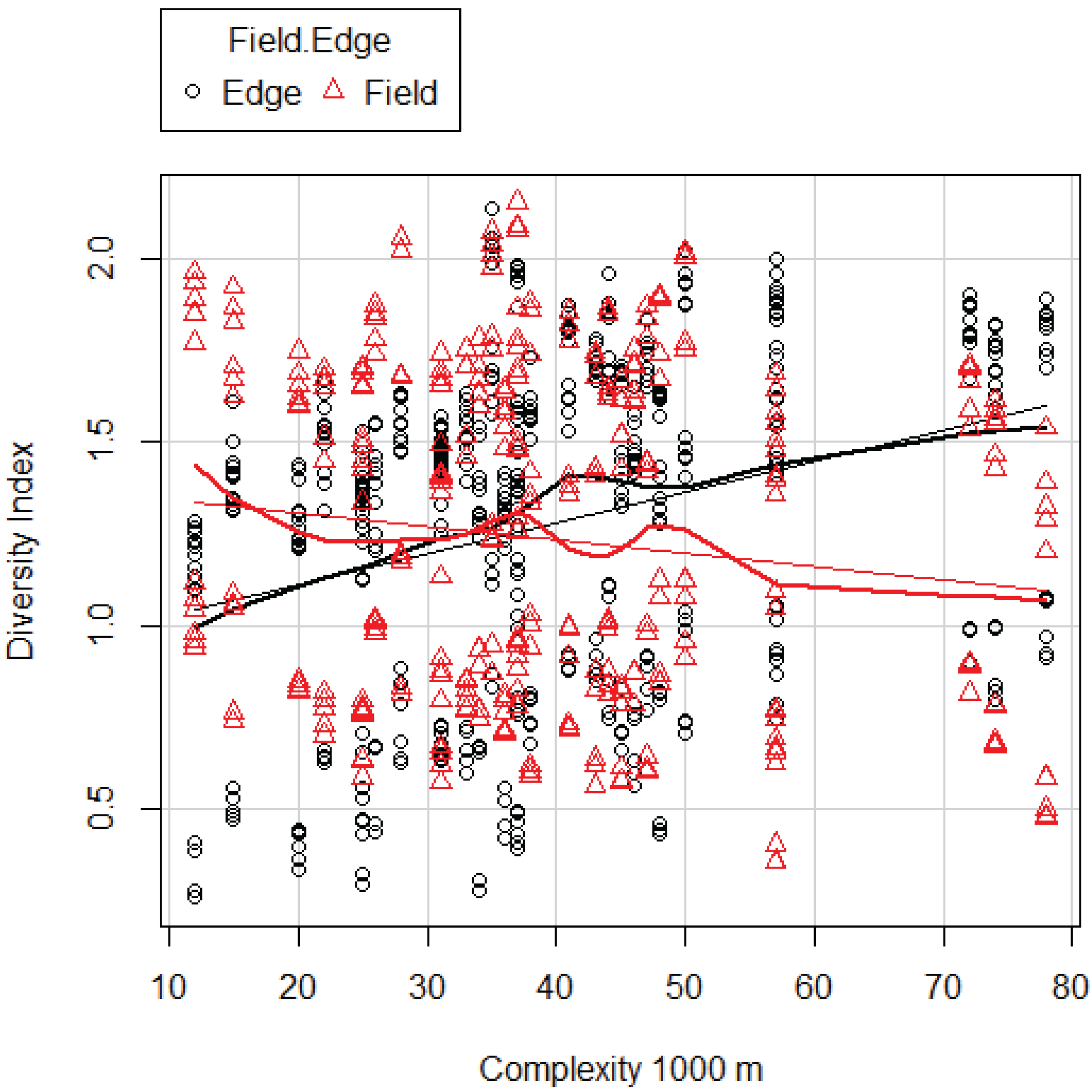
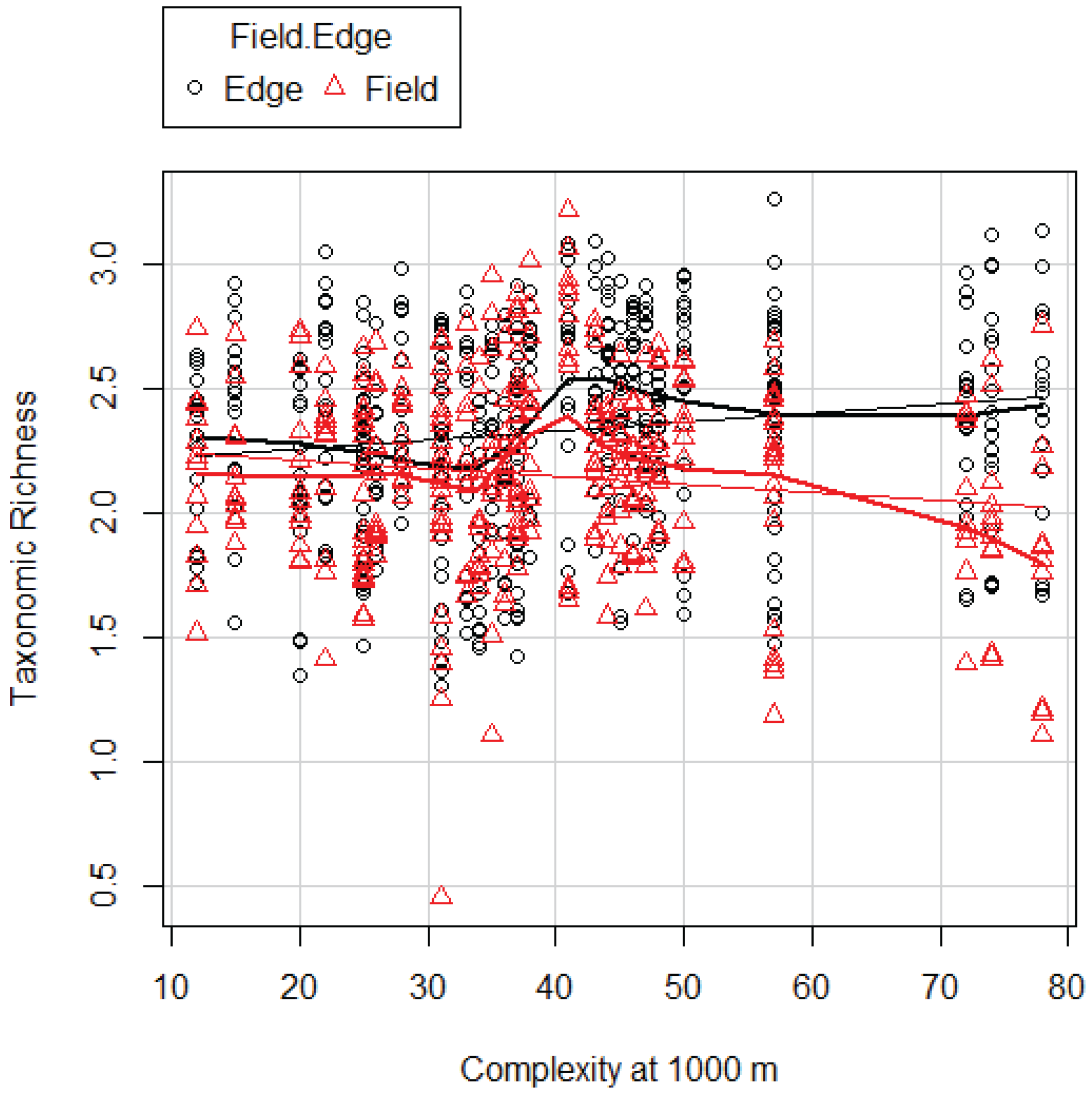
Based on our analysis we developed a snapshot of invertebrate communities early in the growing season. We uncovered significant differences between fields and edges in the way these communities were affected by landscape complexity. We found it was affected by complexity at the landscape scale of 1000 m and 6000 m. DI was affected by complexity at 1000 m. Both TR and DI increased with increasing complexity in FE and surprisingly decreased with increasing complexity in FI. Here we showed that the patterns of both TR and DI in fields and edges were identical as far as the relationship with landscape complexity was concerned.
3.2.1. Difference between FE and FI
The difference in TR between edges (FE) and fields (FI) seems small, however was statistically significant. Greater differences have been reported in a south African study in corn fields [
43] and in European studies that examined a large variety of field edges as they relate to organic farming [
44], ecological compensation meadows [
45] and ditch banks [
20].
Boundaries between “habitat” and “non-habitat” are often clearly identifiable to the human eye and presumably also to invertebrates [
46]. The landscapes in our study had a large percentage of areas that would be considered usable habitat (forests, grassland,
etc.) for invertebrates with the agricultural FI presumably at the lower end of any habitat ranking (
Table 1). It is therefore not surprising that FE had more invertebrates than FI. Agricultural FI recently disturbed by planting would appear to be even more lacking in habitat value. Our results, however, are consistent with the positive values of soil loosened by planting, warmed by exposure to sunlight, and drained of heavy spring rains. Many invertebrates deposit eggs into the soil; larvae feed on underground roots and detritus; they pupate and emerge as adults to mate. Ants and the larvae of some beetles, moths, flies and worms transport below ground materials to above ground consumers [
47]. The agricultural FI thus have habitat value and are not “non-habitat”. Invertebrates that occupy the FI are obviously “adapted” to high disturbance levels and monoculture vegetation. They are frequently generalists that have a small body size and short life-cycle [
48]. These generalists are successful and likely account for TR in the FI. Species requiring pristine environments, undisturbed habitats or that have limited dispersal ranges can be expected to be rare in the FI, but could find conditions in the FE that provide a great range of possible habitable environments with varied vegetation type, height, edge width,
etc.TR and DI are highly correlated but give different information. TR gives equal weight to rare taxa as common taxa; DI gives more emphasis on common taxa with little contribution from rare taxa. DI is a relative measure of evenness [
36]. This is an important component of biodiversity. Having great numbers of a single pest species and few predator species may result in the same TR as having balanced numbers of predator and prey. Knowing that the same patterns persist with TR and DI for FE and FI allows us to address important ecological food webs without harming agricultural yields.
Vegetation in the FE and non-arable areas was not classified as native or non-native in our study. Presumably the non-arable areas had mostly native species, while the FE had varying amounts of native plants. Native invertebrates have evolved in step with the native plants and would optimize interactions. The crops in the FI are non-native. Providing a complex native habitat should provide opportunities for native invertebrate species. Future studies should assess the relationship of invertebrate diversity to native vegetation in the FE.
Differences in agricultural practices may account for divergence from European studies. Studies of FI and FE in Europe deal with a vastly different agricultural system than we studied in central Illinois. Researchers of European agricultural systems have studied cereal crops, peas, potatoes, and sugar beets with small scale crop rotation [
22,
49]. GMO crops are infrequently sown and are prohibited in some nations [
50] while they are the norm for Illinois [
51]. Some FI are quite large in our study area (e.g., 117 ha) while many European study field interiors are considerably smaller, e.g., 22–30 ha (e.g., [
52]). Field edges in Europe are relatively stable and consistent temporally [
20]. In our study area, FE are frequently mown, treated with herbicide, exposed to de-icing chemicals and burned, and therefore show little contrast from FI in our study or in field edges in Europe.
Our study was conducted during a two-week period in early summer, not long after crops had been sown. The period before our study included major disturbance of the soil from cultivation in the FI and potential movement of invertebrates between FI and FE. Invertebrates vary in their timing of emergence from diapause. The crops were growing rapidly but not yet shading the ground between plants as occurs later in the growing season. Patterns observed at this time may not be the same as patterns later in the growing season. Future studies should look at TR and DI across the growing season.
Agricultural areas in many parts of Europe have been active for many centuries if not millennia and in Illinois somewhat less than two centuries [
53,
54]. Data from European studies may not be transferable to studies in Illinois and vice versa. That does not mean, however, that the management techniques that are shown to be effective in Europe would not be equally effective in Illinois. Practices that might be beneficial in Illinois include “beetle banks” [
14], reduction of chemical applications in conservation headlands and field margins [
55], and Ecological Conservation Area wildflower strips [
56].
3.2.2. Landscape Complexity
Most of the fields of our study (
n = 30) were located in an area (6000 m scale) of high non-agricultural complexity of 16%–30% (
n = 14), 30%–40% (
n = 13) and >40% (
n = 3). This is unexpected in an area known as the “corn desert” [
57] and may be related to our selection of structurally diverse FE in proximity to natural areas. Assuming that the intermediate landscape complexity hypothesis [
3] is also applicable in the agricultural areas of the Midwest, this would mean that agri-environmental measures to increase biodiversity would have little effect in our study area: the TR and DI are already optimal for an agricultural area. This idea is supported by the fact that the TR we found in the FI is relatively high and close to the TR in the FE (
ca. 9
vs. ca. 11 taxa per sample on average). However, it should be noted that our study areas may not be representative of the rest of the Midwest.
Local communities are firstly dependent on the regional pool of species [
58]. Local TR is expected to be larger within areas of greater landscape complexity [
22], because increased complexity increases the regional pool from which to draw local communities [
28]. Therefore the higher TR that we found in the FE in more complex landscapes fits this species pool hypothesis.
However, according to the species pool hypothesis, we would also expect higher TR in the FI. The pattern that emerges in our data shows that as TR in the FE increases, the TR in the FI decreases. There are several possible explanations of this deviation from expectation, which could be examined in further studies. First, because of the higher TR in the more complex landscapes, the predation pressure on invertebrates could be higher, either by other invertebrates or by vertebrates that were not measured by us [
54,
59]. In the FE more invertebrate species would be able to escape this increased pressure because of the higher vegetation density, while in the fields the invertebrates are more vulnerable to predation. This would fit Tscharntke and coworkers’ [
60] hypothesis that landscape complexity provides spatial and temporal insurance, which would mean in this case a more efficient regulation of pest species populations in the FI.
Second, another explanation is that invertebrates prefer the FE habitat and non-agricultural landscape elements even when they are able to occupy the FI niche. In this case, the data may reflect species moving into the FI only when they have no other option, e.g., when individual numbers are high in the FE or resources are depleted, but no escape to other landscape elements than FI is possible. When there is other, more suitable habitat available, that is where invertebrates will occur.
Third, it is intriguing to consider that plant-to-plant interactions in more complex areas may provide a defense for the FI [
61,
62]. This might involve a signal sent by plants in the edge in response to herbivory being received by the crops in the FI [
63]. Because the FI are a monoculture, the response spreads through the entire FI providing protection against the herbivores either repelling the herbivores or calling predators or perhaps both [
64,
65]. This effect could be masked in the FE by the dense vegetation.
We tested whether the study areas within the three counties varied significantly in their complexity (
Table 1). The study areas were not randomly selected; they were selected because of their proximity to non-arable land within the agricultural landscape and did not include urban areas within the buffer circles. The three counties were typical of the Illinois landscape, but the study areas selected were probably more complex than the remainder of the land in the counties. These areas were not necessarily representative of either the rest of county, state or even Midwest.
3.2.3. Confounding Variables
A drought period began in the summer of 2011 and continued through 2012 with higher than normal temperatures and lower than normal rainfall. We tested if the difference in TR and DI was significantly different between years (
Table 6). We do not know if the highly significant difference between years was typical of the normal variability of invertebrate populations or a product of the drought conditions.
We measured a number of confounding variables that we felt might impact TR and DI. Crops in the study areas were GM soybean and corn. Corn is usually the first crop to be sown when the fields dry out and start to warm. Soybeans are planted later. The planting dates and subsequent plant growth and shading may have influenced colonization by invertebrates. Price [
66] found that herbivores colonized the soybeans first with no appreciable increase in parasites and predators until the canopy had developed. Botha [
43] found that biodiversity loss was apparent if corn fields were within 30 m of the field margins being sampled. Therefore it was no surprise that crop had an effect in our study.
All FI were tilled before planting and may or may not have been recently treated with glyphosate (broad spectrum herbicide). Herbicides vary in their impact on invertebrates and the impact often depends on the timing and context of the application [
67,
68]. Invertebrates vary greatly in their mobility and dispersal ability. In agricultural landscapes with high disturbance particularly in the planting and harvesting phases the dispersal technique is crucial for survival [
3]. In addition, crossing hard barriers, such as roadways, limits the mobility of arthropods [
69]. These issues are outside the scope of this study but should be acknowledged as having an impact.
As the length of the FE increased, the TR and DI decreased. The edge along the fields may serve as a corridor for migration as well as a refuge during episodes of disturbance. The distance to additional field edges increases vulnerability of invertebrates with low mobility.
The distance to the nearest non-arable space > 1 ha was important to TR but had no significant impact on DI. We did not collect from the nearest non-arable space and cannot say how the TR and DI compared to our study fields. Gonzalez [
27] found both forest cover and proximity affected arthropod assemblages in soybean fields in central Argentina.
We measured a number of other factors which did not significantly contribute to our findings (
Table 2). These included the size of the agricultural field, the width of the FE, the average vegetation height, vegetation standard deviation (sd) and soil type. These were local factors within the agricultural landscape that affect other groups of organisms such as birds [
70,
71], and mammals [
72,
73], but not some reptiles [
74].


 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}