Influence of Age and Nutritional Status on Flight Performance of the Asian Tiger Mosquito Aedes albopictus (Diptera: Culicidae)

Abstract
:1. Introduction
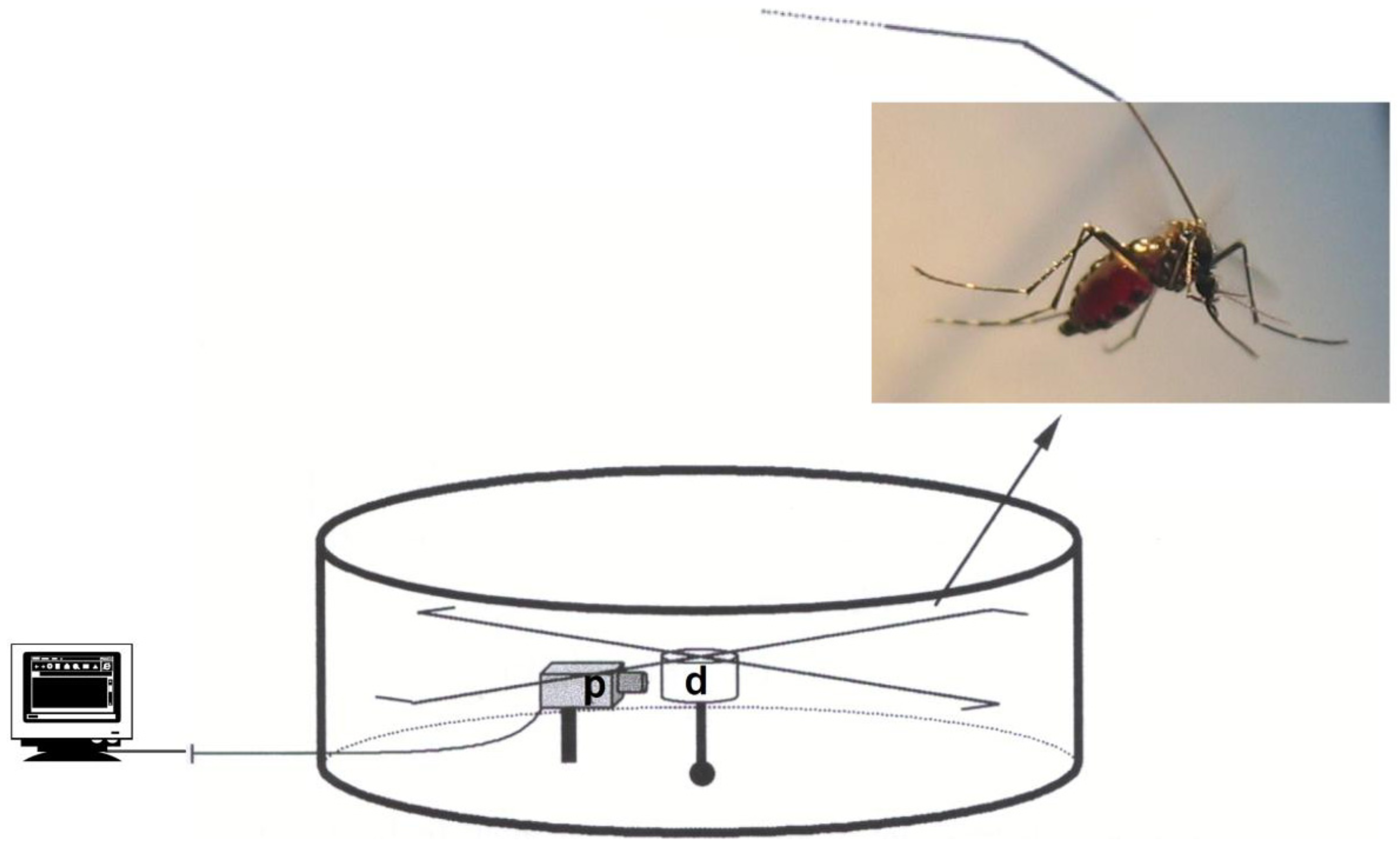
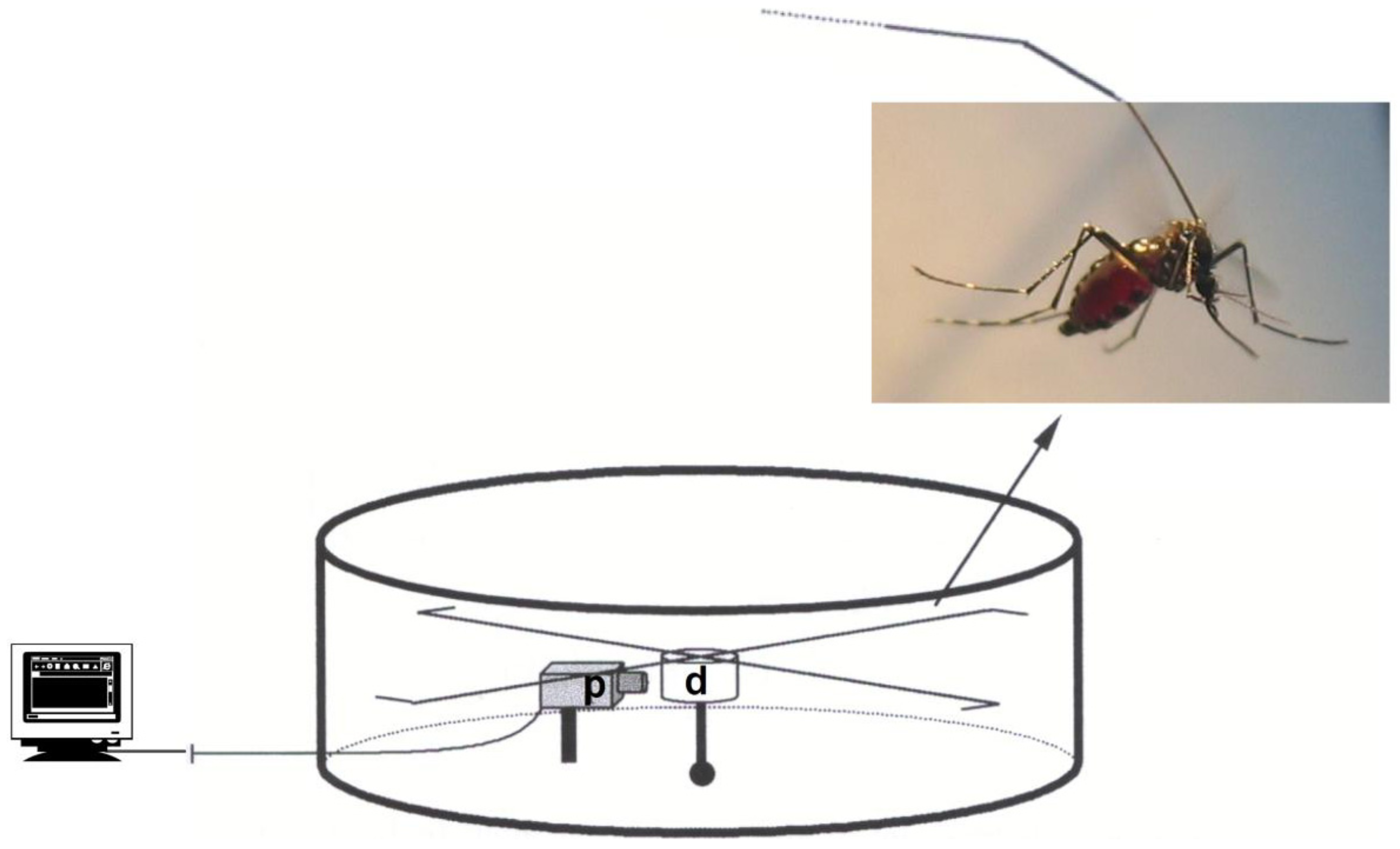
2. Material and Methods
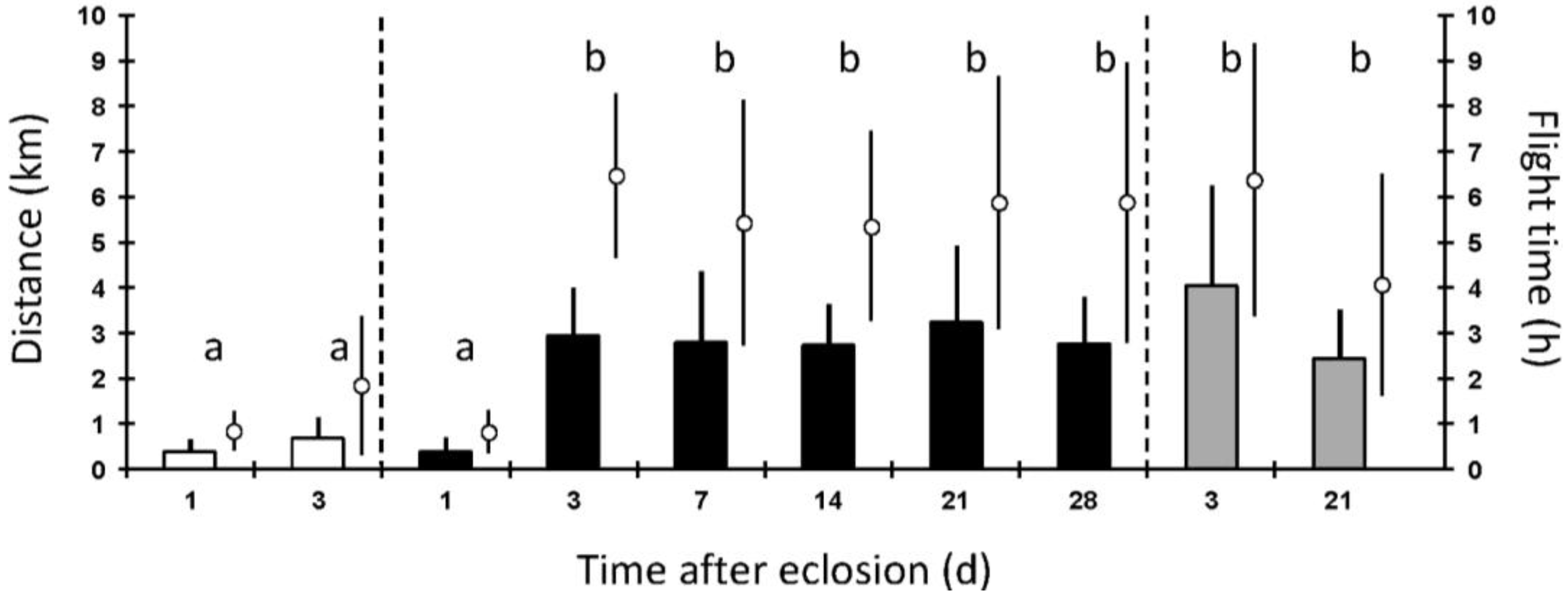
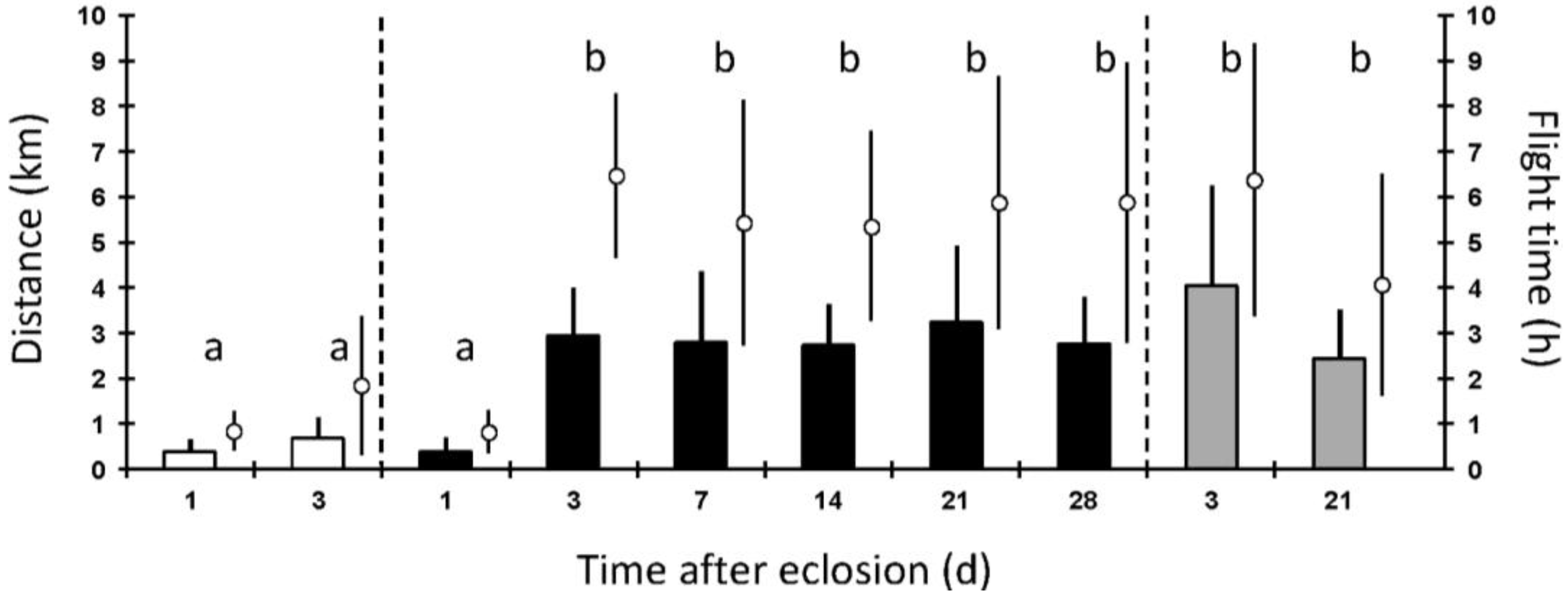
3. Results

{kind=link}
{kind=link}
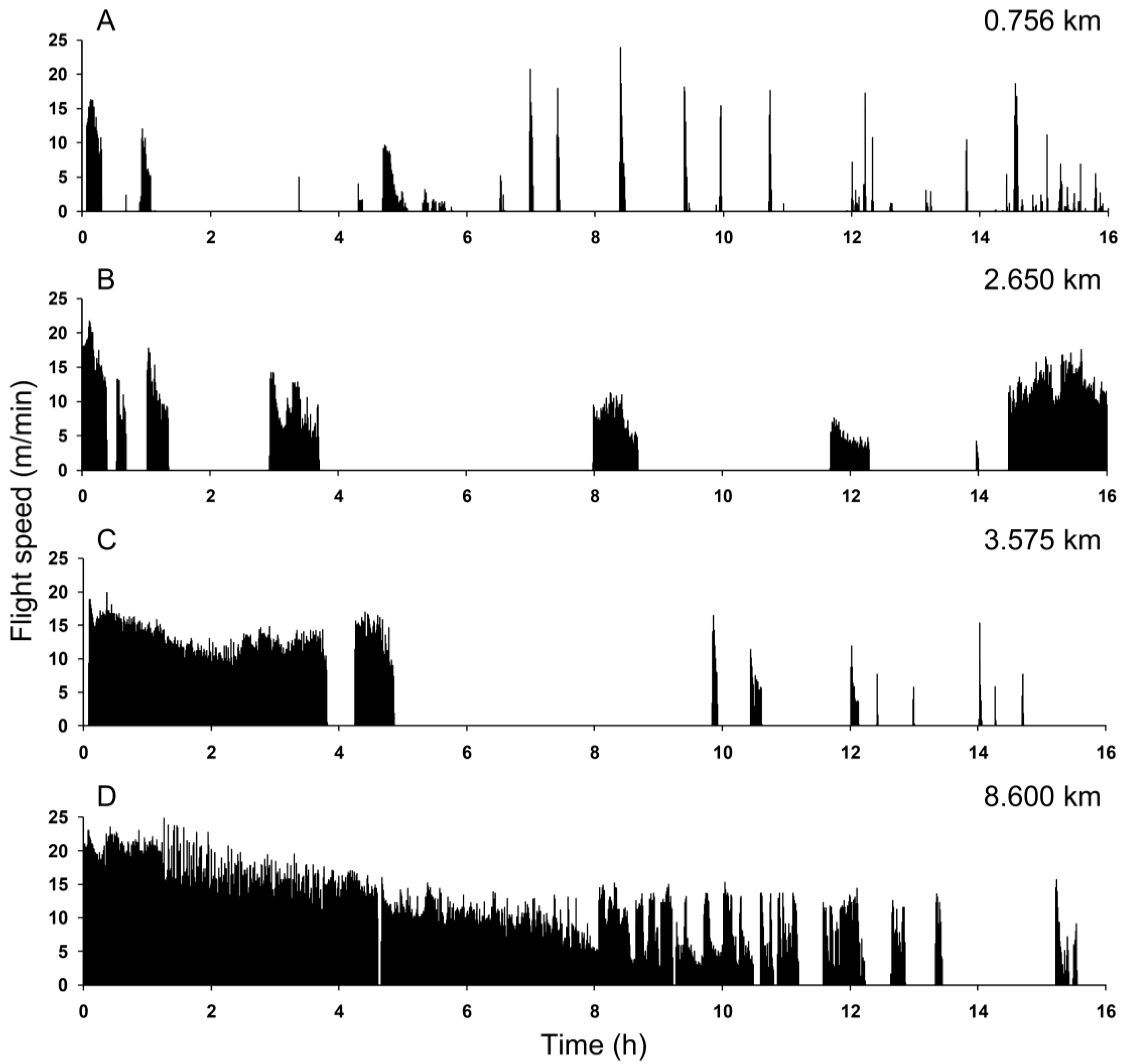{kind=link}
| Maximal continuous flights (h) | |||||
|---|---|---|---|---|---|
| <0.5 | 0.5–1 | 1–2 | 2–4 | >4 | |
| Feeding condition 1 (number of fliers) | % | ||||
| Starved (18) | 83 | 11 | 6 | 0 | 0 |
| Sugar-fed (16) | 6 | 19 | 25 | 37 | 13 |
| Blood-fed (17) | 6 | 6 | 12 | 41 | 35 |
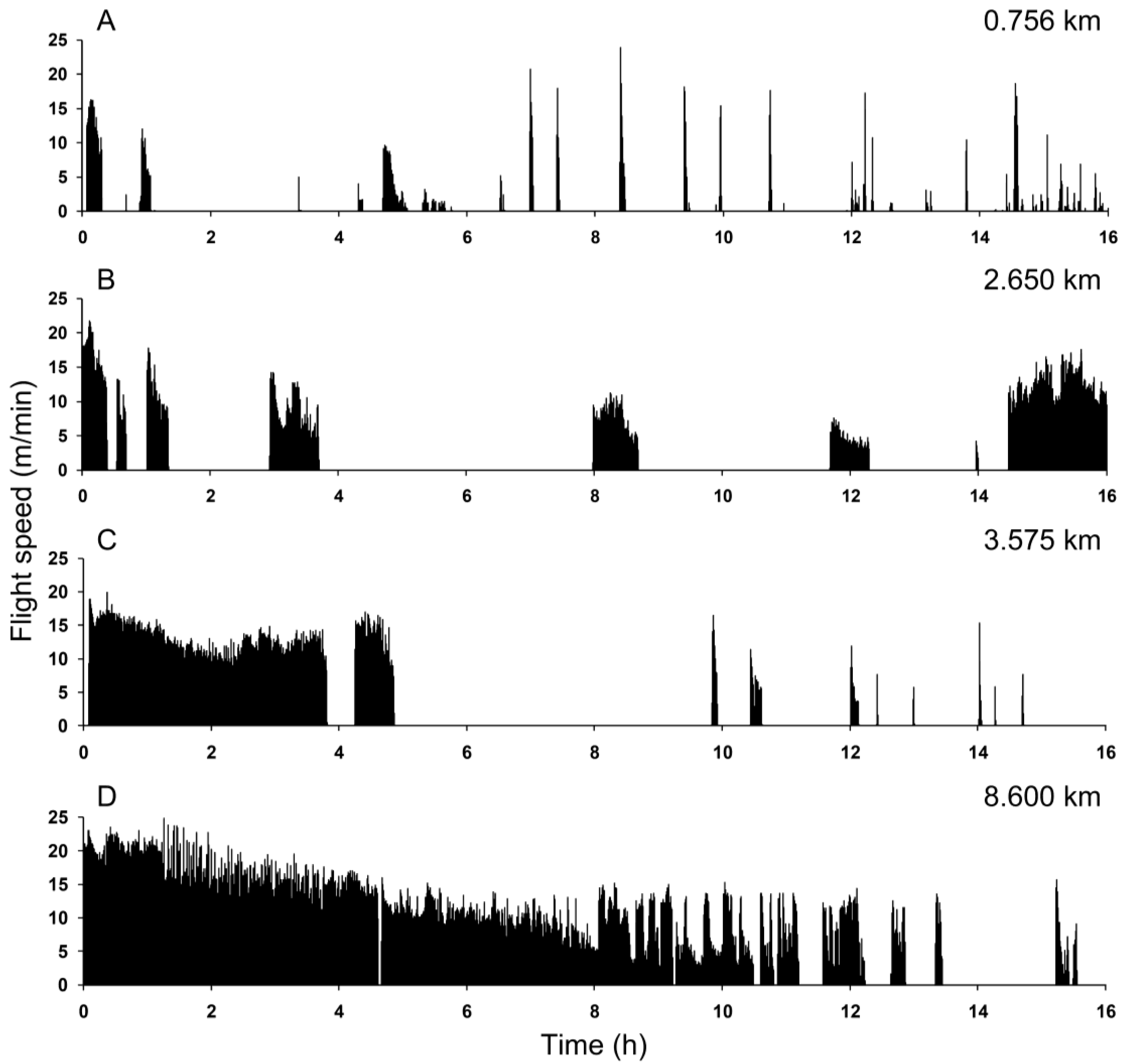
4. Discussion and Conclusions
Acknowledgments
Conflict of Interest
References
- Yee, D.A. Tires as Habitats for Mosquitoes: A Review of Studies within the Eastern United States. J. Med. Entomol. 2009, 45, 581–593. [Google Scholar] [CrossRef]
- Takumi, K.; Scholte, E.J.; Braks, M.; Reusken, C.; Avenell, D.; Medlock, J.M. Introduction, scenarios for establishment and seasonal activity of Aedes albopictus in The Netherlands. Vector Borne Zoonotic Dis. 2009, 9, 191–196. [Google Scholar] [CrossRef]
- Adhami, J.; Reiter, P. Introduction and establishment of Aedes (Stegomyia) albopictus skuse (Diptera: Culicidae) in Albania. J. Am. Mosq. Control Assoc. 1998, 14, 340–343. [Google Scholar]
- Romi, R.; Majori, G. An overview of the lesson learned in almost 20 years of fight against the “tiger” mosquito. Parassitologia 2008, 50, 117–119. [Google Scholar]
- Wymann, M.N.; Flacio, E.; Radczuweit, S.; Patocchi, N.; Lüthy, P. Asian tiger mosquito (Aedes albopictus)—A threat for Switzerland? Euro Surveill. 2008, 13, 8058. [Google Scholar]
- Schaffner, F.; Hendrickx, G.; Scholte, E.J.; Medlock, J.M.; Angelini, P.; Ducheyne, E. Development of Aedes albopictus risk maps; Technical report for European Centre for Disease Prevention and Control: Stockholm, Sweden, 2009; pp. 1–45. [Google Scholar]
- Sprenger, D.; Wuithiranyagool, T. The discovery and distribution of Aedes albopictus in Harris County, Texas. J. Am. Mosq. Control Assoc. 1986, 2, 217–219. [Google Scholar]
- Gratz, N.G. Critical review of the vector status of Aedes albopictus. Med. Vet. Entomol. 2004, 18, 215–227. [Google Scholar] [CrossRef]
- Bonilauri, P.; Bellini, R.; Calzolari, M.; Angelini, R.; Venturi, L.; Fallacara, F.; Cordioli, P.; Angelini, P.; Venturelli, C.; Merialdi, G.; et al. Chikungunya virus in Aedes albopictus, Italy. Emerg. Infect. Dis. 2008, 14, 852–854. [Google Scholar] [CrossRef]
- Lambrechts, L.; Scott, T.W.; Gubler, D.J. Consequences of the expanding global distribution of Aedes albopictus for dengue virus transmission. PLoS Negl. Trop. Dis. 2010, 4, e646. [Google Scholar] [CrossRef]
- Reiter, P.; Fontenille, D.; Paupy, C. Aedes albopictus as an epidemic vector of chikungunya virus: Another emerging problem? Lancet Infect. Dis. 2006, 6, 463–464. [Google Scholar] [CrossRef]
- Gould, E.A.; Gallian, P.; de Lamballerie, X.; Charrel, R.N. First cases of autochthonous dengue fever and chikungunya fever in France: From bad dream to reality! Clin. Microbiol. Infect. 2010, 16, 1702–1704. [Google Scholar] [CrossRef]
- Gage, K.L.; Burkot, T.R.; Eisen, R.J.; Hayes, E.B. Climate and Vectorborne Diseases. Am. J. Prev. Med. 2008, 35, 436–450. [Google Scholar] [CrossRef]
- Sutherst, R.W. Global change and human vulnerability to vector-borne diseases. Clin. Microbiol. Rev. 2004, 17, 136–173. [Google Scholar] [CrossRef]
- Talbalaghi, A.; Moutailler, S.; Vazeille, M.; Failloux, A.B. Are Aedes albopictus or other mosquito species from northern Italy competent to sustain new arboviral outbreaks? Med. Vet. Entomol. 2010, 24, 83–87. [Google Scholar] [CrossRef]
- Kaufmann, C.; Briegel, H. Flight performance of the malaria vectors Anopheles gambiae and Anopheles atroparvus. J. Vector Ecol. 2004, 29, 140–153. [Google Scholar]
- Briegel, H.; Knusel, I.; Timmermann, S.E. Aedes aegypti: Size, reserves, survival, and flight potential. J. Vector Ecol. 2001, 26, 21–31. [Google Scholar]
- Briegel, H.; Waltert, A.; Kuhn, A.R. Reproductive physiology of Aedes (Aedimorphus) vexans (Diptera: Culicidae) in relation to flight potential. J. Med. Entomol. 2001, 38, 557–565. [Google Scholar] [CrossRef]
- Clements, A.N. Flight. In The Biology of Mosquitoes: Sensory, Reception, and Behaviour; CABI: Oxford, UK, 1999; Volume 2, pp. 286–332. [Google Scholar]
- Bargielowski, I.; Kaufmann, C.; Alphey, L.; Reiter, P.; Koella, J. Flight performance and teneral energy reserves of two genetically-modified and one wild-type strain of the yellow fever mosquito Aedes aegypti. Vector Borne Zoonotic Dis. 2012, 12, 1053–1058. [Google Scholar] [CrossRef] [Green Version]
- Briegel, H.; Timmermann, S.E. Aedes albopictus (Diptera: Culicidae): Physiological aspects of development and reproduction. J. Med. Entomol. 2001, 38, 566–571. [Google Scholar] [CrossRef]
- Rowley, W.A.; Graham, C.L.; Williams, R.E. A flight mill system for the laboratory study of mosquito flight. Ann. Entomol. Soc. Am. 1968, 61, 1507–1514. [Google Scholar]
- SPSS Software. Available online: www.spss.com (accessed on 3 July 2013).
- Müller, G.C.; Xue, R.D.; Schlein, Y. Differential attraction of Aedes albopictus in the field to flowers, fruits and honeydew. Acta Trop. 2011, 118, 45–49. [Google Scholar] [CrossRef]
- Xue, R.D.; Barnard, D.R.; Müller, G.C. Effects of body size and nutritional regimen on survival in adult Aedes albopictus (Diptera: Culicidae). J. Med. Entomol. 2010, 47, 778–782. [Google Scholar] [CrossRef]
- Gary, R.E., Jr.; Foster, W.A. Effects of available sugar on the reproductive fitness and vectorial capacity of the malaria vector Anopheles gambiae (Diptera: Culicidae). J. Med. Entomol. 2001, 38, 22–28. [Google Scholar] [CrossRef]
- Honorio, N.A.; Silva Wda, C.; Leite, P.J.; Goncalves, J.M.; Lounibos, L.P.; Lourenco-de-Oliveira, R. Dispersal of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) in an urban endemic dengue area in the State of Rio de Janeiro, Brazil. Mem. Inst. Oswaldo Cruz 2003, 98, 191–198. [Google Scholar]
- Kawada, H.; Takemura, S.Y.; Arikawa, K.; Takagi, M. Comparative study on nocturnal behavior of Aedes aegypti and Aedes albopictus. J. Med. Entomol. 2005, 42, 312–318. [Google Scholar] [CrossRef]
- Liew, C.; Curtis, C.F. Horizontal and vertical dispersal of dengue vector mosquitoes, Aedes aegypti and Aedes albopictus, in Singapore. Med. Vet. Entomol. 2004, 18, 351–360. [Google Scholar] [CrossRef]
- Rowley, W.A.; Graham, C.L. The effect of temperature and relative humidity on the flight performance of female Aedes aegypti. J. Insect Physiol. 1968, 14, 1251–1257. [Google Scholar] [CrossRef]
- Marini, F.; Caputo, B.; Pombi, M.; Tarsitani, G.; della Torre, A. Study of Aedes albopictus dispersal in Rome, Italy, using sticky traps in mark-release-recapture experiments. Med. Vet. Entomol. 2010, 24, 361–368. [Google Scholar] [CrossRef]
- Lacroix, R.; Delatte, H.; Hue, T.; Reiter, P. Dispersal and survival of male and female Aedes albopictus (Diptera: Culicidae) on Reunion Island. J. Med. Entomol. 2009, 46, 1117–1124. [Google Scholar] [CrossRef]
- Bellini, R.; Albieri, A.; Balestrino, F.; Carrieri, M.; Porretta, D.; Urbanelli, S.; Calvitti, M.; Moretti, R.; Maini, S. Dispersal and survival of Aedes albopictus (Diptera: Culicidae) males in Italian urban areas and significance for sterile insect technique application. J. Med. Entomol. 2010, 47, 1082–1091. [Google Scholar] [CrossRef]
- Rozeboom, L.E.; Rosen, L.; Ikeda, J. Observations on oviposition by Aedes (S.) albopictus Skuse and A. (S.) polynesiensis Marks in nature. J. Med. Entomol. 1973, 10, 397–399. [Google Scholar]
- Reiter, P.; Amador, M.A.; Anderson, R.A.; Clark, G.G. Short report: Dispersal of Aedes aegypti in an urban area after blood feeding as demonstrated by rubidium-marked eggs. Am. J. Trop. Med. Hyg. 1995, 52, 177–179. [Google Scholar]
- WHO. Dengue: Guidelines for Diagnosis, Treatment, Prevention and Control; WHO: Geneva, Switzerland, 2009.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kaufmann, C.; Collins, L.F.; Brown, M.R. Influence of Age and Nutritional Status on Flight Performance of the Asian Tiger Mosquito Aedes albopictus (Diptera: Culicidae). Insects 2013, 4, 404-412. https://doi.org/10.3390/insects4030404
Kaufmann C, Collins LF, Brown MR. Influence of Age and Nutritional Status on Flight Performance of the Asian Tiger Mosquito Aedes albopictus (Diptera: Culicidae). Insects. 2013; 4(3):404-412. https://doi.org/10.3390/insects4030404
Chicago/Turabian StyleKaufmann, Christian, Lauren F. Collins, and Mark R. Brown. 2013. "Influence of Age and Nutritional Status on Flight Performance of the Asian Tiger Mosquito Aedes albopictus (Diptera: Culicidae)" Insects 4, no. 3: 404-412. https://doi.org/10.3390/insects4030404