
Foliar Substrate Affects Cuticular Hydrocarbon Profiles and Intraspecific Aggression in the Leafcutter Ant Atta sexdens

Abstract
:1. Introduction
2. Experimental Section
2.1. Diet Groups
2.2. Behavioral Assays

2.3. Chemical Analysis
3. Results
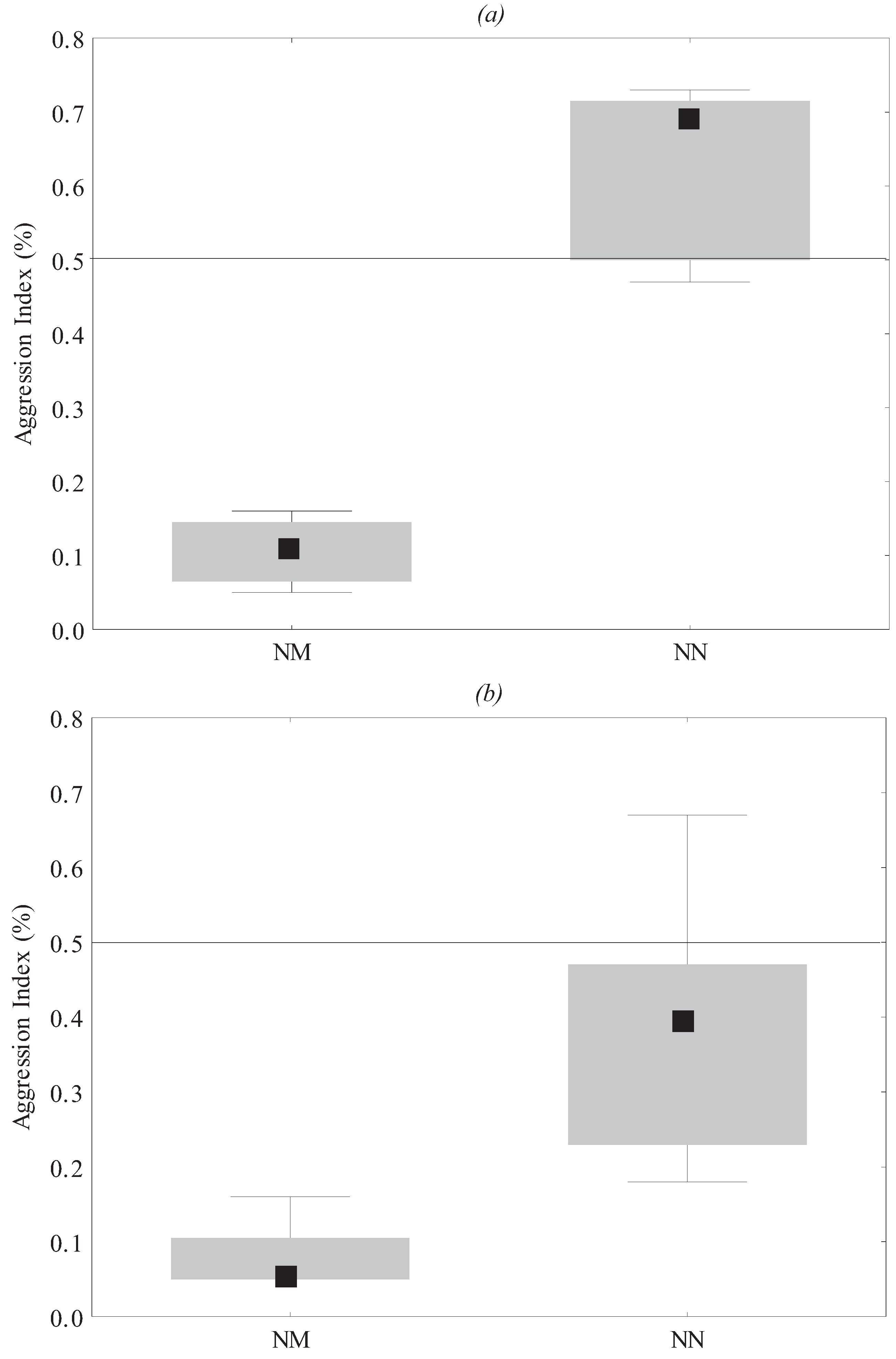
3.1. Behavioral Assays
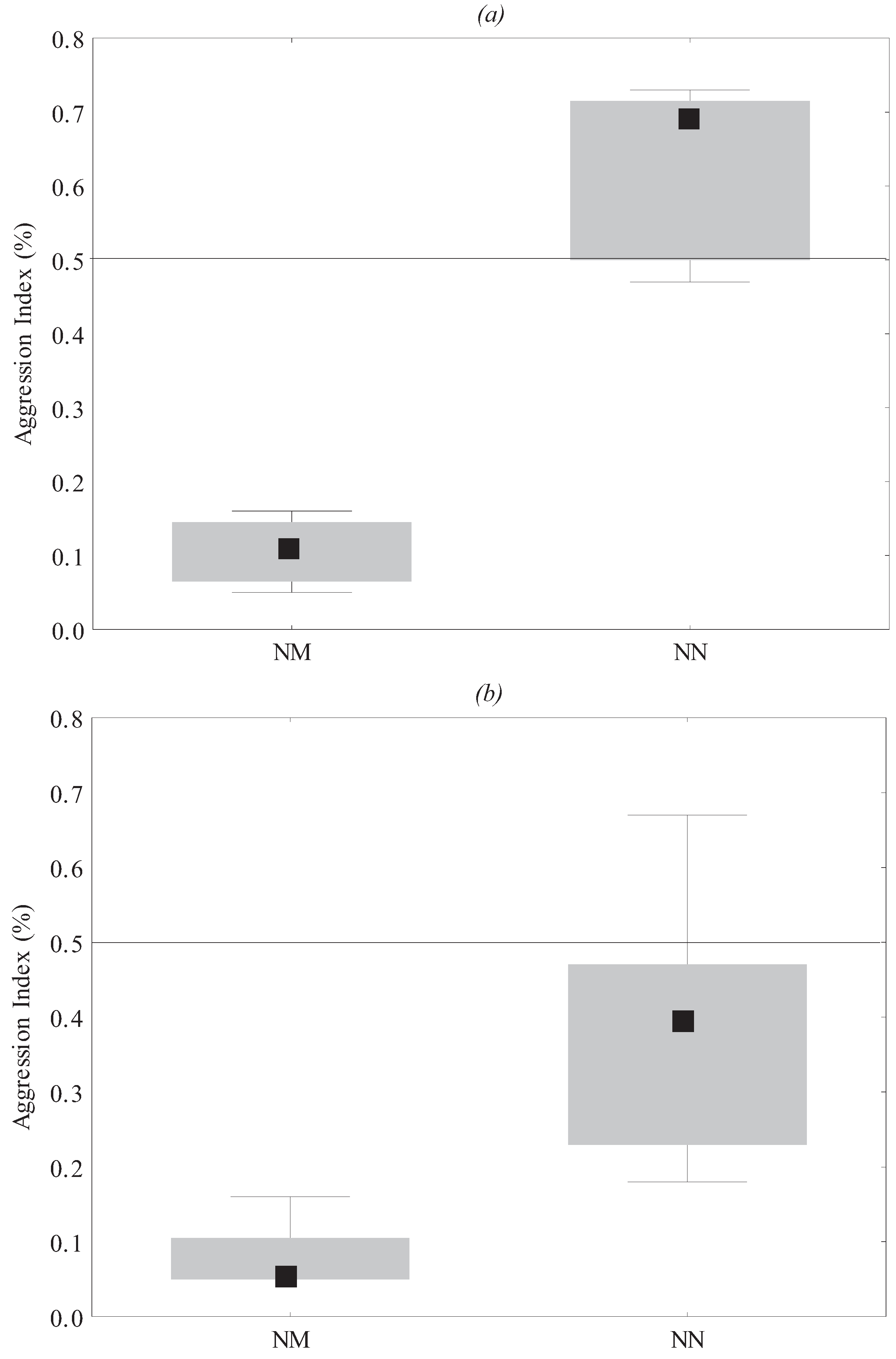
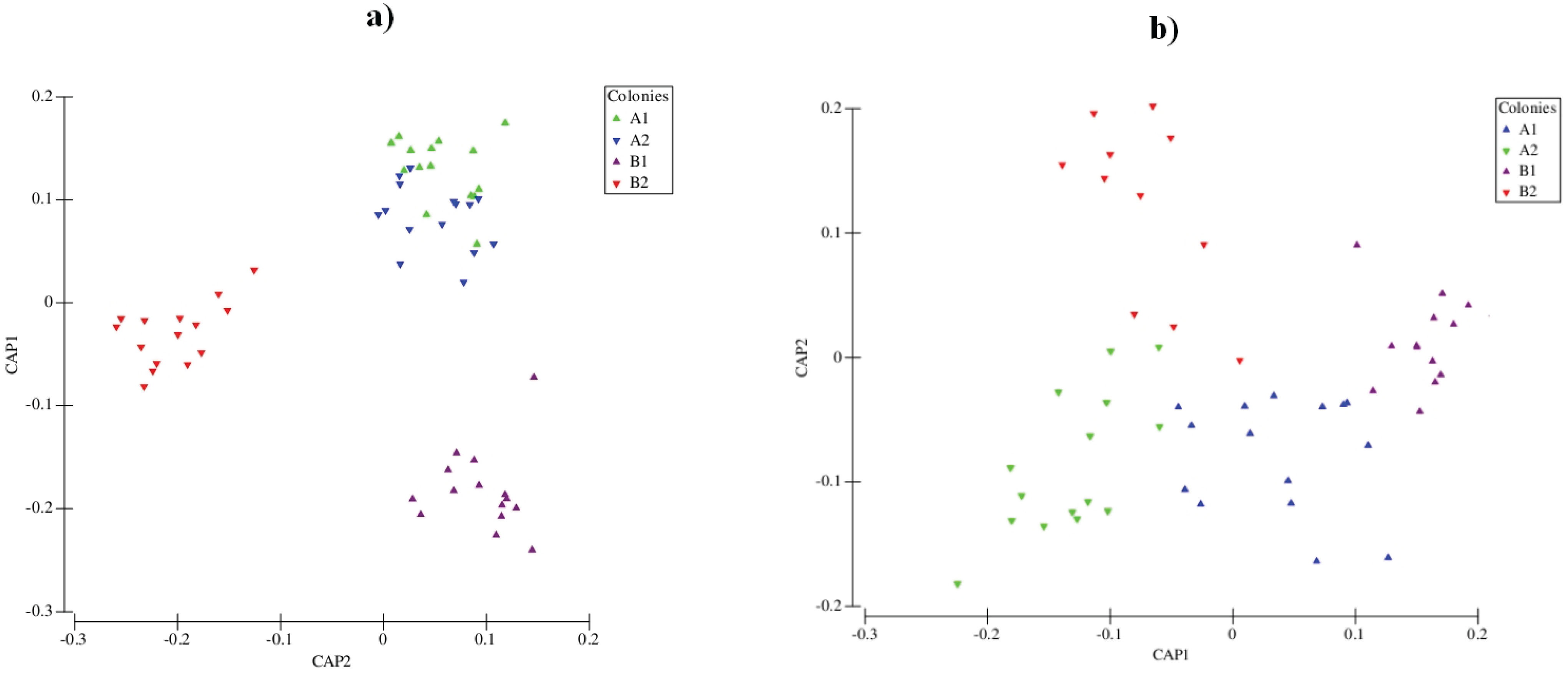
3.2. Chemical Analyses

{kind=link}
{kind=link}
{kind=link}
| Compounds | A1 | A2 | B1 | B2 | ||||
|---|---|---|---|---|---|---|---|---|
| PI | PII | PI | PII | PI | PII | PI | PII | |
| Z-9-C19 | - | - | - | - | 1.04 ± 1.83 | - | - | - |
| Z,Z-C32:2 | - | - | - | - | 0.69 ± 1.24 | - | - | - |
| n-C25 | 5 ± 1.64 | 6.32 ± 2.40 | 4.28 ± 1.65 | 5.97 ± 1.39 | 7.93 ± 2.28 | 6.25 ± 2.14 | 7.47 ± 228 | 6.32 ± 1.21 |
| n-C26 | - | 0.54 ± 0.36 | - | - | 0.59 ± 0.44 | - | 0.78 ± 0.34 | - |
| n-C27 | 10.5 ± 2.85 | 9.49 ± 2.48 | 10.33 ± 3.02 | 9.36 ± 2.20 | 17.0 ± 7.78 | 14.46 ± 2.57 | 10.92 ± 2.35 | 8.49 ± 1.93 |
| n-C29 | 1.8 ± 0.68 | 0.62 ± 0.59 | 1.84 ± 0.88 | 1.23 ± 0.63 | 4.47 ± 3.77 | 4.14 ± 2.04 | 1.69 ± 0.7 | 0.92 ± 0.27 |
| a,b,c-TriMeC30 | - | - | - | - | 0.61 ± 0.56 | - | - | - |
| 3,7,11-trimeC30 | 4.72 ± 1.17 | - | 2.87 ± 1.28 | - | 7.71 ± 2.46 | - | 3.72 ± 1.55 | - |
| n-C31 | 1.03 ± 0.23 | 4.65 ± 1.52 | 0.58 ± 0.36 | 4.17 ± 1.57 | - | 4.41 ± 0.96 | - | 3.92 ± 1.14 |
| 9-meC31 | 0.76 ± 0.35 | - | - | 0.69 ± 0.41 | 0.85 ± 0.72 | - | 0.6 ± 0.55 | - |
| 3,5-dimeC31 | 3.8 ± 1.41 | 3.22 ± 1.07 | 2.26 ± 1.09 | 2.8 ± 1.05 | 4.87 ± 1.86 | 2.48 ± 0.88 | 3.13 ± 1.5 | 3.37 ± 0.96 |
| Unknown | 8.36 ± 1.8 | 3.88 ± 0.67 | 5.67 ± 1.79 | 5.84 ± 1.74 | 8.65 ± 3.53 | 3.6 ± 0.78 | 4.92 ± 2.04 | 5.39 ± 1.57 |
| 3,7,11-trimeC32 | 13.5 ± 3.03 | 8.08 ± 1.71 | 10.15 ± 3.26 | 8.28 ± 2.74 | 14.30 ± 4.76 | 6.97 ± 1.41 | 6.94 ± 2.5 | 8.12 ± 1.85 |
| n-C33 | 1.08 ± 0.26 | - | 0.56 ± 0.39 | - | - | - | - | - |
| 4,8,12-TriMeC33 | 7.07 ± 1.44 | 3 ± 0.55 | 4.82 ± 1.56 | 4.02 ± 1.28 | 5.64 ± 2.87 | 2.55 ± 0.62 | 4.58 ± 1.91 | 5.08 ± 1.39 |
| a,b,c-TriMeC33 | 6.12 ± 1.35 | 3.14 ± 0.75 | 4.08 ± 1.43 | 3.03 ± 1.12 | 4.84 ± 2.38 | 2.52 ± 0.61 | 3.43 ± 1.33 | 4.14 ± 1.29 |
| Unknown | - | 6.07 ± 3.82 | - | 7.99 ± 4.84 | - | 2.62 ± 1.75 | - | 4.55 ± 3.31 |
| Unknown | 1.08 ± 0.29 | - | - | 0.73 ± 0.30 | 0.54 ± 0.52 | - | - | 0.61 ± 0.36 |
| a,b,c-TriMeC34 | 1.53 ± 0.57 | 0.58 ± 0.26 | 0.88 ± 0.61 | 1.1 ± 0.61 | 0.87 ± 0.66 | - | 0.84 ± 0.48 | 0.97 ± 0.25 |
| 3,7,11-trimeC34 | 5.54 ± 1.12 | 2.77 ± 0.57 | 3.68 ± 1.58 | 2.78 ± 1.36 | 4.11 ± 1.99 | 2.13 ± 0.47 | 2.78 ± 1.23 | 3.38 ± 1.01 |
| Unknown | 1.36 ± 1.38 | - | 1.34 ± 1.27 | - | - | - | - | - |
| 4,8,12-trimeC35 | 0.95 ± 0.33 | - | - | - | - | - | 0.71 ± 0.44 | 0.69 ± 0.49 |
| a,b,c-TriMeC35 | 3.49 ± 2.2 | 5.64 ± 1.94 | 6.94 ± 3.21 | 4.31 ± 1.74 | 2.99 ± 2.15 | 7.01 ± 1.07 | 6.79 ± 2.89 | 4.44 ± 1.12 |
| Unknown | 0.5 ± 0.37 | - | - | - | - | - | - | - |
| a,b,c-TriMeC37 | 13.54 ± 5.35 | 19.26 ± 3.98 | 24.33 ± 6.50 | 16.39 ± 4.69 | 8.97 ± 5.36 | 22.83 ± 3.28 | 23.7 ± 7.95 | 19.94 ± 3.10 |
| a,b,c-TriMeC39 | 7.87 ± 3.24 | 22.03 ± 3.39 | 14.88 ± 4.41 | 20.34 ± 4.42 | 3.33 ± 2.34 | 18.04 ± 2.89 | 15.6 ± 6.27 | 19.63 ± 3.28 |
| a,b,c-TriMeC40 | - | - | 0.51 ± 0.63 | - | - | - | 1.4 ± 1.66 | - |
| Phase | Colony Combinations | Euclidean Distance | Pairwise Test (p value) |
|---|---|---|---|
| I | A1 vs. A1 | 2.95 | - |
| I | A1 vs. A2 | 3.85 | 0.0001 |
| I | A1 vs. B1 | 4.48 | 0.0001 |
| I | A1 vs. B2 | 4.28 | 0.0008 |
| I | A2 vs. A2 | 2.8 | - |
| I | A2 vs. B1 | 5.25 | 0.0001 |
| I | A2 vs. B2 | 3.49 | 0.0554 |
| I | B1 vs. B1 | 3.97 | - |
| I | B1 vs. B2 | 5.21 | 0.0001 |
| I | B2 vs. B2 | 3.14 | - |
| II | A1 vs. A1 | 2.58 | 0.0608 |
| II | A1 vs. A2 | 2.94 | 0.0001 |
| II | A1 vs. B1 | 2.99 | 0.0033 |
| II | A1 vs. B2 | 2.69 | - |
| II | A2 vs. A2 | 2.61 | - |
| II | A2 vs. B1 | 3.37 | 0.0001 |
| II | A2 vs. B2 | 2.79 | 0.0439 |
| II | B1 vs. B1 | 1.83 | - |
| II | B1 vs. B2 | 3.05 | 0.0001 |
| II | B2 vs. B2 | 2.16 | - |

4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hamilton, W.D. The genetical evolution of social behaviour. J. Theor. Biol. 1964, 7, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Vander Meer, R.K.; Morel, L. Nestmate recognition in ants. In Pheromone Communication in Social Insects: Ants, Wasps, Bees, and Termites; Vander Meer, R.K., Breed, M.D., Espelie, K.E., Winston, M.L., Eds.; Westview Press: Oxford, UK, 1998; pp. 79–103. [Google Scholar]
- Sherman, P.W.; Reeve, H.K.; Pfeenig, D.W. Recognition systems. In Behavioural Ecology: An Evolutionary Approach; Krebs, J.R., Davies, N.B., Eds.; Blackwell Science: Oxford, UK, 1997; pp. 69–96. [Google Scholar]
- Starks, P.T. Recognition systems: From components to conservation. Ann. Zool. Fenni. 2004, 41, 689–690. [Google Scholar]
- Tibbetts, E.A. Visual signals of individual identity in the paper wasp Polistes fuscatus. Proc. R. Soc. Lond. 2002, 269, 1423–1428. [Google Scholar] [CrossRef]
- D’Ettorre, P.; Heinze, J. Individual recognition in ant queens. Curr. Biol. 2005, 15, 2170–2174. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, M.J.; Tibbetts, E.A. Specialized face learning is associated with individual recognition in paper wasps. Science 2012, 334, 1272–1275. [Google Scholar] [CrossRef]
- Tannure-Nascimento, I.C.; Nascimento, F.S.; Turatti, I.C.; Lopes, N.P.; Trigo, J.R.; Zucchi, R. Colony membership is reflected by variations in cuticular hydrocarbon profile in a Neotropical paper wasp, Polistes satan (Hymenoptera, Vespidae). Genet. Mol. Res. 2007, 6, 390–396. [Google Scholar] [PubMed]
- Caliman-Ferreira, M.J.; Nascimento, F.S.; Turatti, I.C.; Mateus, S.; Lopes, N.P.; Zucchi, R. The cuticular hydrocarbons profiles in the stingless bee Melipona marginata reflect task-related differences. J. Insect Physiol. 2010, 56, 800–804. [Google Scholar] [CrossRef] [PubMed]
- Crozier, R.H.; Pamilo, P. Evolution of Social Insect Colonies: Sex Allocation and Kin Selection; Oxford University Press: Oxford, UK, 1996. [Google Scholar]
- Ozaki, M.; Hefetz, A. Neural mechanisms and information processing in recognition systems. Insects 2014, 5, 722–741. [Google Scholar] [CrossRef]
- Breed, M.D.; Bennett, B. Kin recognition in highly eusocial insects. In Kin Recognition in Animals; Fletcher, D.J.C., Michener, C.D., Eds.; John Wiley and Sons: New York, NY, USA, 1987; pp. 243–285. [Google Scholar]
- Hernandéz, J.V.; López, H.; Jaffe, K. Nestmate recognitions signals of the leaf-cutter ant Atta laevigata. J. Insect Physiol. 2002, 48, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Hernandéz, J.; Goitia, V.W.; Osio, A.; Cabrera, H.L.; Sainz, C.; Jaffe, K. Leaf-cutter ant species (Hymenoptera: Atta) differ in types of cues used to differentiate between self and others. Anim. Behav. 2006, 71, 945–952. [Google Scholar] [CrossRef]
- Blomquist, G.J.; Bagnéres, A.G. Structure and analysis of insect hydrocarbons. In Insect Hydrocarbons; Blomquist, G.J., Ed.; Cambridge University Press: Cambridge, UK, 2010; pp. 19–34. [Google Scholar]
- Whitehouse, M.; Jaffe, K. Nestmate recognition in the leaf-cutting ant Atta laevigata. Insectes Sociaux 1995, 42, 157–166. [Google Scholar] [CrossRef]
- Nielsen, J.; Boomsma, J.J.; Oldham, N.J.; Petersen, H.C.; Morgan, E.D. Colony-level and season-specific variation in cuticular hydrocarbon profiles of individual workers in the ant Formica truncorum. Insectes Sociaux 1999, 46, 58–65. [Google Scholar] [CrossRef]
- Buczkowski, G.; Kumar, R.; Suib, S.L.; Silverman, J. Diet-related modifications of hydrocarbon profiles of the argentine ant, Linepithema. humile, dimishes intercolony aggression. J. Chem. Ecol. 2005, 31, 829–843. [Google Scholar]
- Ichinose, K.; Boulay, R.; Cerdá, X.; Lenoir, A. Influence of queen and diet on nestmate recognition and cuticular hydrocarbon differentiation in a fission-dispersing ant, Aphaenogaster senilis. Zoolog. Sci. 2009, 26, 681–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richard, F.J.; Hunt, J.H. Intracolony chemical communication in social insects. Insect. Soc. 2013, 60, 275–291. [Google Scholar] [CrossRef]
- Siqueira, C.G.; Bacci, M.; Pagnocca, F.C.; Bueno, O.C.; Hebling, M.J.A. Metabolism of plant polysaccharides by Leucoagaricus. gongylophorus, the symbiotic fungus of leaf-cutting ant Atta sexdens L. Appl. Environ. Microbiol. 1998, 64, 4820–4822. [Google Scholar]
- Silva, A.; Bacci, M., Jr.; Siqueira, C.G.; Bueno, F.C.; Pagnocca, F.C.; Hebling, M.J.A. Survival of Atta sexdens workers on different food sources. J. Insect Physiol. 2003, 49, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.J.; Vitikainen, E.; Helanterä, H.; Drijfhout, F.P. Chemical basis of nest-mate discrimination in the ant Formica exsecta. Proc. R. Soc. Lond. 2008, 275, 1271–1278. [Google Scholar] [CrossRef]
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.S.S. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2008, 24, 127–135. [Google Scholar] [CrossRef]
- Carlson, D.A. Dimethyl disulfide derivates of long chain alkenes, alkadienes, and alkatrienes for gas chromatography/mass spectrometry. Anal. Chem. 1989, 61, 1564–1571. [Google Scholar] [CrossRef]
- Lambardi, D.; Chegia, B.; Turilazzi, S.; Boomsma, J.J. Diet-induced aggression among colonies of the leafcutter ant Acromyrmex. echinatior Forel (Hymenoptera Formicidae). Redia 2004, 21, 219–221. [Google Scholar]
- Larsen, J.; Fouks, B.; Bos, N.; D’Ettorre, P.; Nehring, V. Variation in nestmate recognition ability among polymorphic leaf-cutting ant workers. J. Insect Physiol. 2014, 70, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Richard, F.J.; Heftz, A.; Christides, J.P.; Errard, C. Food influence on colonial recognition and chemical signature between nestmates in the fungus-growing ant Acromyrmex subterraneus subterraneus. J. Chem. Ecol. 2004, 14, 9–16. [Google Scholar]
- Jaffé, K. Evolution of territoriality and nestmate recognition in ants. Exp. Suppl. 1987, 54, 295–311. [Google Scholar]
- Jaffé, K.; Marcuse, M. Nestmate recognition and territorial behavior in the ant Odontomachus bauri Emery (Formicidae: Ponerinae). Insects Sociaux 1983, 30, 466–481. [Google Scholar] [CrossRef]
- Jaffé, K.; Sanchez, C. On the nestmate recognition system and territorial marking behaviour in the ant Camponotus rufipes. Insects Sociaux 1984, 31, 302–315. [Google Scholar] [CrossRef]
- Jaffé, K. Nestmate recognition and territoral marking in Sonelopsis geminata and in some Attini. In Fire Ants and Leaf-cutting Ants: Biology and Management; Lofgren, C.S., Vander Meer, R.K., Eds.; Westview Press: Boulder, CO, USA, 1986; pp. 211–222. [Google Scholar]
- Lahav, S.; Soroker, V.; Hefetz, A. Direct behavioral evidence for hydrocarbons as ant recognition discriminators. Naturwissenschaften 1999, 86, 246–249. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valadares, L.; Nascimento, D.; Nascimento, F.S. Foliar Substrate Affects Cuticular Hydrocarbon Profiles and Intraspecific Aggression in the Leafcutter Ant Atta sexdens. Insects 2015, 6, 141-151. https://doi.org/10.3390/insects6010141
Valadares L, Nascimento D, Nascimento FS. Foliar Substrate Affects Cuticular Hydrocarbon Profiles and Intraspecific Aggression in the Leafcutter Ant Atta sexdens. Insects. 2015; 6(1):141-151. https://doi.org/10.3390/insects6010141
Chicago/Turabian StyleValadares, Lohan, Daniela Nascimento, and Fabio S. Nascimento. 2015. "Foliar Substrate Affects Cuticular Hydrocarbon Profiles and Intraspecific Aggression in the Leafcutter Ant Atta sexdens" Insects 6, no. 1: 141-151. https://doi.org/10.3390/insects6010141
APA StyleValadares, L., Nascimento, D., & Nascimento, F. S. (2015). Foliar Substrate Affects Cuticular Hydrocarbon Profiles and Intraspecific Aggression in the Leafcutter Ant Atta sexdens. Insects, 6(1), 141-151. https://doi.org/10.3390/insects6010141