Exceptional Use of Sex Pheromones by Parasitoids of the Genus Cotesia: Males Are Strongly Attracted to Virgin Females, but Are No Longer Attracted to or Even Repelled by Mated Females

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
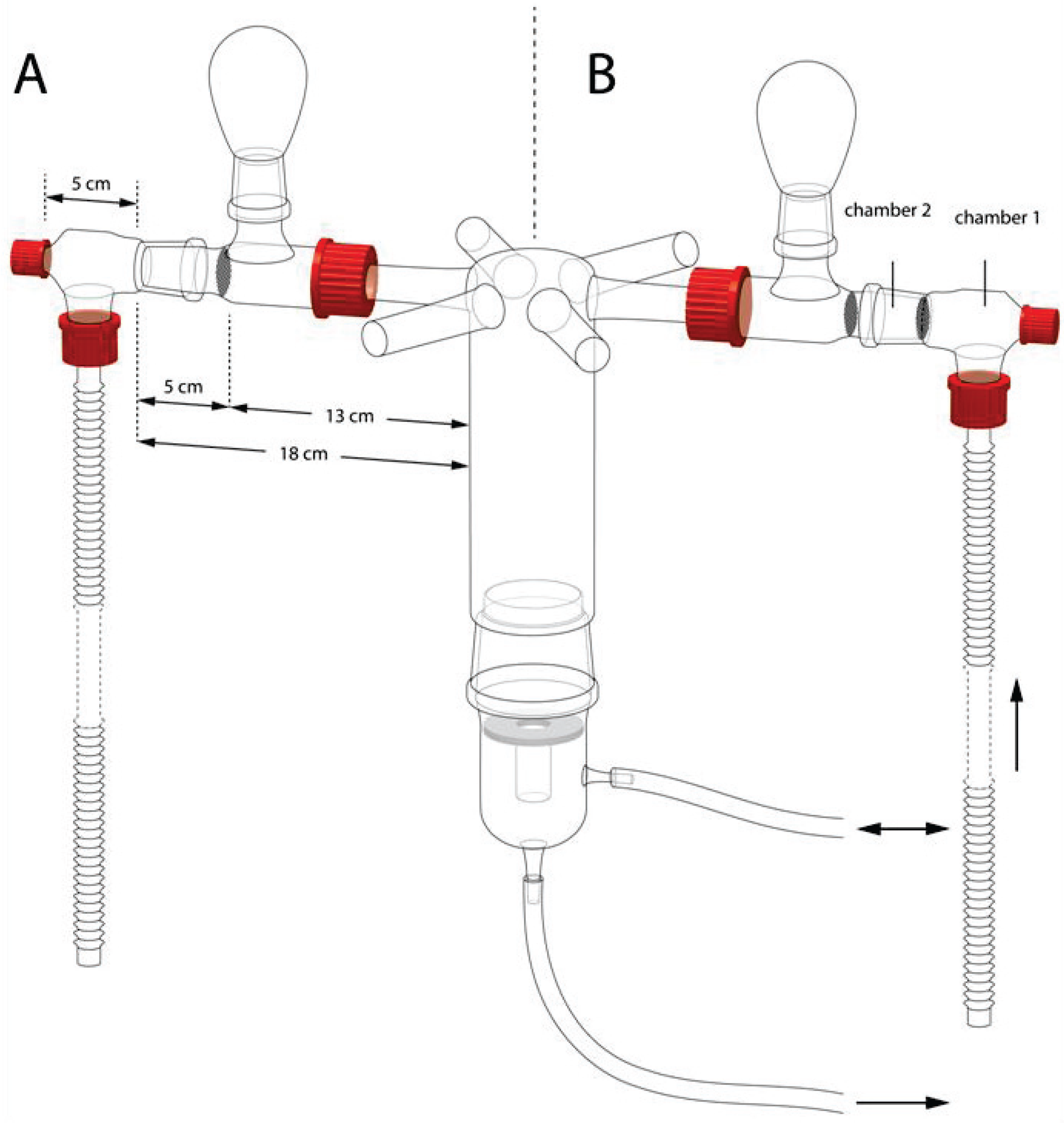
2.1. Wasps
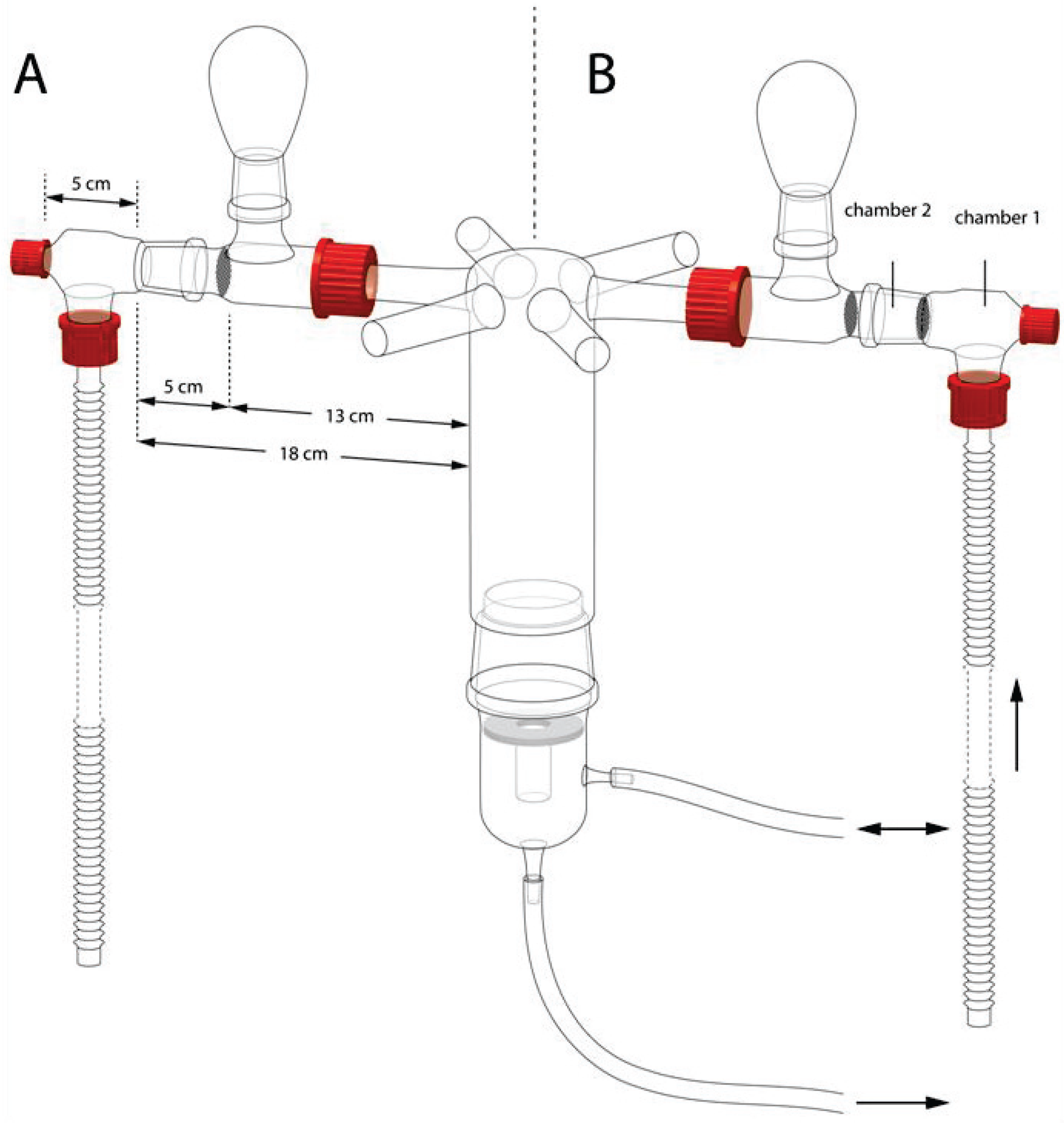
2.2. Bioassay
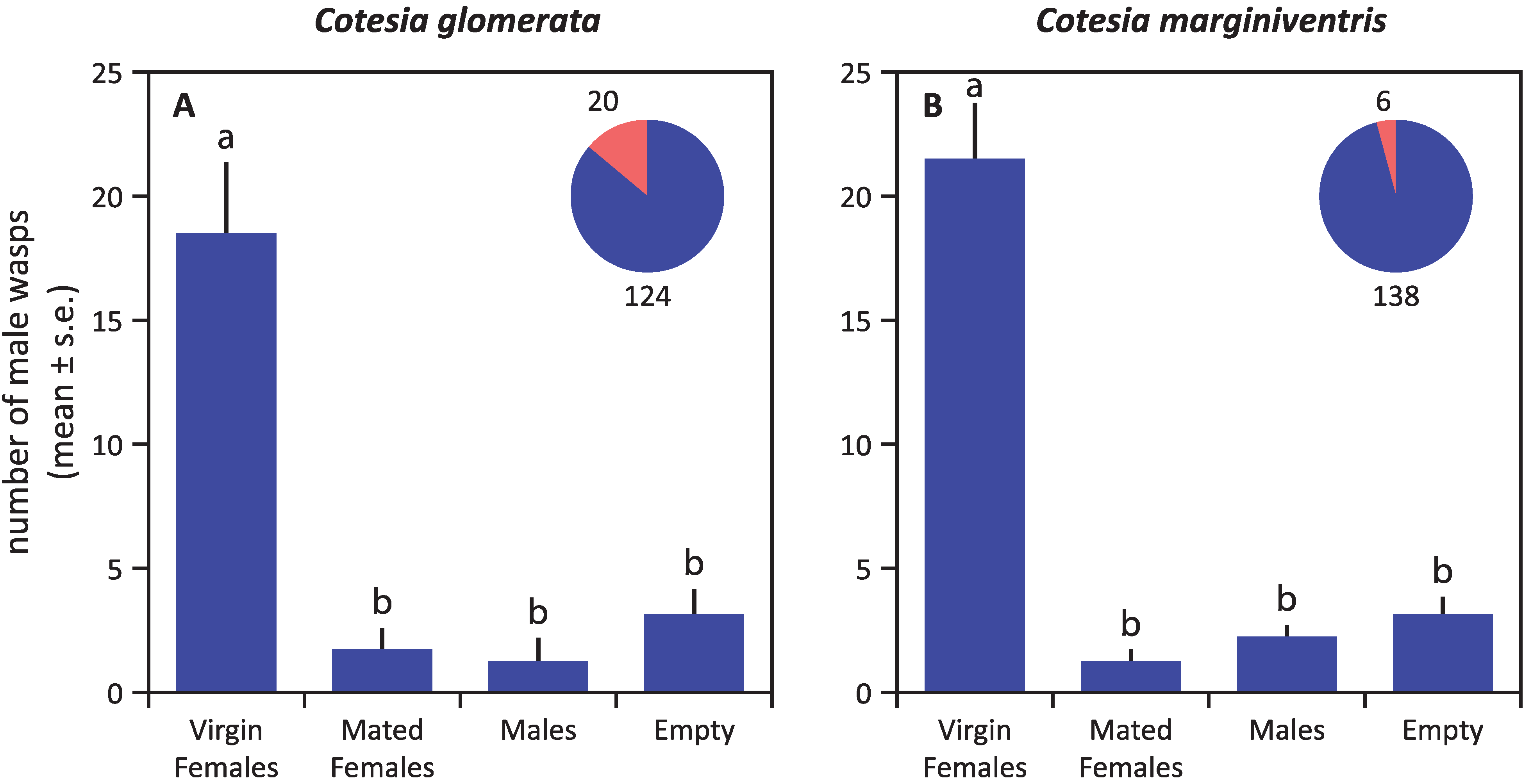
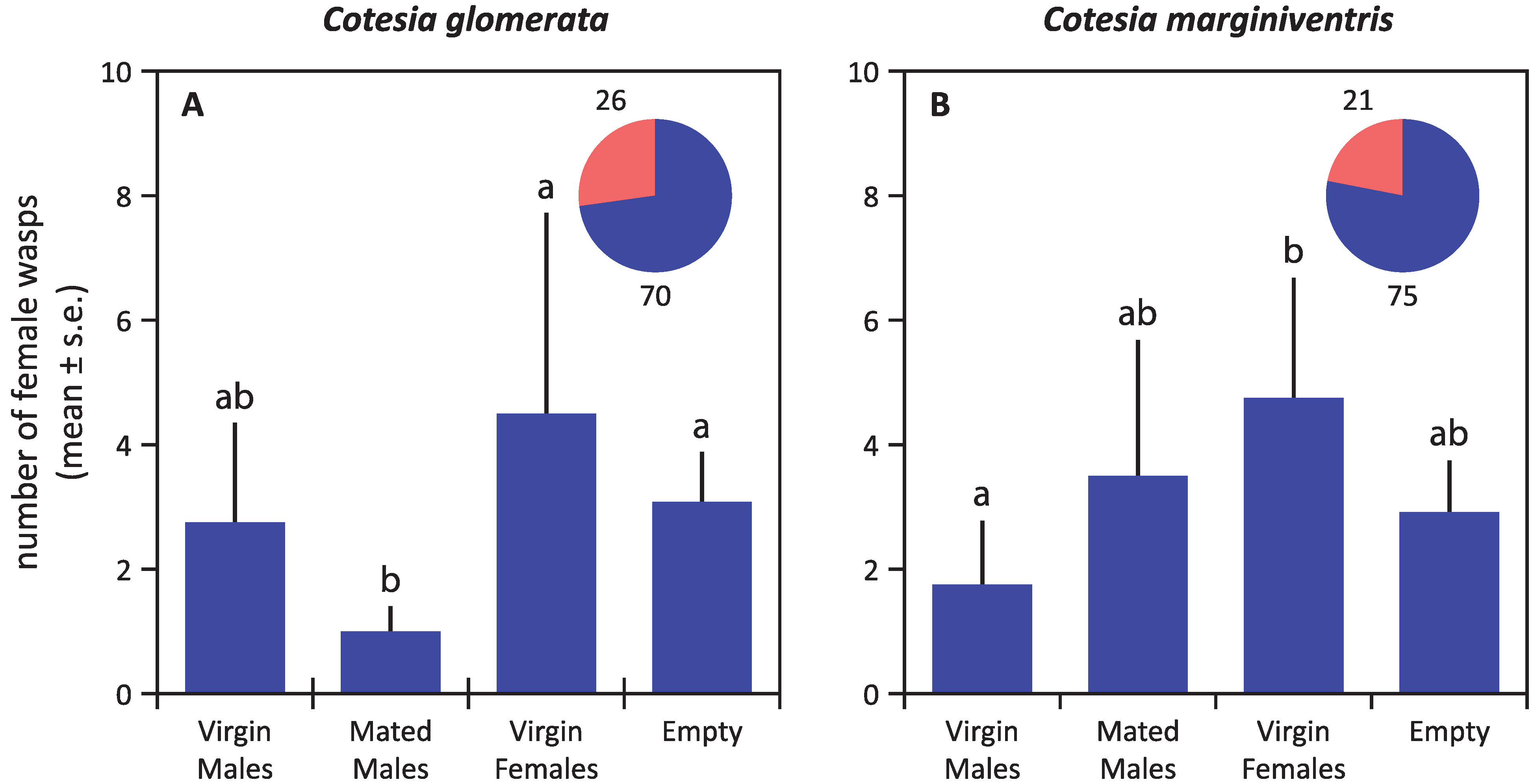
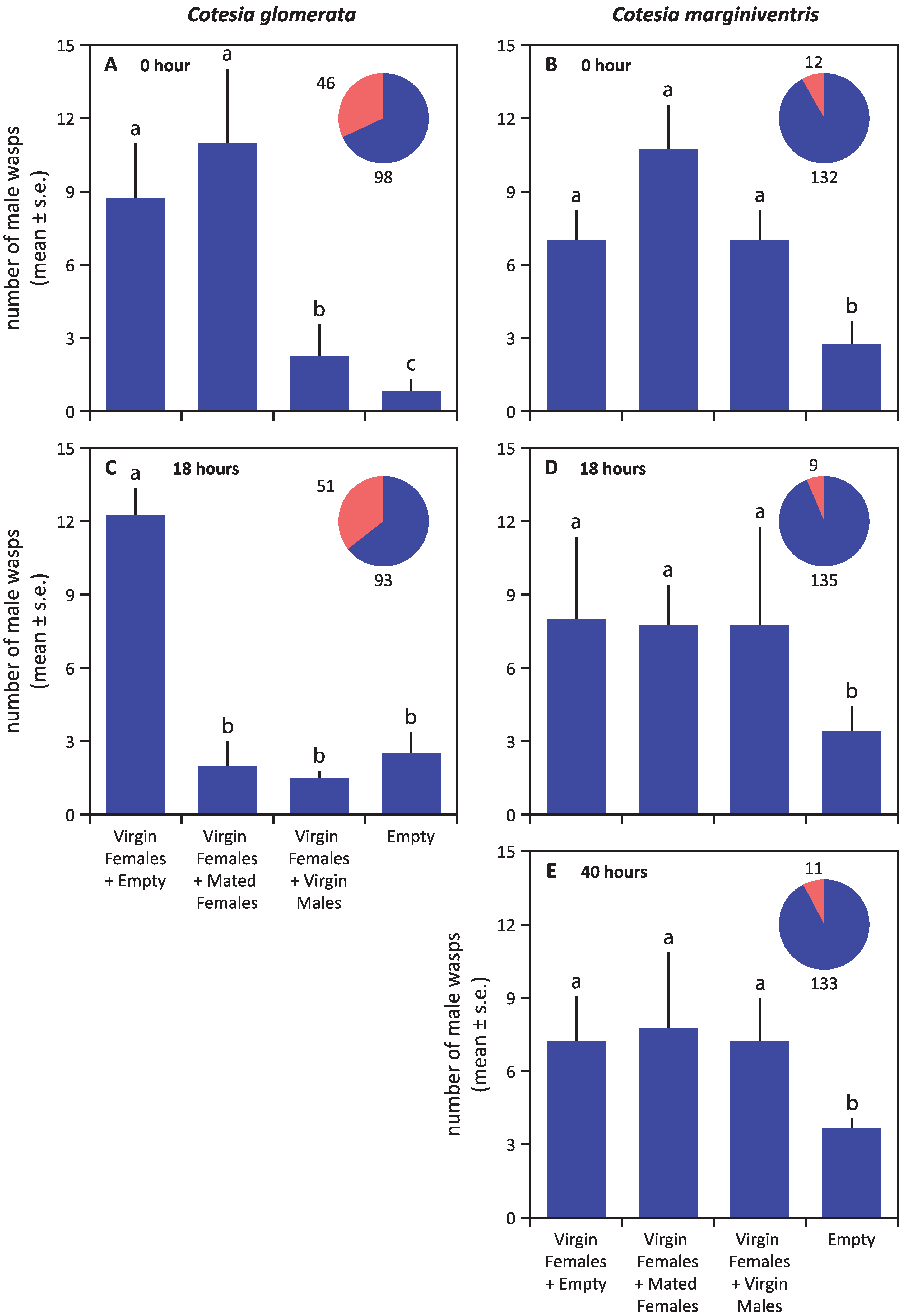
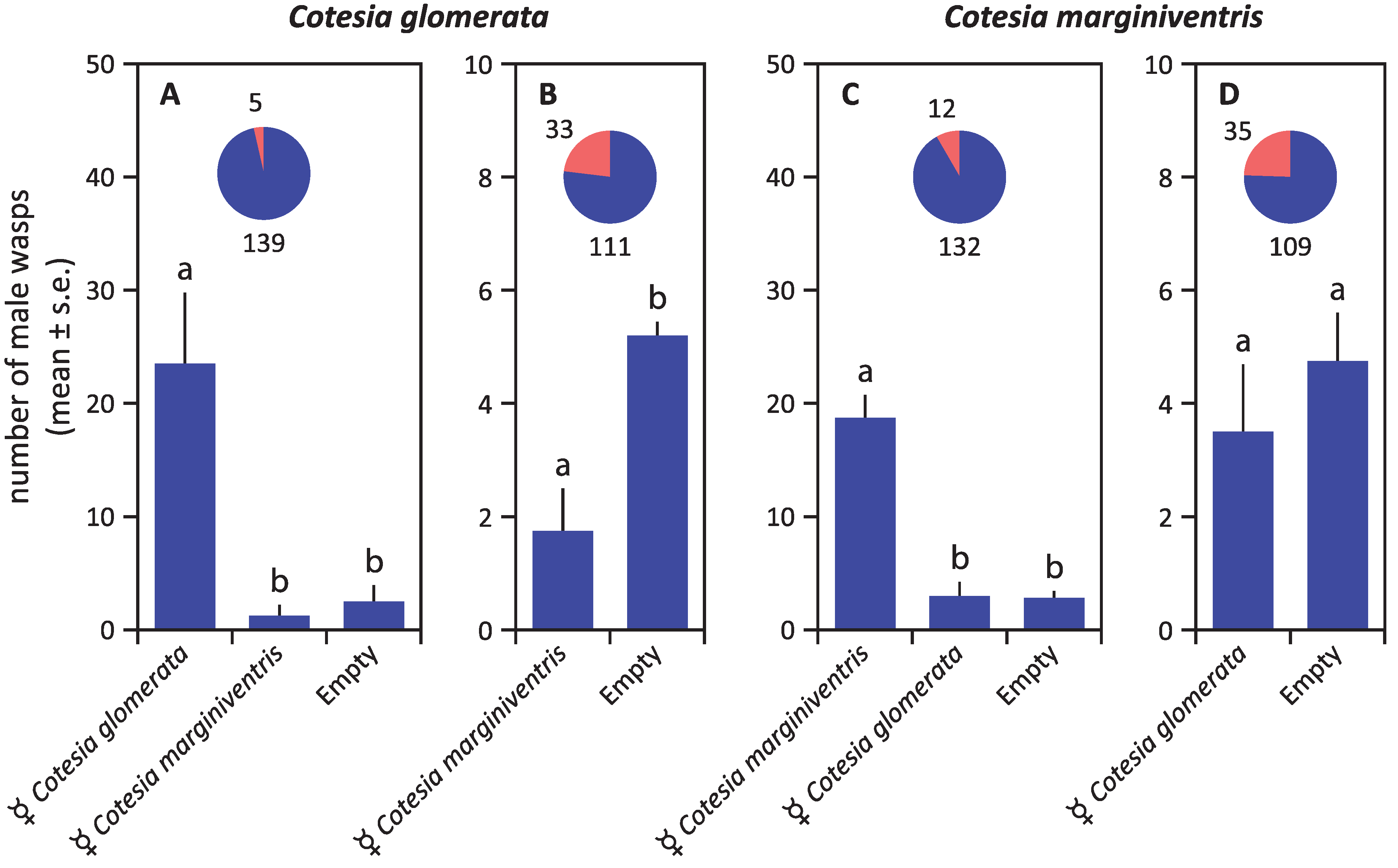
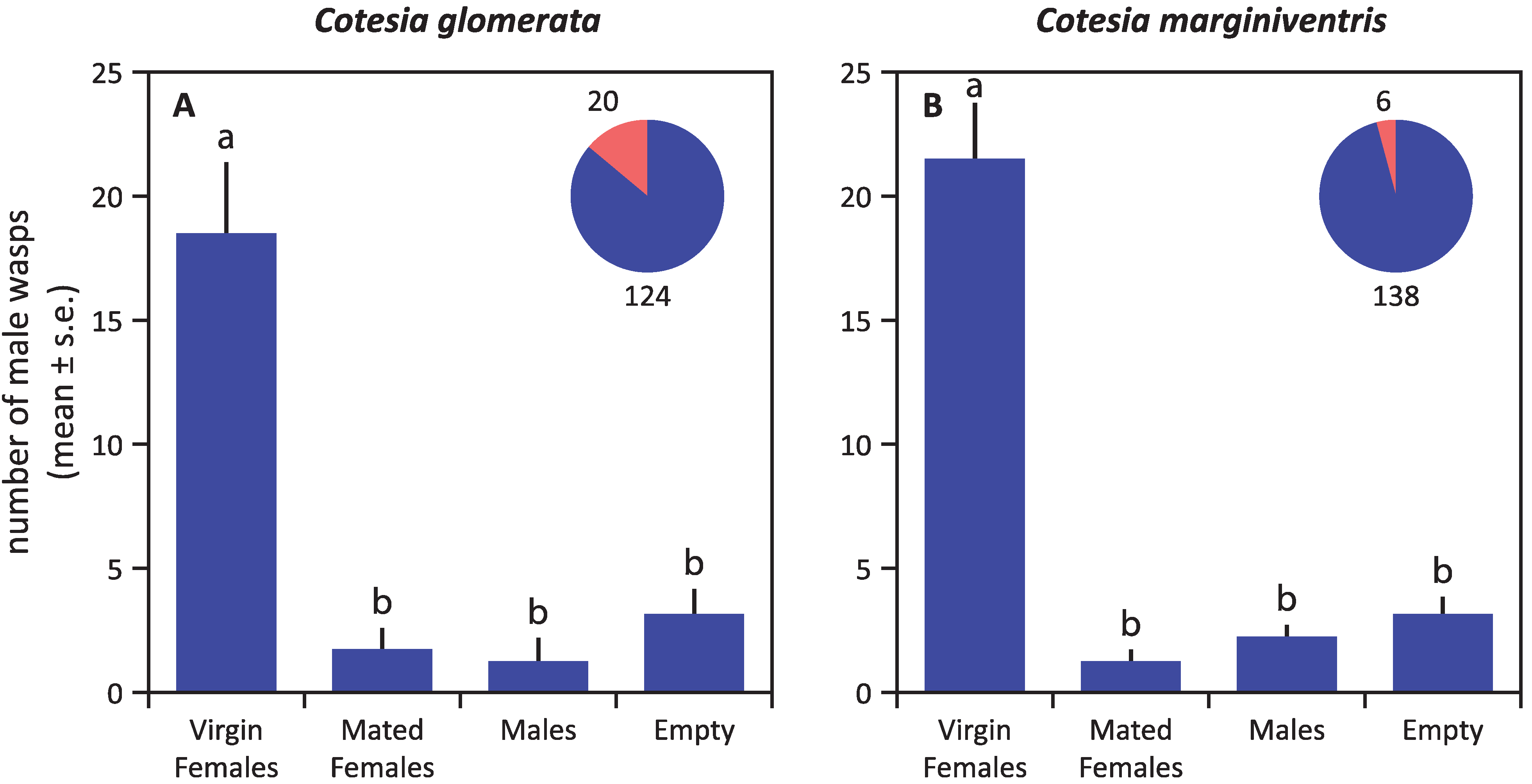
3. Results
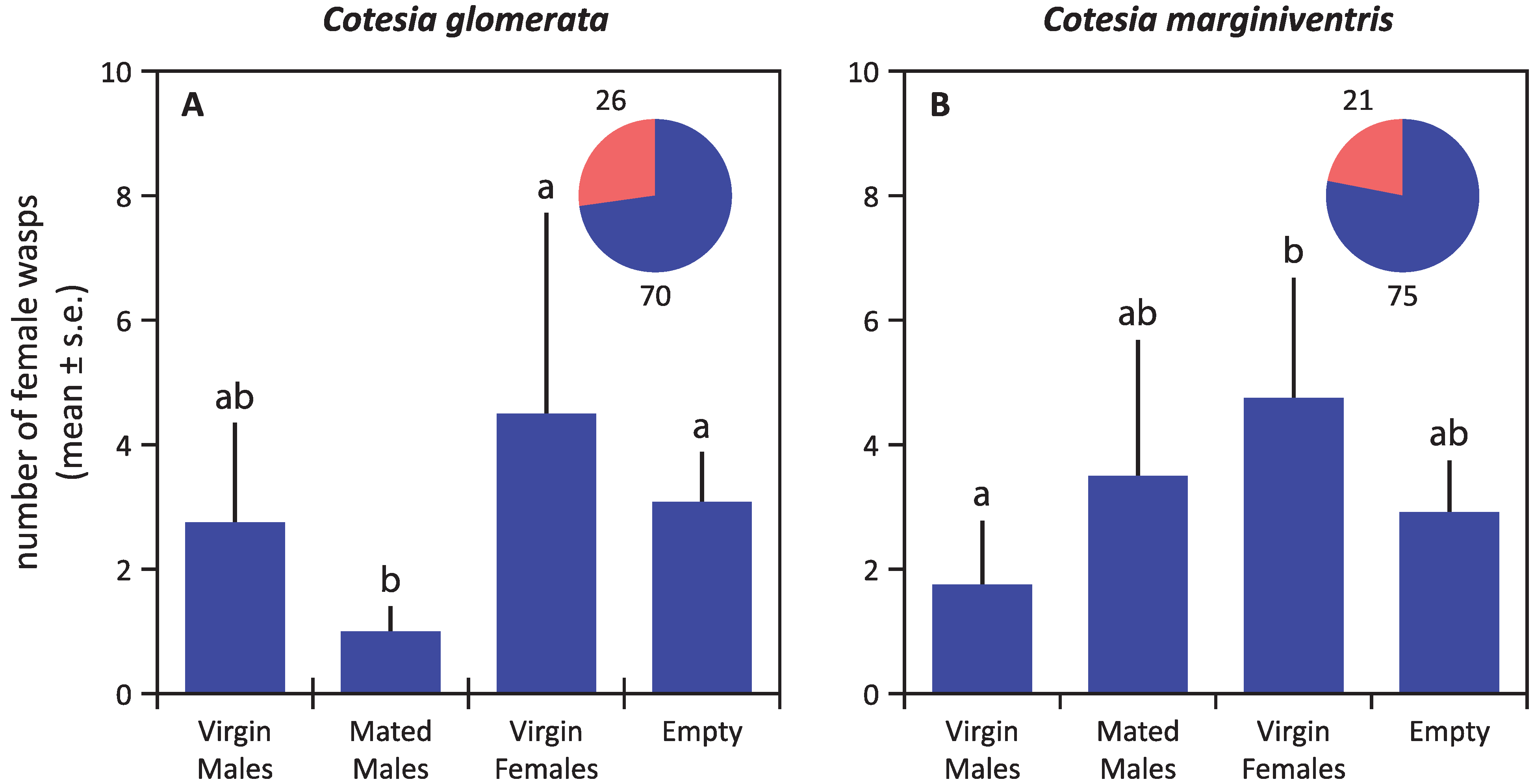
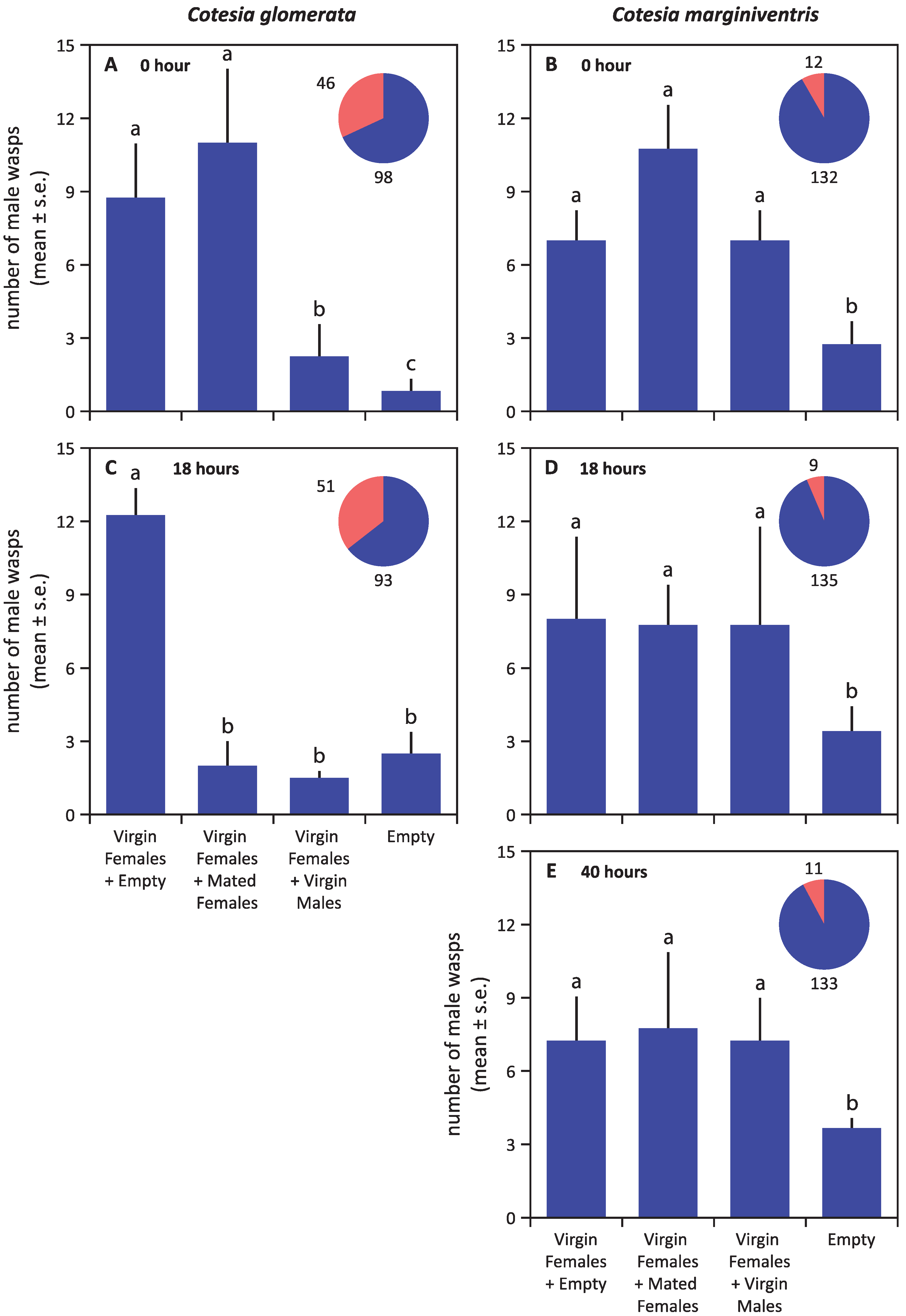
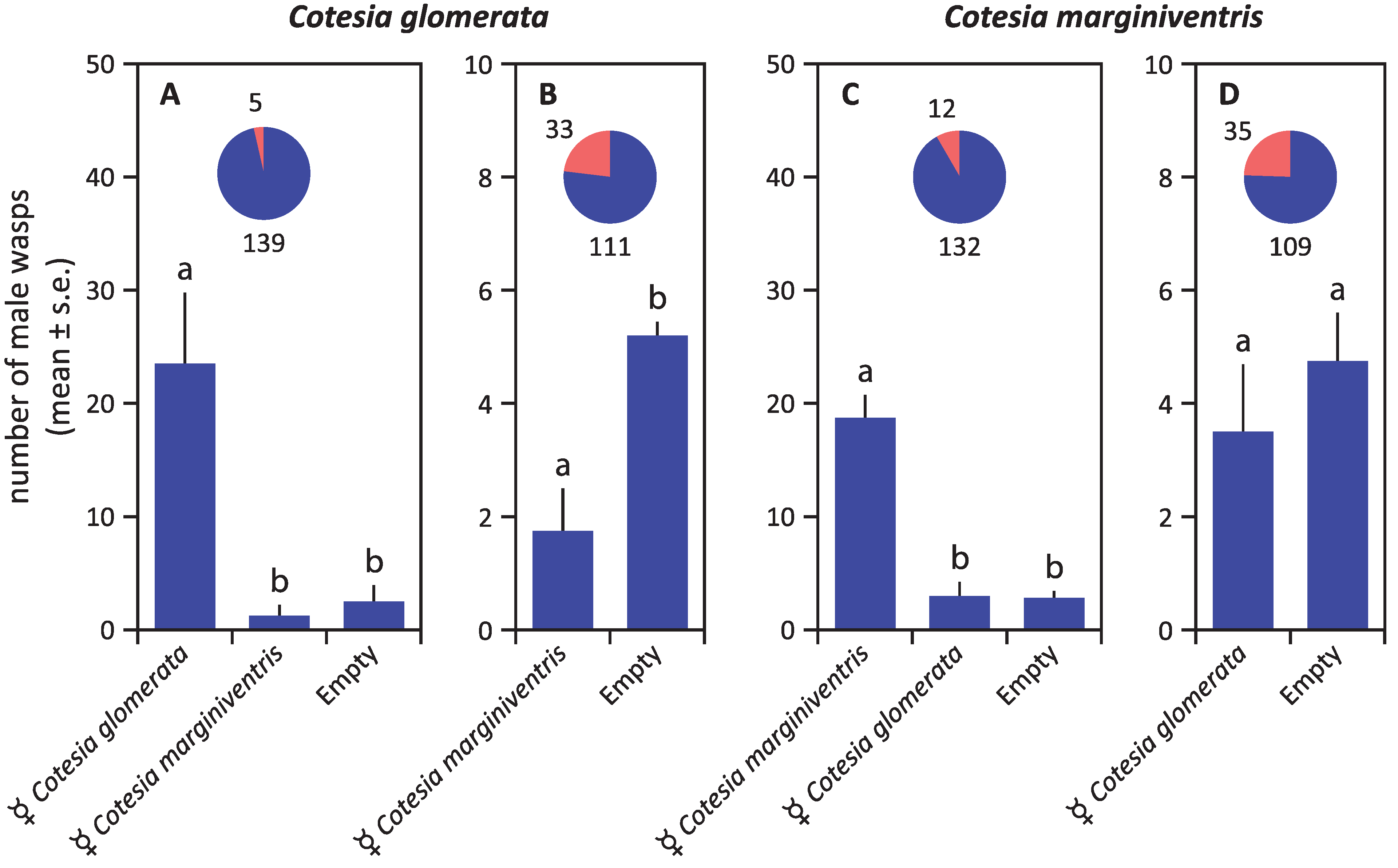
4. Discussion
4.1. Mate Finding Strategy of Cotesia
4.2. Possible Differences in Mating Behaviors between Gregarious and Solitary Parasitoids
4.3. Species Specificity of Sex Pheromones
5. Conclusion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gu, H.N.; Dorn, S. Mating system and sex allocation in the gregarious parasitoid Cotesia glomerata. Anim. Behav. 2003, 66, 259–264. [Google Scholar] [CrossRef]
- Fauvergue, X.; Lo Genco, A.; Lo Pinto, M. Virgins in the wild: Mating status affects the behavior of a parasitoid foraging in the field. Oecologia 2008, 156, 913–920. [Google Scholar] [CrossRef]
- Steiner, S.; Ruther, J. How important is sex for females of a haplodiploid species under local mate competition? Behav. Ecol. 2009, 20, 570–574. [Google Scholar]
- Kant, R.; Minor, M.; Trewick, S. Mating or ovipositing? A crucial decision in the life history of the cabbage aphid parasitoid Diaeretiella rapae (M'Intosh). Ecol. Entomol. 2012, 37, 169–174. [Google Scholar] [CrossRef]
- McNeil, J.N.; Brodeur, J. Pheromone-mediated mating in the aphid parasitoid, Aphidius nigripes (Hymenoptera, Aphididae). J. Chem. Ecol. 1995, 21, 959–972. [Google Scholar] [CrossRef]
- Danci, A.; Gries, R.; Schaefer, P.W.; Gries, G. Evidence for four-component close-range sex pheromone in the parasitic wasp Glyptapanteles flavicoxis. J. Chem. Ecol. 2006, 32, 1539–1554. [Google Scholar] [CrossRef]
- McClure, M.; Whistlecraft, J.; McNeil, J.N. Courtship behavior in relation to the female sex pheromone in the parasitoid, Aphidius ervi (Hymenoptera : Braconidae). J. Chem. Ecol. 2007, 33, 1946–1959. [Google Scholar] [CrossRef]
- Ruther, J.; Stahl, L.M.; Steiner, S.; Garbe, L.A.; Tolasch, T. A male sex pheromone in a parasitic wasp and control of the behavioral response by the female’s mating status. J. Exp. Biol. 2007, 210, 2163–2169. [Google Scholar] [CrossRef]
- Quicke, D.L. Parasitic Wasps; Chapman & Hall Ltd: London, UK, 1997. [Google Scholar]
- Ruther, J. Novel insights into pheromone-mediated communication in parasitic hymenopterans. In Chemical Ecology of Insect Parasitoids; John Wiley & Sons, Ltd: Chichester, UK, 2013; pp. 112–144. [Google Scholar]
- Jang, E.B. Effects of mating and accessory gland injections on olfactory-mediated behavior in the female mediterranean fruit-fly, Ceratitis capitata. J. Insect Physiol. 1995, 41, 705–710. [Google Scholar] [CrossRef]
- Kugimiya, S.; Shimoda, T.; Wajnberg, E.; Uefune, M.; Takabayashi, J. Host-searching responses to herbivory-associated chemical information and patch use depend on mating status of female solitary parasitoid wasps. Ecol. Entomol. 2010, 35, 279–286. [Google Scholar] [CrossRef]
- Si, S.; Xu, S.; Du, J. Pheromonostatic activity of male accessory gland factors in female Helicoverpa assulta. Acta Entomologica Sinica 2000, 43, 120–126. [Google Scholar]
- Werren, J.H. Sex ratio adaptations to local mate competition in a parasitic wasp. Science 1980, 208, 1157–1159. [Google Scholar]
- Matthews, R.W.; Yukawa, J.; Gonzalez, J.M. Sex pheromones in male Melittobia parasitic wasps: Female response to conspecific and congeneric males of 3 species. J. Ethol. 1985, 3, 59–62. [Google Scholar] [CrossRef]
- Kimani, S.W.; Overholt, W.A. Biosystematics of the Cotesia flavipes complex (Hymenoptera: Braconidae): Interspecific hybridization, sex pheromone and mating behavior studies. B. Entomol. Res. 1995, 85, 379–386. [Google Scholar] [CrossRef]
- Niehuis, O.; Buellesbach, J.; Gibson, J.D.; Pothmann, D.; Hanner, C.; Mutti, N.S.; Judson, A.K.; Gadau, J.; Ruther, J.; Schmitt, T. Behavioural and genetic analyses of Nasonia shed light on the evolution of sex pheromones. Nature 2013, 494, 345–348. [Google Scholar] [CrossRef]
- Michel-Salzat, A.; Whitfield, J.B. Preliminary evolutionary relationships within the parasitoid wasp genus Cotesia (Hymenoptera: Braconidae: Microgastrinae): Combined analysis of four genes. Syst. Entomol. 2004, 29, 371–382. [Google Scholar] [CrossRef]
- De Freitas, M.D.T.; Mendonca, A.D.; do Nascimento, R.R.; Sant'ana, A.E.G. Behavioural evidence for a female sex pheromone in Cotesia flavipes (Hymenoptera : Braconidae). Physiol. Entomol. 2004, 29, 183–187. [Google Scholar]
- Field, S.A.; Keller, M.A. Courtship and intersexual signaling in the parasitic wasp Cotesia rubecula (Hymenoptera: Braconidae). J. Insect Behav. 1993, 6, 737–750. [Google Scholar] [CrossRef]
- Tagawa, J. Localization and histology of the female sex pheromone-producing gland in the parasitic wasp, Apanteles glomeratus. J. Insect Physiol. 1977, 23, 49–56. [Google Scholar] [CrossRef]
- Tagawa, J.; Kitano, H. Mating behavior of the braconid wasp, Apanteles glomeratus L (Hymenoptera: Braconidae) in the field. Appl. Entomol. Zool. 1981, 16, 345–350. [Google Scholar]
- Tamò, C.; Roelfstra, L.L.; Guillaume, S.; Turlings, T.C.J. Odour-mediated long-range avoidance of interspecific competition by a solitary endoparasitoid: A time-saving foraging strategy. J. Anim. Ecol. 2006, 75, 1091–1099. [Google Scholar] [CrossRef]
- Turlings, T.C.J.; Davison, A.C.; Tamò, C. A six-arm olfactometer permitting simultaneous observation of insect attraction and odour trapping. Physiol. Entomol. 2004, 29, 45–55. [Google Scholar] [CrossRef]
- Ricard, I.; Davison, A.C. Statistical inference for olfactometer data. J. Roy Stat. Soc. C Appl. Stat. 2007, 56, 479–492. [Google Scholar] [CrossRef]
- Xu, H.; Institute of Biology, University of Neuchâtel, CH-2000 Neuchâtel, Switzerland. Unpublished work. 2014.
- Kawaguchi, M.; Tanaka, T. Biological characteristics of a larval endoparasitoid, Cotesia plutellae (Hymenoptera : Braconidae): Host stage preference, subsequent sex ratio of progeny and mate location of males. Appl. Entomol. Zool. 1999, 34, 213–221. [Google Scholar]
- Mcauslane, H.; Vinson, S.; Williams, H. Influence of host plant on mate location by the parasitoid Campoletis sonorensis (Hymenoptera: Ichneumonidae). Environ. Entomol. 1990, 19, 26–31. [Google Scholar]
- Yu, H.L.; Zhang, Y.J.; Wyckhuys, K.A.G.; Wu, K.M.; Gao, X.W.; Guo, Y.Y. Electrophysiological and behavioral responses of Microplitis mediator (Hymenoptera: Braconidae) to caterpillar-induced volatiles from cotton. Environ. Entomol. 2010, 39, 600–609. [Google Scholar] [CrossRef]
- Tagawa, J.; Hidaka, T. Mating behavior of the braconid wasp, Apanteles glomeratus L (Hymenoptera: Braconidae): Mating sequence and the factor for correct orientation of male to female. Appl. Entomol. Zool. 1982, 17, 32–39. [Google Scholar]
- Nyabuga, F.N.; Volkl, W.; Schworer, U.; Weisser, W.W.; Mackauer, M. Mating strategies in solitary aphid parasitoids: Effect of patch residence time and ant attendance. J. Insect Behav. 2012, 25, 80–95. [Google Scholar] [CrossRef]
- Carroll, M.J.; Schmelz, E.A.; Teal, P.E.A. The attraction of Spodoptera frugiperda neonates to cowpea seedlings is mediated by volatiles induced by conspecific herbivory and the elicitor inceptin. J. Chem. Ecol. 2008, 34, 291–300. [Google Scholar] [CrossRef]
- Degen, T.; Bakalovic, N.; Bergvinson, D.; Turlings, T.C.J. Differential performance and parasitism of caterpillars on maize inbred lines with distinctly different herbivore-induced volatile emissions. PLoS One 2012, 7, e47589. [Google Scholar]
- Yang, G.; Wiseman, B.R.; Espelie, K.E. Movement of neonate fall armyworm (Lepidoptera: Noctuidae) larvae on resistant and susceptible genotypes of corn. Environ. Entomol. 1993, 22, 547–553. [Google Scholar]
- Delisle, J.; Simard, J. Factors involved in the post-copulatory neural inhibition of pheromone production in Choristoneura fumiferana and C. rosaceana females. J. Insect Physiol. 2002, 48, 181–188. [Google Scholar] [CrossRef]
- Marcotte, M.; Delisle, J.; McNeil, J.N. Impact of male mating history on the temporal sperm dynamics of Choristoneura rosaceana and C. fumiferana females. J. Insect Physiol. 2005, 51, 537–544. [Google Scholar] [CrossRef]
- Kingan, T.G.; Thomas-Laemont, P.A.; Raina, A.K. Male accessory gland factors elicit change from virgin to mated behavior in the female corn earworm moth Helicoverpa zea. J. Exp. Biol. 1993, 183, 61–76. [Google Scholar]
- Gilbert, L.E. Postmating female odor in Heliconius butterflies: A male-contributed antiaphrodisiac? Science 1976, 193, 419–420. [Google Scholar]
- Andersson, J.; Borg-Karlson, A.K.; Wiklund, C. Sexual cooperation and conflict in butterflies: A male-transferred anti-aphrodisiac reduces harassment of recently mated females. Proc. Biol. Sci. 2000, 267, 1271–1275. [Google Scholar] [CrossRef]
- Fatouros, N.E.; Huigens, M.E.; van Loon, J.J.A.; Dicke, M.; Hilker, M. Chemical communication: Butterfly anti-aphrodisiac lures parasitic wasps. Nature 2005, 433, 704–704. [Google Scholar] [CrossRef]
- Huigens, M.E.; Pashalidou, F.G.; Qian, M.H.; Bukovinszky, T.; Smid, H.M.; van Loon, J.J.A.; Dicke, M.; Fatouros, N.E. Hitch-hiking parasitic wasp learns to exploit butterfly antiaphrodisiac. Proc. Natl. Acad. Sci. USA 2009, 106, 820–825. [Google Scholar] [CrossRef]
- Scott, D. Sexual mimicry regulates the attractiveness of mated Drosophila melanogaster females. Proc. Natl. Acad. Sci. USA 1986, 83, 8429–8433. [Google Scholar] [CrossRef]
- Scott, D.; Jackson, L.L. Interstrain comparison of male-predominant antiaphrodisiacs in Drosophila melanogaster. J. Insect Physiol. 1988, 34, 863–871. [Google Scholar] [CrossRef]
- Xu, H.; Institute of Biology, University of Neuchâtel, CH-2000 Neuchâtel, Switzerland. Personal observation, 2014.
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Xu, H.; Veyrat, N.; Degen, T.; Turlings, T.C.J. Exceptional Use of Sex Pheromones by Parasitoids of the Genus Cotesia: Males Are Strongly Attracted to Virgin Females, but Are No Longer Attracted to or Even Repelled by Mated Females. Insects 2014, 5, 499-512. https://doi.org/10.3390/insects5030499
Xu H, Veyrat N, Degen T, Turlings TCJ. Exceptional Use of Sex Pheromones by Parasitoids of the Genus Cotesia: Males Are Strongly Attracted to Virgin Females, but Are No Longer Attracted to or Even Repelled by Mated Females. Insects. 2014; 5(3):499-512. https://doi.org/10.3390/insects5030499
Chicago/Turabian StyleXu, Hao, Nathalie Veyrat, Thomas Degen, and Ted C. J. Turlings. 2014. "Exceptional Use of Sex Pheromones by Parasitoids of the Genus Cotesia: Males Are Strongly Attracted to Virgin Females, but Are No Longer Attracted to or Even Repelled by Mated Females" Insects 5, no. 3: 499-512. https://doi.org/10.3390/insects5030499
APA StyleXu, H., Veyrat, N., Degen, T., & Turlings, T. C. J. (2014). Exceptional Use of Sex Pheromones by Parasitoids of the Genus Cotesia: Males Are Strongly Attracted to Virgin Females, but Are No Longer Attracted to or Even Repelled by Mated Females. Insects, 5(3), 499-512. https://doi.org/10.3390/insects5030499