Age-Dependent Developmental Response to Temperature: An Examination of the Rarely Tested Phenomenon in Two Species (Gypsy Moth (Lymantria dispar) and Winter Moth (Operophtera brumata))

Abstract
:1. Introduction
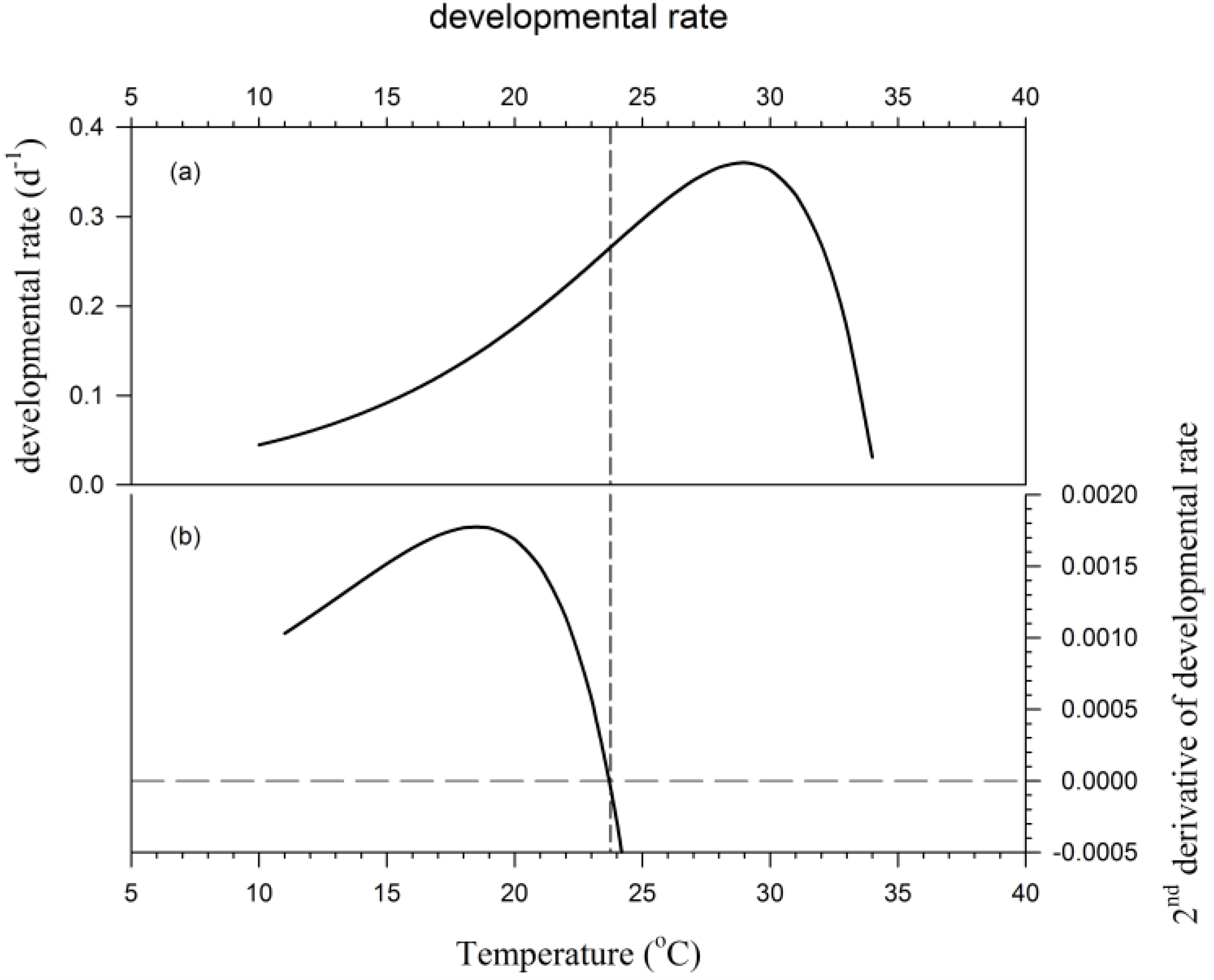
1.1. Phenology Modeling: The Basics
1.2. Stage-Dependent Developmental Response to Temperature
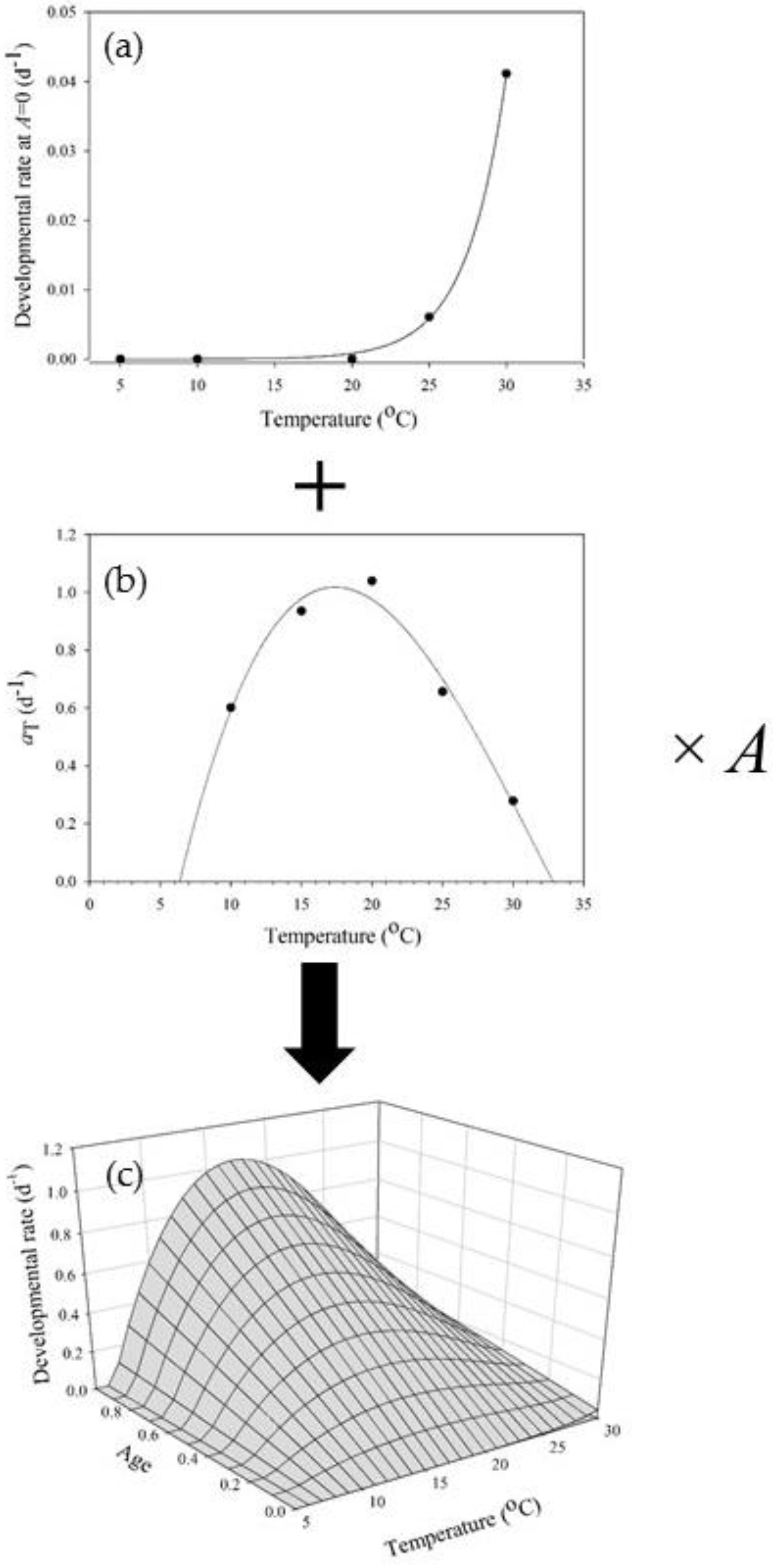
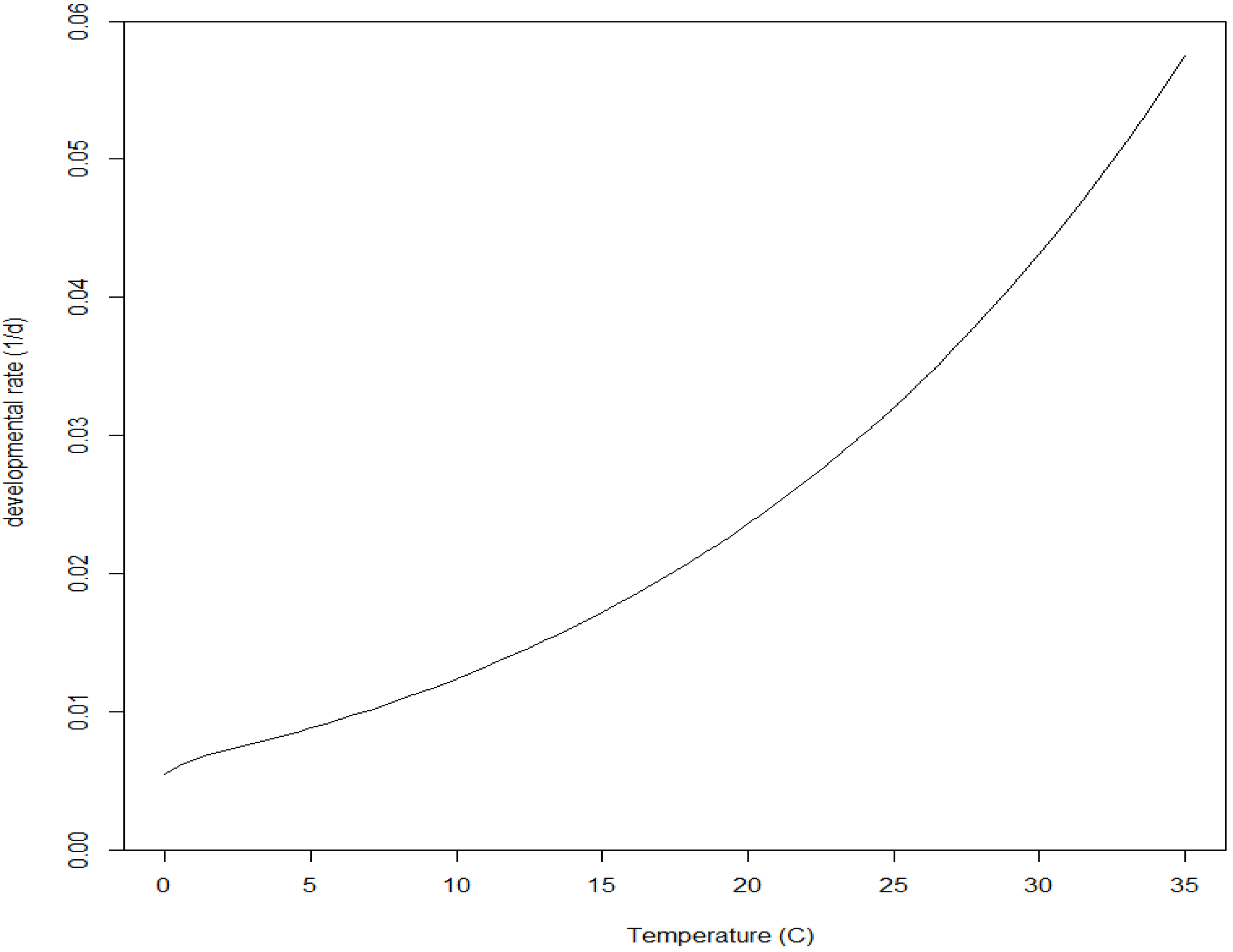
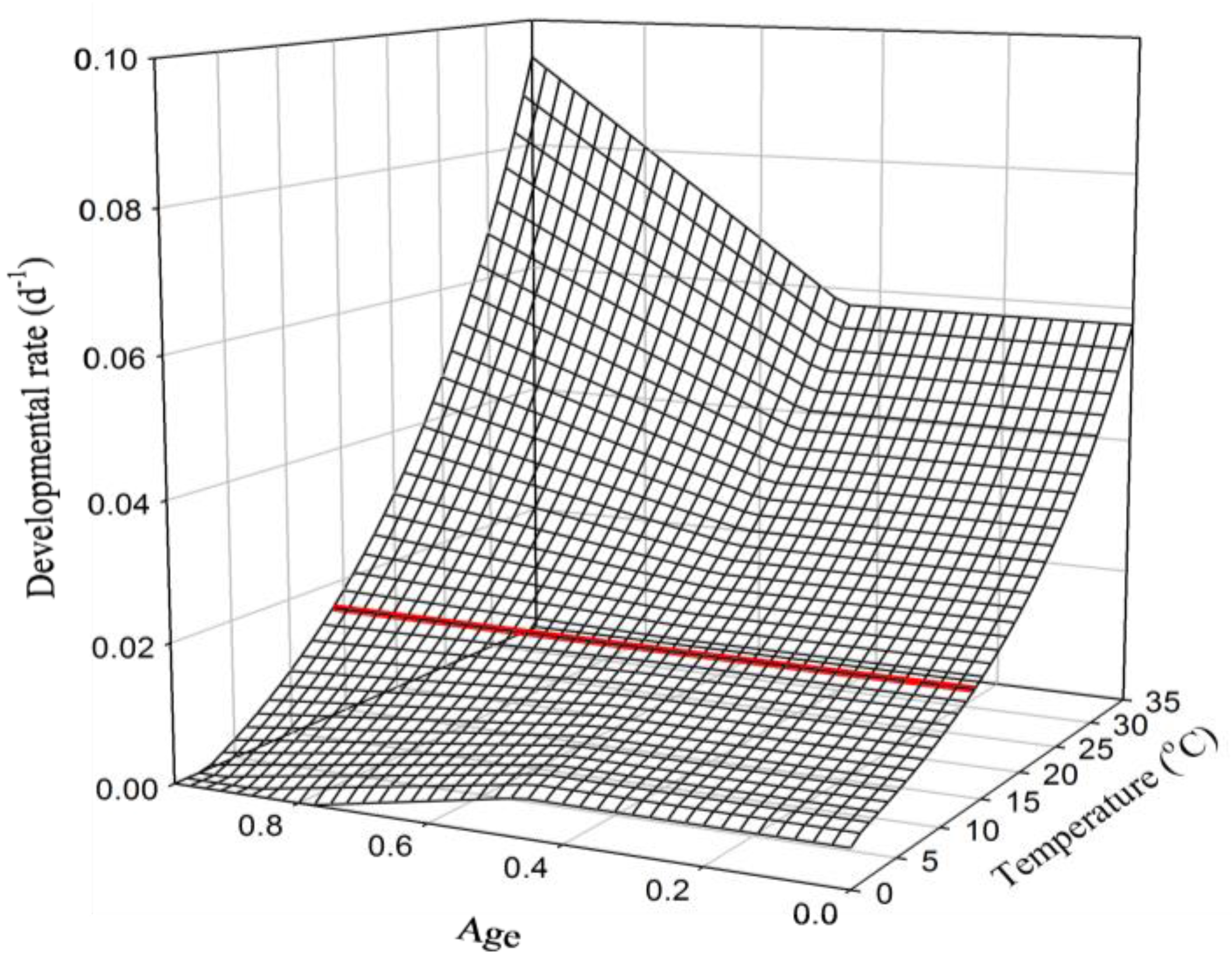
1.3. Age-Dependent Developmental Response to Temperature within a Life Stage
1.4. Age-Dependent Developmental Response to Temperature within a Life Phase (Sub-Stage)
2. Materials and Methods
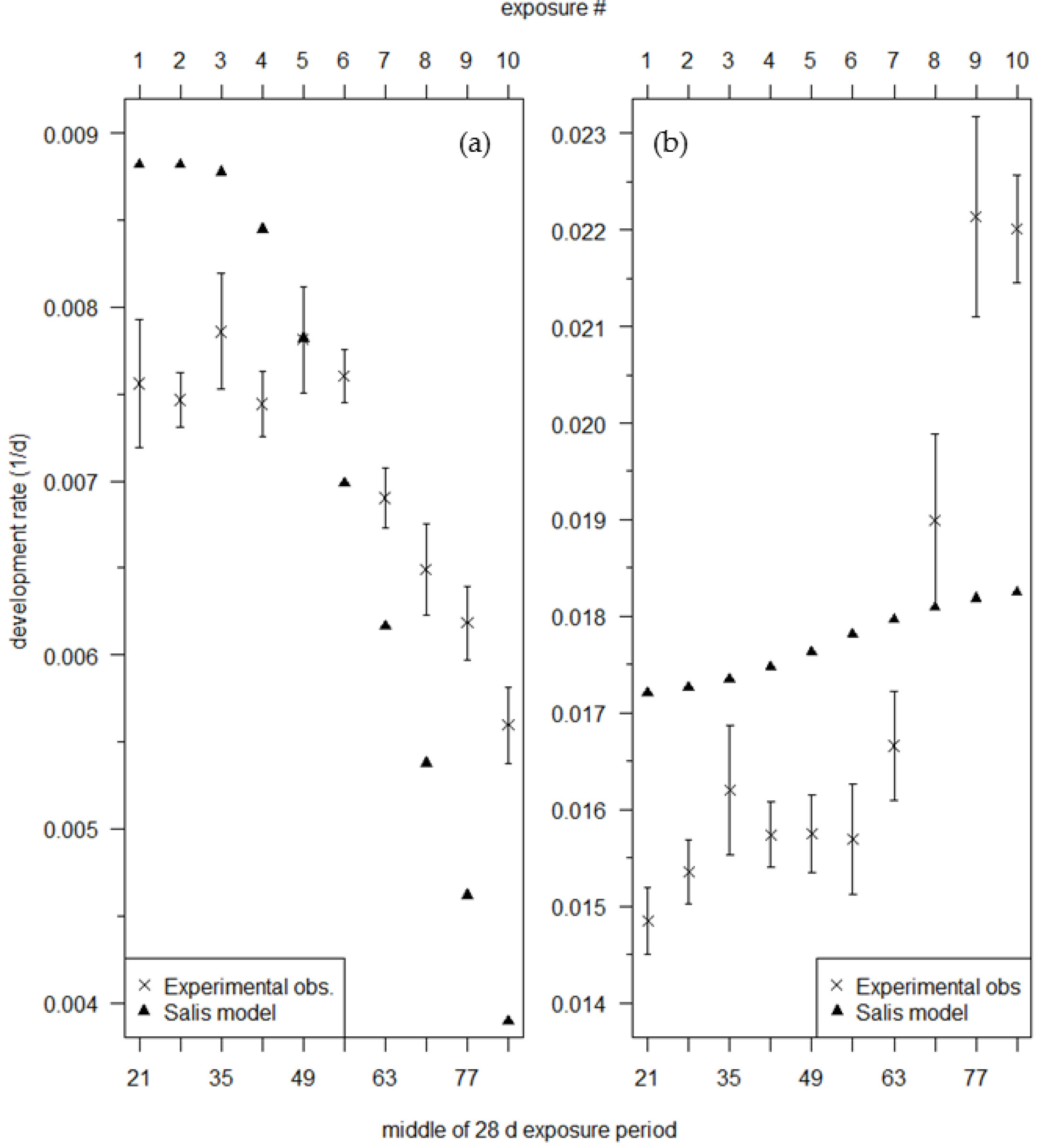
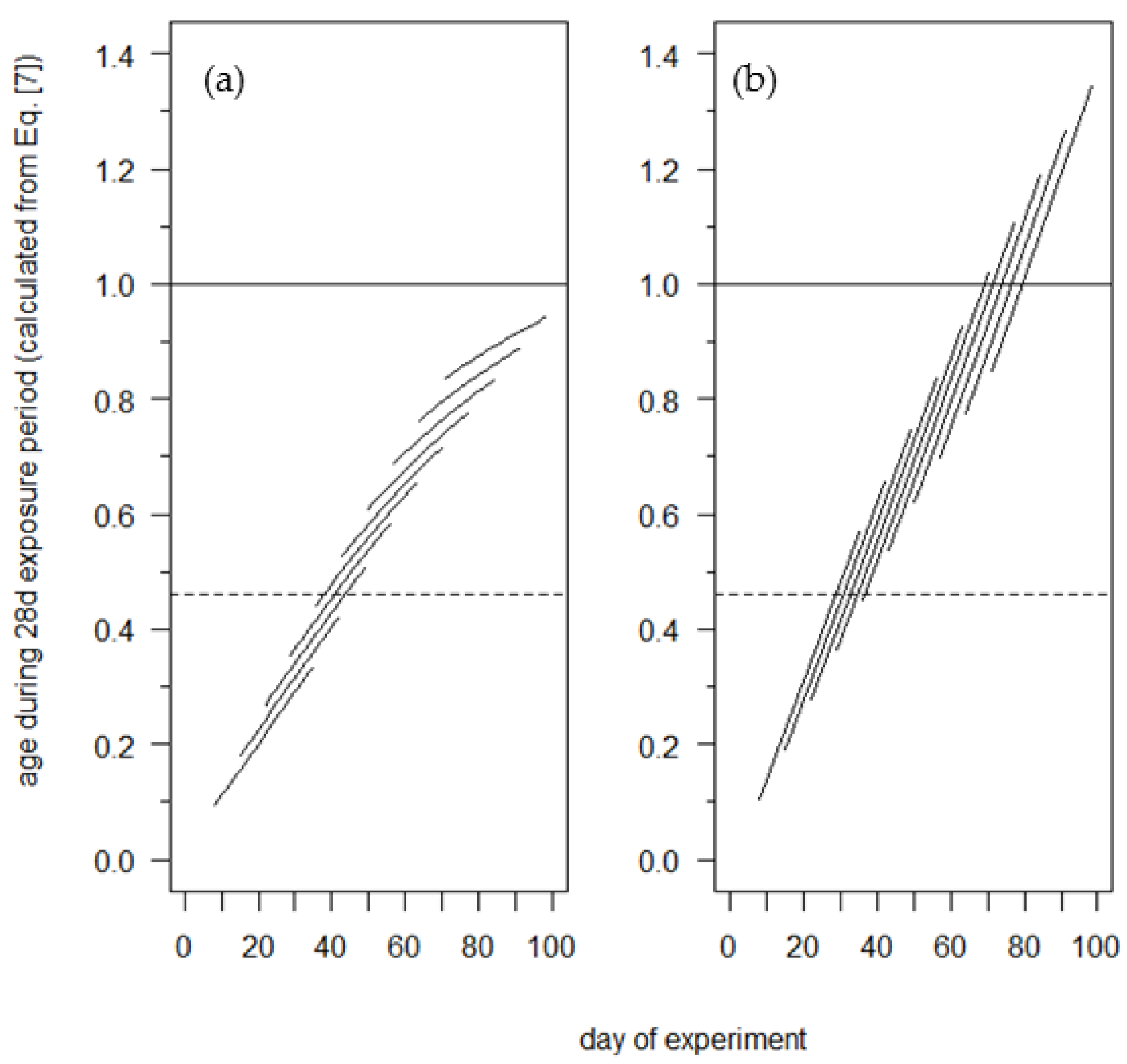
An Examination of the Winter Moth Egg Phenology Model: Discrepancies between Experimentally-Derived Rates and the Model Predictions
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Chen, X. Phenological Data, Networks, and Research: East Asia. In Phenology: An Integrative Environmental Science; Schwartz, M.D., Ed.; Kluwer Academic Publishers: London, UK, 2003; pp. 11–25. [Google Scholar]
- Réaumur, R.A.F. Obserations du thermomètre, Faites à Paris Pendant L’année 1735, Comparées Avec Celles Qui ont été Faites Sous la Ligne, à l’isle de France, à Alger et Quelques Unes de nos Isles de l’Amérique; Mémoires de l’Académie des Sciences de Paris; Académie Royale des Sciences: Paris, France, 1735. (In French) [Google Scholar]
- Damos, P.T.; Savopoulou-Soultani, M. Development and statistical evaluation of models in forecasting moth phenology of major lepidopterous peach pest complex for Integrated Pest Management programs. Crop Prot. 2010, 29, 1190–1199. [Google Scholar] [CrossRef]
- Moerkens, R.; Gobin, B.; Peusen, G.; Helsen, H.; Hilton, R.; Dib, H.; Suckling, D.M.; Leirs, H. Optimizing biocontrol using phenological day degree models: The European earwig in pipfruit orchards. Agric. For. Entomol. 2011, 13, 301–312. [Google Scholar] [CrossRef]
- Gray, D.R. The gypsy moth life stage model: Landscape-wide estimates of gypsy moth establishment using a multi-generational phenology model. Ecol. Model. 2004, 176, 155–171. [Google Scholar] [CrossRef]
- Logan, J.A.; Réqnière, J.; Gray, D.R.; Munson, A.S. Risk assessment in face of a changing environment: Gypsy moth and climate change in Utah. Ecol. Appl. 2007, 17, 101–117. [Google Scholar] [CrossRef]
- Gray, D.R. Risk reduction of an invasive insect by targeting surveillance efforts with the assistance of a phenology model and international maritime shipping routes and schedules. Risk Anal. 2015, 36, 914–925. [Google Scholar] [CrossRef] [PubMed]
- Gray, D.R. Climate change can reduce the risk of biological invasion by reducing propagule pressure. Biol. Invasions 2016. [Google Scholar] [CrossRef]
- Van Asch, M.; Visser, M.E. Phenology of forest caterpillars and their host trees: The importance of synchrony. Ann. Rev. Entomol. 2007, 52, 37–55. [Google Scholar] [CrossRef] [PubMed]
- Salis, L.; Lof, M.; van Asch, M.; Visser, M.E. Modeling winter moth Operophtera brumata egg phenology: Nonlinear effects of temperature and developmental stage on developmental rate. Oikos 2016, 125, 1772–1781. [Google Scholar] [CrossRef]
- Embree, D.G. The diurnal and seasonal pattern of hatching of winter moth eggs, Operophtera brumata (Geometridae: Lepidoptera). Can. Entomol. 1970, 102, 759–768. [Google Scholar] [CrossRef]
- Kimberling, D.N.; Miller, J.C. Effects of temperature on larval eclosion of the winter moth, Operophtera brumata. Entomol. Exp. Appl. 1988, 47, 249–254. [Google Scholar] [CrossRef]
- Visser, M.E.; Holleman, L.J.M. Warmer springs disrupt the synchrony of oak and winter moth phenology. Proc. R. Soc. B Biol. Sci. 2001, 268, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Hibbard, E.L.; Elkinton, J.S. Effect of spring and winter temperatures on winter moth (Geometridae: Lepidoptera) larval eclosion in the Northeastern United States. Environ. Entomol. 2015, 44, 798–807. [Google Scholar] [CrossRef] [PubMed]
- Régnière, J.; Logan, J.A. Animal life cycle models. In Phenology: An Integrative Environmental Science; Schwartz, M.D., Ed.; Kluwer Academic Publishers: Boston, MA, USA, 2003; pp. 238–254. [Google Scholar]
- Schwartz, M.D. Introduction. In Phenology: An Integrative Environmental Science; Schwartz, M.D., Ed.; Kluwer Academic Publishers: Boston, MA, USA, 2003; pp. 1–5. [Google Scholar]
- Chuine, I.; Régnière, J. Process-Based Models of Phenology for Plants and Animals. Ann. Rev. Ecol. Evol. Syst. 2017, 48, 159–182. [Google Scholar] [CrossRef]
- Egerton, F.N. A history of the ecological sciences, part 21: Réaumur and his history of insects. Bull. Ecol. Soc. Am. 2006, 87, 212–224. [Google Scholar] [CrossRef]
- Janisch, E. The influence of temperature on the life-history of insects. Ecol. Entomol. 1932, 80, 137–168. [Google Scholar] [CrossRef]
- Sharpe, P.J.H.; Demichele, D.W. Reaction kinetics of poililotherm development. J. Theor. Biol. 1977, 64, 649–670. [Google Scholar] [CrossRef]
- Schoolfield, R.M.; Sharpe, P.J.H.; Magnuson, C.E. Non-linear regression of biological temperature-dependent rate models based on absolute reaction-rate theory. J. Theor. Biol. 1981, 88, 719–731. [Google Scholar] [CrossRef]
- Sanderson, E.D. The relation of temperature to the growth of insects. J. Econ. Entomol. 1910, 3, 113–189. [Google Scholar] [CrossRef]
- Sanderson, E.D.; Peairs, L.M. The relation of temperature to insect life. In Technical Bulletin; New Hampshire College and Agriculture Experiment Station: Durham, NH, USA, 1913. [Google Scholar]
- Logan, J.A.; Casagrande, P.A.; Liebhold, A.M. Modeling environment for simulation of gypsy moth (Lepidoptera: Lymantriidae) larval phenology. Environ. Entomol. 1991, 20, 1516–1525. [Google Scholar] [CrossRef]
- Sawyer, A.J.; Tauber, M.J.; Tauber, C.A.; Ruberson, J.R. Gypsy moth (Lepidoptera: Lymantriidae) egg development: A simulation analysis of laboratory and field data. Ecol. Model. 1993, 66, 121–155. [Google Scholar] [CrossRef]
- Gray, D.R.; Logan, J.A.; Ravlin, F.W.; Carlson, J.A. Toward a model of gypsy moth egg phenology: Using respiration rates of individual eggs to determine temperature-time requirements of prediapause development. Environ. Entomol. 1991, 20, 1645–1652. [Google Scholar] [CrossRef]
- Gray, D.R.; Ravlin, F.M.; Régnière, J.; Logan, J.A. Further advances toward a model of gypsy moth (Lymantria dispar (L.)) egg phenology: Respiration rates and thermal responsiveness during diapause, and age-dependent developmental rates in postdiapause. J. Insect Physiol. 1995, 41, 247–256. [Google Scholar] [CrossRef]
- Gray, D.R. Age-dependent postdiapause development in the Gypsy Moth (Lepidoptera: Lymantriidae) Life Stage Model. Environ. Entomol. 2009, 38, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Gray, D.R.; Ravlin, F.W.; Braine, J.A. Diapause in the gypsy moth: A model of inhibition and development. J. Insect Physiol. 2001, 47, 173–184. [Google Scholar] [CrossRef]
- Régnière, J. Temperature-dependent development of eggs and larvae of Choristoneura fumiferana (Clem.) (Lepidoptera: Tortricidae) and simulation of its seasonal history. Can. Entomol. 1987, 119, 717–728. [Google Scholar] [CrossRef]
- Wagner, T.L.; Wu, H.-I.; Sharpe, P.J.H.; Schoolfield, R.M.; Coulson, R.N. Modeling insect development rates: A literature review and application of a biophysical model. Ann. Entomol. Soc. Am. 1984, 77, 208–225. [Google Scholar] [CrossRef]
- Andrewartha, H.G.; Birch, L.C. The Distribution and Abundance of Animals; The University of Chicago Press: Chicago, IL, USA, 1954; 782p. [Google Scholar]
- Bale, J.S.; Masters, G.J.; Hodkinson, I.D.; Awmack, C.; Bezemer, T.M.; Brown, V.K.; Butterfield, J.; Buse, A.; Coulson, J.C.; Farrar, J.; et al. Herbivory in global climate change research: Direct effects of rising temperature on insect herbivores. Glob. Chang. Biol. 2002, 8, 1–16. [Google Scholar] [CrossRef]
- Parmesan, C.; Ryrholm, N.; Stefanescu, C.; Hill, J.K.; Thomas, C.D.; Descimon, H.; Huntley, B.; Kaila, L.; Kullberg, J.; Tammaru, T.; et al. Poleward shifts in geographical ranges of butterfly species associated with regional warming. Nature 1999, 399, 579–583. [Google Scholar] [CrossRef]
- Battisti, A.; Stastny, M.; Buffo, E.; Larsson, S. A rapid altitudinal range expansion in the pine processionary moth produced by the 2003 climatic anomaly. Glob. Chang. Biol. 2006, 12, 662–671. [Google Scholar] [CrossRef]
- Crozier, L. Warmer winters drive butterfly range expansion by increasing survivorship. Ecology 2004, 85, 231–241. [Google Scholar] [CrossRef]
- Jepsen, J.U.; Hagen, S.B.; Yoccoz, N.G. Climate change and outbreaks of the geometrids Operophtera brumata and Epirrita autumnata in subarctic birch forest: Evidence of a recent outbreak range expansion [electronic resource]. J. Anim. Ecol. 2008, 77, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.J.; Hagen, S.B.; Ims, R.A.; Yoccoz, N.G. Changes to the elevational limits and extent of species ranges associated with climate change. Ecol. Lett. 2005, 8, 1138–1146. [Google Scholar] [CrossRef] [PubMed]
- Parmesan, C. Ecological and evolutionary responses to recent climate change. Ann. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef]
- Araújo, M.B.; Guisan, A. Five (or so) challenges for species distribution modelling. J. Biogeogr. 2006, 33, 1677–1688. [Google Scholar] [CrossRef]
- Davis, A.J.; Jenkinson, L.S.; Lawton, J.H.; Shorrocks, B.; Wood, S. Making mistakes when predicting shifts in species range in response to global warming. Nature 1998, 391, 783–786. [Google Scholar] [CrossRef] [PubMed]
- Barry, S.; Elith, J. Error and uncertainty in habitat models. J. Appl. Ecol. 2006, 43, 413–423. [Google Scholar] [CrossRef]
- Buisson, L.; Thuiller, W.; Casajus, N.; Lek, S.; Grenouillet, G. Uncertainty in ensemble forecasting of species distribution. Glob. Chang. Biol. 2010, 16, 1145–1157. [Google Scholar] [CrossRef]
- Heikkinen, R.K.; Louto, M.; Araújo, M.B.; Virkkala, R.; Thuiller, W.; Sykes, T. Methods and uncertainties in bioclimatic envelope modelling under climate change. Prog. Phys. Geogr. 2006, 30, 751–777. [Google Scholar] [CrossRef]
- Thuiller, W.; Brotons, L.; Araújo, M.B.; Lavorel, S. Effects of restricting environmental range of data to project current and future species distributions. Ecography 2004, 27, 165–172. [Google Scholar] [CrossRef]
- Chuine, I. Why does phenology drive species distribution? Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3149–3160. [Google Scholar] [CrossRef] [PubMed]
- Tobin, P.C.; Gray, D.R.; Liebhold, A.M. Supraoptimal temperatures influence the range dynamics of a non-native insect. Divers. Distrib. 2014, 20, 813–823. [Google Scholar] [CrossRef]
- Gray, D.R. Using geographically robust models of insect phenology in forestry. In Phenology and Climate Change; Zhang, X., Ed.; InTech: Rijeka, Croatia, 2012; Chapter 1; pp. 3–20. [Google Scholar]
- Gray, D.R. Unwanted spatial bias in predicting establishment of an invasive insect based on simulated demographics. Int. J. Biometeorol. 2014, 58, 949–961. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.C.; Mason, D.P.; Radke, S.L.; Tracewski, K.T. Gypsy moth, Lymantria dispar (L.) (Lepidoptera: Lymantriidae), egg eclosion: Degree-day accumulation. Environ. Entomol. 1983, 12, 929–932. [Google Scholar] [CrossRef]
- Lyons, D.B.; Lysyk, T.J. Development and phenology of eggs of gypsy moth Lymantria dispar (Lepidoptera: Lymantriidae) in Ontario. In Proceedings, Lymantriidae: A Comparison of Features of New and Old World Tussock Moths; Wallner, W.E., McManus, K.A., Eds.; USDA General Technical Report NE-123:351–365; United States Department of Agriculture, Forest Service: Radnor, PA, USA, 1989. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exposure # | Exposure Period (d) 1 | Predicted Hatch Day 2 | Observed Hatch Day 3 |
|---|---|---|---|
| 6 | 42–70 | 69 | na |
| 7 | 49–77 | 71 | na |
| 8 | 56–84 | 74 | 80 |
| 9 | 63–91 | 77 | 80 |
| 10 | 70–98 | 80 | 85 |
| Exposure # | Exposure Period (d) | Development at 5 °C | Development at 15 °C |
|---|---|---|---|
| 1 | 7–35 | 0.21 | 0.42 |
| 2 | 14–42 | 0.21 | 0.43 |
| 3 | 21–49 | 0.22 | 0.45 |
| 4 | 28–56 | 0.21 | 0.44 |
| 5 | 35–63 | 0.22 | 0.44 |
| 6 | 42–70 | 0.21 | 0.44 |
| 7 | 49–77 | 0.19 | 0.47 |
| 8 | 56–84 1 | 0.18 | 0.46 |
| 9 | 63–91 1 | 0.17 | 0.40 |
| 10 | 70–98 1 | 0.16 | 0.33 |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gray, D.R. Age-Dependent Developmental Response to Temperature: An Examination of the Rarely Tested Phenomenon in Two Species (Gypsy Moth (Lymantria dispar) and Winter Moth (Operophtera brumata)). Insects 2018, 9, 41. https://doi.org/10.3390/insects9020041
Gray DR. Age-Dependent Developmental Response to Temperature: An Examination of the Rarely Tested Phenomenon in Two Species (Gypsy Moth (Lymantria dispar) and Winter Moth (Operophtera brumata)). Insects. 2018; 9(2):41. https://doi.org/10.3390/insects9020041
Chicago/Turabian StyleGray, David R. 2018. "Age-Dependent Developmental Response to Temperature: An Examination of the Rarely Tested Phenomenon in Two Species (Gypsy Moth (Lymantria dispar) and Winter Moth (Operophtera brumata))" Insects 9, no. 2: 41. https://doi.org/10.3390/insects9020041
APA StyleGray, D. R. (2018). Age-Dependent Developmental Response to Temperature: An Examination of the Rarely Tested Phenomenon in Two Species (Gypsy Moth (Lymantria dispar) and Winter Moth (Operophtera brumata)). Insects, 9(2), 41. https://doi.org/10.3390/insects9020041