
Propolis Counteracts Some Threats to Honey Bee Health



Abstract
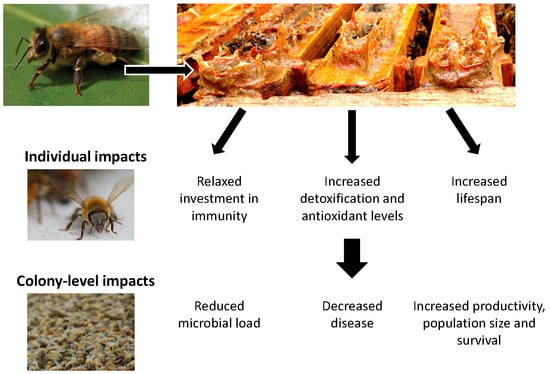
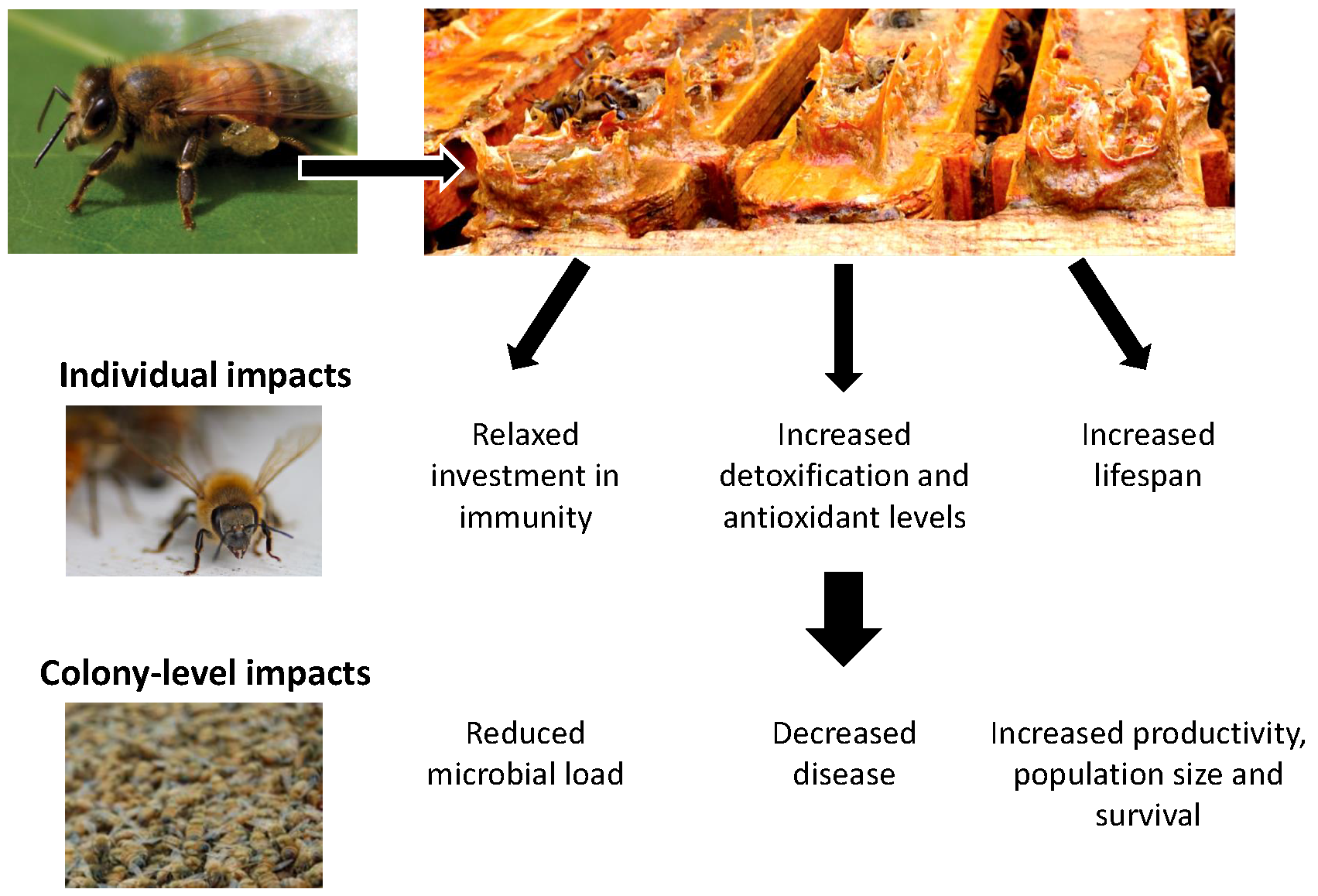
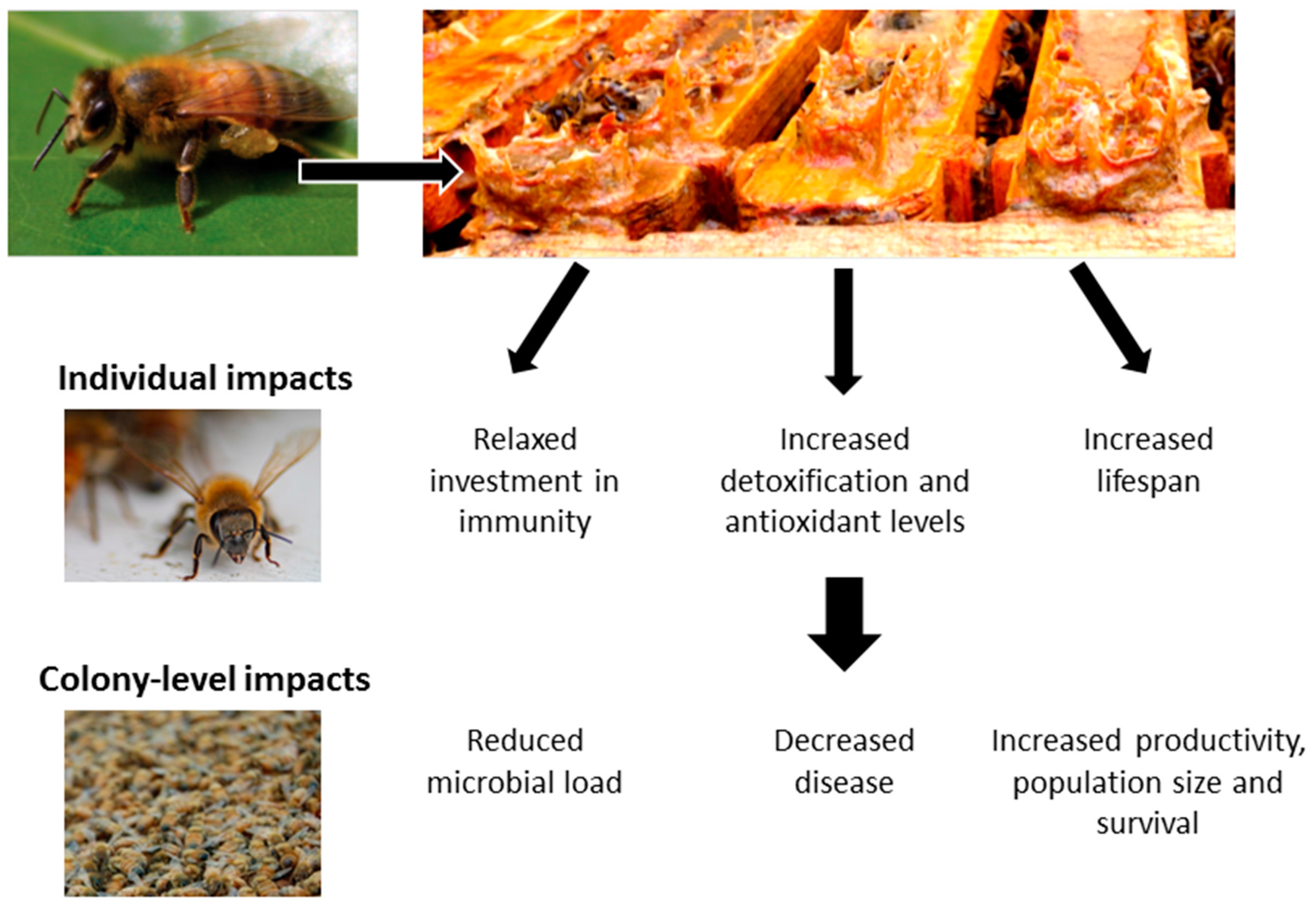
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
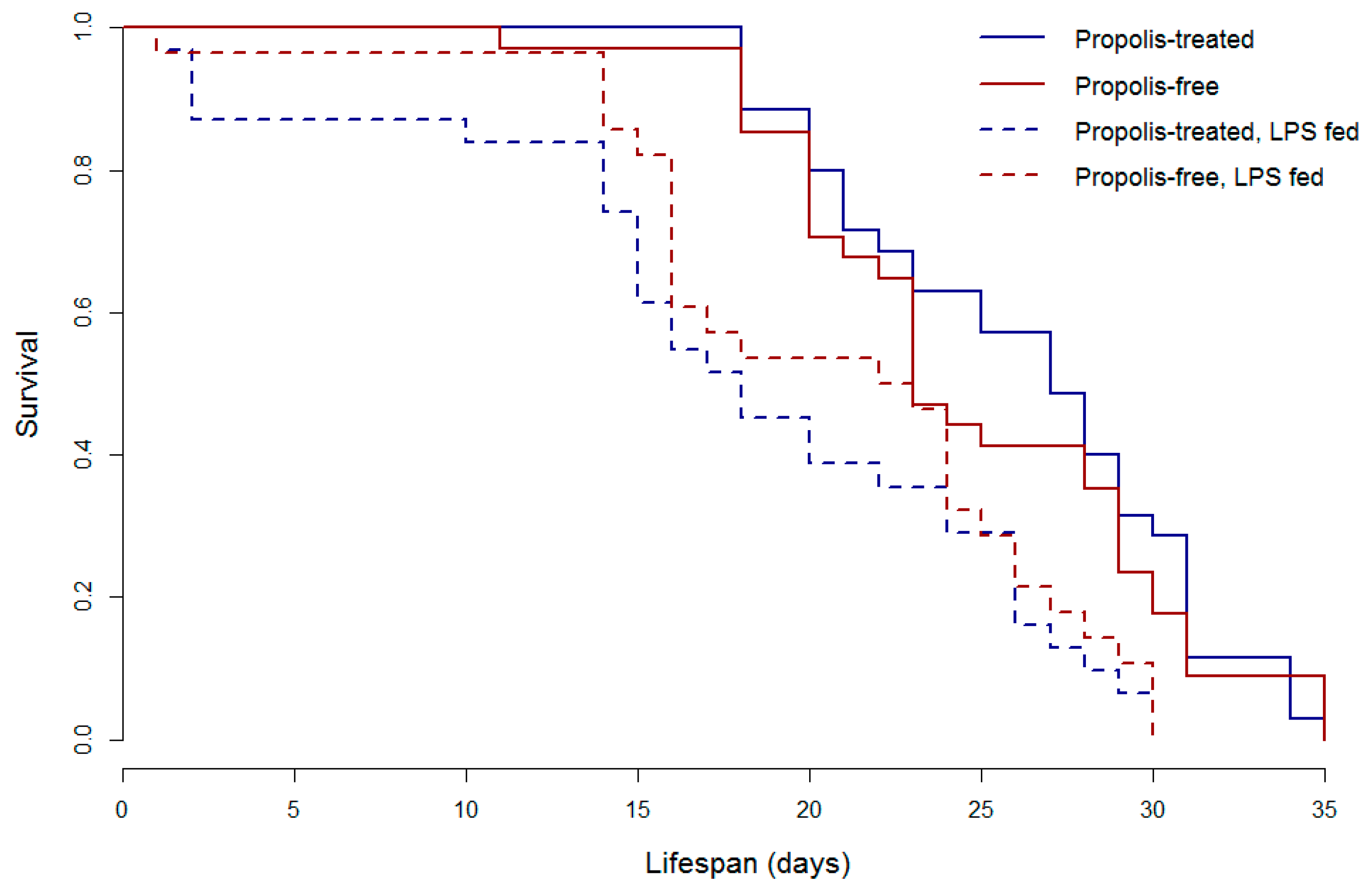{kind=link}
{kind=link}
1. Introduction
1.1. The Role of Propolis in the Hive
1.2. Propolis as Social Immunity
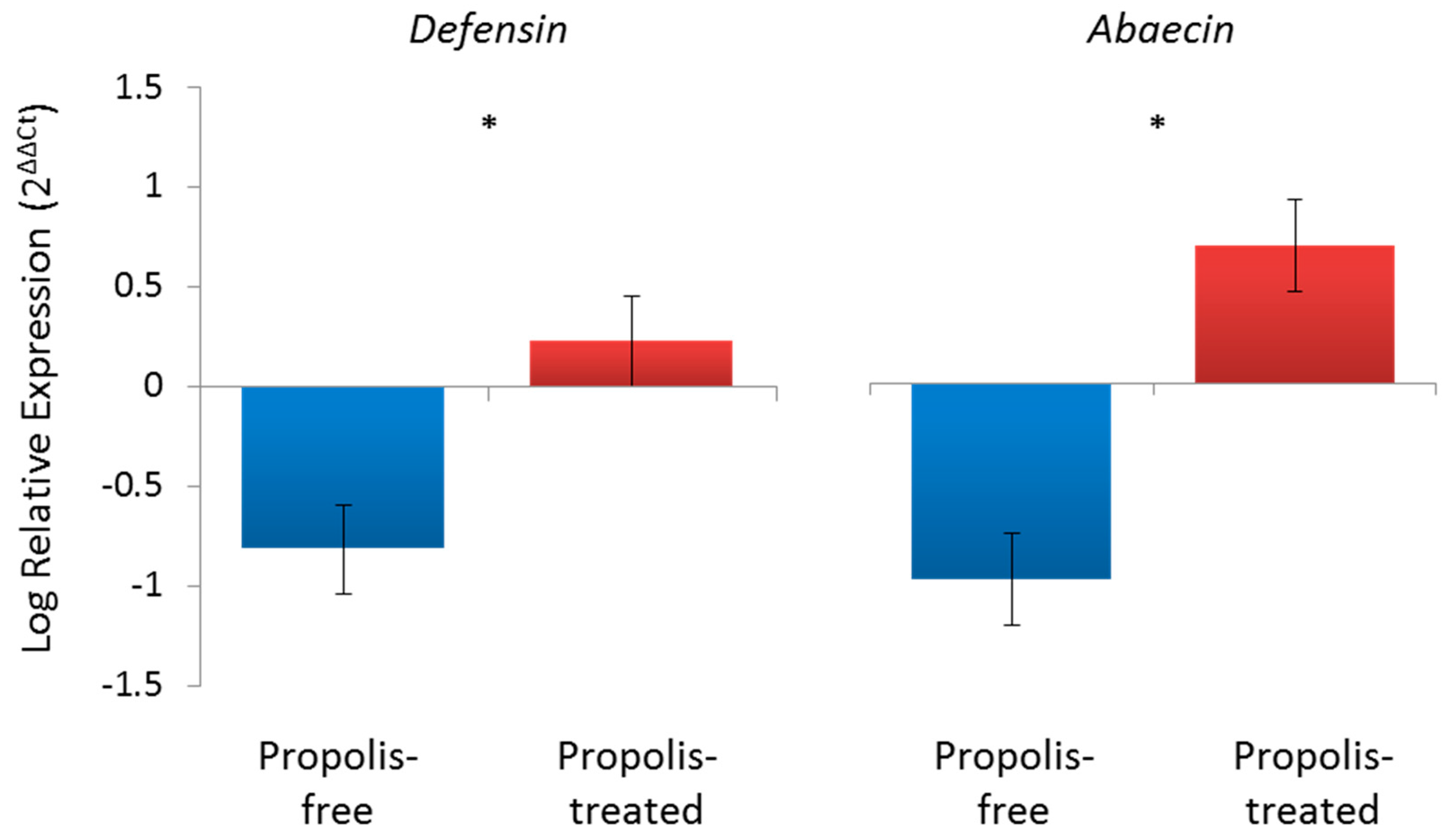
2. Constitutive Effects of Propolis on the Honey Bee Immune System
3. Does Propolis Suppress Immune Function?
4. The Effect of Propolis against Pathogens
5. Effect of Propolis on Other Bee Pathogens, Parasites and Pests
5.1 Nosema Ceranae
5.2 Varroa Destructor
5.3 Viruses
5.4 Other Hive Pests
6. Propolis and Pesticides
7. Propolis and Other Aspects of Bee Health
8. Applications for the Beekeeping Community
9. Conclusions
Acknowledgments
Conflicts of Interest
References
- Evans, J.D.; Schwarz, R.S. Bees brought to their knees: Microbes affecting honey bee health. Trends Microbiol. 2011, 19, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Bayo, F.; Goulson, D.; Pennacchio, F.; Nazzi, F.; Goka, K.; Desneux, N. Are bee diseases linked to pesticides?—A brief review. Environ. Int. 2016, 89–90, 7–11. [Google Scholar] [CrossRef] [PubMed]
- vanEngelsdorp, D.; Meixner, M.D. A historical review of managed honey bee populations in europe and the united states and the factors that may affect them. J. Invertebr. Pathol. 2010, 103, S80–S95. [Google Scholar] [CrossRef] [PubMed]
- Seitz, N.; Traynor, K.S.; Steinhauer, N.; Rennich, K.; Wilson, M.E.; Ellis, J.D.; Rose, R.; Tarpy, D.R.; Sagili, R.R.; Caron, D.M.; et al. A national survey of managed honey bee 2014–2015 annual colony losses in the USA. J. Apic. Res. 2015, 54, 292–304. [Google Scholar] [CrossRef]
- Simone-Finstrom, M.; Spivak, M. Propolis and bee health: The natural history and significance of resin use by honey bees. Apidologie 2010, 41, 295–311. [Google Scholar] [CrossRef]
- Meyer, W. The propolis bees and their activities. Z. Bienenforsch. 1954, 2, 185–200. [Google Scholar] [CrossRef]
- Zhang, C.-P.; Zheng, H.-Q.; Hu, F.-L. Extraction, partial characterization, and storage stability of β-glucosidase from propolis. J. Food Sci. 2011, 76, C75–C79. [Google Scholar] [CrossRef] [PubMed]
- Castella, G.; Chapuisat, M.; Christe, P. Prophylaxis with resin in wood ants. Anim. Behav. 2008, 75, 1591–1596. [Google Scholar] [CrossRef]
- Leonhardt, S.D.; Bluthgen, N. A sticky affair: Resin collection by bornean stingless bees. Biotropica 2009, 41, 730–736. [Google Scholar] [CrossRef]
- Seeley, T.D.; Morse, R.A. Nest of the honey bee (Apis mellifera L.). Insectes Soc. 1976, 23, 495–512. [Google Scholar] [CrossRef]
- Seeley, T.D.; Seeley, R.H.; Akratanakul, P. Colony defense strategies of the honeybees in thailand. Ecol. Monogr. 1982, 52, 43–63. [Google Scholar] [CrossRef]
- Visscher, P.K. Adaptations of honey bees (Apis mellifera) to problems of nest hygiene. Sociobiology 1980, 5, 249–260. [Google Scholar]
- Clark, E.H. Constructive Beekeeping; Ulsaker Printing Company: Fargo, ND, USA, 1918; p. 52. [Google Scholar]
- Simone, M.; Evans, J.D.; Spivak, M. Resin collection and social immunity in honey bees. Evolution 2009, 63, 3016–3022. [Google Scholar] [CrossRef] [PubMed]
- Bastos, E.M.A.F.; Simone, M.; Macedo Jorge, D.; Egea Soares, A.E.; Spivak, M. In vitro study of the antimicrobial activity of brazilian propolis against Paenibacillus larvae. J. Invertebr. Pathol. 2008, 97, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Simone-Finstrom, M.D.; Spivak, M. Increased resin collection after parasite challenge: A case of self-medication in honey bees? PLoS ONE 2012, 7, e34061. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.B.; Spivak, M.; Hegeman, A.D.; Rendahl, A.; Cohen, J.D. Metabolomics reveals the origins of antimicrobial plant resins collected by honey bees. PLoS ONE 2013, 8, e77512. [Google Scholar] [CrossRef] [PubMed]
- Nicodemo, D.; Malheiros, E.B.; De Jong, D.; Couto, R.H.N. Increased brood viability and longer lifespan of honeybees selected for propolis production. Apidologie 2014, 45, 269–275. [Google Scholar] [CrossRef]
- Borba, R.S. Constitutive and Therapeutic Benefits of Plant Resins and a Propolis Envelope to Honey Bee, Apis mellifera L., Immunity and Health. Ph.D. Thesis, University of Minnesota, Ann Arbor, MA, USA, 2015. [Google Scholar]
- Borba, R.S.; Klyczek, K.K.; Mogen, K.L.; Spivak, M. Seasonal benefits of a natural propolis envelope to honey bee immunity and colony health. J. Exp. Biol. 2015, 218, 3689–3699. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.B.; Brinkman, D.; Spivak, M.; Gardner, G.; Cohen, J.D. Regional variation in composition and antimicrobial activity of us propolis against Paenibacillus larvae and Ascosphaera apis. J. Invertebr. Pathol. 2015, 124, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Drescher, N.; Klein, A.-M.; Neumann, P.; Yañez, O.; Leonhardt, S. Inside honeybee hives: Impact of natural propolis on the ectoparasitic mite Varroa destructor and viruses. Insects 2017, 8, 15. [Google Scholar] [CrossRef] [PubMed]
- Cremer, S.; Armitage, S.A.O.; Schmid-Hempel, P. Social immunity. Curr. Biol. 2007, 17, R693–R702. [Google Scholar] [CrossRef] [PubMed]
- Meunier, J. Social immunity and the evolution of group living in insects. Philos. Trans. R. Soc. B-Biol. Sci. 2015, 370, 10. [Google Scholar] [CrossRef] [PubMed]
- Schmid-Hempel, P. Parasites in Social Insects; Princeton University Press: Princeton, NJ, USA, 1998. [Google Scholar]
- Evans, J.D.; Lopez, D.L. Bacterial probiotics induce an immune response in the honey bee (hymenoptera: Apidae). J. Econ. Entomol. 2004, 97, 752–756. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.E.; Sheehan, T.H.; Eckholm, B.J.; Mott, B.M.; DeGrandi-Hoffman, G. An emerging paradigm of colony health: Microbial balance of the honey bee and hive (Apis mellifera). Insectes Soc. 2011, 58, 431–444. [Google Scholar] [CrossRef]
- Evans, J.D.; Aronstein, K.; Chen, Y.P.; Hetru, C.; Imler, J.L.; Jiang, H.; Kanost, M.; Thompson, G.J.; Zou, Z.; Hultmark, D. Immune pathways and defence mechanisms in honey bees Apis mellifera. Insect Mol. Biol. 2006, 15, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Niu, G.; Johnson, R.M.; Berenbaum, M.R. Toxicity of mycotoxins to honeybees and its amelioration by propolis. Apidologie 2011, 42, 79–87. [Google Scholar] [CrossRef]
- Schmid-Hempel, P. Evolutionary ecology of insect immune defenses. Annu. Rev. Entomol. 2005, 50, 529–551. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.D.; Pettis, J.S. Colony-level impacts of immune responsiveness in honey bees, Apis mellifera. Evolution 2005, 59, 2270–2274. [Google Scholar] [CrossRef] [PubMed]
- Moret, Y.; Schmid-Hempel, P. Survival for immunity: The price of immune system activation for bumblebee workers. Science 2000, 290, 1166–1168. [Google Scholar] [CrossRef] [PubMed]
- Wilson-Rich, N.; Spivak, M.; Fefferman, N.H.; Starks, P.T. Genetic, individual, and group facilitation of disease resistance in insect societies. Annu. Rev. Entomol. 2009, 54, 405–423. [Google Scholar] [CrossRef] [PubMed]
- Simone-Finstrom, M. Social immunity and the superorganism: Behavioral defenses protecting honey bee colonies from pathogens and parasites. Bee World 2017, 94, 21–29. [Google Scholar] [CrossRef]
- Erler, S.; Moritz, R.F. Pharmacophagy and pharmacophory: Mechanisms of self-medication and disease prevention in the honeybee colony (Apis mellifera). Apidologie 2016, 47, 389–411. [Google Scholar] [CrossRef]
- Christe, P.; Oppliger, A.; Bancala, F.; Castella, G.; Chapuisat, M. Evidence for collective medication in ants. Ecol. Lett. 2003, 6, 19–22. [Google Scholar] [CrossRef]
- Castella, G.; Chapuisat, M.; Moret, Y.; Christe, P. The presence of conifer resin decreases the use of the immune system in wood ants. Ecol. Entomol. 2008, 33, 408–412. [Google Scholar] [CrossRef]
- Chapuisat, M.; Oppliger, A.; Magliano, P.; Christe, P. Wood ants use resin to protect themselves against pathogens. Proc. R. Soc. B-Biol. Sci. 2007, 274, 2013–2017. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.K.; Paredes-Guzman, J.F.; Aguiar, C.L.; Alencar, S.M.; Fujiwara, F.Y. Chemical constituents in Baccharis dracunculifolia as the main botanical origin of southeastern brazilian propolis. J. Agric. Food Chem. 2004, 52, 1100–1103. [Google Scholar] [CrossRef] [PubMed]
- Bankova, V.; Boudourova-Krasteva, G.; Popov, S.; Sforcin, J.M.; Funari, S.R.C. Seasonal variations of the chemical composition of brazilian propolis. Apidologie 1998, 29, 361–367. [Google Scholar] [CrossRef]
- Sforcin, J.; Fernandes, A.; Lopes, C.; Bankova, V.; Funari, S. Seasonal effect on brazilian propolis antibacterial activity. J. Ethnopharmacol. 2000, 73, 243–249. [Google Scholar] [CrossRef]
- Engels, W.; Kaatz, H.; Zillikens, A.; Simões, Z.P.; Trube, A.; Braun, R.; Dittrich, F. Honey bee reproduction: Vitellogenin and caste-specific regulation of fertility. In Honey Bee Reproduction: Vitellogenin and Caste-Specific Regulation of Fertility; Elsevier Science Publishers B.V.: Amsterdam, The Netherlands, 1990; pp. 495–502. [Google Scholar]
- Amdam, G.V.; Norberg, K.; Hagen, A.; Omholt, S.W. Social exploitation of vitellogenin. Proc. Natl. Acad. Sci. USA 2003, 100, 1799–1802. [Google Scholar] [CrossRef] [PubMed]
- Amdam, G.V.; Simões, Z.L.; Hagen, A.; Norberg, K.; Schrøder, K.; Mikkelsen, Ø.; Kirkwood, T.B.; Omholt, S.W. Hormonal control of the yolk precursor vitellogenin regulates immune function and longevity in honeybees. Exp. Gerontol. 2004, 39, 767–773. [Google Scholar] [CrossRef] [PubMed]
- Smart, M.; Pettis, J.; Rice, N.; Browning, Z.; Spivak, M. Linking measures of colony and individual honey bee health to survival among apiaries exposed to varying agricultural land use. PLoS ONE 2016, 11, e0152685. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.R.; Oliveira, N.T.E.; Camargo, S.C.; Pires, B.G.; Oliveira, C.A.L.; Teixeira, R.A.; Pickler, M.A. Honey and propolis production, hygiene and defense behaviors of two generations of africanized honey bees. Sci. Agric. 2013, 70, 74–81. [Google Scholar] [CrossRef]
- Manrique, A.J.; Soares, A.E.E. Start of africanized honey bee selection program for increased propolis production and its effect on honey production. Interciencia 2002, 27, 312–316. [Google Scholar]
- Nicodemo, D.; De Jong, D.; Couto, R.H.N.; Malheiros, E.B. Honey bee lines selected for high propolis production also have superior hygienic behavior and increased honey and pollen stores. Genet. Mol. Res. 2013, 12, 6931–6938. [Google Scholar] [CrossRef] [PubMed]
- Padilha, A.H.; Sattler, A.; Cobuci, J.A.; McManus, C.M. Genetic parameters for five traits in africanized honeybees using bayesian inference. Genet. Mol. Biol. 2013, 36, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Simone-Finstrom, M.; Hayworth, M. USDA-ARS Honey Bee Breeding, Genetics, and Physiology Laboratory, Baton Rouge, LA 70820, USA. Unpublished data. 2012. [Google Scholar]
- Mallon, E.B.; Brockmann, A.; Schmid-Hempel, P. Immune response inhibits associative learning in insects. Proc. R. Soc. B-Biol. Sci. 2003, 270, 2471–2473. [Google Scholar] [CrossRef] [PubMed]
- Richard, F.J.; Aubert, A.; Grozinger, C.M. Modulation of social interactions by immune stimulation in honey bee, Apis mellifera, workers. BMC Biol. 2008, 6. [Google Scholar] [CrossRef] [PubMed]
- Liao, L.-H.; Wu, W.-Y.; Berenbaum, M. Impacts of dietary phytochemicals in the presence and absence of pesticides on longevity of honey bees (Apis mellifera). Insects 2017, 8, 22. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time pcr data by the comparative ct method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.M.; Mao, W.; Pollock, H.S.; Niu, G.; Schuler, M.A.; Berenbaum, M.R. Ecologically appropriate xenobiotics induce cytochrome p450s in Apis mellifera. PLoS ONE 2012, 7, e31051. [Google Scholar] [CrossRef] [PubMed]
- Mao, W.; Schuler, M.A.; Berenbaum, M.R. Honey constituents up-regulate detoxification and immunity genes in the western honey bee Apis mellifera. Proc. Natl. Acad. Sci. USA 2013, 110, 8842–8846. [Google Scholar] [CrossRef] [PubMed]
- Westra, E.R.; van Houte, S.; Oyesiku-Blakemore, S.; Makin, B.; Broniewski, J.M.; Best, A.; Bondy-Denomy, J.; Davidson, A.; Boots, M.; Buckling, A. Parasite exposure drives selective evolution of constitutive versus inducible defense. Curr. Biol. 2015, 25, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Burdock, G.A. Review of the biological properties and toxicity of bee propolis. Food Chem. Toxicol. 1998, 36, 347–363. [Google Scholar] [CrossRef]
- Bankova, V.; Marcucci, M.C. Standardization of propolis: Present status and perspectives. Bee World 2000, 81, 182–188. [Google Scholar] [CrossRef]
- Bankova, V.S.; de Castro, S.L.; Marcucci, M.C. Propolis: Recent advances in chemistry and plant origin. Apidologie 2000, 31, 3–15. [Google Scholar] [CrossRef]
- Sforcin, J.M.; Bankova, V. Propolis: Is there a potential for the development of new drugs? J. Ethnopharmacol. 2011, 133, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Silva-Carvalho, R.; Baltazar, F.; Almeida-Aguiar, C. Propolis: A complex natural product with a plethora of biological activities that can be explored for drug development. Evid.-Based Complement. Alternat. Med. 2015, 2015, 206439. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Zhang, C.-P.; Wang, K.; Li, G.Q.; Hu, F.-L. Recent advances in the chemical composition of propolis. Molecules 2014, 19, 19610–19632. [Google Scholar] [CrossRef] [PubMed]
- Langenheim, J.H. Plant Resins: Chemistry, Evolution, Ecology and Ethnobiology; Timber Press: Portland, OR, USA, 2003; p. 586. [Google Scholar]
- Alday, E.; Valencia, D.; Carreño, A.L.; Picerno, P.; Piccinelli, A.L.; Rastrelli, L.; Robles-Zepeda, R.; Hernandez, J.; Velazquez, C. Apoptotic induction by pinobanksin and some of its ester derivatives from sonoran propolis in a b-cell lymphoma cell line. Chem.-Biol. Interact. 2015, 242, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Cuesta-Rubio, O.; Frontana-Uribe, B.A.; Ramírez-Apan, T.; Cárdenas, J. Polyisoprenylated benzophenones in cuban propolis: Biological activity of nemorosone. Z. Naturforsch. C 2002, 57, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Aga, H.; Shibuya, T.; Sugimoto, T.; Kurimoto, M.; Nakajima, S. Isolation and identification of antimicrobial compounds in brazilian propolis. Biosci. Biotechnol. Biochem. 1994, 58, 945–946. [Google Scholar] [CrossRef]
- Popova, M.P.; Chinou, I.B.; Marekov, I.N.; Bankova, V.S. Terpenes with antimicrobial activity from cretan propolis. Phytochemistry 2009, 70, 1262–1271. [Google Scholar] [CrossRef] [PubMed]
- Salatino, A.; Weinstein Teixeira, W.; Negri, G.; Message, D. Origin and chemical variation of brazilian propolis. Evid. Based Complement. Alternat. Med. 2005, 2, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Siheri, W.; Igoli, J.O.; Gray, A.I.; Nasciemento, T.G.; Zhang, T.; Fearnley, J.; Clements, C.J.; Carter, K.C.; Carruthers, J.; Edrada-Ebel, R. The isolation of antiprotozoal compounds from libyan propolis. Phytother. Res. 2014, 28, 1756–1760. [Google Scholar] [CrossRef] [PubMed]
- Bilikova, K.; Popova, M.; Trusheva, B.; Bankova, V. New anti-paenibacillus larvae substances purified from propolis. Apidologie 2013, 44, 278–285. [Google Scholar] [CrossRef]
- Wilson, M.B.; Pawlus, A.D.; Brinkman, D.; Gardner, G.; Hegeman, A.D.; Spivak, M.; Cohen, J.D. 3-acyl dihydroflavonols from poplar resins collected by honey bees are active against the bee pathogens Paenibacillus larvae and Ascosphaera apis. Phytochemistry 2017, 138, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Voigt, K.; Rademacher, E. Effect of the propolis components, cinnamic acid and pinocembrin, on Apis mellifera and Ascosphaera apis. J. Apic. Sci. 2015, 59, 89–95. [Google Scholar] [CrossRef]
- Ghisalberti, E.L. Propolis: A review. Bee World 1979, 60, 263–282. [Google Scholar] [CrossRef]
- Marcucci, M.C. Propolis: Chemical composition, biological properties and therapeutic activity. Apidologie 1995, 26, 83–99. [Google Scholar] [CrossRef]
- Gekker, G.; Hu, S.X.; Spivak, M.; Lokensgard, J.R.; Peterson, P.K. Anti-HIV-1 activity of propolis in CD4(+) lymphocyte and microglial cell cultures. J. Ethnopharmacol. 2005, 102, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Sforcin, J.M. Biological properties and therapeutic applications of propolis. Phytother. Res. 2016, 30, 894–905. [Google Scholar] [CrossRef] [PubMed]
- Drescher, N.; Wallace, H.M.; Katouli, M.; Massaro, C.F.; Leonhardt, S.D. Diversity matters: How bees benefit from different resin sources. Oecologia 2014, 176, 943–953. [Google Scholar] [CrossRef] [PubMed]
- Lindenfelser, L.A. Antimicrobial activity of propolis. Am. Bee J. 1967, 107, 90–92. [Google Scholar]
- Lindenfelser, L.A. In vivo activity of propolis against Bacillus larvae. J. Invertebr. Pathol. 1968, 12, 129–131. [Google Scholar] [CrossRef]
- Antunez, K.; Harriet, J.; Gende, L.; Maggi, M.; Eguaras, M.; Zunino, P. Efficacy of natural propolis extract in the control of american foulbrood. Vet. Microbiol. 2008, 131, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Fadhil, N.H.; Powell, J.E.; Kwong, W.K.; Moran, N.A. Long-term exposure to antibiotics has caused accumulation of resistance determinants in the gut microbiota of honeybees. MBio 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Messer, A.C. Fresh dipterocarp resins gathered by megachilid bees inhibit growth of pollen-associated fungi. Biotropica 1985, 17, 175–176. [Google Scholar] [CrossRef]
- Garedew, A.; Lamprecht, I.; Schmolz, E.; Schricker, B. The varroacidal action of propolis: A laboratory assay. Apidologie 2002, 33, 41–50. [Google Scholar] [CrossRef]
- Morse, R.; Flottum, K. Honey Bee Pests Predators and Diseases; AI Root Company: Medina, OH, USA, 1997. [Google Scholar]
- Gilliam, M.; Taber, S.; Lorenz, B.J.; Prest, D.B. Factors affecting development of chalkbrood disease in colonies of honey bees, Apis mellifera, fed pollen contaminated with Ascosphaera apis. J. Invertebr. Pathol. 1988, 52, 314–325. [Google Scholar] [CrossRef]
- Spivak, M.; Reuter, G.S. Resistance to american foulbrood disease by honey bee colonies (Apis mellifera) bred for hygienic behavior. Apidologie 2001, 32, 555–565. [Google Scholar] [CrossRef]
- Rothenbuhler, W.C. Behaviour genetics of nest cleaning in honey bees. I. Responses of four inbred lines to disease-killed brood. Anim. Behav. 1964, 12, 578–583. [Google Scholar] [CrossRef]
- Rothenbuhler, W.C.; Thompson, V.C. Resistance to american foulbrood in honey bees. I. Differential survival of larvae of different genetic lines. J. Econ. Entomol. 1956, 49, 470–475. [Google Scholar] [CrossRef]
- Bambrick, J.F.; Rothenbuhler, W.C. Resistance to american foulbrood in honey bees. 4. Relationship between larval age at innoculation and mortality in a resistant and in a susceptible line. J. Invertebr. Pathol. 1961, 3, 381–390. [Google Scholar]
- Thompson, V.C.; Rothenbuhler, W.C. Resistance to american foulbrood in honey bees. Ii. Differential protection of larvae by adults of different genetic lines. J. Econ. Entomol. 1957, 50, 731–737. [Google Scholar] [CrossRef]
- Rose, R.I.; Briggs, J.D. Resistance to american foulbrood in honey bees ix. Effects of honey-bee larval food on the growth and viability of Bacillus larvae. J. Invertebr. Pathol. 1969, 13, 74–80. [Google Scholar] [CrossRef]
- Seeley, T.D.; Tarpy, D.R. Queen promiscuity lowers disease within honeybee colonies. Proc. R. Soc. B-Biol. Sci. 2007, 274, 67–72. [Google Scholar] [CrossRef] [PubMed]
- de Graaf, D.C.; Alippi, A.M.; Antunez, K.; Aronstein, K.A.; Budge, G.; De Koker, D.; De Smet, L.; Dingman, D.W.; Evans, J.D.; Foster, L.J.; et al. Standard methods for american foulbrood research. J. Apic. Res. 2013, 52, 1–28. [Google Scholar] [CrossRef]
- López-Uribe, M.M.; Fitzgerald, A.; Simone-Finstrom, M. Inducible versus constitutive social immunity: Examining effects of colony infection on glucose oxidase and Defensin-1 production in honey bees. R. Soc. Open Sci. 2017. accepted. [Google Scholar]
- Gherman, B.I.; Denner, A.; Bobis, O.; Dezmirean, D.S.; Marghitas, L.A.; Schluens, H.; Moritz, R.F.A.; Erler, S. Pathogen-associated self-medication behavior in the honeybee Apis mellifera. Behav. Ecol. Sociobiol. 2014, 68, 1777–1784. [Google Scholar] [CrossRef]
- Cox-Foster, D.L.; Conlan, S.; Holmes, E.C.; Palacios, G.; Evans, J.D.; Moran, N.A.; Quan, P.-L.; Briese, T.; Hornig, M.; Geiser, D.M.; et al. A metagenomic survey of microbes in honey bee colony collapse disorder. Science 2007, 318, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Evans, J.D.; Smith, I.B.; Pettis, J.S. Nosema ceranae is a long-present and wide-spread microsporidian infection of the european honey bee (apis mellifera) in the united states. J. Invertebr. Pathol. 2008, 97, 186–188. [Google Scholar] [CrossRef] [PubMed]
- Klee, J.; Besana, A.M.; Genersch, E.; Gisder, S.; Nanetti, A.; Tam, D.Q.; Chinh, T.X.; Puerta, F.; Ruz, J.M.; Kryger, P. Widespread dispersal of the microsporidian Nosema ceranae, an emergent pathogen of the western honey bee, Apis mellifera. J. Invertebr. Pathol. 2007, 96, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Yemor, T.; Phiancharoen, M.; Eric Benbow, M.; Suwannapong, G. Effects of stingless bee propolis on nosema ceranae infected asian honey bees, apis cerana. J. Apic. Res. 2015, 54, 468–473. [Google Scholar] [CrossRef]
- Suwannapong, G.; Maksong, S.; Benbow, M. Stingless bee propolis effects on experimental infection of apis florea with nosema ceranae. J. Agric. Sci. Technol. A 2011, 1, 818–825. [Google Scholar]
- Kwong, W.K.; Moran, N.A. Gut microbial communities of social bees. Nat. Rev. Microbiol. 2016, 14, 374–384. [Google Scholar] [CrossRef] [PubMed]
- Garedew, A.; Schmolz, E.; Lamprecht, I. Microcalorimetric and respirometric investigation of the effect of temperature on the antivarroa action of the natural bee product-propolis. Thermochim. Acta 2003, 399, 171–180. [Google Scholar] [CrossRef]
- Damiani, N.; Daniel Maggi, M.; Brenda Gende, L.; Faverin, C.; Javier Eguaras, M.; Augusto Marcangeli, J. Evaluation of the toxicity of a propolis extract on Varroa destructor (acari: Varroidae) and Apis mellifera (hymenoptera: Apidae). J. Apic. Res. 2010, 49, 257–264. [Google Scholar] [CrossRef]
- Damiani, N.; Fernandez, N.J.; Maldonado, L.M.; Alvarez, A.R.; Eguaras, M.J.; Marcangeli, J.A. Bioactivity of propolis from different geographical origins on Varroa destructor (acari: Varroidae). Parasitol. Res. 2010, 107, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Popova, M.; Reyes, M.; Le Conte, Y.; Bankova, V. Propolis chemical composition and honeybee resistance against Varroa destructor. Nat. Prod. Res. 2014, 28, 788–794. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.; Eischen, F.; Giannasi, D. Chemical composition of north american bee propolis and biological activity towards larvae of greater wax moth (lepidoptera: Pyralidae). J. Chem. Ecol. 1994, 20, 1783–1791. [Google Scholar] [CrossRef] [PubMed]
- Garedew, A.; Schmolz, E.; Lamprecht, I. Effect of the bee glue (propolis) on the calorimetrically measured metabolic rate and metamorphosis of the greater wax moth Galleria mellonella. Thermochim. Acta 2004, 413, 63–72. [Google Scholar] [CrossRef]
- Shaver, T.; Lukefahr, M. Effect of flavonoid pigments and gossypol on growth and development of the bollworm, tobacco budworm, and pink bollworm. J. Econ. Entomol. 1969, 62, 643–646. [Google Scholar] [CrossRef]
- Kato, M. Phenols as indispensible components of the synthetic diet of the silkworm, Bombyx mori. Entomol. Exp. Appl. 1978, 24, 485–490. [Google Scholar] [CrossRef]
- Neumann, P.; Pirk, C.; Hepburn, H.; Solbrig, A.; Ratnieks, F.; Elzen, P.; Baxter, J. Social encapsulation of beetle parasites by cape honeybee colonies (Apis mellifera capensis esch.). Naturwissenschaften 2001, 88, 214–216. [Google Scholar] [PubMed]
- Ellis, J.D.; Hepburn, H.R. A note on mapping propolis deposits in cape honey bee (Apis mellifera capensis) colonies. Afr. Entomol. 2003, 11, 122–124. [Google Scholar]
- Ellis, J. Life behind bars: Why honey bees feed small hive beetles. Am. Bee J. 2002, 142, 267–269. [Google Scholar]
- Ellis, J.D.; Hepburn, H.R.; Ellis, A.M.; Elzen, P.J. Social encapsulation of the small hive beetle (Aethina tumida murray) by european honeybees (Apis mellifera L.). Insectes Soc. 2003, 50, 286–291. [Google Scholar] [CrossRef]
- Hoyt, M. The World of Bees; Coward McCann Inc.: New York, NY, USA, 1965. [Google Scholar]
- Bogdanov, S. Contaminants of bee products. Apidologie 2006, 37, 1–18. [Google Scholar] [CrossRef]
- Bogdanov, S.; Kilchenmann, V.; Imdorf, A. Acaricide residues in some bee products. J. Apic. Res. 1998, 37, 57–67. [Google Scholar] [CrossRef]
- Wallner, K. Varroacides and their residues in bee products. Apidologie 1999, 30, 235–248. [Google Scholar] [CrossRef]
- Zhou, J.; Xue, X.; Li, Y.; Zhang, J.; Chen, F.; Wu, L.; Chen, L.; Zhao, J. Multiresidue determination of tetracycline antibiotics in propolis by using hplc-uv detection with ultrasonic-assisted extraction and two-step solid phase extraction. Food Chem. 2009, 115, 1074–1080. [Google Scholar] [CrossRef]
- Chen, F.; Chen, L.; Wang, Q.; Zhou, J.; Xue, X.; Zhao, J. Determination of organochlorine pesticides in propolis by gas chromatography–electron capture detection using double column series solid-phase extraction. Anal. Bioanal. Chem. 2009, 393, 1073–1079. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Parada, A.; Colazzo, M.; Besil, N.; Geis-Asteggiante, L.; Rey, F.; Heinzen, H. Determination of coumaphos, chlorpyrifos and ethion residues in propolis tinctures by matrix solid-phase dispersion and gas chromatography coupled to flame photometric and mass spectrometric detection. J. Chromatogr. A 2011, 1218, 5852–5857. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Tejada, G.M.; Medina-Peralta, S.; Moguel-Ordóñez, Y.B.; Muñoz-Rodríguez, D. Matrix solid-phase dispersion extraction of organophosphorus pesticides from propolis extracts and recovery evaluation by gc/ms. Anal. Bioanal. Chem. 2011, 400, 885–891. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, T.F.S.; Aquino, A.; Dórea, H.S.; Navickiene, S. Mspd procedure for determining buprofezin, tetradifon, vinclozolin, and bifenthrin residues in propolis by gas chromatography–mass spectrometry. Anal. Bioanal. Chem. 2008, 390, 1425–1430. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.M. Honey bee toxicology. Annu. Rev. Entomol. 2015, 60, 415–434. [Google Scholar] [CrossRef] [PubMed]
- Frazier, M.; Mullin, C.; Frazier, J.; Ashcraft, S. What have pesticides got to do with it? Am. Bee J. 2008, 148, 521–524. [Google Scholar]
- vanEngelsdorp, D.; Evans, J.D.; Donovall, L.; Mullin, C.; Frazier, M.; Frazier, J.; Tarpy, D.R.; Hayes, J., Jr.; Pettis, J.S. “Entombed pollen”: A new condition in honey bee colonies associated with increased risk of colony mortality. J. Invertebr. Pathol. 2009, 101, 147–149. [Google Scholar] [CrossRef] [PubMed]
- Tautz, J. The Buzz about Bees: Biology of a Superorganism; Springer: Berlin, Germany, 2008. [Google Scholar]
- Brütsch, T.; Jaffuel, G.; Vallat, A.; Turlings, T.C.J.; Chapuisat, M. Wood ants produce a potent antimicrobial agent by applying formic acid on tree-collected resin. Ecol. Evol. 2017, 7, 2249–2254. [Google Scholar] [CrossRef] [PubMed]
- Harman, D. Aging: A theory based on free radical and radiation chemistry. J. Gerontol. 1956, 11, 298–300. [Google Scholar] [CrossRef] [PubMed]
- Camici, G.G.; Shi, Y.; Cosentino, F.; Francia, P.; Luscher, T.F. Anti-aging medicine: Molecular basis for endothelial cell-targeted strategies—A mini-review. Gerontology 2011, 57, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Yonar, S.M.; Ural, M.S.; Silici, S.; Yonar, M.E. Malathion-induced changes in the haematological profile, the immune response, and the oxidative/antioxidant status of cyprinus carpio carpio: Protective role of propolis. Ecotoxicol. Environ. Saf. 2014, 102, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, H.; Satoh, K.; Tobe, T.; Yasuda, I.; Ushio, F.; Matsumoto, K.; Endo, K.; Ookubo, C. Free radical scavenging activity of propolis. Redox Rep. 2002, 7, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Farooqui, T.; Farooqui, A.A. Beneficial effects of propolis on human health and neurological diseases. Front. Biosci. 2012, 4, 779–793. [Google Scholar] [CrossRef]
- Seehuus, S.C.; Norberg, K.; Gimsa, U.; Krekling, T.; Amdam, G.V. Reproductive protein protects functionally sterile honey bee workers from oxidative stress. Proc. Natl. Acad. Sci. USA 2006, 103, 962–967. [Google Scholar] [CrossRef] [PubMed]
- Li-Byarlay, H.; Huang, M.H.; Simone-Finstrom, M.; Strand, M.K.; Tarpy, D.R.; Rueppell, O. Honey bee (Apis mellifera) drones survive oxidative stress due to increased tolerance instead of avoidance or repair of oxidative damage. Exp. Gerontol. 2016, 83, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.B.; Roberts, S.P.; Elekonich, M.M. Age and natural metabolically-intensive behavior affect oxidative stress and antioxidant mechanisms. Exp. Gerontol. 2008, 43, 538–549. [Google Scholar] [CrossRef] [PubMed]
- Simone-Finstrom, M.; Li-Byarlay, H.; Huang, M.H.; Strand, M.K.; Rueppell, O.; Tarpy, D.R. Migratory management and environmental conditions affect lifespan and oxidative stress in honey bees. Sci. Rep. 2016, 6, 32023. [Google Scholar] [CrossRef] [PubMed]
- Butler, C. The Honeybee: An Introduction to Her Sense-Physiology and Behaviour; Oxford University Press: London, UK, 1949. [Google Scholar]
- Page, R.E.; Fondrk, M.K. The effects of colony-level selection on the social organization of honey bee (Apis mellifera L.) colonies: Colony-level components of pollen hoarding. Behav. Ecol. Sociobiol. 1995, 36, 135–144. [Google Scholar] [CrossRef]
- Nakamura, J.; Seeley, T.D. The functional organization of resin work in honeybee colonies. Behav. Ecol. Sociobiol. 2006, 60, 339–349. [Google Scholar] [CrossRef]
- Martinez, O.A.; Soares, A.E.E. Melhoramento genético na apicultura comercial para produção da própolis. Rev. Bras. Saúde Prod. Anim. 2012, 13, 982–990. [Google Scholar] [CrossRef]
- Simone-Finstrom, M.; Gardner, J.; Spivak, M. Tactile learning in resin foraging honeybees. Behav. Ecol. Sociobiol. 2010, 64, 1609–1617. [Google Scholar] [CrossRef]
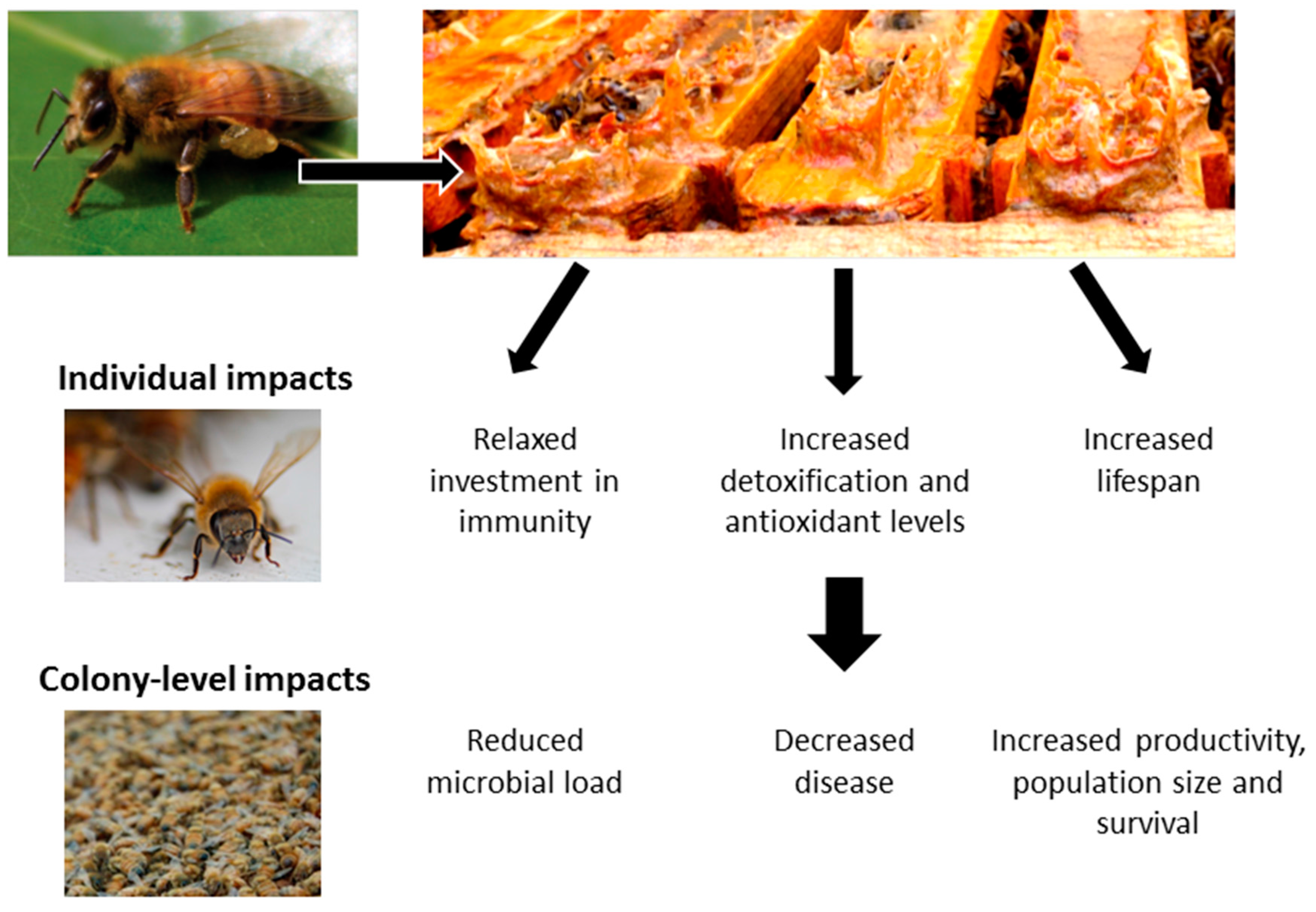

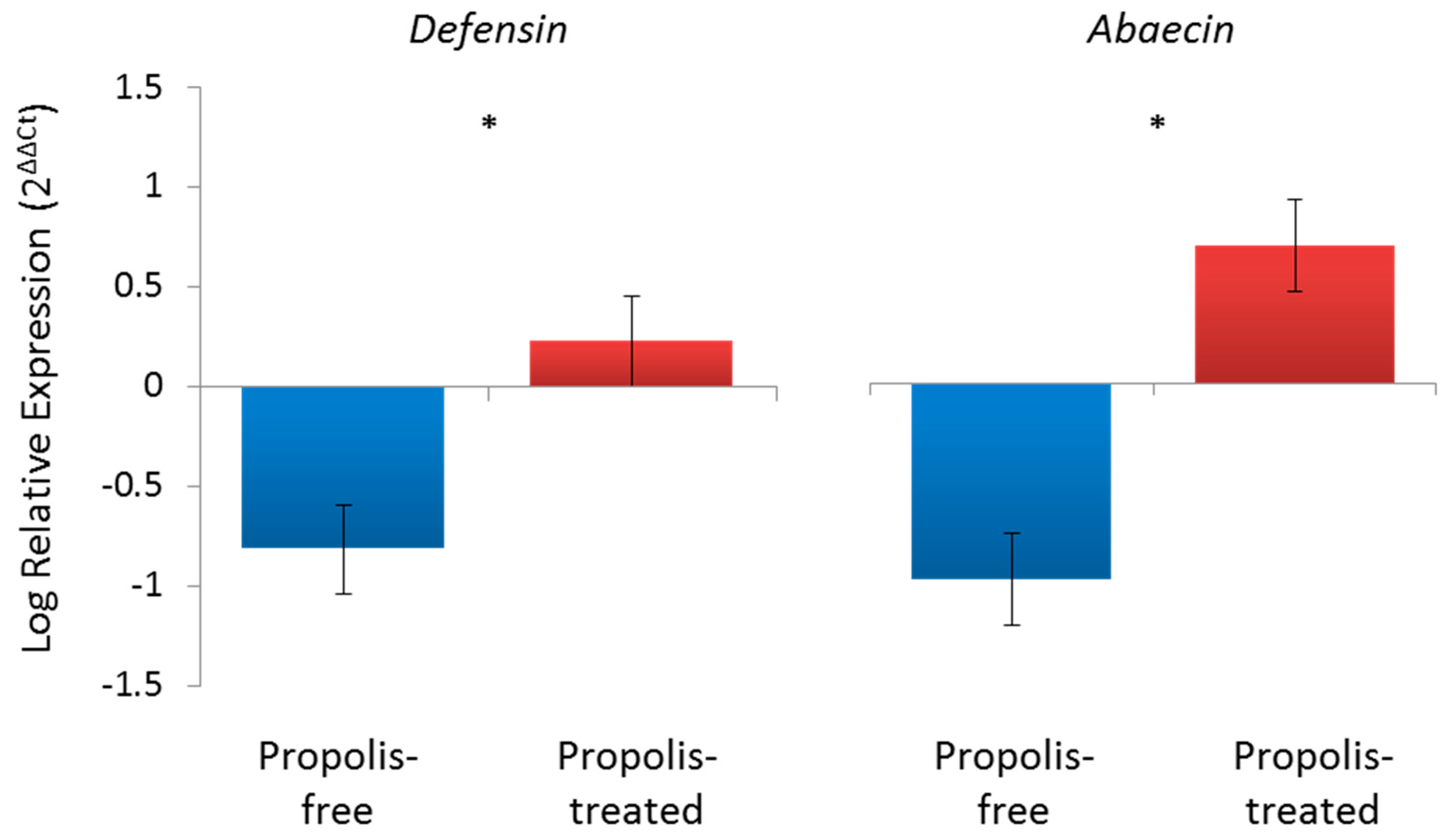
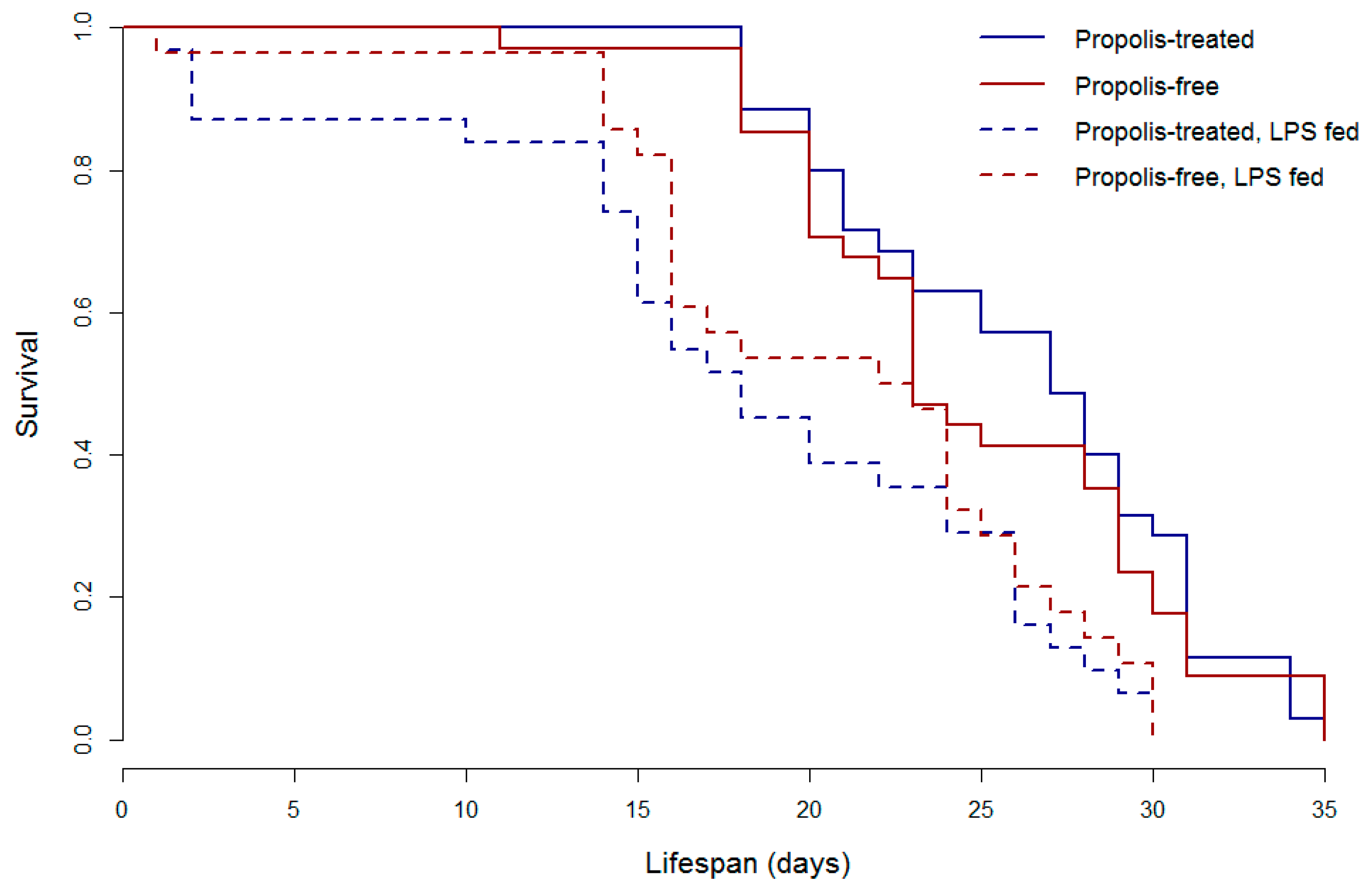

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simone-Finstrom, M.; Borba, R.S.; Wilson, M.; Spivak, M. Propolis Counteracts Some Threats to Honey Bee Health. Insects 2017, 8, 46. https://doi.org/10.3390/insects8020046
Simone-Finstrom M, Borba RS, Wilson M, Spivak M. Propolis Counteracts Some Threats to Honey Bee Health. Insects. 2017; 8(2):46. https://doi.org/10.3390/insects8020046
Chicago/Turabian StyleSimone-Finstrom, Michael, Renata S. Borba, Michael Wilson, and Marla Spivak. 2017. "Propolis Counteracts Some Threats to Honey Bee Health" Insects 8, no. 2: 46. https://doi.org/10.3390/insects8020046