Interaction of a Preventative Fungicide Treatment and Root Rot Pathogen on Ambrosia Beetle Attacks during a Simulated Flood Event



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Field Trials
2.2.1. Trial 1
2.2.2. Trial 2
2.3. Evaluation of Ethanol Emission from Fungicide Treated Redbuds
2.4. Data Analysis
3. Results
3.1. Root Health Scores
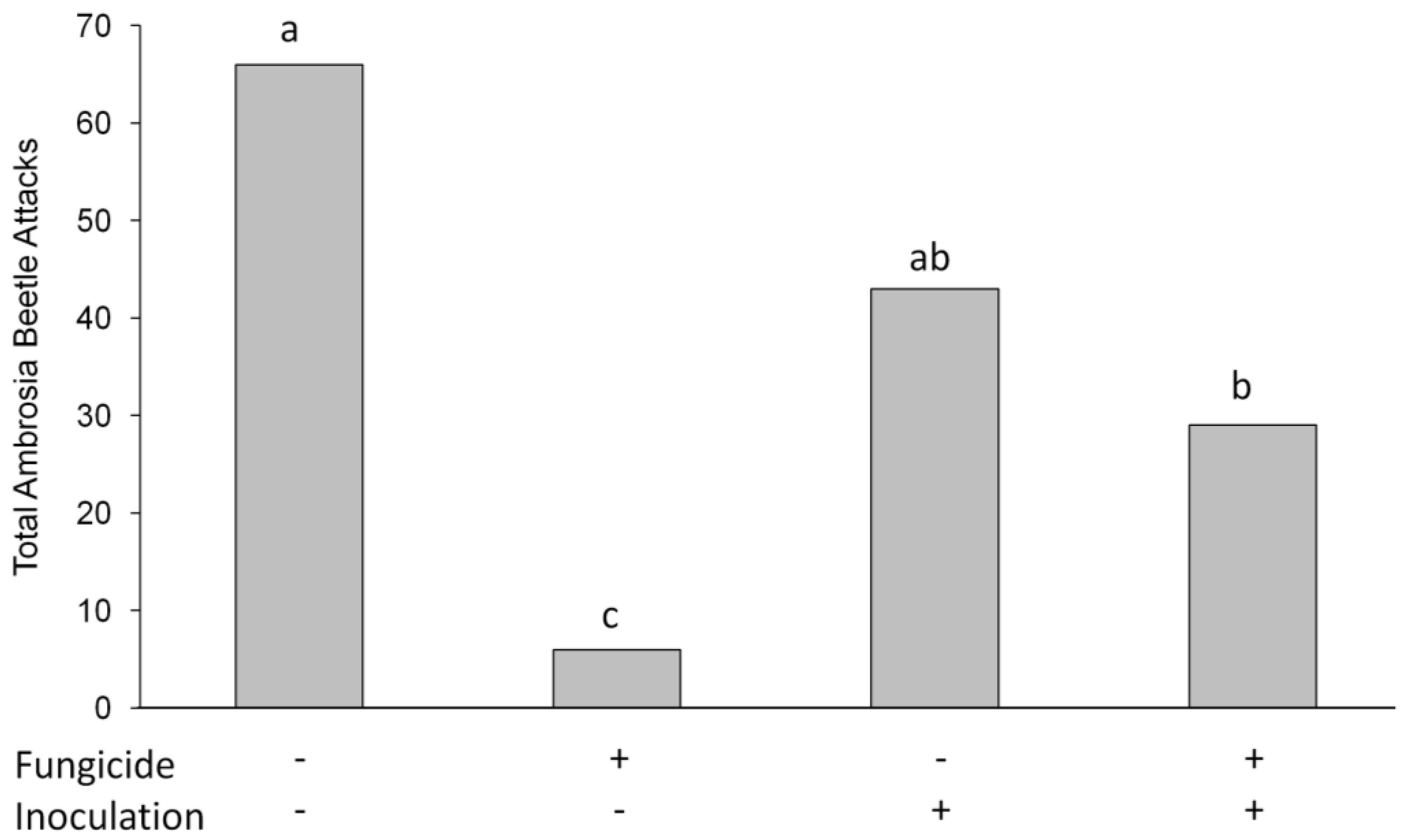
3.2. Ambrosia Beetle Attacks
3.3. Ambrosia Beetle Attacks Resulting in Gallery Formation
3.4. Ambrosia Beetle Galleries with Eggs
3.5. Evaluation of Ethanol Emission from Fungicide Treated Redbuds
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bratkovich, S.; Burban, L.; Katkovich, S.; Locey, C.; Pokorny, J.; Wiest, R. Flooding and Its Effect on Trees; Miscellaneous Publications: Newtown, PA, USA, 1993. [Google Scholar]
- Lebude, A.; Neal, J.; Fair, B.; Ranney, T.; Lauderdale, D.; Weathington, M.; Frank, S.; Meadows, I.; Villani, S.; Fulcher, A.; et al. Managing Storm and Disaster Damage in Landscapes and Nurseries. Available online: http://content.ces.ncsu.edu/managing-storm-and-disaster-damage-in-landscapes-and-nurseries (accessed on 13 June 2018).
- Kozlowski, T.T. Responses of woody plants to flooding and salinity. Tree Physiol. 1997, 1, 1–29. [Google Scholar] [CrossRef]
- Wilcox, W.F.; Mircetich, S.M. Effects of flooding duration on the development of Phytophthora root and crown rots of cherry. Phytopathology 1985, 75, 1451–1455. [Google Scholar] [CrossRef]
- Bourtsoukidis, E.; Kawaletz, H.; Radacki, D.; Schutz, S.; Hakola, H.; Hellen, H.; Noe, S.; Molder, I.; Ammer, C.; Bonn, B. Impact of flooding and drought conditions on the emission of volatile organic compounds of Quercus robur and Prunus serotine. Trees 2014, 28, 193–204. [Google Scholar] [CrossRef]
- Rottenberger, S.; Kleiss, B.; Kuhn, U.; Wolf, A.; Piedade, M.T.F.; Junk, W.; Kesselmeier, J. The exchange of flooding on the exchange of the volatile C2-compounds ethanol, acetaldehyde and acetic acid between leaves of Amazonian floodplain tree species and the atmosphere. Biogeosciences 2008, 5, 1085–1100. [Google Scholar] [CrossRef]
- Ranger, C.M.; Reding, M.E.; Persad, A.B.; Herms, D.A. Ability of stress-related volatiles to attract and induce attacks by Xylosandrus germanus and other ambrosia beetles. Agric. For. Entomol. 2010, 12, 177–185. [Google Scholar] [CrossRef]
- Ranger, C.M.; Reding, M.E.; Gandhi, K.J.K.; Oliver, J.B.; Schultz, P.B.; Canas, L.; Herms, D.A. Species dependent influence of (-)-α-pinene on attraction of ambrosia beetles (Coleoptera: Curculionidae: Scolytinae) to ethanol-baited traps in nursery agroecosystems. J. Econ. Entomol. 2011, 104, 574–579. [Google Scholar] [CrossRef] [PubMed]
- Ranger, C.M.; Gorzlancyk, A.M.; Addesso, K.M.; Oliver, J.B.; Reding, M.E.; Schultz, P.B.; Held, D.W. Conophthorin enhances the electroantennogram and field behavioral response of Xylosandrus germanus (Coleoptera: Curculionidae) to ethanol. Agric. For. Entomol. 2014, 16, 327–334. [Google Scholar] [CrossRef]
- Wood, S.L. The Bark and Ambrosia Beetles of North and Central America (Coleoptera: Scolytidae), A Taxonomic Monograph; Brigham Young University: Provo, UT, USA, 1982; pp. 56–77; 755–849. [Google Scholar]
- Beaver, R.A. Insect-fungus relationships in the bark and ambrosia beetles. In Insect-Fungus Interactions; Wilding, N., Collins, N.M., Hammond, P.M., Webber, J.F., Eds.; Academic Press: Cambridge, MA, USA, 1989; pp. 121–143. ISBN 0127518002. [Google Scholar]
- Ranger, C.M.; Biedermann, P.H.W.; Phuntumart, V.; Beligala, G.U.; Ghosh, S.; Palmquist, D.E.; Mueller, R.; Barnett, J.; Schultz, P.B.; Reding, M.E.; et al. Symbiont selection via alcohol benefits fungus-farming by ambrosia beetles. Proc. Natl. Acad. Sci. USA 2018, 115, 4447–4452. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.B.; Mannion, C.M. Ambrosia beetle (Coleoptera: Scolytidae) species attacking chestnut and captured in ethanol-baited traps in Middle Tennessee. Environ. Entomol. 2001, 30, 909–918. [Google Scholar] [CrossRef]
- Kelsey, R.G.; Beh, M.M.; Shaw, D.C.; Manter, D.K. Ethanol attracts Scolytid beetles to Phytophthora ramorum cankers on coast live oak. J. Chem. Ecol. 2013, 39, 494–506. [Google Scholar] [CrossRef] [PubMed]
- Ranger, C.M.; Reding, M.E.; Schultz, P.B.; Oliver, J.B. Influence of flood-stress on ambrosia beetle host-selection and implications for their management in a changing climate. Agric. For. Entomol. 2013, 15, 56–64. [Google Scholar] [CrossRef]
- Ranger, C.M.; Tobin, P.C.; Reding, M.E. Ubiquitous volatile compound facilitates efficient host location by a non-native ambrosia beetle. Biol. Invasions 2015, 17, 675–686. [Google Scholar] [CrossRef]
- Frank, S.D.; Ranger, C.M. Developing a media moisture threshold for nurseries to reduce tree stress and ambrosia beetle attacks. Environ. Entomol. 2016, 45, 1040–1048. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, K.A.; Johnson, G.R. Ornamental cherry tolerance of flooding and Phytophthora root rot. HortScience 1996, 31, 988–991. [Google Scholar]
- Blaker, N.S.; McDonald, J.D. Predisposing effects of soil moisture extremes on the susceptibility of Rhododendron to Phytophthora root and crown rot. Phytopathology 1981, 71, 831–834. [Google Scholar] [CrossRef]
- Hagan, A.; Mullen, J. Phytophthora Root Rot on Woody Ornamentals. Alabama Cooperative Extension System. 2000. Available online: http://www.aces.edu/pubs/docs/A/ANR-0571/ANR-0571.pdf (accessed on 13 June 2018).
- Benson, D.M.; Jones, R.K. Phytophthora Root Rot and Its Control on Established Woody Ornamentals. 2000. Available online: http://www.ces.ncsu.edu/depts/pp/notes/oldnotes/odin13/od13.htm (accessed on 13 June 2018).
- Solomon, J.D. Guide to Insect Borers in North American Broadleaf Trees and Shrubs; United States Department of Agricluture, Forest Service: Washington, DC, USA, 1995; Volume AH-706, p. 747.
- Kessler, K.J. An apparent symbiosis between Fusarium fungi and ambrosia beetles causes canker on black walnut stems. Plant Dis. Rep. 1974, 58, 1044–1047. [Google Scholar]
- Kovach, J.; Gorsuch, C.S. Survey of ambrosia beetle species infesting South Carolina peach orchards and a taxonomic key for the most common species. J. Agric. Entomol. 1985, 2, 238–247. [Google Scholar]
- A New Nursery and Landscape Pest in Tennessee. Available online: http://www.tnstate.edu/extension/documents/Camphor%20Shot%20Borer%20Final.pdf (accessed on 13 June 2018).
- Ranger, C.M.; Schultz, P.B.; Frank, S.D.; Chong, J.H.; Reding, M.E. Non-native ambrosia beetles as opportunistic exploiters of living but weakened trees. PLoS ONE 2015, 10, e0131496. [Google Scholar] [CrossRef] [PubMed]
- Stouthamer, R.; Rugman-Jones, P.; Eskalen, A.; Kabashima, J.; Dimson, M.; Gonzalez, A.; Arakelian, G.; Hodel, D.; Drill, S. Polypahgous Shot Hole Borer. UC Cooperative Extension. 2017. Available online: http://ucanr.edu/sites/socaloakpests/Polyphagous_Shot_Hole_Borer (accessed on 13 June 2018).
- Mizell, R.F.; Riddle, T.C. Evaluation of insecticides to control the Asian ambrosia beetle, Xylosandrus crassiusculus. In Proceedings of the 49th Annual Southern Nursey Association Research Conference, Atlanta, GA, USA, 11–12 August 2004. [Google Scholar]
- Corcobado, T.; Solla, A.; Madeira, M.A.; Moreno, G. Combined effects of soil properties and Phytophthora cinnamomi infections on Quercus ilex decline. Plant Soil 2013, 373, 403–413. [Google Scholar] [CrossRef]
- Reding, M.E.; Oliver, J.B.; Schultz, P.B.; Ranger, C.M.; Youssef, N.N. Ethanol injection of ornamental trees facilitates testing insecticide efficacy against ambrosia beetles (Coleoptera: Curculionidae: Scolytinae). J. Econ. Entomol. 2013, 106, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Ranger, C.M.; Schultz, P.B.; Reding, M.E.; Frank, S.D.; Palmquist, D.E. Flood stress as a technique to assess preventive insecticide and fungicide treatments for protecting trees against ambrosia beetles. Insects 2016, 7, 40. [Google Scholar] [CrossRef] [PubMed]
- Balba, H. Review of strobilurin fungicide chemicals. J. Environ. Sci. Health Part B 2007, 42, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Herms, S.; Seehaus, K.; Koehle, H.; Conrath, U. A strobilurin fungicide enhances the resistance of tobacco against tobacco mosaic virus and Pseudomonas syringae pv tabaci. Plant Physiol. 2002, 130, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Venancio, W.S.; Rodrigues, M.A.T.; Begliomini, E.; de Souza, N.L. Physiological effects of strobilurin fungicides on plants. Publ. UEPG Ciências Exatas e da Terra, Agrárias e Engenharias 2003, 9, 59–68. [Google Scholar]
- Abdul Jaleel, C.; Lakshmanan, G.M.A.; Gomathinayagam, M.; Panneerselvam, R. Triadimefon induced salt stress tolerance in Withania somnifera and its relationship to antioxidant defense system. S. Afr. J. Bot. 2008, 74, 126–132. [Google Scholar] [CrossRef]
- Han, S.H.; Kang, B.R.; Lee, J.H.; Lee, S.H.; Kim, I.S.; Kim, C.H.; Kim, Y.C. A trifloxystrobin fungicide induces systemic tolerance to abiotic stresses. Plant Pathol. J. 2012, 28, 101–106. [Google Scholar] [CrossRef]
- Grossmann, K.; Retzlaff, G. Bioregulatory effects of the fungicidal strobilurin kresoxim-methyl in wheat (Triticum aestivum). Pestic. Sci. 1997, 50, 11–20. [Google Scholar] [CrossRef]
- Wu, Y.X.; von Tiedemann, A. Impact of fungicides on active oxygen species and antioxidant enzymes in spring barley (Hordeum vulgare L.) exposed to ozone. Environ. Pollut. 2002, 116, 37–47. [Google Scholar] [CrossRef]
- Conrath, U.; Beckers, G.J.M.; Flors, V.; García-Agustín, P.; Jakab, G.; Mauch, F.; Newman, M.A.; Pieterse, C.M.J.; Poinssot, B.; Pozo, M.J.; et al. Priming: Getting ready for battle. Mol. Plant Microbe Interact. 2006, 19, 1062–1071. [Google Scholar] [CrossRef] [PubMed]
- Mayfield, A.E.; Barnard, E.L.; Smith, J.A.; Bernick, S.C.; Elckwort, J.M.; Dreaden, T.J. Effect of propiconazole on laurel wilt disease development in redbay trees and on the pathogen in vitro. Arboric. Urban For. 2008, 34, 317–324. [Google Scholar]
- McPherson, B.A.; Erbilgin, N.; Wood, D.L.; Svihra, P.; Storer, A.J.; Standiford, R.B. Attraction of ambrosia and bark beetles to coast live oaks infected by Phytophthora ramorum. Agric. For. Entomol. 2008, 10, 315–321. [Google Scholar] [CrossRef]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Addesso, K.; Baysal-Gurel, F.; Oliver, J.; Ranger, C.; O’Neal, P. Interaction of a Preventative Fungicide Treatment and Root Rot Pathogen on Ambrosia Beetle Attacks during a Simulated Flood Event. Insects 2018, 9, 83. https://doi.org/10.3390/insects9030083
Addesso K, Baysal-Gurel F, Oliver J, Ranger C, O’Neal P. Interaction of a Preventative Fungicide Treatment and Root Rot Pathogen on Ambrosia Beetle Attacks during a Simulated Flood Event. Insects. 2018; 9(3):83. https://doi.org/10.3390/insects9030083
Chicago/Turabian StyleAddesso, Karla, Fulya Baysal-Gurel, Jason Oliver, Christopher Ranger, and Paul O’Neal. 2018. "Interaction of a Preventative Fungicide Treatment and Root Rot Pathogen on Ambrosia Beetle Attacks during a Simulated Flood Event" Insects 9, no. 3: 83. https://doi.org/10.3390/insects9030083