
Digestive Characteristics of Hyphantria cunea Larvae on Different Host Plants
by
Aoying Zhang
1,2,†, 
Tao Li
1,2,†,
Lisha Yuan
1,2,
Mingtao Tan
1,2,
Dun Jiang
1,2,* and
Shanchun Yan
1,2,* 1
School of Forestry, Northeast Forestry University, Harbin 150040, China
2
Key Laboratory of Sustainable Forest Ecosystem Management-Ministry of Education, Northeast Forestry University, Harbin 150040, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Insects 2023, 14(5), 463; https://doi.org/10.3390/insects14050463
Submission received: 1 April 2023
/
Revised: 11 May 2023
/
Accepted: 12 May 2023
/
Published: 14 May 2023
(This article belongs to the Section Insect Physiology, Reproduction and Development)
Abstract
:Simple Summary
Hyphantria cunea is a cosmopolitan quarantine pest that can sustain on a wide range of host plants. However, the role of the digestive physiology of H. cunea larvae in regulating its multi-host adaptation has not been reported. Herein, we found that α-amylase and trypsin played an important role in nutrient metabolism and host adaptation of H. cunea. Furthermore, H. cunea larvae had highly adaptable compensatory mechanisms for digestion in response to insect digestive enzyme inhibitors. The findings reveal that digestive physiology mediates the multi-host adaptation of H. cunea larvae.
Abstract
Digestive physiology mediates the adaptation of phytophagous insects to host plants. In this study, the digestive characteristics of Hyphantria cunea larvae feeding preferences on different host plants were investigated. The results showed that the body weight, food utilization, and nutrient contents of H. cunea larvae feeding on the high-preference host plants were significantly higher than those feeding on the low-preference host plants. However, the activity of larval digestive enzymes in different host plants presented an opposite trend, as higher α-amylase or trypsin activity was observed in the group feeding on the low-preference host plants than that feeding on the high-preference host plants. Upon treatment of leaves with α-amylase and trypsin inhibitors, the body weight, food intake, food utilization rate, and food conversion rate of H. cunea larvae significantly decreased in all host plant groups. Furthermore, the H. cunea comprised highly adaptable compensatory mechanisms of digestion involving digestive enzymes and nutrient metabolism in response to digestive enzyme inhibitors. Taken together, digestive physiology mediates the adaptation of H. cunea to multiple host plants, and the compensatory effect of digestive physiology is an important counter-defense strategy implemented by H. cunea to resist plant defense factors, especially the insect digestive enzyme inhibitors.
1. Introduction
Hyphantria cunea (Lepidoptera: Erebidae) is a cosmopolitan quarantine pest that originated in North America and is mainly distributed in the United States and southern Canada [1,2]. It was first introduced in Hungary from its origin in 1940 and exploded on a large scale in 1945 [3]. At present, H. cunea has invaded over 32 countries [4]. In China, the H. cunea invasion began in 1979, and since then, it has spread to 613 county-level administrative regions in 14 Chinese provinces. H. cunea exhibits the characteristics of high food intake and prolonged damage time [5]. Upon invasion, H. cunea can eat up the whole plant leaves, inhibit the growth of trees and crops, and eventually result in severe destruction of forest and agriculture ecological balance [6]. Additionally, H. cunea often feeds and pupates on plants in public areas, such as parks and on the sides of roads, thereby affecting the daily life of local residents [7]. The damages caused by H. cunea have seriously threatened agriculture and forestry production and ecological security [8]. In 2022, ten Chinese ministerial-level departments listed the control of H. cunea as a key mission.
H. cunea, as a high polyphagy pest, can sustain on a wide range of host plants. It has been reported that H. cunea can feed on 636 host plants, of which 120 are found in the United States, 300 in Japan, 219 in South Korea, and 234 in Europe [9,10,11,12]. In China, more than 300 host plants have been reported as potential hosts for H. cunea, covering almost all the tree, flower, and crop species [13]. Phytophagous insects exhibit distinct preferences for the host plants [14]. Schowalter et al. (2017) found that H. cunea exhibits a high preference for Morus alba, Platanus acerifolia, Carya cathayensis, Fraxinus pennsylvanica, and Ulmus americana [7]. Jang et al. (2015) found that Cornus kousa and most coniferous tree species are non-feeding species for H. cunea [15]. Numerous studies revealed that during the process of co-evolution between phytophagous insects and plants, insects evolved a digestive physiological regulatory mechanism to achieve the best adaptation for different host plants. As demonstrated by Lazarevi et al. (2017), Lymantria dispar larvae exhibit increased trypsin activity upon feeding on the low-preference host plant, Robinia pseudoacacia, compared with the high-preference host plant, Quercus palustris [16]. Sarate et al. (2012) showed that Helicoverpa armigera exhibits enhanced larval adaptation to pigeon peas and chickpeas (low-preference host plants) through increasing gut amylase activity [17]. Strong host adaptability is also an important aspect of the wide host spectrum for H. cunea [18]. Currently, the studies on the host adaptability of H. cunea mainly focus on the host plant screening, as well as the feeding and oviposition selection behavior of H. cunea to different host plants [19,20]. However, no systematic study has been conducted to explore the adaptive mechanism of H. cunea larvae to multiple host plants from the perspective of digestive physiology.
Digestion and absorption are two important aspects of digestive physiology and can affect the adaptability of phytophagous insects to host plants at varying degrees [21]. The levels of these two aspects are mainly determined by the digestive enzyme activity of phytophagous insects, such as α-amylase and trypsin. In this regard, we hypothesized that digestive physiology based on digestive enzymes mediates the formation of multiple host characteristics of H. cunea. To test the above-mentioned hypothesis, we fed H. cunea larvae with leaves of high-preference (Ulmus pumila, Fraxinus mandshurica, and M. alba), medium-preference (Juglans mandshurica and Betula platyphylla), and low-preference (Albizia kalkora and Tilia amurensis) host plants. Subsequently, the larval growth and food utilization status, content of nutrients, and activity of digestive enzymes were measured. Furthermore, the regulatory roles of digestive physiology on the adaptation of H. cunea to multiple host plants were demonstrated in reverse by the treatment of α-amylase and trypsin inhibitors. The relevant findings of this study helped in determining the correlation between the digestive physiology and multi-host adaptability of H. cunea and provide a theoretical foundation to analyze the outbreak of H. cunea.
2. Materials and Methods
2.1. Experimental Host Plants
According to the results of our previous study [22], the larvae of H. cunea exhibit a high preference for U. pumila (UP), F. mandshurica (FM), and M. alba (MA), a medium preference for J. mandshurica (JM) and B. platyphylla (BP), and a low preference for A. kalkora (AK) and T. amurensis (TA). Therefore, these seven tree species were selected as host plants to test in this study. Except for MA, which was 2 years old (purchased from Baima Mountain Spring Agricultural Development Co., Ltd., Taian, China), the rest of the trees were 3 years old (purchased from Shenyang Ruihe Technology Service Co., Ltd., Shenyang, China) at the time of testing. In May 2022, all plants were planted in pots (400 × 350 mm) containing 10 kg of soil substrate. The soil substrate consisted of sand, turf soil, and vermiculite at a ratio of 1:3:1. A total of 300 host plants per species were planted and maintained under natural light conditions. During the experiment, the plants were watered every two days and weeded over time to ensure normal growth.
2.2. Experimental Insects
In July 2022, egg masses of H. cunea were collected from Hengshui, Hebei province, China. The egg masses were incubated in an incubator (HPG-280HX, Harbin Donglian Electronic Technology Development Co., Ltd., Harbin, China) at a temperature of 25 ± 1 °C, relative humidity of 70 ± 1%, and a photoperiod of 16L:8D [23]. After hatching, the larvae were reared on the artificial diets (purchased from the Ecology and Nature Conservation Institute, Chinese Academy of Forestry, Beijing, China) under the same conditions until the third instar. Third-instar larvae with a molting time of less than 24 h were selected. All larvae were divided into 7 groups and reared on the leaves of 7 host plants, respectively. The temperature, humidity, and photoperiod of the larval growth environment were consistent with the incubation environment. The larvae were not starved before being transferred from artificial diet to the leaves. The larvae were kept in plastic boxes that contain sufficient leaves (133 × 80 × 48 mm). Each box was fed with 5 larvae. To keep the food plentiful and fresh, the leaves were changed once a day.
2.3. Treatment with Insect Digestive Enzyme Inhibitors
Newly molted 4th instar larvae were selected from each host plant group. These larvae from each host plant group were divided into three parts. One part was used as the non-inhibited group (denoted as CK) and continued to be reared on the original plant leaves. The other two parts were fed with leaves of host plants sprayed with either α-amylase inhibitor (denoted as AI) or trypsin inhibitor (denoted as TI), respectively. The α-amylase inhibitor was purchased from Shanghai Renbang Pharmaceutical Technology Co., Ltd., Shanghai, China and the trypsin inhibitor was purchased from Shanghai Beyotime Biotechnology (Shanghai, China). Based on the results of our preliminary experiments, the α-amylase inhibitor and trypsin inhibitor were constituted in distilled water at a concentration of 750 g/L and 200 g/L, respectively. The prepared inhibitor solutions were evenly sprayed onto the leaf surface of each host plant. A total of 1 mL of treatment solution was sprayed on every 5 leaves. After drying at room temperature, the leaves were used to feed the 4th instar larvae of H. cunea. When these larvae reached the 5th instar stage, they were divided into two parts. One part was propagated for the next 48 h and then collected to obtain the whole larval body or midgut tissue samples. The other part was used to determine the larval body weight and food utilization parameters.
2.4. Body Weight and Food Utilization of H. cunea Larvae
Using the method developed by Gao et al. (2022) and Jiang et al. (2018), the food utilization by H. cunea larvae was measured [24,25]. During the experiment, the larval body weight and fresh leaf mass were measured before feeding, and then, the larval body weight, fecal mass, and residual leaf mass were measured after 48 h of feeding. The corrected water loss rate was calculated using the leaves that were not fed by H. cunea larvae. A total of three replicates were set for each group, and each replicate consisted of 10 larvae. The food intake, food consumption rate, conversion rate, and utilization rate of H. cunea larvae were calculated using the following formula:
Food intake (g) = (mass of fresh leaves before feeding − mass of residual leaves after feeding)/(1 − corrected water loss rate);
Food consumption rate (%) = (food intake − fecal discharge quality)/food intake × 100%;
Food conversion rate (%) = (weight after feeding − weight before feeding)/(food intake − feces discharge quality) × 100%;
Food utilization rate (%) = (weight after feeding − weight before feeding)/food intake × 100%.
2.5. Determination of Nutrient Contents in H. cunea Larvae
The contents of total protein (A045-4), total amino acids (A026-1-1), glucose (A54-1-1), trehalose (A49-1-1), and free fatty acids (A042-2-1) in the larvae of H. cunea were determined using the kit. All kits were purchased from Nanjing Jiancheng Bioengineering Institute. Briefly, the larvae in each group, at a ratio of weight (g): volume (mL) = 1:9, were mixed with normal saline and ground using a mortar and pestle. The larvae tissues for trehalose determination were ground with the extracting solution provided in the kit. The homogenate was centrifuged at the speed and time per the manufacturer’s instructions (10 min at 2500 rpm for total protein, glucose and free fatty acids; 10 min at 3500 rpm for total amino acids; 10 min at 8000× g for trehalose). Subsequently, the supernatant was collected for nutrient content determination according to the manufacturer’s instructions. Three replicates were set for each group, and each replicate consisted of three larvae. The total protein, total amino acids, glucose, trehalose, and free fatty acids contents of H. cunea were expressed as mg/g, μmol/mg prot, mmol/g prot, mg/g tissue, and mmol/g prot, respectively.
2.6. Determination of Digestive Enzyme Activity in the Midgut of H. cunea Larvae
The content or activity of the protein (A045-4), α-amylase (C06-1-1), and trypsin (A080-2) in H. cunea larval midgut were determined using the kit. All kits were purchased from Nanjing Jiancheng Bioengineering Institute. The tissue was mixed with normal saline at a ratio of weight (g): volume (mL) = 1:9. Subsequently, the samples were ground using a plastic grinding pestle in a 1.5 mL centrifuge tube. After centrifugation at 2500 rpm for 10 min, the supernatant was collected. The protein concentration and α-amylase and trypsin activities of the supernatant were determined according to the manufacturer’s instructions. Three replicates were set for each group, and each replicate consisted of midgut tissue from five larvae. Both α-amylase and trypsin activities were expressed as U/mg prot.
2.7. Statistical Analysis
The significance of larval body weight, food utilization index, nutrient content index, and activity of digestive enzymes among different host plant groups were analyzed by the one-way analysis of variance (ANOVA) after testing for variance homogeneity and normal distribution or achieving variance homogeneity and normal distribution by log-transforming if necessary (Tables S1 and S2). Independent sample t-test analysis was performed to analyze the significance of each parameter between the non-inhibited and AI treatment groups or between the non-inhibited and TI treatment groups (p < 0.05) (Table S3).
3. Results
3.1. Digestive Enzyme Activity of H. cunea Larvae
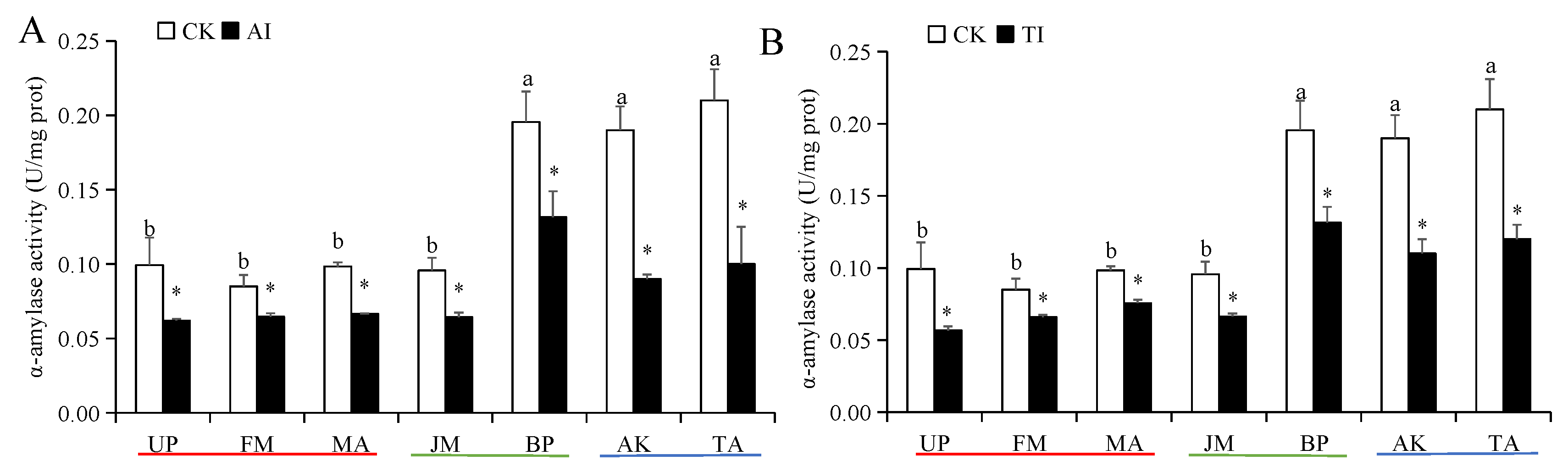
The larval α-amylase and trypsin activities in the BP, AK, and TA groups were significantly higher than those in the UP, FM, MA, and JM groups (Figure 1 and Figure 2). Compared with the CK group, the larval α-amylase activity in all AI-treated host plant groups was significantly decreased (Figure 1A). Among them, those feeding on AI-treated FM groups exhibited the lowest activity. However, the larval trypsin activity was significantly increased in the medium-preference host plant (JM and BP) groups after AI treatment and significantly decreased in the high- and low-preference host plant (UP, FM, MA, AK, and TA) groups after AI treatment (Figure 2A). After TI treatment, the α-amylase and trypsin activities of larvae were significantly inhibited in all host plant groups (Figure 1B and Figure 2B). Of which, the strongest inhibition effect on the two digestive enzyme activities was observed in the TI-treated TA group, and the inhibition rate of α-amylase activity and trypsin activity was 52.38% and 58.42%, respectively (Figure 1B and Figure 2B).
3.2. Growth status of H. cunea Larvae
The body weight of H. cunea larvae that were fed on high-preference host plants was significantly higher than those fed on low-preference host plants (Figure 3). Of these, the larvae that were fed on UP exhibited the highest body weight, and those fed on TA exhibited the lowest body weight. The larval body weight in all AI- or TI-treated host plant groups was significantly suppressed compared to the CK group (Figure 3A,B). The most significant suppression effect of either AI or TI treatment was observed in the UP group, with the inhibition rates as high as 61.68% and 61.85%, respectively (Figure 3A,B).
3.3. Food Utilization of H. cunea Larvae
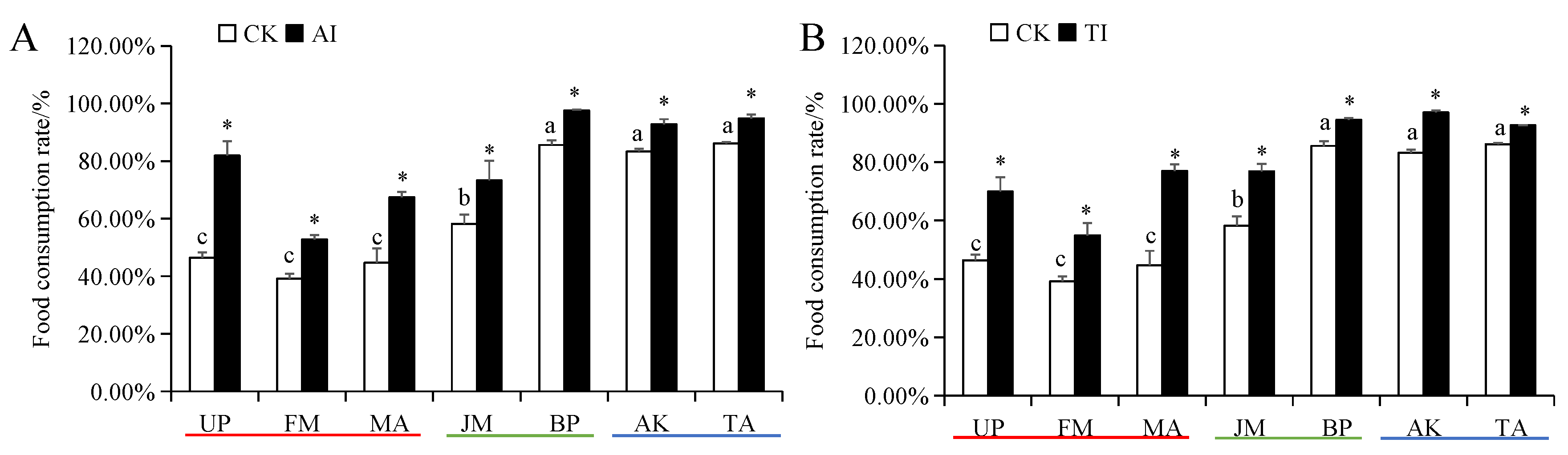
The food intake and food conversion rates of H. cunea larvae in the high-preference host plant (UP, FM, and MA) groups were significantly higher than those in medium- and low-preference host plant groups (Figures S1 and S2). The food utilization rate of H. cunea larvae on high- and medium-preference host plants was significantly higher than those on low-preference host plants (Figure 4). The food consumption rate of H. cunea larvae on high-preference plants was significantly lower than those on medium- and low-preference host plants (Figure 5). Compared with the CK group, after UP, FM, MA, JM, BP, AK, and TA groups were treated with AI or TI, the larval food intake, food conversion rate and food utilization rate of H. cunea larvae exhibited a significant decrease (Figures S1 and S2 and Figure 4), whereas the larval food consumption rate was significantly elevated (Figure 5).
3.4. Nutrient Contents of H. cunea Larvae
The contents of various nutrients (total protein, total amino acids, glucose, trehalose, and free fatty acids) in larvae that were fed on the high-preference host plants were significantly higher than those fed on the low-preference host plants (Table 1). Upon AI or TI treatment, the total protein content of larvae significantly increased in the AK and medium-preference host plant groups (JM and BP) but significantly decreased in the high-preference host plant groups; the total amino acids content of larvae was significantly increased in the MA group, but significantly decreased in the FM, medium- and low-preference host plant groups (JM, BP, AK, and TA); the larval contents of glucose and trehalose in all plant groups were significantly decreased; the larval content of free fatty acids was significantly increased in the JM and high-preference host plant groups (UP, FM, and MA), but significantly decreased in the BP and low-preference host plant groups (AK and TA).
4. Discussion
Generally, phytophagous insects use their own digestive physiological mechanisms to improve the efficiency of food utilization for adapting to the host plants [26,27]. In the present study, the body weight, food utilization rate, and nutrient contents (e.g., total protein and total amino acids) of H. cunea larvae feeding on the high-preference host plants were found to be significantly higher than those feeding on the low-preference host plants. This supports the results of our previous study [22], indicating that H. cunea exhibits different preferences for the host plants. It is worth mentioning that the food consumption rate of H. cunea larvae in the low-preference host plant group was significantly higher than those in the high-preference host plant group. The food consumption rate is an important indicator that characterizes the food absorption capacity of phytophagous insects [25,28]. It indicates that H. cunea enhances the level of larval adaptation to the low-preference host plants by increasing the absorption capacity. However, inconsistent with the expected results, the α-amylase and trypsin activities in the midgut of H. cunea larvae feeding on the high-preference host plants were significantly lower than those feeding on the low-preference host plants. A possible reason is that H. cunea larvae are more easily able to digest and absorb the high-preference host plants than the low-preference host plants. Namely, it only needs relatively few digestive enzymes to efficiently absorb the nutrients from the high-preference host plants. Similarly, Chai et al. (2016) found that the amylase and cellulase activities in the gut of Agasicles hygrophila larvae were higher when feeding on the non-palatable host plants than those feeding on the palatable host plants [29]. A reverse trend was also reported by Shen et al. (2022), who found that among the five host plant groups, Dendrolimus houi larvae feeding on the high-preference host plants exhibited the highest digestive enzyme activity [30]. Our results, together with these previous findings, suggest that the response of digestive enzymes in phytophagous insects to the host plants with different preferences varies depending on the insect species or the content of endogenous inhibitors in these plants.
To further clarify the physiological function of digestive enzymes, we perturbed the digestion physiology of H. cunea larvae by spraying the inhibitors of digestive enzymes onto the leaf surface and then analyzed the adaptability of H. cunea to different host plants. Our results revealed that the body weight, food utilization status, several nutrients content, and α-amylase activity of H. cunea larvae upon AI treatment were significantly reduced. The function of α-amylase is to convert the starch into oligosaccharides, a precursor for glucose abortion [31,32,33]. These results suggest that the absence of α-amylase or functional impairment of α-amylase inhibits normal nutritional metabolism and attenuates the growth of H. cunea larvae. Trypsin is a key enzyme involved in the process of food protein hydrolysis [34]. The inhibition analysis revealed that the growth and digestive physiology trends of H. cunea larvae in the TI treatment group were consistent with those in the AI treatment group. It further indicates that the absence of trypsin may lead to inefficient uptake and utilization of nutrients (e.g., total protein) from the host plants by H. cunea, which ultimately leads to the growth retardation of H. cunea larvae. In addition to digestive physiology disorders, another possible reason for the reduced growth of H. cunea larvae caused by AI and TI is that inhibitor treatment reduces the palatability of plant leaves, as inhibitor-treated larvae in all host plant groups consume significantly less food. However, the food consumption rate increased following the addition of inhibitors across the board, irrespective of high and low preference towards host plants. This suggests once again that H. cunea larvae may be trying to adapt to adverse environmental conditions by improving their absorption capacity. This ability has also been reported in H. cunea larvae adaptation to other stressors, such as secondary metabolites and heavy metals stress [35,36,37]. Interestingly, treatment with digestive enzyme inhibitors attenuated differences in larval adaptation between high- and low-preference host plant groups, as shown by the fact that the AI and TI treatment reduced the difference in larval body weight from 65.77–135.17 mg to 38.66–74.14 mg between high and low feeding groups. A large number of studies have shown that compared with low-preference host plants, high-preference host plants have higher nutrient content and lower endogenous digestive enzyme inhibitor content [38,39,40]. After treatment with exogenous enzyme inhibitors, the H. cunea larvae could not effectively absorb nutrients from plants with high preference. This has led to a narrowing of the differences in the degree of adaptation. Another reason may be that digestive disorders lead to weakened digestive and taste nervous system functions, but further research is needed. Taken together, our results demonstrate that α-amylase and trypsin play an important role in nutrient metabolism and host adaptation of H. cunea.
Numerous studies have demonstrated that phytophagous insects are capable of regulating their digestive physiology to meet nutrient requirements [41,42,43]. This is a strategy used by insects to adapt to adverse conditions [44,45]. In the present study, the α-amylase activity in larval midgut was found to be significantly reduced in all AI-treated host plant groups, but the trypsin activity was significantly increased in the AI-treated JM and BP groups. Similar compensatory effects of digestive enzymes have been reported in other Lepidopteran insects. For example, Bezerra et al. (2017) found that Ephestia kuehniella exhibited a significant increase in chymotrypsin activity in vivo after treatment with TI [46]. Kuwar et al. (2019) also found that the trypsin activity of H. armigera larvae that were fed on soybean upon TI treatment was significantly increased [47]. These findings suggest that if the host plant has a high content of an insect digestive enzyme inhibitor, phytophagous insects can still regulate the activities of other digestive enzymes to compensate for the adverse effects of such inhibitors to promote the absorption and utilization of food nutrients [48]. Additionally, the nutrient content analysis revealed similar compensatory trends in the nutrient content of H. cunea larvae after treatment with AI or TI. For example, the total amino acids and free fatty acids contents were significantly higher in both AI- and TI-treated MA groups. It is further shown that H. cunea larvae have highly adaptable compensatory mechanisms for digestion, including digestive enzymes and nutrient metabolism, in response to unfavorable conditions. The insect digestive enzyme inhibitors secreted by plants are a common class of chemical defense compounds against phytophagous insects [47,49]. The compensatory effect exhibited by H. cunea to the digestive enzyme inhibitors may be implicated as a counter-defense strategy in response to their endogenous plant resistance factors. It is not difficult to conclude that the highly adaptable digestive physiological compensation effect may be the reason for the multi-host adaptability and host expansion of H. cunea larvae.
5. Conclusions
The digestive physiology of H. cunea plays an integral role in its adaptation to multiple host plants. The H. cunea larvae can improve larval food utilization and absorption efficiency on host plants by regulating the activity of larval digestive enzymes. This helps H. cunea maximize its adaptation to a variety of host plants and ensure its spread and host expansion. Additionally, H. cunea exhibits a highly adaptable digestive physiological compensation effect involving digestive enzymes and nutrient metabolism. This is implicated as a counter-defense strategy evolved by H. cunea in response to the endogenous resistance factors of host plants, especially the inhibitors of insect digestive enzymes. The findings of this study provide new insights into potential strategies to analyze the spread and host expansion of H. cunea.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/insects14050463/s1; Figure S1: Food intake of H. cunea larvae; Figure S2: Food conversion rate of H. cunea larvae; Table S1: Multiple comparisons; Table S2: Test of homogeneity of variances; Table S3: Independent samples test.
Author Contributions
Investigation, A.Z. and L.Y.; methodology, L.Y. and T.L.; validation, T.L. and M.T.; data curation, M.T.; visualization, L.Y., M.T. and T.L.; writing—original draft preparation, A.Z. and D.J.; writing—review and editing, S.Y. and D.J.; funding acquisition, S.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Project funded by the National Key R & D Program of China (2021YFD1400300).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare that they have no competing interest.
References
- Zhang, X.; Fan, Z.Z.; Zhang, R.; Kong, X.B.; Liu, F.; Fang, J.X.; Zhang, S.F.; Zhang, Z. Bacteria-mediated RNAi for managing fall webworm, Hyphantria cunea: Screening target genes and analyzing lethal effect. Pest Manag. Sci. 2023, 79, 1261–1628. [Google Scholar] [CrossRef] [PubMed]
- Trajkovic, A.; Zikic, V. Stuck in the caterpillars’ web: A half-century of biocontrol research and application on gregarious Lepidopteran pests in Europe. Sustainability 2023, 15, 2881. [Google Scholar] [CrossRef]
- Sullivan, G.T.; Karaca, I.; Ozman-Sullivan, S.K.; Kara, K. Tachinid (Diptera:Tachinidae) parasitoids of overwintered Hyphantria cunea (Drury) (Lepidoptera: Arctiidae) pupae in hazelnut plantations in Samsun province, Turkey. J. Entomol. Res. Soc. 2012, 14, 21–30. [Google Scholar]
- Ge, X.Z.; He, S.Y.; Zhu, C.Y.; Wang, T.; Xu, Z.C.; Zong, S.X. Projecting the current and future potential global distribution of Hyphantria cunea (Lepidoptera: Arctiidae) using CLIMEX. Pest. Manag. Sci. 2019, 75, 160–169. [Google Scholar] [CrossRef]
- Zhao, X.D.; Geng, Y.S.; Hu, T.Y.; Li, W.X.; Liang, Y.Y.; Hao, D.J. Comparing the performance of Hyphantria cunea (Lepidoptera: Arctiidae) on artificial and natural diets: Feasibility of mass-rearing on artificial diets. J. Econ. Entomol. 2023, 116, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Bai, P.H.; Wang, H.M.; Liu, B.S.; Li, M.; Liu, B.M.; Gu, X.S.; Tang, R. Botanical volatiles selection in mediating electrophysiological responses and reproductive behaviors for the fall webworm moth Hyphantria cunea. Front. Physiol. 2020, 11, 486. [Google Scholar] [CrossRef] [PubMed]
- Schowalter, T.D.; Ring, D.R. Biology and management of the fall webworm, Hyphantria cunea (Lepidoptera: Erebidae). J. Integr. Pest Manag. 2017, 8, 7. [Google Scholar] [CrossRef]
- Tang, X.G.; Yuan, Y.D.; Liu, X.F.; Zhang, J.C. Potential range expansion and niche shift of the invasive Hyphantria cunea between native and invasive countries. Ecol. Entomol. 2021, 46, 910–925. [Google Scholar] [CrossRef]
- Cao, L.J.; Wei, S.J.; Hoffmann, A.A.; Wen, J.B.; Chen, M. Rapid genetic structuring of populations of the invasive fall webworm in relation to spatial expansion and control campaigns. Divers. Distrib. 2016, 22, 1276–1287. [Google Scholar] [CrossRef]
- Kim, D.E.; Kil, J.H. A report on the occurrence of and crop damage caused by Hyphantria cunea (Drury) with in Korea. Korean J. Appl. Entomol. 2012, 53, 285–293. [Google Scholar] [CrossRef]
- Chen, Q.; Zhao, H.; Wen, M.; Li, J.; Zhou, H.; Wang, J.; Zhou, Y.; Liu, Y.; Du, L.; Kang, H.; et al. Genome of the webworm Hyphantria cunea unveils genetic adaptations supporting its rapid invasion and spread. BMC Genom. 2020, 21, 242. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.I.; Park, Y.S. Dispersal patterns of exotic forest pests in South Korea. Insect Sci. 2012, 19, 535–548. [Google Scholar] [CrossRef]
- Wang, W.X.; Zhou, L.F.; Dong, G.P.; Chen, F.M. Isolation and identification of entomopathogenic fungi and an evaluation of their actions against the larvae of the fall webworm, Hyphantria cunea (Drury) (Lepidoptera: Arctiidae). BioControl 2020, 65, 101–111. [Google Scholar] [CrossRef]
- Zhang, Z.J.; Zhang, S.S.; Niu, B.L.; Ji, D.F.; Liu, X.J.; Li, M.W.; Bai, H.; Palli, S.R.; Wang, C.Z.; Tan, A.J. A determining factor for insect feeding preference in the silkworm, Bombyx mori. PLoS Biol. 2019, 17, e3000162. [Google Scholar] [CrossRef]
- Jang, T.; Rho, M.S.; Koh, S.H.; Lee, K.P. Host-plant quality alters herbivore responses to temperature: A case study using the generalist Hyphantria cunea. Entomol. Exp. Appl. 2015, 154, 120–130. [Google Scholar] [CrossRef]
- Lazarevic, J.; Jankovic-Tomanic, M.; Savkovic, U.; Dordevic, M.; Milanovic, S.; Stojkovic, B. Host-associated divergence in the activity of digestive enzymes in two populations of the gypsy moth Lymantria dispar (Lepidoptera: Erebidae). J. Entomol. Sci. 2017, 20, 189–194. [Google Scholar] [CrossRef]
- Sarate, P.J.; Tamhane, V.A.; Kotkar, H.M.; Ratnakaran, N.; Susan, N.; Gupta, V.S.; Giri, A.P. Developmental and digestive flexibilities in the midgut of a polyphagous pest, the cotton bollworm, Helicoverpa armigera. J. Insect Sci. 2012, 12, 238–240. [Google Scholar] [CrossRef]
- Wu, N.N.; Zhang, S.F.; Li, X.W.; Cao, Y.H.; Liu, X.J.; Wang, Q.H.; Liu, Q.; Liu, H.H. Fall webworm genomes yield insights into rapid adaptation of invasive species. Nat. Ecol. Evol. 2019, 3, 105–115. [Google Scholar] [CrossRef]
- Li, L.S.; Yuan, Y.F.; Wu, L.; Chen, M. Effects of host plants on the feeding behavior and detoxification enzyme activities in Hyphantria cunea (Lepidoptera: Arctiidae) larvae. Acta Ecol. Sin. 2018, 61, 232–239. [Google Scholar]
- Mason, P.A.; Wilkes, S.R.; Lill, J.T.; Singer, M.S. Abundance trumps quality: Bi-trophic performance and parasitism risk fail to explain host use in the fall webworm. Oikos 2011, 120, 1509–1518. [Google Scholar] [CrossRef]
- Holtof, M.; Lenaerts, C.; Cullen, D.; Vanden Broeck, J. Extracellular nutrient digestion and absorption in the insect gut. Cell Tissue Res. 2019, 377, 397–414. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.M.; Fang, J.; Zhao, J.Q.; Wang, K.; Ren, Y.J.; Meng, Z.J.; Yan, S.C. Effects of 8 host young trees on growth, development and nutritional efficiency of Hyphantria cunea larvae. For. Res. 2023, 35, 124–131. [Google Scholar]
- Yan, S.C.; Tan, M.T.; Zheng, L.; Wu, H.F.; Wang, K.; Chai, R.S.; Jiang, D. Defense response of Fraxinus mandshurica seedlings to Hyphantria cunea larvae under Cd stress: A contradiction between attraction and resistance. Sci. Total Environ. 2023, 859, 160390. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.L.; Pan, Z.Y.; Meng, X. The Effect of quercetin on the growth, development, nutrition utilization, and detoxifification enzymes in Hyphantria cunea Drury (Lepidoptera: Arctiidae). Forests 2022, 13, 1945. [Google Scholar] [CrossRef]
- Jiang, D.; Dong, X.W.; Yan, S.C. Heavy metal accumulation/excretion in and food utilization of Lymantria dispar larvae fed with Zn- or Pb-stressed Populus alba berolinensis leaves. Environ. Entomol. 2018, 47, 1329–1336. [Google Scholar] [CrossRef]
- Terra, W.R.; Barroso, I.G.; Dias, R.O.; Ferreira, C. Molecular physiology of insect midgut. Adv. Insect Physiol. 2019, 56, 366–418. [Google Scholar]
- Chen, S.X.; Yang, F.; Fang, M.; Yao, L.; Zheng, R.W.; Tang, Q.F. Effect of host plants on the biology and digestive physiology of Spodoptera frugiperda. Interciencia 2022, 47, 328–334. [Google Scholar]
- Teimouri, N.; Sendi, J.J.; Zibaee, A.; Khosravi, R. Feeding indices and enzymatic activities of carob moth Ectomyelois ceratoniae (Zeller) (Lepidoptera: Pyrallidae) on two commercial pistachio cultivars and an artificial diet. J. Saudi Soc. Agric. Sci. 2015, 14, 76–82. [Google Scholar] [CrossRef]
- Chai, Y.P.; Zhang, J.Y.; Wang, Y.X.; Jia, D.; Ma, R.Y.; Guo, Y.Q. Effects of starvation and different plant feedings on the digestive enzyme activities in theadults of alligator weed flea beetle Agasicles hygrophila. J. Integr. Plant Biol. 2016, 43, 138–144. [Google Scholar]
- Shen, H.Y.; Liang, Y.; Chen, Z.H.; Li, C.L.; Zhang, Z.H.; Weng, H.F.; Huang, B.; Zhang, F.P.; Liang, G.H. Effects of different host plants on the growth, reproduction and physiological enzyme activity of Dendrolimus houi Lajonquiere (Lepidoptera:Lasiocampidae). For. Res. 2022, 35, 63–70. [Google Scholar]
- Ashouri, S.; Pourabad, R.F. Regulation of gene expression encoding the digestive alpha-amylase in the larvae of Colorado potato beetle, Leptinotarsa decemlineata (Say) in response to plant protein extracts. Gene 2021, 766, 145159. [Google Scholar] [CrossRef] [PubMed]
- Da Lage, J.-L. The Amylases of insects. Int. J. Insect Sci. 2018, 10, 1179543318804783. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.J.; Song, Y.; Jiang, X.M.; He, L.; Wei, L.Y.; Zhao, Z.W. Synergism of feeding and digestion regulated by the neuropeptide F system in Ostrinia furnacalis Larvae. Cells 2023, 12, 194. [Google Scholar] [CrossRef] [PubMed]
- Lazarevic, J.; Jankovic-Tomanic, M. Dietary and phylogenetic correlates of digestive trypsin activity in insect pests. Entomol. Exp. Appl. 2015, 157, 123–151. [Google Scholar] [CrossRef]
- Tan, M.T.; Wu, H.F.; Yan, S.C.; Jiang, D. Evaluating the toxic effects of tannic acid treatment on Hyphantria cunea larvae. Insects 2022, 13, 872. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Tan, M.T.; Yan, S.C.; Jiang, D. Cadmium exposure-triggered growth retardation in Hyphantria cunea larvae involves disturbances in food utilization and energy metabolism. Ecotoxicol. Environ. Saf. 2023, 256, 114886. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.S.; Zheng, L.; Tan, M.T.; Wu, H.F.; Yan, S.C.; Jiang, D. The susceptibility of Hyphantria cunea larvae to microbial pesticides Bacillus thuringiensis and Mamestra brassicae nuclear polyhedrosis virus under Cd stress. Pestic. Biochem. Physiol. 2023, 191, 105383. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.L.; Zhou, L.F.; Wang, Q.Y.; Ding, J.Q. Plant chemistry determines host preference and performance of an invasive insect. Front. Plant Sci. 2020, 11, 594663. [Google Scholar] [CrossRef]
- Vengateswari, G.; Arunthirumeni, M.; Shivaswamy, M.S.; Shivakumar, M.S. Effect of host plants nutrients, antioxidants, and phytochemicals on growth, development, and fecundity of Spodoptera litura (Fabricius) (Lepidoptera: Noctuidae). Int. J. Trop. Insect Sci. 2022, 42, 3161–3173. [Google Scholar] [CrossRef]
- Gou, Y.P.; Quandahor, P.; Zhang, Y.X.; Coulter, J.A.; Liu, C.Z. Host plant nutrient contents influence nutrient contents in Bradysia cellarum and Bradysia impatiens. PLoS ONE 2020, 15, e0226471. [Google Scholar] [CrossRef]
- Hafeez, M.; Li, X.W.; Zhang, J.M.; Zhang, Z.J.; Huang, J.; Wang, L.K.; Khan, M.M.; Shah, S.K.; Fernandez-Grandon, G.M.; Lu, Y.B. Role of digestive protease enzymes and related genes in host plant adaptation of a polyphagous pest, Spodoptera frugiperda. Insect Sci. 2021, 28, 611–626. [Google Scholar] [CrossRef] [PubMed]
- Babamir-Satehi, A.; Habibpour, B.; Aghdam, H.R.; Hemmati, S.A. Interaction between feeding efficiency and digestive physiology of the pink stem borer, Sesamia cretica Lederer (Lepidoptera: Noctuidae), and biochemical compounds of different sugarcane cultivars. Arthropod-Plant Interact. 2022, 16, 309–316. [Google Scholar] [CrossRef]
- Barzin, S.; Naseri, B.; Fathi, S.A.A.; Razmjou, J.; Aeinehchi, P. Feeding efficiency and digestive physiology of Trogoderma granarium Everts (Coleoptera: Dermestidae) on different rice cultivars. J. Stored Prod. Res. 2019, 84, 101511. [Google Scholar] [CrossRef]
- Mendonca, E.G.; Barros, R.D.; Cordeiro, G.; Silva, C.R.; Campos, W.G.; de Oliveira, J.A.; Oliveira, M.G.D. Larval development and proteolytic activity of Anticarsia gemmatalis Hubner (Lepidoptera: Noctuidae) exposed to different soybean protease inhibitors. Arch. Insect Biochem. Physiol. 2020, 103, e21637. [Google Scholar] [CrossRef] [PubMed]
- Juma, G.; Le, R.B.; Calatayud, P.A. Assortments of digestive enzymes induced in first instar larvae of Busseola fusca feeding on different plants. Int. J. Insect Sci. 2019, 11, 1179543319843521. [Google Scholar] [CrossRef]
- Bezerra, C.; Oliveira, C.T.; Macedo, M. Inga vera trypsin inhibitor interferes in the proteolytic activity and nutritional physiology of Ephestia kuehniella larvae. Entomol. Exp. Appl. 2017, 165, 109–119. [Google Scholar] [CrossRef]
- Kuwar, S.S.; Pauchet, Y.; Heckel, D.G. Effects of class-specific, synthetic, and natural proteinase inhibitors on life-history traits of the cotton bollworm Helicoverpa armigera. Arch. Insect Biochem. Physiol. 2020, 103, e21647. [Google Scholar] [CrossRef]
- Macedo, M.L.R.; de Oliveira, C.F.R.; Costa, P.M.; Castelhano, E.C.; Silva-Filho, M.C. Adaptive mechanisms of insect pests against plant protease inhibitors and future prospects related to crop protection: A review. Protein Pept. Lett. 2015, 22, 149–163. [Google Scholar] [CrossRef]
- Zhu-Salzman, K.; Zeng, R. Insect Responses to Plant Defensive Protease Inhibitors. Annu. Rev. Entomol. 2015, 60, 233–252. [Google Scholar] [CrossRef]
Figure 1.
The α-amylase activities of H. cunea larvae treated with AI (A) or TI (B). Data in the figure are presented as the mean ± SE (N = 3); lowercase letters indicate the difference between different host plant control groups (p < 0.05); * indicates the significant difference between the non-inhibited and AI treatment groups or between the non-inhibited and TI treatment groups for the same plants (p < 0.05).
Figure 1.
The α-amylase activities of H. cunea larvae treated with AI (A) or TI (B). Data in the figure are presented as the mean ± SE (N = 3); lowercase letters indicate the difference between different host plant control groups (p < 0.05); * indicates the significant difference between the non-inhibited and AI treatment groups or between the non-inhibited and TI treatment groups for the same plants (p < 0.05).

Figure 2.
The trypsin activities of H. cunea larvae treated with AI (A) or TI (B). Data in the figure are presented as the mean ± SE (N = 3); lowercase letters indicate the difference between different host plant control groups (p < 0.05); * indicates the significant difference between the non-inhibited and AI treatment groups or between the non-inhibited and TI treatment groups for the same plants (p < 0.05).
Figure 2.
The trypsin activities of H. cunea larvae treated with AI (A) or TI (B). Data in the figure are presented as the mean ± SE (N = 3); lowercase letters indicate the difference between different host plant control groups (p < 0.05); * indicates the significant difference between the non-inhibited and AI treatment groups or between the non-inhibited and TI treatment groups for the same plants (p < 0.05).

Figure 3.
Body weight of H. cunea larvae treated with AI (A) or TI (B). Note: Data in the figure are presented as the mean ± SE (N = 3); lowercase letters indicate the difference between different host plant control groups (p < 0.05); * indicates the significant difference between the non-inhibited and AI treatment groups or between the non-inhibited and TI treatment groups for the same plants (p < 0.05).
Figure 3.
Body weight of H. cunea larvae treated with AI (A) or TI (B). Note: Data in the figure are presented as the mean ± SE (N = 3); lowercase letters indicate the difference between different host plant control groups (p < 0.05); * indicates the significant difference between the non-inhibited and AI treatment groups or between the non-inhibited and TI treatment groups for the same plants (p < 0.05).

Figure 4.
Food utilization rate of H. cunea larvae treated with AI (A) or TI (B). Data in the figure are presented as the mean ± SE (N = 3); lowercase letters indicate the difference between different host plant control groups (p < 0.05); * indicates the significant difference between the non-inhibited and AI treatment groups or between the non-inhibited and TI treatment groups for the same plants (p < 0.05).
Figure 4.
Food utilization rate of H. cunea larvae treated with AI (A) or TI (B). Data in the figure are presented as the mean ± SE (N = 3); lowercase letters indicate the difference between different host plant control groups (p < 0.05); * indicates the significant difference between the non-inhibited and AI treatment groups or between the non-inhibited and TI treatment groups for the same plants (p < 0.05).

Figure 5.
Food consumption rate of H. cunea larvae treated with AI (A) or TI (B). Data in the figure are presented as the mean ± SE (N = 3); lowercase letters indicate the difference between different host plant control groups (p < 0.05); * indicates the significant difference between the non-inhibited and AI treatment groups or between the non-inhibited and TI treatment groups for the same plants (p < 0.05).
Figure 5.
Food consumption rate of H. cunea larvae treated with AI (A) or TI (B). Data in the figure are presented as the mean ± SE (N = 3); lowercase letters indicate the difference between different host plant control groups (p < 0.05); * indicates the significant difference between the non-inhibited and AI treatment groups or between the non-inhibited and TI treatment groups for the same plants (p < 0.05).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Nutrients content of H. cunea larvae.
| Nutrient Content | UP | FM | MA | JM | BP | AK | TA | |
|---|---|---|---|---|---|---|---|---|
| Total protein (mg/g) | CK | 10.53 ± 0.65 ab | 11.55 ± 0.41 ab | 10.02 ± 0.25 b | 12.11 ± 0.41 a | 7.37 ± 0.63 c | 7.66 ± 0.66 c | 7.08 ± 0.88 c |
| AI | 8.19 ± 0.18 * | 9.60 ± 0.47 * | 7.33 ± 0.61 * | 14.83 ± 0.20 * | 13.77 ± 1.24 * | 10.73 ± 0.27 * | 8.18 ± 0.53 | |
| TI | 7.70 ± 0.75 * | 9.12 ± 0.54 * | 8.08 ± 0.60 * | 17.05 ± 0.66 * | 14.02 ± 1.11 * | 11.32 ± 1.14 * | 8.63 ± 0.30 | |
| Total amino acids (μmol/mg prot) | CK | 1.92 ± 0.19 a | 1.90 ± 0.12 a | 1.80 ± 0.05 ab | 1.83 ± 0.13 ab | 1.52 ± 0.19 abc | 1.41 ± 0.13 c | 1.18 ± 0.05 c |
| AI | 2.31 ± 0.20 * | 1.20 ± 0.07 * | 2.00 ± 0.01 * | 1.09 ± 0.14 * | 0.85 ± 0.11 * | 0.96 ± 0.01 * | 1.02 ± 0.01 * | |
| TI | 1.94 ± 0.19 | 1.26 ± 0.14 * | 2.30 ± 0.04 * | 1.07 ± 0.02 * | 0.96 ± 0.07 * | 0.84 ± 0.10 * | 0.86 ± 0.03 * | |
| Glucose (μmol/g prot) | CK | 110.30 ± 3.41 a | 120.90 ± 11.36 a | 110.90 ± 3.31 a | 82.20 ± 15.37 b | 55.20 ± 1.56 b | 62.00 ± 6.61 b | 67.60 ± 3.80 b |
| AI | 40.70 ± 1.97 * | 85.50 ± 1.22 * | 88.90 ± 4.83 * | 48.10 ± 3.68 * | 27.80 ± 1.74 * | 41.70 ± 7.66 * | 37.70 ± 1.50 * | |
| TI | 62.40 ± 3.06 * | 76.80 ± 7.96 * | 81.95 ± 4.97 * | 43.50 ± 6.60 * | 36.90 ± 6.09 * | 52.60 ± 1.57 * | 53.90 ± 1.80 * | |
| Trehalose (mg/g tissue) | CK | 8.69 ± 0.30 a | 9.26 ± 0.29 b | 7.23 ± 0.46 c | 5.75 ± 0.13 d | 2.50 ± 0.21 e | 1.24 ± 0.11 f | 3.82 ± 0.41 g |
| AI | 2.43 ± 0.12 * | 4.50 ± 0.52 * | 1.81 ± 0.16 * | 3.60 ± 0.44 * | 0.74 ± 0.35 * | 0.53 ± 0.15 * | 0.60 ± 0.18 * | |
| TI | 2.48 ± 0.24 * | 4.21 ± 0.37 * | 3.50 ± 0.28 * | 3.34 ± 0.58 * | 0.85 ± 0.57 * | 0.38 ± 0.04 * | 0.78 ± 0.23 * | |
| Free fatty acids (μmol/g prot) | CK | 114.60 ± 11.28 a | 55.00 ± 3.40 b | 105.60 ± 7.98 a | 49.00 ± 11.30 b | 6.30 ± 1.28 c | 11.20 ± 1.60 c | 9.40 ± 2.00 c |
| AI | 152.90 ± 6.38 * | 79.40 ± 11.24 * | 223.90 ± 6.15 * | 80.20 ± 5.09 * | 1.70 ± 0.21 * | 2.60 ± 0.35 * | 3.10 ± 0.54 * | |
| TI | 157.20 ± 13.98 * | 79.20 ± 7.09 * | 269.80 ± 9.98 * | 84.10 ± 8.50 * | 2.80 ± 0.91 * | 2.80 ± 1.94 * | 3.10 ± 1.58 * |
Data in the table are presented as the mean ± SE (N = 3); lowercase letters indicate the difference between different host plant control groups (p < 0.05); * Indicates the significant difference between the non-inhibited and AI treatment groups or between the non-inhibited and TI treatment groups for the same nutrients in the same plants (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, A.; Li, T.; Yuan, L.; Tan, M.; Jiang, D.; Yan, S. Digestive Characteristics of Hyphantria cunea Larvae on Different Host Plants. Insects 2023, 14, 463. https://doi.org/10.3390/insects14050463
AMA Style
Zhang A, Li T, Yuan L, Tan M, Jiang D, Yan S. Digestive Characteristics of Hyphantria cunea Larvae on Different Host Plants. Insects. 2023; 14(5):463. https://doi.org/10.3390/insects14050463
Chicago/Turabian StyleZhang, Aoying, Tao Li, Lisha Yuan, Mingtao Tan, Dun Jiang, and Shanchun Yan. 2023. "Digestive Characteristics of Hyphantria cunea Larvae on Different Host Plants" Insects 14, no. 5: 463. https://doi.org/10.3390/insects14050463
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.