Selective Capture of Transcribed Sequences: A Promising Approach for Investigating Bacterium-Insect Interactions

Abstract
:1. Introduction
2. In-vitro Profiling Techniques
3. Approaches in Bacteria-Host Interactions
3.1. Mutagenesis Based Methods
3.2. Hybridization Based Methods
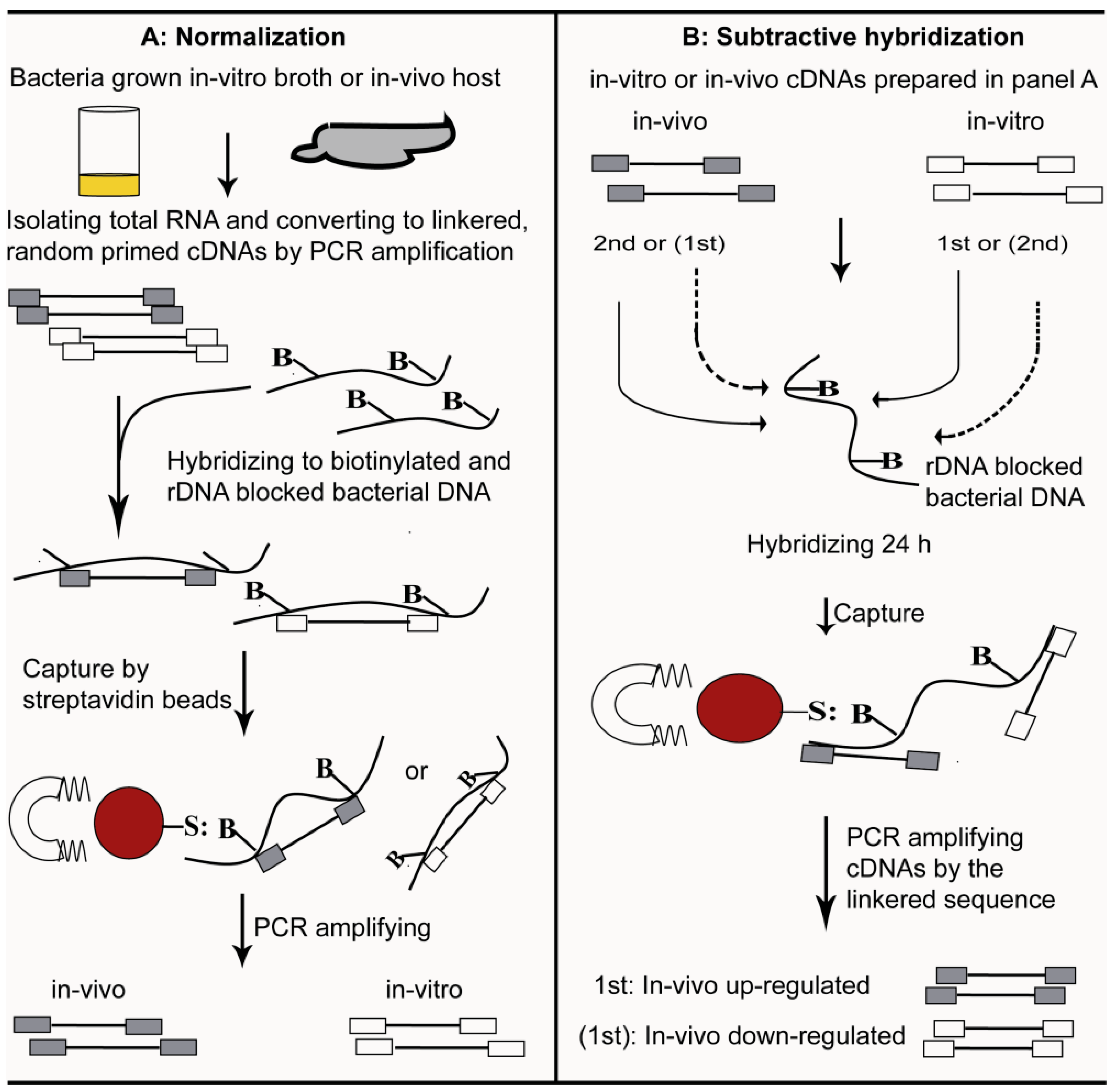
4. Selective Capture of Transcribed Sequences
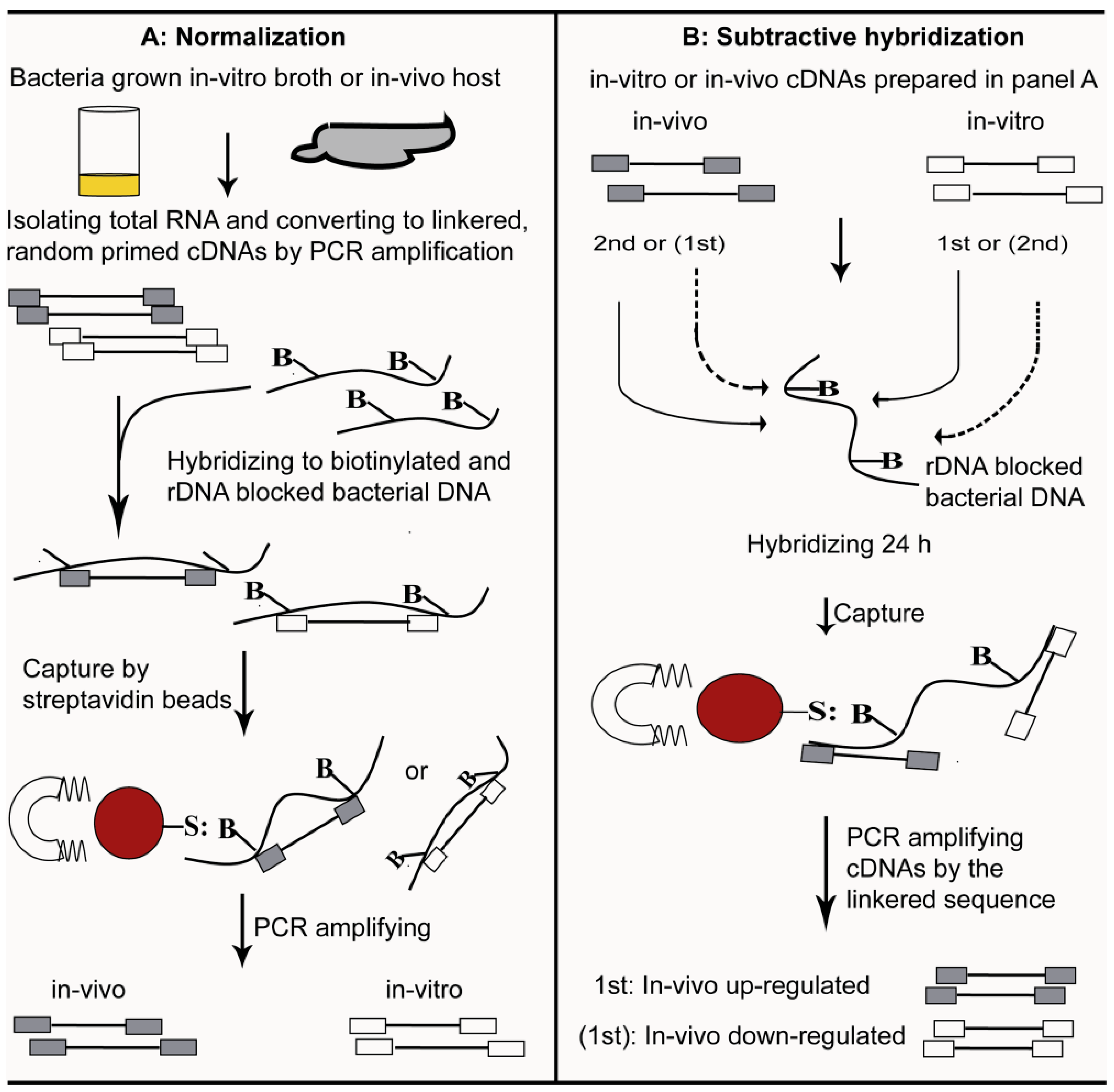
5. SCOTS versus other Methods

{kind=link}
| Approach | Bacteria Requirements | Animal Model | mRNA Requirements | Genetic Information |
|---|---|---|---|---|
| 2-D Gel | High titer | Not needed | Not needed | Not needed |
| DDRT-PCR | High titer | Not needed | High quality & quantity | Needed |
| Microarray | High titer | Not needed | High quality & quantity | Needed |
| IVET | Transformable | Needed | Not needed | Not needed |
| DFI | Transformable | Needed | Not needed | Not needed |
| STM | Transformable | Needed | Not needed | Not needed |
| GAMBIT | Transformable | Needed | Not needed | Needed |
| SCOTS | No restriction | Not needed | No restriction | Not needed |
6. Conclusions
References
- Gygi, S.P.; Rist, B.; Gerber, S.A.; Turecek, F.; Gelb, M.H.; Aebersold, R. Quantitative analysis of complex protein mixtures using isotopecoded affinity tags. Nat. Biotechnol. 1999, 17, 994–999. [Google Scholar]
- Boyce, J.D.; Cullen, P.A.; Adler, B. Genomic-scale analysis of bacterial gene and protein expression in the host. Emerg. Infect. Dis. 2004, 10, 1357–1362. [Google Scholar] [CrossRef]
- Schmidt, F.; Donahoe, S.; Hagens, K.; Mattow, J.; Schaible, U.; Kaufmann, S.; Aebersold, R.; Jungblut, P.R. Complementary analysis of the Mycobacterium tuberculosis proteome by two-dimensional electrophoresis and isotope coded affinity tag technology. Mol. Cell Proteomics 2004, 3, 24–42. [Google Scholar]
- Zuo, X.; Echan, L.; Hembach, P.; Tang, H.Y.; Speicher, K.D.; Santoli, D.; Speicher, D.W. Towards global analysis of mammalian proteomes using sample prefractionation prior to narrow pH range two-dimensional gels and using one-dimensional gels for insoluble and large proteins. Electrophoresis 2001, 22, 1603–1615. [Google Scholar] [CrossRef]
- Hamdan, M.; Righetti, P.G. Assessment of protein expression by means of 2-D gel electrophoresis with and without mass spectrometry. Mass Spectrom. Rev. 2003, 22, 272–284. [Google Scholar] [CrossRef]
- Lehoux, D.E.; Sanschagrin, F.; Levesque, R.C. Discovering essential and infection-related genes. Curr. Opin. Microbiol. 2001, 4, 515–519. [Google Scholar]
- Judson, N.; Mekalanos, J.J. TnAraOut, a transposon-based approach to identify and characterize essential bacterial genes. Nat. Biotechnol. 2000, 18, 740–745. [Google Scholar] [CrossRef]
- Reich, K.A.; Chovan, L.; Hessler, P. Genome scanning in Haemophilus influenzae for identification of essential genes. J. Bacteriol. 1999, 181, 4961–4968. [Google Scholar]
- Wong, S.M.; Mekalanos, J.J. Genetic footprinting with mariner-based transposition in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2000, 97, 10191–10196. [Google Scholar]
- Hare, R.S.; Walker, S.S.; Dorman, T.E.; Greene, J.R.; Guzman, L.M.; Kenney, T.J.; Sulavik, M.C.; Baradaran, K.; Houseweart, C.; Yu, H.; et al. Genetic footprinting in bacteria. J. Bacteriol. 2001, 183, 1694–1706. [Google Scholar]
- Akerley, B.J.; Rubin, E.J.; Lampe, D.J.; Mekalanos, J.J. PCR-mediated detection of growth-attenuated mutants in large pools generated by in vitro transposon mutagenesis. Atlanta, GA, USA, 17–21 May 1998; American Society for Microbiology: Atlanta, GA, USA, 1998. Presented at the 98th General Meeting of the American Society for Microbiology. [Google Scholar]
- Liang, P.; Pardee, A.B. Differential display of eukaryotic messenger RNA by means of the polymerase chain reaction. Science 1992, 257, 967–971. [Google Scholar]
- Rindi, L.; Lari, N.; Garzelli, C. Search for genes potentially involved in Mycobacterium tuberculosis virulence by mRNA differential display. Biochem. Biophys. Res. Commun. 1999, 258, 94–101. [Google Scholar] [CrossRef]
- Rivera-Marrero, C.A.; Burroughs, M.A.; Masse, R.A.; Vannberg, F.O.; Leimbach, D.L.; Roman, J.; Murtagh, J.J., Jr. Identification of genes differentially expressed in Mycobacterium tuberculosis by differential display PCR. Microb. Pathog. 1998, 25, 307–316. [Google Scholar] [CrossRef]
- Sturtevant, J. Applications of differential-display reverse transcription-PCR to molecular pathogenesis and medical mycology. Clin. Microbiol. Rev. 2000, 13, 408–427. [Google Scholar] [CrossRef]
- Mahan, M.J.; Slauch, J.M.; Mekalanos, J.J. Selection of bacterial virulence genes that are specifically induced in host tissues. Science 1993, 259, 686–688. [Google Scholar]
- Angelichio, M.J.; Camilli, A. In vivo expression technology. Infect. Immun. 2002, 70, 6518–6523. [Google Scholar] [CrossRef]
- Valdivia, R.H.; Falkow, S. Bacterial genetics by flow cytometry: Rapid isolation of Salmonella typhimurium acid-inducible promoters by differential fluorescence induction. Mol. Microbiol. 1996, 22, 367–378. [Google Scholar]
- Münch, A.; Stingl, L.; Jung, K.; Heermann, R. Photorhabdus luminescens genes induced upon insect infection. BMC Genomics 2008, 9, 229. [Google Scholar] [CrossRef]
- Hensel, M.; Shea, J.E.; Gleeson, C.; Jones, M.D.; Dalton, E.; Holden, D.W. Simultaneous identification of bacterial virulence genes by negative selection. Science 1995, 269, 400–403. [Google Scholar]
- Mecsas, J. Use of signature-tagged mutagenesis in pathogenesis studies. Curr. Opin. Microbiol. 2002, 5, 33–37. [Google Scholar]
- Lehoux, D.E.; Levesque, R.C. Detection of genes essential in specific niches by signature-tagged mutagenesis. Curr. Opin. Biotechnol. 2000, 11, 434–439. [Google Scholar] [CrossRef]
- Shea, J.E.; Santangelo, J.D.; Feldman, R.G. Signature-tagged mutagenesis in the identification of virulence genes in pathogens. Curr. Opin. Microbiol. 2000, 3, 451–458. [Google Scholar]
- Autret, N.; Charbit, A. Lessons from signature-tagged mutagenesis on the infectious mechanisms of pathogenic bacteria. FEMS Microbiol. Rev. 2005, 29, 703–717. [Google Scholar] [CrossRef]
- Saenz, H.L.; Dehio, C. Signature-tagged mutagenesis: Technical advances in a negative selection method for virulence gene identification. Curr. Opin. Microbiol. 2005, 8, 612–619. [Google Scholar]
- Boyce, J.D.; Wilkie, I.; Harper, M.; Paustian, M.L.; Kapur, V.; Adler, B. Genomic-scale analysis of Pasteurella multocida gene expression during growth within liver tissue of chickens with fowl cholera. Microbes Infect. 2004, 6, 290–298. [Google Scholar] [CrossRef]
- Boyce, J.D.; Wilkie, I.; Harper, M.; Paustian, M.L.; Kapur, V.; Adler, B. Genomic scale analysis of Pasteurella multocida gene expression during growth within the natural chicken host. Infect. Immun. 2002, 70, 6871–6879. [Google Scholar]
- Xu, Q.; Dziejman, M.; Mekalanos, J.J. Determination of the transcriptome of Vibrio cholerae during intraintestinal growth and midexponential phase in vitro. Proc. Natl. Acad. Sci. USA 2003, 100, 1286–1291. [Google Scholar]
- Paustian, M.L.; May, B.J.; Kapur, V. Pasteurella multocida gene expression in response to iron limitation. Infect. Immun. 2001, 69, 4109–4115. [Google Scholar]
- Schembri, M.A.; Kjaergaard, K.; Klemm, P. Global gene expression in Escherichia coli biofilms. Mol. Microbiol. 2003, 48, 253–267. [Google Scholar] [CrossRef]
- Alland, D.; Kramnik, I.; Weisbrod, T.R.; Otsubo, L.; Cerny, R.; Miller, L.P.; Jacobs, W.R., Jr.; Bloom, B.R. Identification of differentially expressed mRNA in prokaryotic organisms by customized amplification libraries (DECAL): The effect of isoniazid on gene expression in Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 1998, 95, 13227–13232. [Google Scholar]
- Narasimhan, S.; Santiago, F.; Koski, R.A.; Brei, B.; Anderson, J.F.; Fish, D.; Fikrig, E. Examinatin of the Borrelia burgdorferi transcriptome in lxodes scapularis during feeding. J. Bacteriol. 2002, 184, 3122–3125. [Google Scholar]
- Narasimhan, S.; Camaino, M.J.; Liang, F.T.; Santiago, F.; Laskowski, M.; Philipp, M.T.; Pachner, A.R.; Radolf, J.D.; Fikrig, E. Borrelia burgdorferi transcriptome in the central nervous system of non-human primates. Proc. Natl. Acad. Sci. USA 2003, 100, 15953–15958. [Google Scholar]
- Shelburne, S.A.; Musser, J.M. Virulence gene expression in vivo. Curr. Opin. Microbiol. 2004, 7, 283–289. [Google Scholar]
- Graham, J.E.; Clark-Curtiss, J.E. Identification of Mycobacterium tuberculosis RNAs synthesized in response to phagocytosis by human macrophages by selective capture of transcribed sequences (SCOTS). Proc. Natl. Acad. Sci. USA 1999, 96, 11554–11559. [Google Scholar]
- Liu, S.; Graham, J.E.; Bigelow, L.; Morse, P.D., 2nd; Wilkinson, B.J. Identification of Listeria monocytogenes genes expressed in response to growth at low temperature. Appl. Environ. Microbiol. 2002, 68, 1697–1705. [Google Scholar] [CrossRef]
- Graham, J.E.; Peek, R.M., Jr.; Krishna, U.; Cover, T.L. Global analysis of Helicobacter pylori gene expression in human gastric mucosa. Gastroenterology 2002, 123, 1637–1648. [Google Scholar] [CrossRef]
- Dozois, C.M.; Daigle, F.; Curtiss, R., 3rd. Identification of pathogen-specific and conserved genes expressed in vivo by an avian pathogenic Escherichia coli strain. Proc. Natl. Acad. Sci. USA 2003, 100, 247–252. [Google Scholar]
- Baltes, N.; Gerlach, G.F. Identification of genes transcribed by Actinobacillus pleuropneumoniae in necrotic porcine lung tissue by using selective capture of transcribed sequences. Infect. Immun. 2004, 72, 6711–6716. [Google Scholar] [CrossRef]
- An, R.; Sreevatsan, S.; Grewal, P.S. Comparative in vivo gene expression of the closely related bacteria Photorhabdus temperata and Xenorhabdus koppenhoeferi upon infection of the same insect host, Rhizotrogus majalis. BMC Genomics 2009, 10, 433. [Google Scholar]
- An, R.; Sreevatsan, S.; Grewal, P.S. Moraxella osloensis gene expression in the slug host Deroceras reticulatum. BMC Microbiology 2008, 8, 19. [Google Scholar] [CrossRef]
- Li, W.; Liu, L.; Qiu, D.; Chen, H.; Zhou, R. Identification of Streptococcus suis serotype 2 genes preferentially expressed in the natural host. Int. J. Med. Microbiol. 2010, 300, 482–488. [Google Scholar] [CrossRef]
- Emboule, L.; Daigle, F.; Meyer, D.F.; Mari, B.; Pinarello, V.; Sheikboudou, C.; Magnone, V.; Frutos, R.; Viari, A.; Barbry, P.; et al. Innovative approach for transcriptomic analysis of obligate intracellular pathogen: Selective capture of transcribed sequences of Ehrlichia ruminantium. BMC Mol. Biol. 2009, 10, 111. [Google Scholar] [CrossRef]
- Zhou, Z.; Zheng, J.; Tian, W.; Li, J.; Zhang, W.; Zhang, J.; Meng, X.; Hu, S.; Bi, D.; Li, Z. Identification of Riemerella anatipestifer genes differentially expressed in infected duck livers by the selective capture of transcribed sequences technique. Avian Pathol. 2009, 38, 321–329. [Google Scholar] [CrossRef]
- Jin, H.; Wan, Y.; Zhou, R.; Li, L.; Luo, R.; Zhang, S.; Hu, J.; Langford, P.R.; Chen, H. Identification of genes transcribed by Haemophilus parasuis in necrotic porcine lung through the selective capture of transcribed sequences (SCOTS). Environ. Microbiol. 2008, 10, 3326–3336. [Google Scholar]
- Daigle, F.; Hou, J.Y.; Clark-Curtiss, J.E. Microbial gene expression elucidated by selective capture of transcribed sequences (SCOTS). Methods Enzymol. 2002, 358, 108–122. [Google Scholar]
- Haydel, S.E.; Benjamin, W.H., Jr.; Dunlap, N.E.; Clark-Curtiss, J.E. Expression, autoregulation, and DNA binding properties of the Mycobacterium tuberculosis TrcR response regulator. J. Bacteriol. 2002, 184, 2192–2203. [Google Scholar] [CrossRef]
- Faucher, S.P.; Porwollik, S.; Dozois, C.M.; McClelland, M.; Daigle, F. Transcriptome of Salmonella enterica serovar Typhi within macrophages revealed through the selective capture of transcribed sequences. Proc. Natl. Acad. Sci. USA 2006, 103, 1906–1911. [Google Scholar]
- Guerrero-Ferreira, R.C.; Nishiguchi, M.K. Differential gene expression in bacterial symbionts from loliginid squids demonstrates variation between mutualistic and environmental niches. Environ. Microbiol. Rep. 2010, 2, 514–523. [Google Scholar] [CrossRef]
- An, R.; Grewal, P.S. Molecular mechanisms of persistence of mutualistic bacteria Photorhabdus in the entomopathogenic nematode host. PLoS One 2010, 5, e13154. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
An, R.; Grewal, P.S. Selective Capture of Transcribed Sequences: A Promising Approach for Investigating Bacterium-Insect Interactions. Insects 2012, 3, 295-306. https://doi.org/10.3390/insects3010295
An R, Grewal PS. Selective Capture of Transcribed Sequences: A Promising Approach for Investigating Bacterium-Insect Interactions. Insects. 2012; 3(1):295-306. https://doi.org/10.3390/insects3010295
Chicago/Turabian StyleAn, Ruisheng, and Parwinder S. Grewal. 2012. "Selective Capture of Transcribed Sequences: A Promising Approach for Investigating Bacterium-Insect Interactions" Insects 3, no. 1: 295-306. https://doi.org/10.3390/insects3010295