Macroalgal Endophytes from the Atlantic Coast of Canada: A Potential Source of Antibiotic Natural Products?

Abstract
:1. Introduction
2. Experimental Section
2.1. Algal Collection
2.2. Surface Sterilization of Algae and Culture Techniques

| Algal Species | Sterilant Immersion Duration (Seconds) | |
|---|---|---|
| Bleach (5.25%) | Ethanol (70%) | |
| Chondrus crispus | 5 | 10 |
| Devaleraea ramentacea | 5 | 10 |
| Mastocarpus stellatus | 5 | 15 |
| Palmaria palmata | 0 | 15 |
| Polysiphonia lanosa | 0 | 10 |
| Porphyra purpurea | 0 | 10 |
| Porphyra umbilicalis | 0 | 10 |
| Ascophyllum nodosum | 0 | 15 |
| Fucus spiralis | 0 | 15 |
| Fucus vesiculosus | 0 | 15 |
| Saccharina latissima | 5 | 15 |
| Spongomorpha arcta | 0 | 20 |
| Ulva intestinalis | 0 | 20 |
| Ulva lactuca | 0 | 20 |
2.3. Isolation of Endophytes
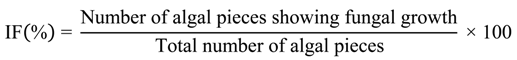
2.4. Identification of Fungi
2.5. Preparation of Extracts
2.6. Antibacterial and Antifungal Activity Assay
2.7. Statistical Analyses
3. Results and Discussion
3.1. Isolation of Endophytic Fungi from Marine Algae
| Seaweed Species | Total Algae Segments | Pieces with Endophytic Growth | Isolation Frequency (%) | Distinct Fungal Taxa |
|---|---|---|---|---|
| Chondrus crispus | 246 | 2 | 0.08 | 2 |
| Devaleraea ramentacea | 108 | 46 | 43 | 13 |
| Mastocarpus stellatus | 98 | 32 | 33 | 9 |
| Palmaria palmata | 167 | 146 | 87 | 7 |
| Polysiphonia lanosa | 67 | 12 | 18 | 5 |
| Porphyra purpurea | 85 | 17 | 20 | 2 |
| Porphyra umbilicalis | 217 | 156 | 72 | 3 |
| Ascophyllum nodosum | 152 | 16 | 11 | 1 |
| Fucus spiralis | 192 | 16 | 8 | 4 |
| Fucus vesiculosus | 109 | 6 | 6 | 1 |
| Saccharina latissima | 1138 | 95 | 8 | 13 |
| Spongomorpha arcta | 184 | 49 | 27 | 11 |
| Ulva intestinalis | 117 | 24 | 21 | 5 |
| Ulva lactuca | 201 | 15 | 8 | 3 |
| Total | 3081 | 632 | 26 | 79 |
| Host species | Isolate | Species | Accession number |
|---|---|---|---|
| Chondrus crispus | AF8-053 | Aspergillus sp. I | KF572134 |
| AF8-129 | Penicillium crustosum I | KF572146 | |
| Devaleraea ramentacea | AF8-079 | Cladosporium sp. | KF572136 |
| AF8-081 | Septate Pigmented I | - | |
| AF8-083 | Septate Pigmented II | - | |
| AF8-085 | Sterile Hyaline I | - | |
| AF8-087 | Sterile Hyaline II | - | |
| AF8-089 | Botrytis sp. I | - | |
| AF8-091 | Penicillium decumbens I | KF572138 | |
| AF8-093 | Septate Pigmented III | - | |
| AF8-095 | Aureobasidium pullulans I | - | |
| Devaleraea ramentacea | AF8-109 | Trametes versicolor | KF572133 |
| AF8-111 | White Fluffy II | - | |
| AF8-113 | Coniothyrium sp. | KF572141 | |
| AF8-115 | Botrytis sp. II | - | |
| Mastocarpus stellatus | AF8-055 | Penicillium sp. | - |
| AF8-057 | Red Yeast I | - | |
| AF8-059 | Septate Pigmented IV | - | |
| AF8-061 | Red Yeast II | - | |
| AF8-063 | Sterile Hyaline III | - | |
| AF8-065 | Sterile Hyaline IV | - | |
| AF8-067 | Coelomycete I | - | |
| AF8-101 | Penicillium decumbens II | KF572139 | |
| Palmaria palmata | AF8-069 | Sterile Hyaline V | - |
| AF8-071 | Hypoxylon sp. | KF572135 | |
| AF8-073 | Penicillium decumbens III | KF572149 | |
| AF8-075 | Penicillium chrysogenum I | KF572137 | |
| AF8-077 | Helicomyces sp. | - | |
| AF8-103 | Penicillium crustosum II | KF572147 | |
| AF8-105 | Botrytis sp. III | - | |
| AF8-107 | Sterile Hyaline VI | - | |
| AF8-123 | Sterile Hyaline VII | - | |
| Polysiphonia lanosa | AF8-097 | Sterile Hyaline VIII | - |
| AF8-099 | Aureobasidium pullulans II | KF572140 | |
| AF8-117 | Sterile Hyaline IX | - | |
| AF8-119 | Botryotinia fuckeliana | KF572148 | |
| AF8-121 | Sterile Hyaline X | - | |
| Porphyra purpurea | AF8-125 | Sterile Hyaline XI | - |
| AF8-127 | Sterile Hyaline XII | - | |
| Porphyra umbilicalis | AF8-049 | Sterile Hyaline XIII | - |
| AF8-051 | Sterile Hyaline XIV | - | |
| AF8-131 | Aspergillus sydowii | KF572145 | |
| Ascophyllum nodosum | AF1-141B | Aspergillus sp. II | - |
| Fucus spiralis | AF1-021A | Black Hyaline I | - |
| AF1-021B | Septate Pigmented V | - | |
| AF1-021D | Pigmented Hyaline I | - | |
| AF1-021G | Sterile Hyaline XV | - | |
| Fucus vesiculosus | AF2-059A | Sterile Hyaline XXI | - |
| Saccharina latissima | AF1-029A2 | White Hyaline IV | - |
| AF1-029B2 | Black Hyaline II | - | |
| AF1-073C | Penicillium chrysogenum II | KF572144 | |
| AF1-073D | Aspergillus sp. III | - | |
| AF1-073E | White Hyaline I | - | |
| AF1-073G | Sterile Beige I | - | |
| AF1-073N | Sterile Beige II | - | |
| AF2-055B | Penicillium soppii I | KF572151 | |
| AF2-055F | Sterile Hyaline XVI | - | |
| AF2-055G | White Hyaline II | - | |
| AF2-055O | Penicillium chrysogenum III | KF572150 | |
| AF2-055P | Pigmented Hyaline IV(Red) | - | |
| Spongomorpha arcta | AF1-037C2 | Sterile Hyaline XVII | - |
| AF1-037F | Sterile Beige III | - | |
| AF1-153A | Pigmented Hyaline VII | - | |
| AF1-153B | Sterile Hyaline XVIII | - | |
| AF1-153C | White Hyaline III | - | |
| AF1-153D | Penicillium spinulosum | KF572143 | |
| AF1-153H | Black Hyaline III | - | |
| AF1-153J | Pigmented Hyaline VI | - | |
| AF1-153L | Pigmented Hyaline V | - | |
| AF1-153M | Penicillium sp. | KF572152 | |
| AF1-153N | Penicillium soppii II | - | |
| Ulva intestinalis | AF2-063C | Sterile Hyaline XIX | - |
| AF2-063E | Septate Pigmented VIII | - | |
| AF2-063F | Pigmented Hyaline VIII | - | |
| AF2-063G | Aspergillus sp. V | - | |
| AF2-063H | Septate Pigmented VI | - | |
| Ulva lactuca | AF1-033A2 | Septate Pigmented VII | - |
| AF1-033B | Sterile Hyaline XX | - | |
| AF1-033C | Penicillium chrysogenum IV | KF572142 |
3.2. Antimicrobial Screening Results of Algal-Derived Fungal Endophytes
| Fungal Taxa | Host Alga | Extract 1 Source | Inhibition (%) 2 | ||
|---|---|---|---|---|---|
| SA 3 | PA 3 | CA 3 | |||
| Aspergillus sp. I | C. crispus | Mycelia | - | 18 ± 1 | - |
| Aspergillus sp. I | C. crispus | Media | 28 ± 3 | - | 15 ± 3 |
| Penicillium crustosum I | C. crispus | Media | 43 ± 1 | - | - |
| Cladosporium sp. | D. ramentacea | Mycelia | 36 ± 1 | 9 ± 1 | - |
| Septate Pigmented I | D. ramentacea | Mycelia | - | - | 17 ± 2 |
| Sterile Hyaline I | D. ramentacea | Mycelia | - | 20 ± 1 | - |
| Sterile Hyaline I | D. ramentacea | Media | 29 ± 2 | - | - |
| Botrytis sp. I | D. ramentacea | Mycelia | 94 ± 1 | - | - |
| P. decumbens I | D. ramentacea | Mycelia | 36 ± 4 | - | 98 ± 1 |
| P. decumbens I | D. ramentacea | Media | - | - | 99 ± 1 |
| Septate Pigmented III | D. ramentacea | Mycelia | 47 ± 4 | - | - |
| Aureobasidium pullulans I | D. ramentacea | Media | 82 ± 4 | 51 ± 2 | 13 ± 2 |
| Trametes versicolor | D. ramentacea | Mycelia | - | 17 ± 2 | - |
| White Fluffy II | D. ramentacea | Media | - | - | 12 ± 1 |
| Coniothyrium sp. | D. ramentacea | Mycelia | 31 ± 1 | - | - |
| Coniothyrium sp. | D. ramentacea | Media | 25 ± 2 | 28 ± 2 | - |
| Botrytis sp. II | D. ramentacea | Mycelia | - | - | 14 ± 1 |
| Botrytis sp. II | D. ramentacea | Media | 97 ± 1 | - | - |
| Red Yeast I | M. stellatus | Mycelia | 31 ± 2 | - | - |
| Septate Pigmented IV | M. stellatus | Mycelia | - | 16 ± 3 | - |
| Septate Pigmented IV | M. stellatus | Media | 23 ± 1 | - | 70 ± 3 |
| P. decumbens II | M. stellatus | Media | - | - | 12 ± 1 |
| Sterile Hyaline V | P. palmata | Mycelia | - | 11 ± 1 | 15 ± 4 |
| Sterile Hyaline V | P. palmata | Media | - | - | 16 ± 1 |
| Hypoxylon sp. | P. palmata | Mycelia | - | 13 ± 3 | 30 ± 3 |
| Hypoxylon sp. | P. palmata | Media | - | - | 18 ± 2 |
| P. decumbens III | P. palmata | Mycelia | - | - | 16 ± 5 |
| P. chrysogenum I | P. palmata | Media | 58 ± 3 | - | - |
| Helicomyces sp. | P. palmata | Mycelia | 21 ± 1 | - | - |
| Helicomyces sp. | P. palmata | Media | - | - | 21 ± 5 |
| P. crustosum II | P. palmata | Mycelia | 82 ± 5 | - | 22 ± 3 |
| P. crustosum II | P. palmata | Media | 39 ± 2 | - | - |
| Botrytis sp. III | P. palmata | Media | 92 ± 1 | - | - |
| Sterile Hyaline VI | P. palmata | Mycelia | - | - | 13 ± 2 |
| Sterile Hyaline VI | P. palmata | Media | 19 ± 6 | - | - |
| Sterile Hyaline VII | P. palmata | Media | 90 ± 2 | - | 24 ± 2 |
| Aureobasidium pullulans II | P. lanosa | Mycelia | 49 ± 1 | 34 ± 3 | - |
| Sterile Hyaline IX | P. lanosa | Media | - | - | 40 ± 5 |
| Botryotinia fuckeliana | P. lanosa | Mycelia | 96 ± 1 | - | 30 ± 2 |
| Botryotinia fuckeliana | P. lanosa | Media | 41 ± 2 | - | - |
| Sterile Hyaline X | P. lanosa | Mycelia | - | - | 20 ± 11 |
| Sterile Hyaline XI | P. purpurea | Mycelia | 22 ± 1 | - | - |
| Sterile Hyaline XI | P. purpurea | Media | 34 ± 9 | - | - |
| Sterile Hyaline XII | P. purpurea | Mycelia | 50 ± 1 | - | - |
| Sterile Hyaline XIII | P. umbilicalis | Media | - | - | 33 ± 1 |
| Sterile Hyaline XIV | P. umbilicalis | Mycelia | - | 14 ± 1 | 27 ± 6 |
| Sterile Hyaline XIV | P. umbilicalis | Media | 22 ± 5 | - | - |
| Aspergillus sydowii | P. umbilicalis | Media | 51 ± 3 | - | - |
| Aspergillus sp. II | A. nodosum | Mycelia | 66 ± 4 | - | - |
| Aspergillus sp. II | A. nodosum | Media | 85 ± 1 | - | 15 ± 3 |
| Black Hyaline I | F. spiralis | Media | 41 ± 2 | - | - |
| Septate Pigmented V | F. spiralis | Mycelia | 39 ± 3 | 8 ± 1 | - |
| Septate Pigmented V | F. spiralis | Media | 27 ± 3 | - | - |
| Pigmented Hyaline I | F. spiralis | Mycelia | 53 ± 4 | - | - |
| White Hyaline IV | S. latissima | Media | - | - | 30 ± 1 |
| Black Hyaline II | S. latissima | Mycelia | - | - | 13 ± 5 |
| P. chrysogenum II | S. latissima | Mycelia | 25 ± 4 | - | - |
| P. chrysogenum II | S. latissima | Media | 31 ± 4 | - | - |
| Aspergillus sp. III | S. latissima | Mycelia | 25 ± 1 | - | - |
| Sterile Beige II | S. latissima | Media | - | - | 14 ± 3 |
| White Hyaline II | S. latissima | Mycelia | 28 ± 3 | - | 27 ± 5 |
| White Hyaline II | S. latissima | Media | 21 ± 1 | - | 18 ± 9 |
| Pigmented Hyaline IV(Red) | S. latissima | Mycelia | - | 12 ± 1 | - |
| Pigmented Hyaline IV(Red) | S. latissima | Media | - | - | 17 ± 1 |
| Sterile Hyaline XVII | S. arcta | Mycelia | - | - | 13 ± 1 |
| Pigmented Hyaline VII | S. arcta | Media | - | 15 ± 3 | - |
| White Hyaline III | S. arcta | Mycelia | 27 ± 7 | - | - |
| White Hyaline III | S. arcta | Media | - | - | 13 ± 3 |
| Penicillium spinulosum | S. arcta | Mycelia | 65 ± 4 | - | - |
| Penicillium spinulosum | S. arcta | Media | 37 ± 1 | - | - |
| Pigmented Hyaline V | S. arcta | Media | 87 ± 3 | - | 82 ± 1 |
| Septate Pigmented VIII | S. arcta | Mycelia | - | 17 ± 1 | - |
| Septate Pigmented VIII | S. arcta | Media | 36 ± 2 | - | - |
| Penicillium soppii II | S. arcta | Media | - | 9 ± 3 | - |
| Sterile Hyaline XIX | U. intestinalis | Media | - | 9 ± 2 | 22 ± 2 |
| Septate Pigmented VI | U. intestinalis | Mycelia | - | - | 13 ± 2 |
| Septate Pigmented VII | U. lactuca | Mycelia | - | - | 18 ± 2 |
| Sterile Hyaline XX | U. lactuca | Mycelia | - | 24 ± 3 | - |
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Appelbaum, P.C. 2012 and beyond: Potential for the start of a second pre-antibiotic era? J. Antimicrob. Chemother. 2012, 67, 2062–2068. [Google Scholar] [CrossRef]
- Ashkenazi, S. Beginning and possibly the end of the antibiotic era. J. Paediatr. Child Health 2012, 49, 179–182. [Google Scholar]
- Spellberg, B.; Bartlett, J.G.; Gilbert, D.N. The future of antibiotics and resistance. N. Engl. J. Med. 2013, 368, 299–302. [Google Scholar] [CrossRef]
- Chopra, I. The 2012 garrod lecture: Discovery of antibacterial drugs in the 21st century. J. Antimicrob. Chemother. 2013, 68, 496–505. [Google Scholar] [CrossRef]
- Butler, M.S.; Cooper, M.A. Screening strategies to identify new antibiotics. Curr. Drug Targets 2012, 13, 373–387. [Google Scholar] [CrossRef]
- Debbab, A.; Aly, A.H.; Proksch, P. Endophytes and associated marine derived fungi-ecological and chemical perspectives. Fungal Divers. 2012, 57, 45–83. [Google Scholar] [CrossRef]
- Suryanarayanan, T.S. Fungal Endosymbionts of Seaweeds. In Biology of Marine Fungi; Raghukumar, C., Ed.; Springer: Berlin, Germany, 2012; pp. 53–69. [Google Scholar]
- Jones, E.B.G.; Pang, K.-L.; Stanley, S.J. Fungi from Marine Algae. In Marine Fungi and Fungal-Like Organisms; Jones, E.B.G., Pang, K.-L., Eds.; Walter de Gruyter: Berlin, Germany, 2012; pp. 329–344. [Google Scholar]
- Suryanarayanan, T.S.; Venkatachalam, A.; Thirunavukkarasu, N.; Ravishankar, J.P.; Doble, M.; Geetha, V. Internal mycobiota of marine macroalgae from the tamilnadu coast: Distribution, diversity and biotechnological potential. Bot. Mar. 2010, 53, 457–468. [Google Scholar]
- Schulz, B.; Boyle, C.; Draeger, S.; Rommert, A.K.; Rohn, K. Endophytic fungi: A source of novel biologically active secondary metabolites. Mycol. Res. 2002, 106, 996–1004. [Google Scholar] [CrossRef]
- Flewelling, A.J.; Johnson, J.A.; Gray, C.A. Isolation and bioassay screening of fungal endophytes from north atlantic marine macroalgae. Bot. Mar. 2013, 56, 287–297. [Google Scholar]
- Petrini, O.; Fisher, P.; Petrini, L. Fungal endophytes of bracken (Pteridium aquilinum), with some reflections on their use in biological control. Sydowia 1992, 44, 282–293. [Google Scholar]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Dobranic, J.K.; Johnson, J.A.; Alikhan, Q.R. Isolation of endophytic fungi from eastern larch (Larix laricina) leaves from new-brunswick, Canada. Can. J. Microbiol. 1995, 41, 194–198. [Google Scholar] [CrossRef]
- Suryanarayanan, T.S.; Kumaresan, V.; Johnson, J.A. Foliar fungal endophytes from two species of the mangrove Rhizophora. Can. J. Microbiol. 1998, 44, 1003–1006. [Google Scholar]
- Bates, C.R.; Saunders, G.; Chopin, T. Historical versus contemporary measures of seaweed biodiversity in the bay of fundy. Botany 2009, 87, 1066–1076. [Google Scholar] [CrossRef]
- South, G.R. A checklist of marine algae of eastern canada, 2nd revision. Can. J. Bot. 1984, 62, 680–704. [Google Scholar] [CrossRef]
- Fries, N.; Thorentolling, K. Identity of fungal endophyte of Ascophyllum with Mycosphaerella ascophylli established by means of fluorescent-antibody technique. Bot. Mar. 1978, 21, 409–411. [Google Scholar]
- Fries, N. Physiological characteristics of Mycosphaerella ascophylli, a fungal endophyte of the marine brown alga Ascophyllum nodosum. Physiol. Plant. 1979, 45, 117–121. [Google Scholar] [CrossRef]
- Fries, L. Ascophyllum nodosum (Phaeophyta) in Axenic culture and its response to the endophytic fungus Mycosphaerella ascophylli and epiphytic bacteria. J. Phycol. 1988, 24, 333–337. [Google Scholar]
- Garbary, D.J.; Gautam, A. The Ascophyllum/Polysiphonia/Mycosphaerella symbiosis. I. Population ecology of Mycosphaerella from Nova Scotia. Bot. Mar. 1989, 32, 181–186. [Google Scholar]
- Garbary, D.J.; Burke, J.; Tian, L.N. The Ascophyllum/Polysiphonia/Mycosphaerella symbiosis. II. Aspects of the ecology and distribution of Polysiphonia lanosa in Nova Scotia. Bot. Mar. 1991, 34, 391–401. [Google Scholar]
- Stanley, S.J. Observations on the seasonal occurance of marine endophytic and parasitic fungi. Can. J. Bot. 1992, 70, 2089–2096. [Google Scholar] [CrossRef]
- Garbary, D.J.; London, J.F. The Ascophyllum/Polysiphonial/Mycosphaerella symbiosis. V. Fungal infection protects A. nosodum from desiccation. Bot. Mar. 1995, 38, 529–533. [Google Scholar]
- Osterhage, C.; Kaminsky, R.; Konig, G.M.; Wright, A.D. Ascosalipyrrolidinone A, an antimicrobial alkaloid, from the obligate marine fungus Ascochyta salicorniae. J. Org. Chem. 2000, 65, 6412–6417. [Google Scholar] [CrossRef]
- Abdel-Lateff, A.; Fisch, K.M.; Wright, A.D.; Konig, G.M. A new antioxidant isobenzofuranone derivative from the algicolous marine fungus Epicoccum sp. Planta Med. 2003, 69, 831–834. [Google Scholar] [CrossRef]
- Deckert, R.; Garbary, D. Ascophyllum and its symbionts. VI. Microscopic characterization of the Ascophyllum nodosum (Phaeophyceae), Mycophycias ascophylli (Ascomycetes) symbiotum. Algae 2005, 20, 225–232. [Google Scholar] [CrossRef]
- Garbary, D.J.; Jamieson, M.M.; Fraser, S.J.; Ferguson, C.A.; Cranston, P.S. Ascophyllum (Phaeophyceae) and its symbionts. IX. A novel symbiosis between Halocladius variabilis (Chironomidae, Insecta) and Elachista fucicola (Elachistaceae, Phaeophyceae) from marine rocky shores of Nova Scotia. Symbiosis 2005, 40, 61–68. [Google Scholar]
- Schulz, B.; Draeger, S.; dela Cruz, T.E.; Rheinheimer, J.; Siems, K.; Loesgen, S.; Bitzer, J.; Schloerke, O.; Zeeck, A.; Kock, I.; et al. Screening strategies for obtaining novel, biologically active, fungal secondary metabolites from marine habitats. Bot. Mar. 2008, 51, 219–234. [Google Scholar]
- Xu, H.; Deckert, R.J.; Garbary, D.J. Ascophyllum and its symbionts. X. Ultrastructure of the interaction between A. nodosum (Phaeophyceae) and Mycophycias ascophylli (Ascomycetes). Botany 2008, 86, 185–193. [Google Scholar] [CrossRef]
- Webber, F.C. Observations on the structure, life history and biology of Mycosphaerella ascophylli. Trans. Br. Mycol. Soc. 1967, 50, 583–601. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Flewelling, A.J.; Ellsworth, K.T.; Sanford, J.; Forward, E.; Johnson, J.A.; Gray, C.A. Macroalgal Endophytes from the Atlantic Coast of Canada: A Potential Source of Antibiotic Natural Products? Microorganisms 2013, 1, 175-187. https://doi.org/10.3390/microorganisms1010175
Flewelling AJ, Ellsworth KT, Sanford J, Forward E, Johnson JA, Gray CA. Macroalgal Endophytes from the Atlantic Coast of Canada: A Potential Source of Antibiotic Natural Products? Microorganisms. 2013; 1(1):175-187. https://doi.org/10.3390/microorganisms1010175
Chicago/Turabian StyleFlewelling, Andrew J., Katelyn T. Ellsworth, Joseph Sanford, Erica Forward, John A. Johnson, and Christopher A. Gray. 2013. "Macroalgal Endophytes from the Atlantic Coast of Canada: A Potential Source of Antibiotic Natural Products?" Microorganisms 1, no. 1: 175-187. https://doi.org/10.3390/microorganisms1010175
APA StyleFlewelling, A. J., Ellsworth, K. T., Sanford, J., Forward, E., Johnson, J. A., & Gray, C. A. (2013). Macroalgal Endophytes from the Atlantic Coast of Canada: A Potential Source of Antibiotic Natural Products? Microorganisms, 1(1), 175-187. https://doi.org/10.3390/microorganisms1010175