
Insect Cuticular Hydrocarbons as Dynamic Traits in Sexual Communication

{kind=link}
Abstract
:1. Introduction
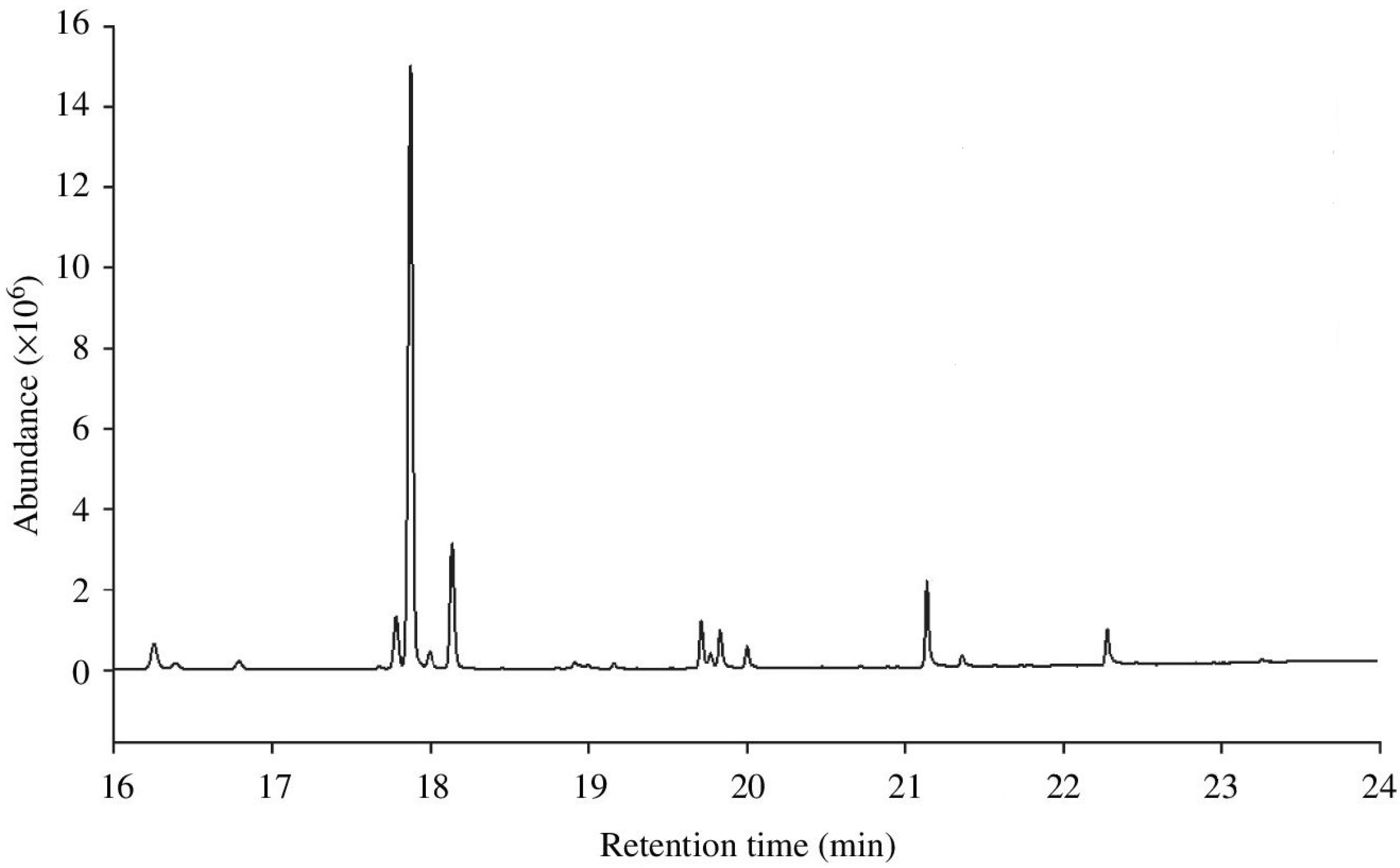
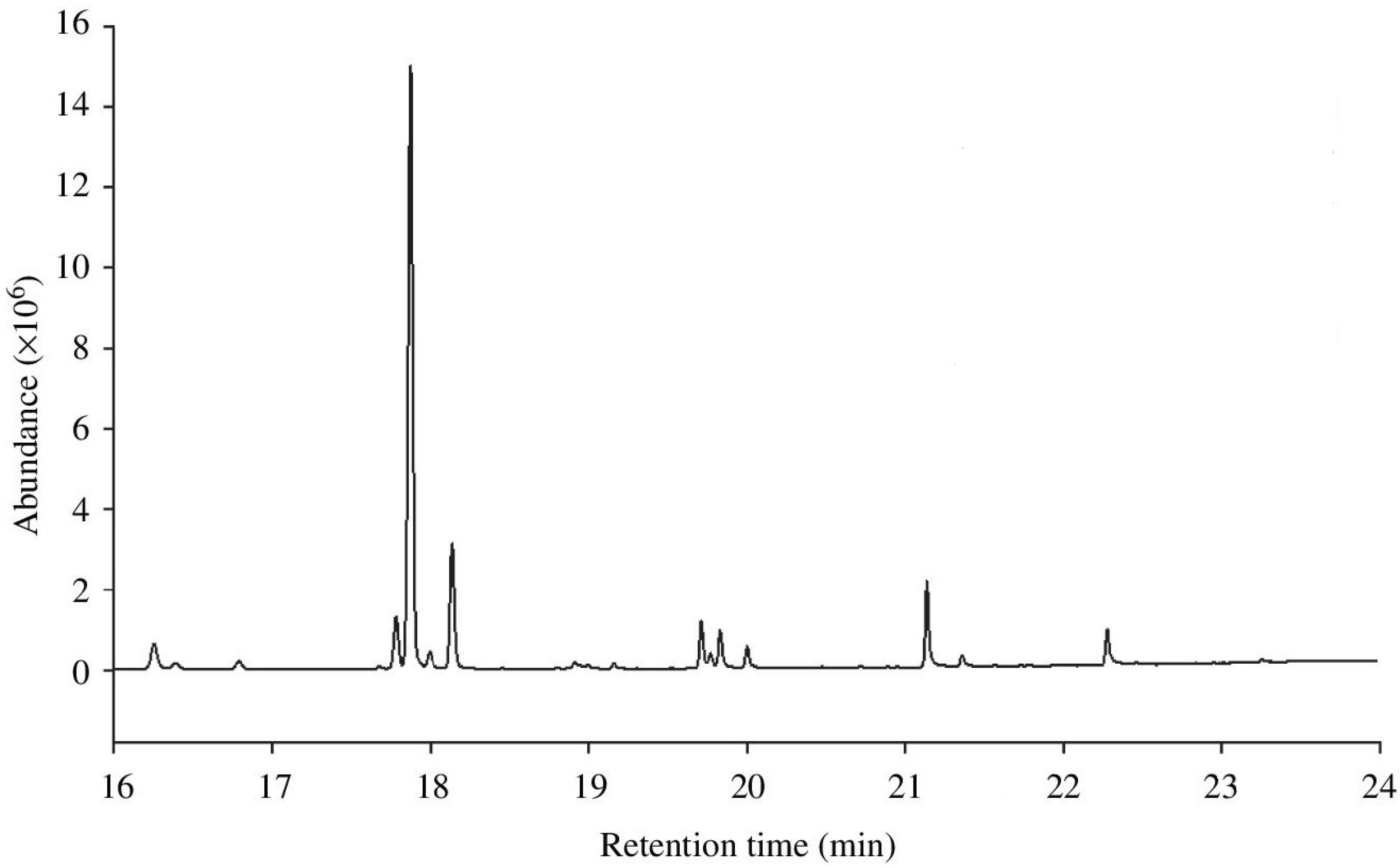
2. Variation in CHCs across Abiotic Environments
2.1. CHCs as Condition-Dependent Sexual Signals
2.2. Potential Trade-Offs between Signalling CHCs and Waterproofing CHCs
2.3. The Consequences of Abiotic Environmental Variation in CHCs for Sexual Signalling
3. The Role of the Social Environment in CHC Expression
4. Conclusions
Acknowledgements
Conflicts of Interest
References
- Ferveur, J.F. Cuticular hydrocarbons: Their evolution and roles in Drosophila pheromonal communication. Behav. Gen. 2005, 35, 279–295. [Google Scholar] [CrossRef] [PubMed]
- Hadley, N.F. Cuticular lipids of terrestrial plants and arthropods: A comparison of their structure, composition, and waterproofing function. Biol. Rev. 1981, 56, 23–47. [Google Scholar] [CrossRef]
- Blomquist, G.J. Biosynthesis of cuticular hydrocarbons. In Insect Hydrocarbons; Blomquist, G.J., Bagneres, A.G., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 35–51. [Google Scholar]
- Singer, T.L. Roles of hydrocarbons in the recognition systems of insects. Am. Zool. 1998, 38, 394–405. [Google Scholar] [CrossRef]
- Martin, S.J.; Drijfhout, F.P. How reliable is the analysis of complex cuticular hydrocarbon profiles by multivariate statistical methods? J. Chem. Ecol. 2009, 35, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Blomquist, G.J. Structure and analysis of insect hydrocarbons. In Insect Hydrocarbons; Blomquist, G.J., Bagneres, A.G., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 19–34. [Google Scholar]
- Ferveur, J.F.; Cobb, M. Behavioral and evolutionary roles of cuticular hydrocarbons in Diptera. In Insect Hydrocarbons; Blomquist, G.J., Bagneres, A.G., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 19–34. [Google Scholar]
- Neems, R.M.; Butlin, R.K. Variation in cuticular hydrocarbons across a hybrid zone in the grasshopper. Proc. R Soc. Lond. B 1995, 257, 135–140. [Google Scholar] [CrossRef]
- Jallon, J.M.; David, J.R. Variations in cuticular hydrocarbons among the eight species of the Drosophila melanogaster subgroup. Evolution 1987, 41, 294–302. [Google Scholar] [CrossRef]
- Niehuis, O.; Bullesbach, J.; Judson, A.K.; Schmitt, T.; Gadau, J. Genetics of cuticular hydrocarbon differences between males of the parasitoid wasps Nasonia giraulti and Nasonia vitripennis. Heredity 2011, 107, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Mas, F.; Jallon, J.M. Sexual isolation and cuticular hydrocarbon differences between Drosophila santomea and Drosophila yakuba. J. Chem. Ecol. 2005, 31, 2747–2752. [Google Scholar] [CrossRef] [PubMed]
- Higgie, M.; Chenoweth, S.F.; Blows, M.W. Natural selection and the reinforcement of mate recognition. Science 2000, 290, 519–521. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, T.D. Pheromones and Animal Behaviour; Cambridge University Press: New York, NY, USA, 2003. [Google Scholar]
- Ingleby, F.C.; Hosken, D.J.; Flowers, K.; Hawkes, M.F.; Lane, S.M.; Rapkin, J.; House, C.M.; Hunt, J. Environmental heterogeneity, multivariate sexual selection and genetic constraints on cuticular hydrocarbons in Drosophila simulans. J. Evol. Biol. 2014, 27, 700–713. [Google Scholar] [CrossRef] [PubMed]
- Bontonou, G.; Wicker-Thomas, C. Sexual communication in the Drosophila genus. Insects 2014, 5, 439–458. [Google Scholar] [CrossRef]
- Carlson, D.A.; Mayer, M.S.; Silhacek, D.L.; James, J.D.; Beroza, M.; Bierl, B.A. Sex attractant pheromone of the house fly: Isolation, identification and synthesis. Science 1971, 174, 76–78. [Google Scholar] [CrossRef] [PubMed]
- Antony, C.; Davis, T.L.; Carlson, D.A.; Pechine, J.M.; Jallon, J.M. Compared behavioural responses of male Drosophila melanogaster to natural and synthetic aphrodisiacs. J. Chem. Ecol. 1985, 11, 1617–1629. [Google Scholar] [CrossRef] [PubMed]
- Cobb, M.; Jallon, J.M. Pheromones, mate recognition and courtship stimulation in the Drosophila melanogaster species sub-group. Anim. Behav. 1990, 39, 1058–1067. [Google Scholar] [CrossRef]
- Gibbs, A.G.; Louie, A.K.; Ayala, J.A. Effects of temperature on cuticular lipids and water balance in a desert Drosophila: Is thermal acclimation beneficial? J. Exp. Biol. 1998, 201, 71–80. [Google Scholar] [PubMed]
- Foley, B.R.; Telonis-Scott, M. Quantitative genetic analysis suggests causal association between cuticular hydrocarbon composition and desiccation survival in Drosophila melanogaster. Heredity 2011, 106, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Golebiowski, M.; Bogus, M.I.; Paszkiewicz, M.; Stepnowski, P. Cuticular lipids of insects as potential biofungicides: Methods of lipid composition analysis. Anal. Bioanal. Chem. 2011, 399, 3177–3191. [Google Scholar] [CrossRef] [PubMed]
- Byrne, P.G.; Rice, W.R. Evidence for adaptive male mate choice in the fruit fly Drosophila melanogaster. Proc. R Soc. Lond. B 2006, 273, 917–922. [Google Scholar] [CrossRef] [PubMed]
- Chenoweth, S.F.; Blows, M.W. Contrasting mutual sexual selection on homologous signal traits in Drosophila serrata. Am. Nat. 2005, 165, 281–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, M.A.; Dobler, S.; Larson, E.L.; Juarez, D.; Schlarbaum, T.; Monsen, K.J.; Francke, W. Profiles of cuticular hydrocarbons mediate male mate choice and sexual isolation between hydridising Chrysochus. Chemoecology 2007, 17, 87–96. [Google Scholar] [CrossRef]
- Thomas, M.L.; Simmons, L.W. Cuticular hydrocarbons influence female attractiveness to males in the Australian field cricket, Teleogryllus oceanicus. J. Evol. Biol. 2010, 23, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Rundle, H.D.; Chenoweth, S.F.; Blows, M.W. The diversification of mate preferences by natural and sexual selection. J. Evol. Biol. 2009, 22, 1608–1615. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.L.; Simmons, L.W. Sexual selection on cuticular hydrocarbons in the Australian field cricket, Teleogryllus oceanicus. BMC Evol. Biol. 2009, 9. [Google Scholar] [CrossRef] [PubMed]
- Ingleby, F.C.; Hunt, J.; Hosken, D.J. Genotype-by-environment interactions for female mate choice of male cuticular hydrocarbons in Drosophila simulans. PLoS ONE 2013, 8, e67623. [Google Scholar] [CrossRef] [PubMed]
- Grillet, M.; Dartevelle, L.; Ferveur, J.F. A Drosophila male pheromone affects female sexual receptivity. Proc. R Soc. Lond. B 2006, 273, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Jennings, J.H.; Etges, W.J.; Schmitt, T.; Hoikkala, A. Cuticular hydrocarbons of Drosophila montana: Geographic variation, sexual dimorphism and potential roles as pheromones. J. Insect Physiol. 2014, 61, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Bretman, A.; Westmancoat, J.D.; Gage, M.J.G.; Chapman, T. Males use multiple, redundant cues to detect mating rivals. Curr. Biol. 2011, 21, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Lane, S.M.; Solino, J.H.; Mitchell, C.; Blount, J.D.; Okada, K.; Hunt, J.; House, C.M. Rival male chemical cues evoke changes in male pre- and post-copulatory investment in a flour beetle. Behav. Ecol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Marcillac, F.; Ferveur, J.F. A set of female pheromones affects reproduction before, during and after mating in Drosophila. J. Exp. Biol. 2004, 207, 3927–3933. [Google Scholar] [CrossRef] [PubMed]
- Steiger, S.; Peschke, K.; Francke, W.; Müller, J.K. The smell of parents: Breeding status influences cuticular hydrocarbon pattern in the burying beetle Nicrophorus vespilloides. Proc. R. Soc. Lond. B 2007, 274, 2211–2220. [Google Scholar] [CrossRef] [PubMed]
- Yew, J.Y.; Dreisewerd, K.; Oliveira, C.C.; Etges, W.J. Male-specific transfer and fine scale spatial differences of newly identified cuticular hydrocarbons and triacylglycerides in a Drosophila species pair. PLoS ONE 2011, 6, e16898. [Google Scholar] [CrossRef] [PubMed]
- Gosden, T.P.; Shastri, K.L.; Innocenti, P.; Chenoweth, S.F. The B-matrix harbors significant and sex-specific constraints on the evolution of multicharacter sexual dimorphism. Evolution 2012, 66, 2106–2116. [Google Scholar] [CrossRef] [PubMed]
- Buellesbach, J.; Gadau, J.; Beukeboom, L.W.; Echinger, F.; Raychoudhury, R.; Werren, J.H.; Schmitt, T. Cuticular hydrocarbon divergence in the jewel wasp Nasonia: Evolutionary shifts in chemical communication channels? J. Evol. Biol. 2013, 26, 2467–2478. [Google Scholar] [CrossRef] [PubMed]
- Rowe, L.; Houle, D. The lek paradox and the capture of genetic variance by condition dependent traits. Proc. R Soc. Lond. B 1996, 263, 1415–1421. [Google Scholar] [CrossRef]
- Cotton, S.; Small, J.; Pomiankowski, A. Sexual selection and condition-dependent mate preferences. Curr. Biol. 2006, 16, R755–R765. [Google Scholar] [CrossRef] [PubMed]
- Blows, M.W. Interaction between natural and sexual selection during the evolution of mate recognition. Proc. R Soc. Lond. B 2002, 269, 1113–1118. [Google Scholar] [CrossRef] [PubMed]
- Gosden, T.P.; Chenoweth, S.F. On the evolution of heightened condition dependence of male sexual displays. J. Evol. Biol. 2011, 24, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Delcourt, M.; Rundle, H.D. Condition dependence of a multicomponent sexual display trait in Drosophila serrata. Am. Nat. 2011, 177, 812–823. [Google Scholar] [CrossRef] [PubMed]
- Weddle, C.B.; Mitchell, C.; Bay, S.K.; Sakaluk, S.K.; Hunt, J. Sex-specific genotype-by-environment interactions for cuticular hydrocarbon expression in decorated crickets, Gryllodes sigillatus: Implications for the evolution of signal reliability. J. Evol. Biol. 2012, 25, 2112–2125. [Google Scholar] [CrossRef] [PubMed]
- Ingleby, F.C.; Hosken, D.J.; Flowers, K.; Hawkes, M.F.; Lane, S.M.; Rapkin, J.; Dworkin, I.; Hunt, J. Genotype-by-environment interactions for cuticular hydrocarbon expression in Drosophila simulans. J. Evol. Biol. 2013, 26, 94–107. [Google Scholar] [CrossRef] [PubMed]
- Etges, W.J.; Oliveira, C.C. Premating isolation is determined by larval rearing substrates in cactophilic Drosophila mojavensis. Ecol. Evol. 2014, 4, 2033–2045. [Google Scholar] [PubMed]
- Van Homrigh, A.; Higgie, M.; McGuigan, K.; Blows, M.W. The depletion of genetic variance by sexual selection. Curr. Biol. 2007, 17, 528–532. [Google Scholar] [CrossRef] [PubMed]
- Izzo, A.; Wells, M.; Huang, Z.; Tibbetts, E. Cuticular hydrocarbons correlate with fertility, not dominance, in a paper wasp, Polistes dominulus. Behav. Ecol. Sociobiol. 2010, 64, 857–864. [Google Scholar] [CrossRef]
- Liang, D.; Silverman, J. You are what you eat: Diet modifies cuticular hydrocarbons and nestmate recognition in the Argentine ant, Linepiithema humile. Naturwissenschaften 2000, 87, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Sorvari, J.; Theodora, P.; Turillazzi, S.; Hakkarainen, H.; Sundstrom, L. Food resources, chemical signaling, and nest mate recognition in the ant Formica aquilonia. Behav. Ecol. 2007, 19, 441–447. [Google Scholar] [CrossRef]
- Fedina, T.Y.; Kuo, T.H.; Dreisewerd, K.; Dierick, H.A.; Yew, J.Y.; Pletcher, S.D. Dietary effects on cuticular hydrocarbons and sexual attractiveness in Drosophila. PLoS ONE 2012, 7, e49799. [Google Scholar] [CrossRef] [PubMed]
- Steiger, S.; Ower, G.D.; Stokl, J.; Mitchell, C.; Hunt, J.; Sakaluk, S.K. Sexual selection on cuticular hydrocarbons of male sagebush crickets in the wild. Proc. R Soc. Lond. B 2013, 280, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Frentiu, F.D.; Chenoweth, S.F. Clines in cuticular hydrocarbons in two Drosophila species with independent population histories. Evolution 2010, 64, 1784–1794. [Google Scholar] [CrossRef] [PubMed]
- Savarit, F.; Ferveur, J.F. Temperature affects the ontogeny of sexually dimorphic cuticular hydrocarbons in Drosophila melanogaster. J. Exp. Biol. 2002, 205, 3241–3249. [Google Scholar] [PubMed]
- Roff, D.A. Evolutionary Quantitative Genetics; Chapman and Hall: New York, NY, USA, 1997. [Google Scholar]
- Hoffmann, A.A.; Merilä, J. Heritable variation and evolution under favourable and unfavourable conditions. Trends Evol. Ecol. 1999, 14, 96–101. [Google Scholar] [CrossRef]
- Greenfield, M.D.; Rodríguez, R.L. Genotype-environment interaction and the reliability of mating signals. Anim. Behav. 2004, 68, 1461–1468. [Google Scholar] [CrossRef]
- Rundle, H.D.; Chenoweth, S.F.; Blows, M.W. The roles of natural and sexual selection during adaptation to a novel environment. Evolution 2006, 60, 2218–2225. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.A. The Genetical Theory of Natural Selection; Clarendon Press: London, UK, 1930. [Google Scholar]
- Jia, F.Y.; Greenfield, M.D. When are good genes good? Variable outcomes of female choice in wax moths. Proc. R Soc. Lond. B 1997, 264, 1057–1063. [Google Scholar] [CrossRef]
- Ritchie, M.G.; Saarikettu, M.; Livingstone, S.; Hoikkala, A. Characterization of female preference functions for Drosophila montana courtship song and a test of the temperature coupling hypothesis. Evolution 2001, 55, 721–727. [Google Scholar] [CrossRef]
- Ingleby, F.C.; Hunt, J.; Hosken, D.J. Heritability of male attractiveness persists despite evidence for unreliable sexual signals in Drosophila simulans. J. Evol. Biol. 2013, 26, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Jennions, M.D.; Petrie, M. Variation in mate choice and mating preferences: A review of causes and consequences. Biol. Rev. 1997, 72, 283–327. [Google Scholar] [CrossRef] [PubMed]
- Delcourt, M.; Blows, M.W.; Rundle, H.D. Quantitative genetics of female mate preferences in an ancestral and a novel environment. Evolution 2010, 64, 2758–2766. [Google Scholar] [CrossRef] [PubMed]
- Petfield, D.; Chenoweth, S.F.; Rundle, H.D.; Blows, M.W. Genetic variance in female condition predicts indirect genetic variance in male sexual display traits. Proc. Natl. Acad. Sci. USA 2005, 102, 6045–6050. [Google Scholar] [CrossRef] [PubMed]
- Gershman, S.N.; Tournishey, E.; Rundle, H.D. Time flies: Time of day and social environment affect cuticular hydrocarbon sexual displays in Drosophila serrata. Proc. R Soc. Lond. B 2015, 281. [Google Scholar] [CrossRef] [PubMed]
- Kent, C.; Azanchi, R.; Smith, B.; Formosa, A.; Levine, J.D. Social context influences chemical communication in D. melanogaster males. Curr. Biol. 2008, 18, 1384–1389. [Google Scholar] [CrossRef] [PubMed]
- Krupp, J.J.; Kent, C.; Billeter, J.C.; Azanchi, R.; So, A.K.C.; Schonfeld, J.A.; Smith, B.P.; Lucas, C.; Levine, J.D. Social experience modifies pheromone expression and mating behavior in male Drosophila melanogaster. Curr. Biol. 2008, 18, 1373–1383. [Google Scholar] [CrossRef] [PubMed]
- Chenoweth, S.F.; Rundle, H.D.; Blows, M.W. Experimental evidence for the evolution of indirect genetic effects: Changes in the interaction effect coefficient, psi, due to sexual selection. Evolution 2010, 64, 1849–1856. [Google Scholar] [CrossRef] [PubMed]
- Bloch, G.; Hazan, E.; Rafaeli, A. Circadian rhythms and endocrine functions in adult insects. J. Insect Physiol. 2013, 59, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Wolf, J.B.; Brodie, E.D.; Moore, A.J. Interacting phenotypes and the evolutionary process. II. Selection resulting from social interactions. Am. Nat. 1999, 153, 254–266. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ingleby, F.C. Insect Cuticular Hydrocarbons as Dynamic Traits in Sexual Communication. Insects 2015, 6, 732-742. https://doi.org/10.3390/insects6030732
Ingleby FC. Insect Cuticular Hydrocarbons as Dynamic Traits in Sexual Communication. Insects. 2015; 6(3):732-742. https://doi.org/10.3390/insects6030732
Chicago/Turabian StyleIngleby, Fiona C. 2015. "Insect Cuticular Hydrocarbons as Dynamic Traits in Sexual Communication" Insects 6, no. 3: 732-742. https://doi.org/10.3390/insects6030732
APA StyleIngleby, F. C. (2015). Insect Cuticular Hydrocarbons as Dynamic Traits in Sexual Communication. Insects, 6(3), 732-742. https://doi.org/10.3390/insects6030732