. Bull. 2006, 46, 972–982. [Google Scholar] [CrossRef]
- Mancia, A.; Abelli, L.; Fossi, M.C.; Panti, C. Skin distress associated with xenobiotics exposure: An epigenetic study in the Mediterranean fin whale (Balaenoptera physalus). Mar. Gen. 2021, 57, 100822. [Google Scholar] [CrossRef]
- Pinzone, M.; Budzinski, H.; Tasciotti, A.; Ody, D.; Lepoint, G.; Schnitzler, J.; Scholl, G.; Thomé, J.P.; Tapie, N.; Eppe, G.; et al. POPs in free-ranging pilot whales, sperm whales and fin whales from the Mediterranean Sea: Influence of biological and ecological factors. Environ. Res. 2015, 142, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Notarbartolo di Sciara, G.; Castellote, M.; Druon, J.N.; Panigada, S. Fin Whales, Balaenoptera physalus: At Home in a Changing Mediterranean Sea? Adv. Mar. Biol. 2016, 75, 75–101. [Google Scholar]
- Castellote, M.; Clark, C.W.; Lammers, M.O. Acoustic and behavioural changes by fin whales (Balaenoptera physalus) in response to shipping and airgun noise. Biol. Conserv. 2012, 147, 115–122. [Google Scholar] [CrossRef]
- Gambaiani, D.; Mayol, P.; Isaac, S.; Simmonds, M. Potential impacts of climate change and greenhouse gas emissions on Mediterranean marine ecosystems and cetaceans. J. Mar. Biol. Assoc. United Kingd. 2009, 89, 179–201. [Google Scholar] [CrossRef] [Green Version]
- Borri, M. Una rete per i delfini. Cetacea Inf. 1995, 4, 42–44. [Google Scholar]
- Di Lorenzo, A.; Olivieri, V.; Internullo, E.; Bortolotto, A.; Manfrini, V.; Guccione, S.; Piscione, I.; Di Nardo, W.; Tringali, M. GeoCetus: Sistema informativo geografico per la gestione di una banca dati online degli spiaggiamenti di cetacei lungo le coste italiane. Biol. Mar. Mediterr. 2013, 20, 256–257. [Google Scholar]
- Tomilin, A.G. Cetacea. In Mammals of the U.S.S.R. and Adjacent Countries; Heptner, V.G., Ed.; Israel Program for Scientific Translations: Jerusalem, Israel, 1957; Volume 9, 756p. [Google Scholar]
- Robineau, D. Cétacés de France. Faune de France; Féderation Française des Sociétés de Sciences Naturelle: Paris, France, 2005; Volume 89, 646p. [Google Scholar]
- Pyenson, N.D.; Goldbogen, J.A.; Shadwick, R.E. Mandible allometry in extant and fossil Balaenopteridae (Cetacea: Mammalia): The largest vertebrate skeletal element and its role in rorqual lunge feeding. Biol. J. Linn. Soc. 2013, 108, 586–599. [Google Scholar] [CrossRef] [Green Version]
- Mogoe, T.; Bando, T.; Maeda, H.; Kato, H.; Ohsumi, S. Biological Observations of Fin Whales Sampled by JARPAII in the Antarctic; SC/F14/J10; IWC: Cambridge, UK, 2014. [Google Scholar]
- Rossi, A.; Panigada, S.; Arrigoni, M.; Zanardelli, M.; Cimmino, C.; Marangi, L.; Manfredi, P.; Santangelo, G. Demography and conservation of the Mediterranean fin whale (Balaenoptera physalus): What clues can be obtained from photo-identification data. Theor. Biol. Forum 2014, 107, 123–142. [Google Scholar] [PubMed]
- Ohsumi, S. Examination on age determination of the fin whale. Sci. Rep. Whales Res. Inst. 1964, 18, 49–88. [Google Scholar]
- Geraci, J.R.; Lounsbury, V.J. Marine Mammals Ashore: A Field Guide for Strandings, 2nd ed.; Texas A&M University Sea Grant Publication: College Station, TX, USA, 2005; Volume 1, p. 344. [Google Scholar]
- Fioravanti, T.; Maio, N.; Latini, L.; Splendiani, A.; Guarino, F.M.; Mezzasalma, M.; Petraccioli, A.; Cozzi, B.; Mazzariol, S.; Centelleghe, C.; et al. Nothing is as it seems: Genetic analyses on stranded fin whales unveil the presence of a fin-blue whale hybrid in the Mediterranean Sea (Balaenopteridae). Eur. Zool. J. 2022, 89, 590–600. [Google Scholar] [CrossRef]
- Zeileis, A.; Leisch, F.; Hornik, K.; Kleiber, C. Strucchange: An R Package for Testing for Structural Change in Linear Regression Models. J. Stat. Softw. 2002, 7, 1–38. [Google Scholar] [CrossRef]
- Zeileis, A.; Kleiber, C.; Krämer, W.; Hornik, K. Testing and Dating of Structural Changes in Practice. Comput. Stat. Data Anal. 2003, 44, 109–123. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 28 July 2022).
- Hamed, K.H. Exact distribution of the Mann-Kendall trend test statistic for persistent data. J. Hydrol. 2009, 365, 86–94. [Google Scholar] [CrossRef]
- Getis, A.; Ord, J.K. The Analysis of Spatial Association by Use of Distance Statistics. In Perspectives on Spatial Data Analysis. Advances in Spatial Science; Anselin, L., Rey, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar] [CrossRef]
- Ord, J.K.; Getis, A. Local Spatial Autocorrelation Statistics: Distributional Issues and an Application. Geogr. Anal. 1995, 27, 286–306. [Google Scholar] [CrossRef]
- Bearzi, G.; Pierantonio, N.; Affronte, M.; Holcer, D.; Maio, N.; Notarbartolo di Sciara, G. Overview of sperm whale Physeter microcephalus mortality events in the Adriatic Sea, 1555–2009. Mamm. Rev. 2011, 41, 276–293. [Google Scholar] [CrossRef]
- UNEP/MAP. Mediterranean Strategy for Sustainable Development 2016–2025; Plan Bleu, Regional Activity Centre: Valbonne, France, 2016. [Google Scholar]
- Pace, D.S.; Giacomini, G.; Campana, I.; Paraboschi, M.; Pellegrino, G.; Silvestri, M.; Alessi, J.; Angeletti, D.; Cafaro, V.; Pavan, G.; et al. An integrated approach for cetacean knowledge and conservation in the central Mediterranean Sea using research and social media data sources. Aquatic. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 1302–1323. [Google Scholar] [CrossRef]
- Moore, J.E.; Barlow, J.P. Declining Abundance of Beaked Whales (Family Ziphiidae) in the California Current Large Marine Ecosystem. PLoS ONE 2013, 8, e52770. [Google Scholar] [CrossRef] [Green Version]
- Peltier, H.; Dabin, W.; Daniel, P.; Canneyt, O.V.; Dorémus, G.; Huon, M.; Ridoux, V. The significance of stranding data as indicators of cetacean population at sea: Modelling the drift of cetacean carcasses. Ecol. Indic. 2012, 18, 278–290. [Google Scholar] [CrossRef]
- Panigada, S.; Lauriano, G.; Burt, L.; Pierantonio, N.; Donovan, G. Monitoring winter and summer abundance of cetaceans in the Pelagos Sanctuary (northwestern Mediterranean Sea) through aerial surveys. PLoS ONE 2011, 6, e22878. [Google Scholar] [CrossRef] [PubMed]
- Panigada, S.; Araujo, H.; Belmont, J.; Cañadas, A.; David, L.; Di-Méglio, N.; Dorémus, G.; Gonzalvo, J.; Holčer, D.; Laran, S.; et al. The ACCOBAMS Survey Initiative: The first synoptic survey of the Mediterranean Sea. In Proceedings of the World Marine Mammal Conference, Barcelona, Spain, 9–12 December 2019. [Google Scholar]
- Pierantonio, N.; Bearzi, G. Review of fin whale mortality events in the Adriatic Sea (1728–2012), with a description of a previously unreported killing. Mar. Biodiv. Rec. 2012, 5, e109. [Google Scholar] [CrossRef]
- Stephens, G.; Akkaya Bas, A.; Hardy, J.; Awbery, T.; Rudd, L.; Arac, N.; Lyne, P. Sightings and stranding reports of fin whales (Balaenoptera physalus) in the Levantine Sea. J. Cetacean Res. Manag. 2021, 22, 55–59. [Google Scholar] [CrossRef]
- Sciacca, V.; Caruso, F.; Beranzoli, L.; Chierici, F.; De Domenico, E.; Embriaco, D.; Favali, P.; Giovanetti, G.; Larosa, G.; Marinaro, G.; et al. Annual Acoustic Presence of Fin Whale (Balaenoptera physalus) Offshore Eastern Sicily, Central Mediterranean Sea. PLoS ONE 2015, 10, e0141838. [Google Scholar] [CrossRef] [Green Version]
- Zellar, R.; Pulkkinen, A.; Moore, K.; Rousseaux, C.S.; Reeb, D. Oceanic and Atmospheric Correlations to Cetacean Mass Stranding Events in Cape Cod, Massachusetts, USA. Geophys. Res. Lett. 2021, 48, e2021GL093697. [Google Scholar] [CrossRef]
- Notarbartolo di Sciara, G.; Bearzi, G. Cetacean direct killing and live capture in the Mediterranean Sea. In Cetaceans in the Mediterranean and Black Seas: State of Knowledge and Conservation Strategies; Notarbartolo di Sciara, G., Ed.; ACCOBAMS: Monaco City, Monaco, 2002. [Google Scholar]
- Druon, J.N.; Panigada, S.; David, L.; Gannier, A.; Mayol, P.; Arcangeli, A.; Cañadas, A.; Laran, S.; Di Méglio, N.; Gauffier, P. Potential feeding habitat of fin whales in the western Mediterranean Sea: An environmental niche model. Mar. Ecol. Prog. Ser. 2012, 464, 289–306. [Google Scholar] [CrossRef] [Green Version]
- Gambell, R. Fin Whale Balaenoptera physalus (Linnaeus, 1758). In Handbook of Marine Mammals. The Sirenians and Baleen Whales; Ridgway, S.H., Harrison, R., Eds.; Academic Press: London, UK, 1985; Volume 3, pp. 171–192. [Google Scholar]
- Zanardelli, M.; Airoldi, S.; Bérubé, M.; Borsani, J.F.; Di-Meglio, N.; Gannier, A.; Hammond, P.S.; Jahoda, M.; Lauriano, G.; Notarbartolo di Sciara, G.; et al. Long-term photo-identification study of fin whales in the Pelagos Sanctuary (NW Mediterranean) as a baseline for targeted conservation and mitigation measures. Aquatic. Conserv. 2022, 32, 1457–1470. [Google Scholar] [CrossRef]
- Herr, H.; Viquerat, S.; Devas, F.; Lees, A.; Wells, L.; Gregory, B.; Giffords, T.; Beecham, D.; Meyer, B. Return of large fin whale feeding aggregations to historical whaling grounds in the Southern Ocean. Sci. Rep. 2022, 12, 9458. [Google Scholar] [CrossRef]
- Arrigoni, M.; Manfredi, P.; Panigada, S.; Bramanti, L.; Santangelo, G. Life-history tables of the Mediterranean fin whale from stranding data. Mar. Ecol. 2011, 32, 1–9. [Google Scholar] [CrossRef]
- Bearzi, G.; Pierantonio, N.; Bonizzoni, S.; Notarbartolo di Sciara, G.; Demma, M. Perception of a cetacean mass stranding in Italy: The emergence of compassion. Aquat. Conserv. Mar. Freshw. Ecosyst. 2010, 20, 644–654. [Google Scholar] [CrossRef]
- Dolman, S.; Williams-Grey, V.; Asmutis-Silvia, R.; Isaac, S. Vessel Collisions and Cetaceans: What Happens if They Don’t Miss the Boat; Science Report; Whale and Dolphin Conservation Society: Wiltshire, UK, 2006. [Google Scholar]
- Laist, D.W.; Knowlton, A.R.; Mead, J.G.; Collet, A.S.; Podesta, M. Collisions between ships and whales. Mar. Mam. Sci. 2001, 17, 35–75. [Google Scholar] [CrossRef]
- Jefferson, T.A.; Stacey, P.J.; Baird, R.W. A review of Killer Whale interactions with other marine mammals: Predation to co-existence. Mamm. Rev. 1991, 21, 151–180. [Google Scholar] [CrossRef]
- Ford, J.K.B.; Reeves, R.R. Fight or flight: Antipredator strategies of baleen whales. Mamm. Rev. 2008, 38, 50–86. [Google Scholar] [CrossRef]
- Williams, R.; Lusseauc, D.; Hammond, P.H. Estimating relative energetic costs of human disturbance to killer whales (Orcinus orca). Biol. Conserv. 2006, 133, 301–311. [Google Scholar] [CrossRef]
- Van der Hoop, J.; Corkeron, P.; Moore, M. Entanglement is a costly life-history stage in large whales. Ecol. Evol. 2017, 7, 92–106. [Google Scholar] [CrossRef] [Green Version]
- Fossi, M.C.; Casini, S.; Marsili, L. Potential toxicological hazard due to endocrine-disrupting chemicals on Mediterranean top predators: State of art, gender differences and methodological tools. Environ. Res. 2007, 104, 174–182. [Google Scholar] [CrossRef]
- Fossi, M.C.; Panti, C.; Guerranti, C.; Coppola, D.; Giannetti, M.; Marsili, L.; Minutoli, R. Are baleen whales exposed to the threat of microplastics? A case study of the Mediterranean fin whale (Balaenoptera physalus). Mar. Pollut. Bull. 2012, 64, 2374–2379. [Google Scholar] [CrossRef]
- Cózar, A.; Sanz-Martín, M.; Martí, E.; González-Gordillo, J.I.; Ubeda, B.; Gálvez, J.Á.; González-Gordillo, J.I.; Ubeda, B.; Gálvez, J.Á.; Irigoien, X.; et al. Plastic Accumulation in the Mediterranean Sea. PLoS ONE 2015, 10, e0121762. [Google Scholar] [CrossRef] [Green Version]
- Fossi, M.C.; Marsili, L.; Baini, M.; Giannetti, M.; Coppola, D.; Guerranti, C.; Caliani, I.; Minutoli, R.; Lauriano, G.; Finoia, M.G.; et al. Fin whales and microplastics: The Mediterranean Sea and the Sea of Cortez scenarios. Environ. Pollut. 2016, 209, 68–78. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Description | Value | % |
|---|---|---|---|
| No. of animals | 67 | ||
| Sex | Male | 12 | 17.91 |
| Female | 10 | 14.92 | |
| Unknown | 45 | 67.16 | |
| Life stage | Fetus/Calf/Newborn | 8 | 11.94 |
| Immature | 16 | 23.88 | |
| Sub-adult/Adult/Mature | 29 | 43.28 | |
| NA | 14 | 20.89 | |
| Total length (m) | Minimum | 5.50 | |
| Maximum | 21.50 | ||
| Cause of death | Hypothetical | 19 | 28.35 |
| Unknown | 48 | 71.64 | |
| Hypothetical human-related cause of death | Killing * | 16 | 84.21 |
| Bycatch | 2 | 10.52 | |
| Ship strike | 1 | 5.26 | |
| Season | Autumn | 16 | 23.88 |
| Winter | 16 | ||
| NA | 14 | 20.89 | |
| Summer | 14 | ||
| Spring | 7 | 10.44 | |
| Region | Liguria | 17 | 25.37 |
| Tuscany | 15 | 22.38 | |
| Sardinia | 10 | 14.92 | |
| Lazio | 7 | 10.44 | |
| Campania | 5 | 7.46 | |
| Puglia | 5 | ||
| Sicily | 3 | 4.47 | |
| Calabria | 2 | 2.98 | |
| Friuli-Venezia Giulia | 2 | ||
| Abruzzo | 1 | 1.49 | |
| Carcass/Body conservation status | Alive (Code 1)/dead (Code 2) | 2 | 2.98 |
| NA | 65 | 97.01 |
| Variable | Description | Value | % |
|---|---|---|---|
| No. of animals | 112 | ||
| Sex | Male | 27 | 24.10 |
| Female | 38 | 33.92 | |
| Unknown | 47 | 41.96 | |
| Life stage | Fetus/Calf/Newborn | 32 | 28.57 |
| Immature | 47 | 41.96 | |
| Sub-adult/Adult/Mature | 24 | 21.42 | |
| NA | 9 | 8.03 | |
| Total length (m) * | Minimum | 2.70 | |
| Maximum | 19.77 | ||
| Cause of death | Hypothetical | 35 | 31.25 |
| Unknown | 77 | 68.75 | |
| Hypothetical cause of death | Human-related | 26 | 74.28 |
| Natural/Biological | 9 | 25.71 | |
| Human related cause of death | Ship strike | 23 | 88.46 |
| Bycatch/entanglement | 3 | 11.53 | |
| Season | Autumn | 34 | 30.35 |
| Spring | 28 | 25.00 | |
| Summer | 25 | 22.32 | |
| Winter | 25 | ||
| Region | Tuscany | 28 | 25.00 |
| Sardinia | 20 | 17.85 | |
| Liguria | 18 | 16.07 | |
| Calabria | 11 | 9.82 | |
| Campania | 9 | 8.03 | |
| Sicily | 9 | ||
| Puglia | 6 | 5.35 | |
| Lazio | 5 | 4.46 | |
| Emilia-Romagna | 2 | 1.78 | |
| Friuli-Venezia Giulia | 1 | 0.89 | |
| Marche | 1 | ||
| Uncertain location | 2 | 1.78 | |
| Carcass/Body conservation (°) | Advanced decomp. (Code 4) | 54 | 48.21 |
| NA | 34 | 30.35 | |
| Fresh (Code 2) | 15 | 13.39 | |
| Alive (Code 1)/dead (Code 2) | 8 | 7.14 |
| Death Cause/Life Stage | Sub-Adult/Adult/Mature | Immature | Fetus/Calf/Newborn |
|---|---|---|---|
| Natural/Biological | 3 | 2 | 4 |
| Human related | 6 | 17 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manfrini, V.; Pierantonio, N.; Giuliani, A.; De Pascalis, F.; Maio, N.; Mancia, A. Fin Whale (Balaenoptera physalus) Mortality along the Italian Coast between 1624 and 2021. Animals 2022, 12, 3111. https://doi.org/10.3390/ani12223111
Manfrini V, Pierantonio N, Giuliani A, De Pascalis F, Maio N, Mancia A. Fin Whale (Balaenoptera physalus) Mortality along the Italian Coast between 1624 and 2021. Animals. 2022; 12(22):3111. https://doi.org/10.3390/ani12223111
Chicago/Turabian StyleManfrini, Valerio, Nino Pierantonio, Alessandro Giuliani, Federico De Pascalis, Nicola Maio, and Annalaura Mancia. 2022. "Fin Whale (Balaenoptera physalus) Mortality along the Italian Coast between 1624 and 2021" Animals 12, no. 22: 3111. https://doi.org/10.3390/ani12223111
APA StyleManfrini, V., Pierantonio, N., Giuliani, A., De Pascalis, F., Maio, N., & Mancia, A. (2022). Fin Whale (Balaenoptera physalus) Mortality along the Italian Coast between 1624 and 2021. Animals, 12(22), 3111. https://doi.org/10.3390/ani12223111