

The Structure of Rice Stemborer Assemblages: A Review of Species’ Distributions, Host Ranges, and Interspecific Interactions

1
EcoLaVerna Integral Restoration Ecology, Bridestown, Kildinan, T56 P499 Co. Cork, Ireland
2
Centre for Pesticide Suicide Prevention, University/BHF Centre for Cardiovascular Science, University of Edinburgh, Edinburgh EH16 4TJ, UK
3
Escuela de Agronomía, Facultad de Ciencias Agrarias y Forestales, Universidad Católica del Maule, Casilla 7-D, Curicó 3430000, Chile
Insects 2023, 14(12), 921; https://doi.org/10.3390/insects14120921
Submission received: 9 November 2023
/
Revised: 30 November 2023
/
Accepted: 30 November 2023
/
Published: 2 December 2023
(This article belongs to the Topic Arthropod Biodiversity: Ecological and Functional Aspects)
Abstract
:Simple Summary
Rice is attacked by range of stem-boring moths and flies. Despite their ecological and economic importance and the considerable scientific literature on their impacts and management, relatively little attention has been given to their coexistence mechanisms. This review lists the stemborer species associated with rice and determines their prevalence in rice crops based on published reports. The approximate distributions, host plant associations, modes of attacking rice, and details of the bionomics and behaviors of the economically important species are presented. Furthermore, using published data based on rice stem dissections, the varying structures of rice stemborer assemblages are described. Species richness is mainly determined by latitude, geography, and climate. Based on historical trends and information from stem dissections, possible mechanisms by which stemborers divide the rice crop, and thereby avoid competition, are determined. These include partitioning the resource by fidelity to rice and host plant range; by season and climate; by plant age, crop duration, and anatomy; and by the proximity and extent of alternative food resources (e.g., grasslands or other crops). Stemborer assemblages typically include a dominant primary moth species, one or more secondary species, and occasional species that are normally rare and probably spill over from adjacent grasslands. The dominance of primary species may change regionally, depending on environmental conditions including prevailing rice production systems.
Abstract
This review describes global rice stemborer assemblages based on published species distributions, apparent host preferences, and reported shifts in assemblage composition in response to environmental factors. At least 56 moth (Lepidoptera: Crambidae, Pyralidae, Noctuidae) and fly (Diptera: Diopsidae, Chloropidae) species have been associated with rice; however, only 21 species are of potential, large-scale economic importance with a further 2 species of localized concern; most of the remaining species’ associations with rice are based on dubious records without economic impacts on rice production. A list of stemborer–host associations indicates that rice stemborers are largely oligophagous on grasses (Poaceae), but a few species are polyphagous (also attacking Cyperaceae, Typhaceae, and some Eudicotyledon plants). Total stemborer abundance is determined by rice cropping patterns and management. Assemblage species richness is determined by geographical location, surrounding habitat (particularly as regards secondary and occasional species), and season. Evidence suggests that stemborer assemblage structure is largely determined through conditional interspecific competition. Regional assemblages typically include a single dominant lepidopteran species (primary species) that is largely restricted to rice and for which the climate is optimal; one or more secondary species that vary based on the age of rice attacked, rice anatomy, and the proximity to other habitats (including other crops); and occasional species that probably spill over from adjacent grasslands. The co-occurrence of lepidopteran with dipteran rice stemborers requires further research attention.
1. Introduction
Rice is produced on about 160 million ha globally, mainly in tropical and subtropical lowlands. It is the second most important cereal crop (after maize—Zea mays L.) and constitutes the largest terrestrial biome in much of tropical South and Southeast Asia [1]. The rice biome also dominates much of the lowland coastal regions of tropical and subtropical Africa and the Americas, as well as the river deltas of southern Europe [2,3]. Although this biome is dominated by domesticated rice (mainly Oryza sativa L., but also Oryza glaberrima Steud. in parts of Africa), a wide range of other cereals and grasses also occur in rice production landscapes [4,5]. This considerable resource (grasses and cereal crops) hosts a diversity of arthropods that are overwhelmingly beneficial to rice ecosystems as regulators of herbivore populations and of weed biomass, as pollinators of crops and wildflowers, or as food for livestock and wildlife [5,6]. However, a relatively small number of rice-associated arthropods are also considered economically challenging because they damage rice and potentially reduce yields [7].
Among the most damaging rice herbivores are the stemborers. These include the larvae of a number of moths and flies that attack rice by boring into the plant stem where they feed and develop [7,8,9]. Stemborers can damage or kill rice tillers, giving rise to characteristic, straw-like vegetative tillers known as deadhearts, or, during the reproductive stage, result in sterile or unfilled panicles known as whiteheads [10,11,12,13]. In most rice-producing regions, stemborer assemblages consist of several species, of which a small number are associated with the greatest damage [7,9]. However, the relative economic importance of species can change over time. For example, since its introduction in the 1930s, Chilo partellus (Swinhoe) has become a major pest of rice and other cereals in Africa and has been associated with the declining abundance of native African stemborers [14,15]; in the 1980s, Eoreuma loftini (Dyar) became a major pest of rice and other crops after range expansion in Mexico and into the southern USA [16,17,18], also affecting the abundance of other stemborers [19]. Assemblages of native stemborers can also sometimes shift in species’ dominance; for example, in Japan, Korea, and China, dominance shifted from Scirpophaga incertulas (Walker) to Chilo suppressalis (Walker) beginning in the 1960s [20,21]. In contrast, in Luzon (Philippines), S. incertulas increased in abundance after the 1970s while the abundance of C. suppressalis decreased [22]. In some parts of Malaysia, dominance shifted from Chilo polychrysus (Meyrick) to S. incertulas during the 1970s [23,24], and in parts of the Philippines from S. incertulas to Scirpophaga innotata (Walker) during the 1980s [22,25]. Furthermore, in North America, Chilo plejadellus Zincken gained abundance relative to Diatraea saccharalis (Fabricius) in Texas and Louisiana during the 1970s [26]; at about that same time, C. plejadellus abundance and damage increased on wild rice (Zizania spp.) in Minnesota (USA) [27].
It is still largely unknown why shifts in stemborer assemblage structure have occurred; however, some authors have implicated changing agronomic practices and the progressive adoption of new ‘types’ of rice variety (i.e., varieties that share a common phenology or common anatomical traits) over wide areas [20,28,29]. Shifting rice production practices have also been implicated in driving regional changes in relative damage to rice from established species. For example, throughout the 1980s, the abundance and damage from Maliarpha separatella Ragonot declined dramatically on Madagascar when farmers shifted from long-duration to short-duration rice varieties [30] and, in China, damage from both C. suppressalis and S. incertulas increased in the 1980s and 1990s, at the same time that a majority of farmers transitioned from inbred to hybrid rice varieties [31]. Some authors have implicated changing climatic conditions as determining or facilitating shifts in cereal stemborer assemblages [21,22,32], and recent evidence suggests that temperature can determine the outcome of interspecific competition between stemborer species on a common host [15]. These shifts in species abundance and dominance indicate that stemborer assemblages are structured and restructured through changes in the quality or availability of the rice resource relative to other host plant resources, and by conditional interspecific competition.
To better understand rice stemborer resource partitioning and possible coexistence mechanisms, this paper reviews current knowledge of rice stemborer assemblages. Using published records, it assesses whether assemblages exhibit any common patterns across regions in terms of species richness, and apparent assemblage structures. As an initial step, the host ranges of stemborers are assessed and, based on this appraisal, as well as on species’ apparent fidelity to rice, the stemborers are categorized as primary (often occurs as the dominant—most abundant—species in rice, and rice is the main host), secondary (usually subordinate in rice, i.e., rarely dominant; rice is often not the main host), and occasional rice herbivores (oligophagous and polyphagous species that sometimes occur in rice). The approximate distributions of the primary and secondary herbivores are then mapped and information on assemblage compositions in different regions is presented. Furthermore, together with behavioral, bionomic, and sampling data from dissected rice plants, possible resource partitioning mechanisms are identified. The review, therefore, clarifies the nature and extent of species interactions with practical implications for rice crop management and conservation biological control, and it outlines some preliminary rules of assembly for rice stemborers.
2. Review Methods
An initial search of the literature was conducted at the end of June 2023 using the terms ‘stemborer’ and ‘rice’ in Google Scholar. Google Scholar was used because it retrieves a wide variety of sources from the literature including peer-reviewed scientific papers, books and book chapters, national and regional reports, conference proceedings, and unpublished theses. This wide range of documents was used to map the approximate distributions of stemborer species. After the initial search, a series of further searches were conducted using the Latin names of rice stemborer species as listed by Khan et al. (1991) [9] and Pathak and Khan (1994) [7] together with the term ‘rice’. Furthermore, the taxonomy of the species was updated and any reports of new species described from rice were added to the list. Where possible, the original reports implicating each species as a rice herbivore were checked and the status of each species’ relations to rice were revised.
All reported field observations of the main rice stemborers species were assessed for taxonomic updates and were georeferenced. The referenced locations were then plotted and approximate species distributions traced by including all referenced points and other cited distributions. In some cases, detailed species distributions were already available [9]. Because there were a relatively large number of studies as well as available distribution data from Asia and the Americas, approximate distributions were traced without reference to national or regional boundaries (for Asia—distributions were overlaid on approximate rice distribution maps); however, many of the distribution records for African stemborers only list the species as present in specific countries; therefore, the mapped distributions for Africa were mainly based on country records for each species.
The literature on the potential hosts of the main rice stemborers was screened for host species records. The taxonomic information related to each species was updated when required. Information from the retrieved literature was used to assess the nature of interactions (mode of attack) between the stemborers and host plants, particularly rice, to determine possible coexistence mechanisms in rice fields. At the same time, information on species biometrics and behaviors were noted to determine further, possible niche- partitioning mechanisms. Because comparative biometric data were often incomplete across stemborer species, missing data on female minimum pupal weights and wingspans were estimated using linear regressions between female pupal length, pupal weight, and wingspan for those species for which complete data were available.
To limit the review text, much of the retrieved information is presented as a series of comprehensive Supplementary Tables. It is hoped that these will serve as a resource for further research into rice stemborer management. Because of the already extensive literature on stemborers—parts of which may have been overlooked during this review, and intense, recent research interest in these species [13], the tables should be continually revised and updated. Much of the information that appears in the Supplementary Tables is summarized in the main body of the review.
3. Results and Discussion
3.1. The Economically Important Rice Stemborers
Khan et al. (1991) [9] and Pathak and Khan (1994) [7] listed a total of 46 species of lepidopteran stemborers and 5 species of dipteran stemborers that attack rice globally. However, many of the listed species are only weakly associated with rice in the source documents because reports were often based on light-trapping or sweep-netting in rice habitats and not from the rearing of larvae collected from rice samples [9,33]. As such, many of these species might be more closely associated with wild grasses or other weeds that occur in the rice fields [34,35], or with other cereal crops [34], than they are with rice. Furthermore, the taxonomy of several of the listed species has since been revised—with species often split into two or more taxa [34,35,36,37], and none of the new taxa have yet been directly associated with rice. In the case of Diopsidae flies (henceforth diopsids), a number of species from the lists of Khan et al. (1991) [9] and Pathak and Khan (1994) [7] have been amalgamated; however, diopsid taxonomy remains understudied and there is still confusion as regards the key species attacking rice and their relative distributions [9,38,39]. A small number of species can now also be added to the original lists of Khan et al. (1991) [9] and Pathak and Khan (1994) [7]. These include moth and fly species that have since been reared from rice or have emerged as rice pests in localized areas, as well as a small number of reported species that were overlooked on the lists.
Based on the above criteria, Table 1 presents a revised list of 21 stemborers (with information on Sesamia nonagrioides (Lefèbvre) divided by subspecies) that are clearly associated with economic damage to rice plants over relatively large areas. A further 35 species with recent clarifications of status or with recent updates are included in Table S1. Of the 56 species listed in Table S1, there is a lack of clear evidence for rice herbivory among at least 24 of the species. Further information on rice stemborer assemblages, including species of localized interest, is presented in Section 3.3.
3.2. Stemborer Host Ranges
Several authors have listed the plants associated with stemborers (see Tables S2 and S3). However, there has been no systematic evaluation of host ranges across stemborer species and some of the more substantial host species lists have been derived from disparate studies, using a range of methods (e.g., oviposition experiments, rearing bioassays, field sampling, seed box tests, etc.). Further research on comparative host ranges is warranted. Based on the accumulated evidence presented in Table S2 and summarized in Table 2, all primary and secondary rice stemborers are at least oligophagous on grasses (Poaceae). A number of species have also been reported from monocotyledonous sedges and reeds (Table 2 and Table S2). Many stemborer species have been associated with one or more potential trap plants (Table S2). Trap plants are species that are used by stemborers for oviposition, but where larvae fail to develop to adults [140,141,142,143,144]. Some trap plants may be more attractive to stemborers than rice [145].
The more extensive host species lists have been compiled for the invasive species C. partellus, C. suppressalis, D. saccharalis, and E. loftini; and the major pest species S. incertulas, S. nonagrioides, and S. inferens (Table 2 and Table S2). However, these species have also received the greatest research attention (Table S3). The species D. saccharalis, E. lignosellus, S. incertulas, S. nonagrioides, S. calamistis, and S. inferens are associated with a diversity of hosts including a range of taxonomically distinct crops, sedges, and reeds, indicating that these species are somewhat polyphagous. A number of the secondary rice pest species from Table 1 are mainly associated with other crops including maize (e.g., S. nonagrioides, C. partellus, S. inferens), sugarcane (e.g., C. auricilius, S. nonagrioides botanephaga), sorghum (e.g., C. partellus), or with wild grasses (e.g., C. calamistis, C. polychrysus) (Table 1 and Table S1). Despite their ability to feed on other plants, the abundance of many of the primary rice stemborers is largely determined by the availability of rice [22,27,40,191].
3.3. Stemborer Distribution Ranges and Assemblage Structures
Species distributions and assemblage structures based on stem dissections are discussed by region in the following sections with distribution maps provided for the 21 species from Table 1.
3.3.1. Africa and the Mediterranean
A total of 29 stemborer species have been associated with rice in Africa; however, there are no recent records from rice for 16 of these species (Table S1); this includes Busseola fusca Fuller, a major pest of maize that was likely misidentified from rice in the original source information [34,192], and Eldana saccharina Walker, for which a direct association with rice appears doubtful [100]. A further four species (Ancylolomia chrysographellus Kollar and Redtenbacher, Adelpherupa costipunctalis Maes, Adelpherupa flavescens Hampson, and Chilo aleniellus Strand) have been reared from rice or associated with damage to rice, but without further, recent reports [9,35,139]. Of the remaining species, some, although widespread, damage rice in relatively localized regions: For example, S. nonagrioides that occurs around the Adriatic Sea and Persian Gulf, although it attacks rice in southern Europe, is more commonly associated with maize [40,52,193]. The species has also been reported from sugarcane in Iran apparently without attacking adjacent rice fields [194,195]. In West Africa, the subspecies Sesamia nonagrioides botanephaga Tams and Bowden is also more prevalent in maize and sugarcane [32,101,166]. Similarly, the introduced species C. partellus is a major pest of rice in some parts of Africa [38,48]; however, it is mainly a pest of maize and sorghum in its native range in Asia [196,197], although it occasionally damages Asian rice [118]. Figure 1 indicates the approximate distributions of the nine economically important rice stemborers in Africa, as well as the European and Central Asian distributions of invasive or naturalized C. suppressalis [198,199]. Based on distribution records, rice stemborer assemblages in tropical West Africa may include four or five lepidopteran stemborers and one or both of the main diopsid stemborers. In East Africa, assemblages can include four or five of the main species, including lepidopterans and diopsids (Figure 1).
Diopsids are prevalent stemborers in West African rice fields [56,58,59,96] and were the most abundant species in dissected rice stems in a study from Burkina Faso [96] (Figure 2A,B), but not in a study from Cameroon [200]. Among the lepidopteran stemborers, M. separatella and C. zacconius are often the most abundant species in lowland, rain-fed rice, with S. calamistis or S. diffusilineus largely occurring as secondary species [46,54,96,201] (Figure 2C–E). Meanwhile, in East Africa, M. separatella is often the dominant species [30,47,98]; but C. partellus is dominant at sites in Tanzania, with M. separatella, C. zacconius, or C. calamistis as secondary species [48,98] (Figure 2F). Diopsids appear less prevalent from East African studies, although they occur widely throughout the region [38] (Figure 1 and Figure 2).
The relative abundance of African stemborers is affected by season (Figure 2A,B) and habitat (Figure 2C–E), without affecting dominance (i.e., the rank abundance of species is maintained, particularly for the dominant species). January et al. (2021) [48] have shown that the abundance of S. calamistis relative to C. partellus increases with soil nitrogen levels and seeding density, and Leonard et al. (2015) [98] indicate varietal and sowing-date effects on the relative abundance of C. partellus, M. separatella, and S. calamistis in Tanzania—in both studies, this did not affect the dominance of C. partellus. Although not statistically significant, rice varieties and insecticide treatments also affected the relative abundance of lepidopteran stemborers in field plots in Nigeria, without affecting dominance [54].
3.3.2. Asia and Oceania
A total of 18 stemborer species have been associated with rice in Asia and Oceania (Table S1). Scirpophaga gilviberbis Zeller and A. chrysographellus, which have been associated with rice in Africa, also occur in Asia [33]. Of these 20 species, there are no direct records, or no recent records for 9 of the species feeding on rice. This includes a single report, from the 1950s, of Niphadoses palleucus Common attacking rice at a site in north Western Australia [122] and a single report of Saluria inficita (Walker) from rice in the Philippines [22]. Scirpophaga nivella (Fabricus) Lewvanich likely invaded New Caledonia and Fiji with the spread of rice [33]; however, there have been no recent reports of the species attacking rice in the Pacific region or in its native range in mainland Asia [9]. Seven species occur as primary or secondary species in rice (Table 1). Two further species are of localized economic importance (Table S1): Scirpophaga fusciflua Hampson occurs as a minor pest of rice in south India; however, in recent years it has been reported as the dominant species in rice in Himanchal Pradesh [123,128,205]. The Chloropidae fly Anatrichus erinaceus Loew has recently been associated with rice in Uttar Pradesh (India), where it caused up to 30% damage to rice in some localities [116].
Figure 1 indicates that tropical rice fields in Asia could include between three and six of the main stemborer species (see also Figure S1); however, dissections of rice stems indicate no more than five species in a given region: S. incertulas, C. suppressalis, and S. innotata are frequently the dominant species in Asian rice stemborer assemblages (Figure 2K–Y) [75,206]. Chilo polychrysus has also been reported as dominant at locations in Malaysia, particularly during older surveys (i.e., 1970s) [23,24,75,119,207] (Figure 2P). Stem dissections indicate that dominance is strongly influenced by habitat and region (Figure 2K–U), including altitude [208]. Furthermore, Zhu et al. (2002) [204] and Horgan et al. (2021) [73] indicate that this dominance can shift depending on season (Figure 2V,W) (see also Litsinger et al. (2011) [22]) and rice variety (Figure 2X,Y). Stem dissections suggest that assemblage composition is largely unaffected by crop establishment practices (drum-seeded, direct-seeded, transplanted, system of rice intensification—SRI [209]) and fertilizer inputs [72,73]. Furthermore, closely related hybrid, inbred, and sterile rice lines had little effect on stemborer assemblage structure in field plots in the Philippines [210]. The prevalence of Sesamia inferens (Walker) in dissected rice stems from north western India and the Punjab suggests that the abundance of this species is affected by cropping patterns including rice–wheat systems (Figure 2M,N) [29].
3.3.3. North America, South America, and the Caribbean
Relatively few stemborer species damage rice in the Americas, where only six species have been reported (Table S1). Of these, only Diatraea lineolata (Walker) has no recent records from rice (Table S1). This species mainly occurs in maize and sugarcane [19,129]. In the Americas, the maximum diversity of rice stemborers occurs in the southern USA, Mexico, Central America, and the northern parts of South America (three–four co-occurring species) (Figure 3). Elasmopalpus lignosellus (Zeller) is the most widespread species, occurring from the Great Lakes to central Chile (Figure 3). The species is highly polyphagous (Table 2 and Table S2) and damages upland rice seedlings in northern Brazil [93]. Chilo plejadellus is restricted to the eastern USA and south eastern Canada. Around the Great Lakes, it damages wild rice (Zizania spp.) [27]. Although C. plejadellus occurs in low numbers, it was often the predominant species in Louisiana rice fields during the 1970s [26,79] when it increased in abundance relative to D. saccharalis, possibly in response to the adoption of relatively thick-stemmed rice varieties [78,79]. Diatraea saccharalis is widespread in the tropical and subtropical Americas, extending from Florida and the Gulf of Mexico (Texas, Louisiana) to southern Brazil and Uruguay. Two species with a more restricted distribution are Rupela albinella (Cramer), which occurs in Central America and tropical South America (Figure 3) as the dominant species in rice (Figure 2G,H), and E. loftini, which occurs mainly in Mexico, but has recently extended its range into the southern USA [16,17,19,211,212]. Eoreuma loftini is now the dominant species in Louisiana and Texas rice fields [86,87], largely displacing D. lineolata and D. saccharalis in rice and other crops in its expanded range [19] (Figure 2I,J).
3.4. Stemborer Abundance and Species Richness
Notable shifts in the relative abundance of rice stemborer species over time suggest that their assemblages are ultimately structured through interspecific competition. Large scale changes in agriculture (e.g., crop diversity and distribution, crop rotations, pesticide and fertilizer inputs, irrigation regimes, land clearing) [48,213,214,215,216,217,218,219,220] and rice cropping practices (e.g., extent of rice production, production intensity, crop duration, synchronization of cropping) affect resource availability, setting the carrying capacity of the environment for stemborers and influencing total stemborer abundance and consequent damage [22,27,30,31,33,40,191,221,222]. Some of these changes apparently favor some species more than others (see Section 3.5).
Regionally, the potential species richness of rice stemborer assemblages is determined by latitude, altitude, and associated habitats, and, for islands, by the proximity to a continental mainland (Figure 1, Figure 3 and Figure S1) [32,62,67,223,224]. In-field stemborer richness will be strongly influenced by the occurrence, at low densities, of occasional stem-boring species, many of which are oligophagous grass-feeders—including species that predominantly occur in other cereal crops, or polyphagous species that often occur in upland rice or in relatively dry rice production environments (Table 1 and Table S1). For example, when occasional species are included, rice in some parts of tropical Asia could be affected by up to nine stemborer species (Figure S1). Many of the occasional species have no negative economic impacts on rice production; however, some may have beneficial functions by reducing weed biomass or as alternative hosts of the important natural enemies in rice production systems [182,223,225,226]. It is also possible that some of these occasional rice-feeding stemborers could become significant rice pests in specific rice production systems or if introduced to new regions, as likely occurred with C. partellus in Africa or S. fusciflua and A. erinaceus in India [14,116,123,128,150]. However, the apparent rarity of occasional species, particularly in recent African studies, might also point to a general decline in biodiversity associated with cereal crop expansion and grassland clearing.
Stemborer species richness in rice is also strongly affected by the spread of crop-associated invasive species including C. partellus, C. suppressalis, S. innotata, S. nivella, D. saccharalis, and E. loftini [18,33,64,185,191,198,199,212,227,228,229,230]. Many of these species caused shifts in assemblage structures in parts of their expanded ranges, often becoming the dominant species in rice (e.g., Figure 2F,I,J). This may be due to a release from natural enemies or to other competitive advantages over native rice stemborer species.
3.5. Resource Partitioning and Potential Coexistence Mechanisms
Shifts in species dominance over time and in response to large-scale changes to rice production practices reveal possible assemblage structuring mechanisms [14,19,20,22]. Stem dissections from field experiments, although relatively uncommon, offer further evidence of resource partitioning (Figure 2), particularly where these can be linked to comparative reports on stemborer bionomics (Table S4) and behaviors (vis-à-vis modes of attack: Table 1). However, stemborer assemblages are often characterized by the overwhelming dominance of a single species capable of ‘displacing’ potential competitors [14,19,22]. Despite such dominance, apparently weaker competitors, with very similar modes of attack, still occur at low or moderate densities in the rice crop (Figure 2), sometimes even in the same rice plants [22,73]. This suggests that interspecific competition is seldom sufficient to exclude competitors—despite all stemborer species occurring inside the stem lumen. This paradox may be explained by a high level of aggression, intraguild predation, and cannibalism [71,73,178] that reduce intraspecific competition and limit interspecific encounters inside the rice plant [231]; and by the intraspecific aggregation of egg masses and larvae in the rice crop [134,232,233]
Oligophagy plays a large role in stemborer species’ coexistence (Figure 4). Crop plants offer a highly favorable resource for most herbivores because of their high nutrient content (due to fertilizers), their relatively consistent access to water, and often low anti-herbivore defenses [72,73]. Oligophagy allows the survival of a number of stemborer species during periods when rice is unavailable [41,66,234]. It may also allow weaker competitors to maintain populations in the rice landscape during periods when some superior competitor is most abundant [235]. For example, the recent invasions of Fall Armyworm, Spodoptera frugiperda (J. E. Smith) in Africa and Asia, have been associated with a declining abundance of native stemborers in maize [236,237,238,239,240,241], even though S. frugiperda is not a stemborer, but a leaf chewer. This is partly due to predation of neonate stemborers before they enter the stem by S. frugiperda at high post-introduction densities—as shown in laboratory experiments [238,240,241]. Furthermore, S. frugiperda larvae have been show to induce maize defenses that subsequently reduce stemborer fitness (i.e., plant-mediated interspecific competition [238]). These interactions are predicted to shift maize stemborers from maize to relatively enemy-free or competitor-free crops such as sorghum [237,239]. Proximity to weedy grasslands or alternative crop hosts have also been suggested to influence the relative abundance of stemborers in rice, including determining the dominance of species in the assemblage [23,56,58,98,179]. However, this hypothesis is difficult to test. Although many of the primary rice stemborers are oligophagous, they occur predominantly in rice and their abundance is determined by the presence and extent of rice production [22,40,191]. Such primary pests are likely the best adapted to rice and, therefore, the strongest competitors for the rice resource over a wide range of environments. Therefore, whereas oligophagy maintains secondary and occasional stemborers in the rice environment, other partitioning mechanisms likely play a greater role in the co-occurrence of the primary species.
Evidence from Asia indicates that shifts in the dominance of the primary rice stemborers depend on production systems (e.g., the availability of standing water in the system) and climate (Figure 4). For example, seasonal and regional shifts in the dominance of S. incertulas, C. suppressalis, and S. innotata in the Philippines [25,204] suggest increasing advantages for the latter two species because of relatively dryer environments. Indeed, S. innotata can undergo prolonged diapause during drought conditions [25]. Litsinger et al. (2011) [22] suggest that S. innotata largely displaced S. incertulas and C. suppressalis in the southern Philippines after a particularly dry El Niño period. Similarly, C. polychrysus and S. inferens occur in deepwater systems during dryer periods of production and are displaced by S. incertulas when the systems are flooded [29,69,71]. In Africa, diopsid flies and S. calamistis are often abundant in wet, lowland environments [38,56,58,59,60]. Regional differences in weather and climate have also been implicated in the coexistence of S. nonagrioides botanephaga and S. calamistis in Côte d’Ivoire, with the former species occurring predominantly in wet coastal zones and the latter in upland savannas [32]. Similarly, C. diffusilineus occurs more in lowland rice fields with C. zacconius in the highlands [242].
The influence of weather and climate on the distribution and abundance of a number of rice stemborers has been studied using climate models and species’ temperature profiles [21,243,244,245]. Based on >50 years of historical weather data from southern China, Shi et al. (2012) [21] suggest that a declining abundance of S. incertulas could be attributed to increasing global temperatures. This hypothesis is consistent with the affinity of S. incertulas for wetter environments; however, the study did not include the possibility for displacement of the species by other stemborers, for which increasing temperatures might represent an advantage. For example, in experiments that manipulated stemborers under a range of temperature regimes, S. calamistis and B. fusca performed relatively poorly in the presence of C. partellus on maize plants and in ‘artificial stems’ under relatively high temperatures (25–30 °C) compared to lower temperatures [15,246], thereby, further indicating how interspecific competition conditioned by climate might determine relative abundance.
The time of maximum occurrence in the crop often differs among co-existing stemborer species (Figure 4). Maximum occurrence depends on oviposition preferences, larval dispersal behaviors, and intra- or interspecific aggression. Most species occur during the vegetative stages of crop development and some have poor fitness on older plants when the stems are tough and the carbon to nitrogen ratios increase. Among the earliest colonizers of rice fields are S. incertulas, S. innotata, and the diopsids [22,59] (Figure 4). Diopsids lay eggs individually over many days, in contrast to the lepidopterans that produce egg masses each with 10s or 100s of eggs (Table S4) [59]. Furthermore, the diopsids may kill rice leaves by feeding on meristems, without causing stem death [38] (Table 1). Therefore, it is unclear whether diopsids directly compete with lepidopteran stemborers. Leonard et al. (2015) [98] suggests that C. partellus can dominate rice stemborer assemblages in Tanzania because it occurs earlier in rice and has faster development. Indeed, recent experiments have shown that C. partellus performs relatively well in the presence of other stemborers and S. frugiperda because of its relatively fast growth rate and, consequently, limited exposure to competitors [236,246,247,248]. Elasmopalpus lignosellus is capable of damaging pre-tillering seedlings in upland rice systems [93]. Late colonizers include S. nonagrioides, S. calamistis, and S. inferens [38,41,214,224,249] (Figure 4). A recent study has shown that volatiles can determine the attractiveness of the host plant for ovipositing stemborers; however, in experiments with S. calamistis and C. partellus, the induced volatiles tended to increase the attractiveness of maize, thereby functioning as a plant-mediated facilitator of host attack [250]—and, therefore, likely increases interspecific aggregation. It is possible that similarly induced volatiles might also function to draw-in natural enemies, such as egg parasitoids [251,252,253]. Environmental effects on rice development can further influence stemborer assemblages; for example, upland rice has thicker stems at early vegetative stages that likely reduces its suitability for diopsids [60], and nitrogenous fertilizers prolong rice maturation and increase stem thickness, thereby favoring relatively late colonizers and larger species (see below) [72,73,254].
Stemborers also differ in the locations they occupy in rice stems and the number of stems they occupy during infestation; for example, C. suppressalis occupies higher internodes than S. incertulas and S. inferens, both of which occur predominantly at the base of the stems (Table 1). Rice anatomy interacts with stemborer bionomics and life histories to determine these features of colonization. For example, the rice lumen must be sufficiently wide to accommodate stemborer development, suggesting that stemborers may be restricted by stem thickness based on the minimum size of female larvae or pupae. Because stem thickness increases with plant age, smaller stemborer species are predicted to occur earlier during crop development, or have a more rapid development than relatively larger species. Some (i.e., S. inferens, S. nonagrioides), but not all (i.e., S. calamistis), late colonizers are relatively large (Figure 5). Shifts in dominance between D. saccharalis and C. plejadellus in Louisiana [78,79] and between S. incertulas and S. suppressalis in Asia [22,31] have been attributed to the large-scale planting of varieties with relatively thick stems. Oliver et al. 1975 [78] found that pupae of C. plejadellus were often larger than D. saccharalis; furthermore, S. suppressalis are often heavier than S. incertulas [72,73]; however, in general, evidence to support the hypothesis for changing species’ dominance related to stem thickness is weak. Horgan et al. (2021) [73] also suggested that C. suppressalis and S. incertulas likely segregate on the basis of rice-tillering patterns, with the former more abundant on low tillering varieties, which also often have thicker stems. However, no such patterns were observed among stemborers on fertile and sterile rice, despite profuse tillering of the sterile rice [210]. This suggests that a combination of factors, including tillering, stem thickness, and others, might determine species’ preferences.
Authors have indicated that a declining abundance of M. separatella in Madagascar [30], C. polychrysus in Malaysia [24], and S. innotata in Australia [22] coincided with the planting of short-duration varieties. This suggests that the species failed to complete their final generation in the rice, effectively functioning as an ecological trap. It is difficult to link crop duration with stemborer development times because systematic knowledge of rice growth and larval development times under varying temperatures is still not available (Table S4) and competition between rice herbivores across a range of temperatures can be influenced by the temperature optima of the rice varieties on which they develop [280,281]. Short duration varieties are often part of double- or triple-cropping systems, which might increase the abundance of those species with relatively short larval periods and rapid development [31]. However, this hypothesis remains untested.
4. Conclusions
Despite the extent of rice landscapes globally, only a few stemborer species (e.g., the moths C. agamemnon, C. partellus, C. suppressalis, C. zacconius, D. sacharralis, M. separatella, S. incertulas, S. innotata, and the fly species D. macrophthalma: Table 1) are of economic importance in rice production. These species can be divided into three or four principal groups, depending on whether diopsids are included as part of the same assemblage. Among lepidopteran stemborers, assemblages are structured by the presence of a single highly dominant species that is mainly adapted to feed on rice, for which the cropping system (including water availability and temperatures) is optimal and crop duration is sufficiently long to allow full larval development. Assemblages may or may not include one or more oligophagous or polyphagous secondary species that partition the resource by age, anatomy (e.g., stem thickness, tiller number), or proximity to key habitats and crops (e.g., S. inferens in wheat–rice systems). Finally, assemblages may include occasional oligophagous or polyphagous species that are of little economic importance to rice production and likely represent a spillover from native grasslands or other habitats. Assemblage structure is influenced by rice crop management. The abundance of stemborers is affected by rice cropping area, cropping intensity, fertilizer use, and crop synchronization, but species dominance appears resilient against changes in these production factors, although the relative abundance of species may be affected. A number of invasive species dominate stemborer assemblages in some regions. This is likely due to a release from natural enemies or other competitors after introduction. Much of our current understanding of stemborer assemblage structuring is based on empirical evidence. There is a need for further, process-oriented research to address several current hypotheses.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/insects14120921/s1, Table S1: List of stemborers species associated with rice; Table S2: Stemborer–host plant associations; Table S3: Data sources for Table S2; Table S4: Bionomic data for key rice stemborers; Figure S1: Species richness of stemborer assemblages attacking rice in Asia. The green area indicates the main rice growing regions of Asia, including around the Caspian Sea. Species richness is based on distributions presented in Figure 1 and in the source information cited in Table S1.
Funding
This research received no external funding.
Data Availability Statement
No new data were created; tabulated data from existing publications are included, together with source references, in the Supplementary Materials.
Conflicts of Interest
The author declares no conflict of interest.
References
- Seck, P.A.; Diagne, A.; Mohanty, S.; Wopereis, M.C.S. Crops that feed the world 7: Rice. Food Secur. 2012, 4, 7–24. [Google Scholar] [CrossRef]
- Van Nguyen, N.; Ferrero, A. Meeting the challenges of global rice production. Paddy Water Environ. 2006, 4, 1–9. [Google Scholar] [CrossRef]
- Ellis, E.C.; Ramankutty, N. Putting people in the map: Anthropogenic biomes of the world. Front. Ecol. Environ. 2008, 6, 439–447. [Google Scholar] [CrossRef]
- Fried, O.; Westphal, C.; Schellenberg, J.; Grescho, V.; Kühn, I.; Van Sinh, N.; Settele, J.; Bergmeier, E. Vascular plant species diversity in Southeast Asian rice ecosystems is determined by climate and soil conditions as well as the proximity of non-paddy habitats. Agric. Ecosyst. Environ. 2021, 314, 107346. [Google Scholar] [CrossRef]
- Bambaradeniya, C.N.B.; Edirisinghe, J.P.; De Silva, D.N.; Gunatilleke, C.V.S.; Ranawana, K.B.; Wijekoon, S. Biodiversity associated with an irrigated rice agro-ecosystem in Sri Lanka. Biodivers. Conserv. 2004, 13, 1715–1753. [Google Scholar] [CrossRef]
- Lupi, D.; Rocco, A.; Rossaro, B. Benthic macroinvertebrates in Italian rice fields. J. Limnol. 2013, 72, 184–200. [Google Scholar] [CrossRef]
- Pathak, M.D.; Khan, Z.R. Insect Pests of Rice; International Rice Research Institute: Los Baños, Philippines, 1994. [Google Scholar]
- Heinrichs, E.A. Biology and Management of Rice Insects; International Rice Research Institute: Los Baños, Philippines, 1994. [Google Scholar]
- Khan, Z.R.; Litsinger, J.A.; Barrion, A.T.; Villanueva, F.F.D.; Fernandez, N.J.; Taylo, L.D. World Bibliography of Rice Stem Borers 1794–1990; International Rice Research Institute: Los Baños, Philippines, 1991. [Google Scholar]
- Ingram, W. The lepidopterous stalk borers associated with Gramineae in Uganda. Bull. Entomol. Res. 1958, 49, 367–383. [Google Scholar] [CrossRef]
- Oñate, B.T. Estimation of stem borer damage in rice fields. Philipp. Stat. 1965, 14, 201–221. [Google Scholar]
- Otake, A. An analytical study on the injury to rice plants by the rice stem borer, Chilo suppressalis Walker (Lepidopter: Pyralidae) in its first generation. Appl. Entomol. Zool. 1967, 2, 173–186. [Google Scholar] [CrossRef]
- Horgan, F.G. Revisiting research and methods in stemborer-rice interactions for integration into future breeding programs. CABI Rev. 2024, in press. [Google Scholar]
- Kfir, R. Competitive displacement of Busseola fusca (Lepidoptera: Noctuidae) by Chilo partellus (Lepidoptera: Pyralidae). Ann. Entomol. Soc. Am. 1997, 90, 619–624. [Google Scholar] [CrossRef]
- Ntiri, E.S.; Calatayud, P.-A.; Van Den Berg, J.; Schulthess, F.; Le Ru, B.P. Influence of temperature on intra- and interspecific resource utilization within a community of lepidopteran maize stemborers. PLoS ONE 2016, 11, e0148735. [Google Scholar] [CrossRef]
- Johnson, K.J.R. Identification of Eoreuma loftini (Dyar) (Lepidoptera: Pyralidae) in Texas, 1980: Forerunner for other sugarcane boring pest immigrants from Mexico? Bull. Entomol. Soc. Am. 1984, 30, 47–52. [Google Scholar] [CrossRef]
- Rodríguez-del-Bosque, L.; Pantaleón-Paulino, G. Eoreuma loftini (Lepidoptera: Crambidae) expanded its geographic range into southern Veracruz and northern Oaxaca, México. Southwest. Entomol. 2012, 37, 525–528. [Google Scholar] [CrossRef]
- Wilson, B.E.; Beuzelin, J.M.; Reagan, T.E. Population distribution and range expansion of the invasive Mexican Rice Borer (Lepidoptera: Crambidae) in Louisiana. Environ. Entomol. 2017, 46, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-del-Bosque, L.; Reyes-Méndez, C. Eoreuma loftini displaced Diatraea lineolata and D. saccharalis (Lepidoptera: Crambidae) as the main corn stalkborer in northern Tamaulipas, México. Southwest. Entomol. 2013, 38, 75–78. [Google Scholar] [CrossRef]
- Kiritani, K. Recent population trends of Chilo suppressalis in temperate and sub-tropical Asia. Int. J. Trop. Insect Sci. 1990, 11, 555–562. [Google Scholar] [CrossRef]
- Shi, P.J.; Zhong, L.; Sandhu, H.S.; Ge, F.; Xu, X.M.; Chen, W. Population decrease of Scirpophaga incertulas Walker (Lepidoptera Pyralidae) under climate warming. Ecol. Evol. 2012, 2, 58–64. [Google Scholar] [CrossRef]
- Litsinger, J.A.; Barrion, A.T.; Canapi, B.L.; Lumaban, M.D.; dela Cruz, C.G.; Pantua, P.C. Philippines rice stemborers: A review. Philipp. Ent. 2011, 25, 1–47. [Google Scholar]
- Khari, N.A.M.; Ab Hamid, S. Abundance and infestation of Rice Stem Borer in North Malaysia. Malays. Appl. Biol. 2022, 51, 165–177. [Google Scholar] [CrossRef]
- Khoo, S. Pest outbreaks in the tropics. J. Plant Prot. Trop. 1986, 3, 13–24. [Google Scholar]
- Litsinger, J.A.; Alviola, A.L.; Dela Cruz, C.G.; Canapi, B.L.; Batay-An, E.H.; Barrion, A.T. Rice white stemborer Scirpophaga innotata (Walker) in southern Mindanao, Philippines. I. Supplantation of yellow stemborer S. incertulas (Walker) and pest status. Int. J. Pest Manag. 2006, 52, 11–21. [Google Scholar] [CrossRef]
- Oliver, B.; Gifford, J.; Trahan, G. Differential infestation of rice lines by the rice stalk borer. J. Econ. Entomol. 1972, 65, 711–713. [Google Scholar] [CrossRef]
- Peterson, A.G.; Noetzel, D.M.; Sargent, J.E.; Hanson, P.E.; Johnson, C.B.; Soemawinata, A.T. Insects of Wild Rice in Minnesota; University of Minnesota: Minneapolis, MN, USA, 1981. [Google Scholar]
- Luo, G.-H.; Luo, Z.-X.; Zhang, Z.-L.; Sun, Y.; Lu, M.-H.; Shu, Z.-L.; Tian, Z.-H.; Hoffmann, A.A.; Fang, J.-C. The response to flooding of two overwintering rice stem borers likely accounts for their changing impacts. J. Pest Sci. 2021, 94, 451–461. [Google Scholar] [CrossRef]
- Singh, R.A.; Singh, C.V.; Singh, G. Drought-induced shifting of stem borer species in shallow deepwater rice. Int. Rice Res. Notes 2005, 30, 1. [Google Scholar]
- Bianchi, G.; Rasoloarison, B.; Genini, M. Noxiousness of the African white stem borer Maliarpha separatella Rag (Pyralidae, Phycitinae) in irrigated paddy fields at Lake Alaotra (Madagascar). Insect Sci. Its Appl. 1993, 14, 667–673. [Google Scholar] [CrossRef]
- Horgan, F.G.; Crisol, E. Hybrid rice and insect herbivores in Asia. Entomol. Exp. Et Appl. 2013, 148, 1–19. [Google Scholar] [CrossRef]
- Moyal, P. Why Sesamia nonagrioides (Lefèbvre)(Lepidoptera: Noctuidae) Is a Pest Only in Portions of Its Geographic Range: The Influence of Climate. Available online: https://hal.science/hal-01087819 (accessed on 15 June 2023).
- Lewvanich, A. A Revision of the Old World Species of the Scirpophaga complex (Lepidoptera: Pyralidae): A Taxonomic and Zoogeographic Study, with a Discussion on Their Affinities; University of London: London, UK, 1971. [Google Scholar]
- Calatayud, P.-A.; Le Ru, B.P.; Van den Berg, J.; Schulthess, F. Ecology of the African Maize Stalk Borer, Busseola fusca (Lepidoptera: Noctuidae) with special reference to insect-plant interactions. Insects 2014, 5, 539–563. [Google Scholar] [CrossRef]
- Maes, K.V.N. Revision of the genus Adelpherupa Hampson (Lepidoptera: Pyraloidea, Crambidae, Schoenobiinae) with the description of five new species. J. Nat. Hist. 2002, 36, 1707–1724. [Google Scholar] [CrossRef]
- Kergoat, G.J.; Toussaint, E.F.A.; Capdevielle-Dulac, C.; Clamens, A.-L.; Ong’amo, G.; Conlong, D.; van Den Berg, J.; Cugala, D.; Pallangyo, B.; Mubenga, O.; et al. Integrative taxonomy reveals six new species related to the Mediterranean Corn Stalk Borer Sesamia nonagrioides (Lefèbvre) (Lepidoptera, Noctuidae, Sesamiina). Zool. J. Linn. Soc. 2015, 175, 244–270. [Google Scholar] [CrossRef]
- Moyal, P.; Le RÜ, B.; Van Den Berg, J.; Ratnadass, A.; Cugala, D.; Matama-Kauma, T.; Pallangyo, B.; Conlong, D.; Defabachew, B. Morphological reinforcement, ancient introgressive hybridization and species delimitation in African stem-borer species of the genus Sesamia Guenée (Lepidoptera: Noctuidae). Syst. Entomol. 2011, 36, 421–434. [Google Scholar] [CrossRef]
- January, B.; Rwegasira, G.M.; Tefera, T. Rice stem borer species in Tanzania: A review. J. Basic Appl. Zool. 2020, 81, 1–9. [Google Scholar] [CrossRef]
- Joshi, R.C.; Ukwungwu, M.N. Stalk-eyed fly (SEF) damage to lowland irrigated rices in Nigeria. Int. Rice Res. Newsl. 1992, 17, 25–26. [Google Scholar]
- Melamed-Madjar, V. Status of Chilo agamemnon Bles. in Israel and the probable reasons for the decrease in its populations. Int. J. Trop. Insect Sci. 1990, 11, 541–545. [Google Scholar] [CrossRef]
- Metwally, M. Ecological studies on lepidopterous stem borers associated with the main graminaceous weeds and crops at kafr El-Sheikh region. J. Plant Prot. Pathol. 2010, 1, 319–330. [Google Scholar] [CrossRef]
- Soliman, A.M.; Hendawy, A.S.; El-Hefny, A.S.; Sherif, M.R. Determination of economic threshold level for Chilo agamemnon Bles. infestation in rice plants, based on simulated dead hearts. Egypt. J. Agric. Res. 2016, 94, 59–69. [Google Scholar] [CrossRef]
- Zaazou, H.M.; El-Nahal, A.K.M.; Bishara, M.A. The effect of certain cultural practices on the infestation of rice by Chilo agamemnon Bles. and Tryporyza incertulas Wlk. (Lepidoptera: Pyralidae). Bull. Soc. Entomol. D’Egypte 1970, 54, 149–154. [Google Scholar]
- Saleh, H.; Bleih, E.M. Influence of sowing dates and nitrogen fertilization levels on rice yield and insect infestation of Sakha super 300 rice cultivar. Egypt. Acad. J. Biol. Sci. H. Bot. 2021, 12, 75–86. [Google Scholar]
- Vercambre, B.; Bordat, D.; Djiba, S. Occurrence and distinction of the two principal species of Chilo in rice, in Casamance, Senegal. Agron. Trop. 1990, 45, 131–138. [Google Scholar]
- Djiba, S. Le devenir de Chilo spp. (Lepido., Pyralidae) en saison sèche dans la région de Casamance. Sahel IPM 1995, 2, 18–21. [Google Scholar]
- Ho, D.T.; Njokah, J.J.; Kibuka, J.G. Studies on rice stemborers in Kenya with emphasis on Maliarpha separatella Rag. Insect Sci. Its Appl. 1983, 4, 65–73. [Google Scholar] [CrossRef]
- January, B.; Rwegasira, G.M.; Tefera, T. Impacts of plant spacing and nitrogen fertiliser on incidences and density of Spotted and African Pink Stem Borers in Tanzania. Int. J. Pest Manag. 2021, 67, 241–251. [Google Scholar] [CrossRef]
- Adewoye, B.B.; Anikwe, J.C.; Makanjuola, W.A. Incidence and relative abundance of rice stem borers in three selected rice fields in Lagos and Ogun States, Nigeria. Bio-Research 2021, 19, 1258–1269. [Google Scholar] [CrossRef]
- Ukwungwu, M.N.; Odebiyi, J.A. Host plant resistance in rice to the African Striped Borer, Chilo zacconius Bles. (Lepidoptera: Pyralidae). Insect Sci. Its Appl. 1990, 11, 639–647. [Google Scholar] [CrossRef]
- Togola, A.; Nwilene, F.E.; Agbaka, A.; Anato, F.; Agunbiade, T.A.; Chougourou, D.C. Farmer knowledge of rice stem borers and their damage in various ecological zones of Benin (West Africa). Cah. Agric. 2010, 19, 262–266. [Google Scholar] [CrossRef]
- Ntanos, D.A.; Koutroubas, S.D. Evaluation of rice for resistance to Pink Stem Borer (Sesamia nonagrioides Lefebre). Field Crops Res. 2000, 66, 63–71. [Google Scholar] [CrossRef]
- Ukwungwu, M.N. Seasonal changes in the stem borer (SB) Maliarpha separatella populations. Int. Rice Res. Newsl. 1987, 12, 34–35. [Google Scholar]
- Ukwungwu, M.N.; Odebiyi, J.A. Yield losses in resistant and susceptible varieties of rice in Nigeria due to Chilo zacconius and other stem borers. Trop. Pest Manag. 1984, 30, 291–295. [Google Scholar] [CrossRef]
- Wopereis, M.C.S.; Defoer, T.; Idinoba, P.; Diack, S.; Dugué, M.J. Participatory Learning and Action Research (PLAR) for Integrated Rice Management (IRM) in Inland Valleys of Sub-Saharan Africa: Technical Manual; AfricaRice Training Series; Africa Rice Center: Cotonou, Benin, 2009. [Google Scholar]
- Chiasson, H. Determination of key factors affecting the population dynamics of Diopsis longicornis and D. apicalis (Diptera: Diopsidae), pests of rice in the Republic of Guinée, West Africa. Ph.D. Thesis, McGill University, Montréal, QC, Canada, 1990. [Google Scholar]
- Bocco, R.; Elie, D.; Gandonou, C. Diopsids (Diopsis thoracica and D. apicalis) damaging rice production in Africa: A review. Int. J. Curr. Res. Biosci. Plant Biol. 2017, 4, 33–41. [Google Scholar]
- Chiasson, H.; Hill, S. Population density, development and behaviour of Diopsis longicornis and D. apicalis (Diptera: Diopsidae) on rice in the Republic of Guinee. Bull. Entomol. Res. 1993, 83, 5–13. [Google Scholar] [CrossRef]
- Umeh, E.D.N.; Joshi, R.C.; Ukwungwu, M.N. Biology, status and management of rice insect pests in Nigeria. Crop Prot. 1992, 11, 408–413. [Google Scholar] [CrossRef]
- Sauphanor, B. Some factors of upland rice tolerance to stem-borers in West Africa. Insect Sci. Its Appl. 1985, 6, 429–434. [Google Scholar] [CrossRef]
- Chu, Y.I. Ecology of rice borers in Taiwan. In Proceedings of the Symposium on Rice Insects, Tokyo, Japan, 19–24 July 1971; Tropical Agriculture Research Centre, Ministry of Agriculture and Environment: Tokyo, Japan, 1971; pp. 155–162. [Google Scholar]
- Hattori, I.; Siwi, S. Rice stemborers in Indonesia. JARQ 1986, 20, 25–30. [Google Scholar]
- Huang, Q.; Yang, J.; Ling, Y.; Jiang, X.; Yu, X.; Rong, Y.; Mo, Y.; Wang, G.; Fu, C.; Long, L. Feeding and oviposition preferences of Chilo auricilius Dudgeon to four species of host plants. J. South. Agric. 2016, 47, 55–58. [Google Scholar]
- Sallam, M.S.; Allsopp, P.G. Preparedness for Borer Incursion; Bureau of Sugar Experiment Stations: Bundaberg, QLD, Australia, 2002. [Google Scholar]
- Yi, X.; Shi, S.; Wang, P.; Chen, Y.; Lu, Q.; Wang, T.; Zhou, X.; Zhong, G. Characterizing potential repelling volatiles for “push-pull” strategy against stem borer: A case study in Chilo auricilius. BMC Genom. 2019, 20, 751. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Huang, P.; Xiong, C. Studies on the occurrence of Chilo auricilius Dudgeon in Yibing Prefecture, Shichuan. Insect Knowl. 1985, 22, 104–106. [Google Scholar]
- Katti, G.; Shanker, C.; Padmakumari, A.P.; Pasalu, I.C. Rice Stem Borers in India: Species Composition and Distribution; Directorate of Rice Research: Hyderabad, India, 2011. [Google Scholar]
- Husain, M.; Begum, N. Seasonal stem borer (SB) population fluctuations in Mymensingh, Bangladesh. Int. Rice Res. Newsl. 1985, 10, 22. [Google Scholar]
- Catling, H.; Islam, Z.; Pattrasudhi, R. Seasonal occurrence of the yellow stem borer Scirpophaga incertulas (Walker) on deepwater rice in Bangladesh and Thailand. Agric. Ecosyst. Environ. 1984, 12, 47–71. [Google Scholar] [CrossRef]
- Rahman, M.T.; Khalequzzaman, M.; Khan, M.A.R. Assessment of infestation and yield loss by stem borers on variety of rice. J. Asia-Pac. Entomol. 2004, 7, 89–95. [Google Scholar] [CrossRef]
- Catling, H.; Islam, Z. Studies on the ecology of the Yellow Stem Borer, Scirpophaga incertulas (Walker)(Pyralidae), in deepwater rice in Bangladesh. Crop Prot. 1995, 14, 57–67. [Google Scholar] [CrossRef]
- Horgan, F.G.; Romena, A.M.; Bernal, C.C.; Almazan, M.L.P.; Ramal, A.F. Stem borers revisited: Host resistance, tolerance, and vulnerability determine levels of field damage from a complex of Asian rice stemborers. Crop Prot. 2021, 142, 105513. [Google Scholar] [CrossRef]
- Horgan, F.G.; Romena, A.M.; Bernal, C.C.; Almazan, M.L.P.; Ramal, A.F. Differences between the strength of preference–performance coupling in two rice stemborers (Lepidoptera: Pyralidae, Crambidae) promotes coexistence at field-plot scales. Environ. Entomol. 2021, 50, 929–939. [Google Scholar] [CrossRef]
- Rubia-Sanchez, E.G.; Nurhasyim; Diah; Heong, K.L.; Zalucki, M.; Norton, G.A. White stem borer damage and grain yield in irrigated rice in West Java, Indonesia. Crop Prot. 1997, 16, 665–671. [Google Scholar] [CrossRef]
- Pujari, D.; Bora, D.K.; Patgiri, P.; Sarma, S. Relative abundance of different stem borer species in Ahu and Sall rice in Jorhat, Assam. Oryza 2008, 45, 44–47. [Google Scholar]
- Li, C. The biology and ecology of the white rice stemborer, in northern Australia [Northern Territory]. Technical Bulletin 171; Northern Territories Department of Primary Industry and Fisheries: Darwin, NT, Australia, 1991. [Google Scholar]
- Department of Primary Industries. Rice Field Guide to Pests, Diseases and Weeds in Southern New South Wales; Department of Primary Industries: Yanco, NSW, Australia, 2013.
- Oliver, B.; Gifford, J. Weight differences among stalk borer larvae collected from rice lines showing resistance in field studies. J. Econ. Entomol. 1975, 68, 134. [Google Scholar] [CrossRef]
- Oliver, B.; Gifford, J.; Trahan, G. Reaction of selected rice lines to stalk borers. J. Econ. Entomol. 1973, 66, 794–796. [Google Scholar] [CrossRef]
- da Silva Martins, J.F.; Van Tan, N.; da Silveira Pinheiro, B. Resistência de arroz de sequeiro à broca-do-colmo e sua associação com características morfológicas das plantas. Pesqui. Agropecuária Bras. 1981, 16, 187–192. [Google Scholar]
- Pantoja, A. Diatraea saccharalis (F.) in rice plants in Puerto Rico. J. Agric. Univ. Puerto Rico 1994, 78, 195. [Google Scholar] [CrossRef]
- Roldan, E.L.; Beuzelin, J.M.; Vanweelden, M.T.; Cherry, R.H. The Sugarcane Borer (Lepidoptera: Crambidae) infests rice at low population levels in Florida. J. Econ. Entomol. 2020, 113, 538–542. [Google Scholar] [CrossRef] [PubMed]
- Hummelen, P.J. Relations between Two Rice Borers in Surinam, Rupela albinella (Cr.) and Diatraea saccharalis (F.), and Their Hymenopterous Larval Parasites; Veenman and Zonen: Wageningen, The Netherlands, 1974. [Google Scholar]
- Ferreira, E.; Barrigossi, J.A.F.; dos Santos, A.B. Perdas de produção pela broca-do-colmo (Diatraea saccharalis Farb. 1794) (Lepidoptera: Pyralidae) em genótipos de arroz de terras altas. Pesqui. Agropecuária Trop. 2004, 34, 99. [Google Scholar]
- Hamm, J.C.; Sidhu, J.K.; Stout, M.J.; Hummel, N.A.; Reagan, T.E. Oviposition behavior of Diatraea saccharalis (Lepidoptera: Crambidae) on different rice cultivars in Louisiana. Environ. Entomol. 2012, 41, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Villegas, J.M.; Way, M.O.; Pearson, R.A.; Stout, M.J. Integrating soil silicon amendment into management programs for insect pests of drill-seeded rice. Plants 2017, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Villegas, J.M.; Wilson, B.E.; Stout, M.J. Assessment of tolerance and resistance of inbred rice cultivars to combined infestations of rice water weevil and stemborers. Entomol. Exp. Et Appl. 2021, 169, 629–639. [Google Scholar] [CrossRef]
- Reay-Jones, F.P.F.; Wilson, L.T.; Showler, A.T.; Reagan, T.E.; Way, M.O. Role of oviposition preference in an invasive crambid impacting two graminaceous host crops. Environ. Entomol. 2007, 36, 938–951. [Google Scholar] [CrossRef] [PubMed]
- Beuzelin, J.M.; Wilson, L.T.; Showler, A.T.; Meszaros, A.; Wilson, B.E.; Way, M.O.; Reagan, T.E. Oviposition and larval development of a stem borer, Eoreuma loftini, on rice and non-crop grass hosts. Entomol. Exp. Et Appl. 2013, 146, 332–346. [Google Scholar] [CrossRef]
- Beuzelin, J.M.; Wilson, B.E.; VanWeelden, M.T.; Meszaros, A.; Way, M.O.; Stout, M.J.; Reagan, T.E. Biology and management of the Mexican Rice Borer (Lepidoptera: Crambidae) in rice in the United States. J. Integr. Pest Manag. 2016, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Van Dither, J.B.M. A method of assessing rice yield losses caused by the stem borers Rupela albinella and Diatrea saccharalis in Surinam and the aspect of economic thresholds. Entomophaga 1971, 16, 185–191. [Google Scholar] [CrossRef]
- Pantoja, A.; Matta, J. Rupela albinella (Lepidoptera: Pyralidae) oviposition behavior and egg distribution on rice plants. J. Entomol. Sci. 1993, 28, 249–253. [Google Scholar] [CrossRef]
- Ferreira, E.; da Silva Martins, J.F.; Zimmermann, F.J.P. Resistência de cultivares e linhagens de arroz à broca-do-colo. Pesqui. Agropecuária Bras. 1979, 14, 317–321. [Google Scholar]
- Gill, H.K.; Capinera, J.L.; McSorley, R. Lesser Cornstalk Borer, Elasmopalpus lignosellus (Zeller)(Insecta: Lepidoptera: Pyralidae). Document EENY-155; Institute of Food and Agricultural Sciences-Extension, University of Florida: Gainesville, FL, USA, 2011; Available online: https://entnemdept.ufl.edu/creatures/field/lesser_cornstalk_borer.htm (accessed on 15 June 2023).
- Schulten, G.; Feijen, H. A new species of Trichogramma (Hymenoptera: Trichogrammatidae) from Malawi, parasitizing eggs of Chilo diffusilineus (de Joannis). Entomol. Berichten. Deel 1982, 42, 142–144. [Google Scholar]
- Ba, N.M.; Dakouo, D.; Nacro, S.; Karamage, F. Seasonal abundance of lepidopteran stemborers and diopsid flies in irrigated fields of cultivated (Oryza sativa) and wild rice (Oryza longistaminata) in western Burkina Faso. Int. J. Trop. Insect Sci. 2008, 28, 30–36. [Google Scholar] [CrossRef]
- Akinsola, E.; Agyen-Sampong, M. The ecology, bionomics and control of rice stem-borers in West Africa. Int. J. Trop. Insect Sci. 1984, 5, 69–77. [Google Scholar] [CrossRef]
- Leonard, A.; Rwegasira, G.M. Abundance and spatial dispersion of rice stem borer species in Kahama, Tanzania. J. Insect Sci. 2015, 15, 1–5. [Google Scholar] [CrossRef]
- Alam, M.S. A survey of rice pests in Nigeria. Trop. Pest Manag. 1992, 38, 115–118. [Google Scholar] [CrossRef]
- Atkinson, P.R. Distribution and natural hosts of Eldana saccharina Walker in Natal, its oviposition sites and feeding pattern. Proc. S. Afr. Sugar Technol. Assoc. 1979, 53, 111–115. [Google Scholar]
- Harris, K.M. Lepidopterous stem borers of cereals in Nigeria. Bull. Entomol. Res. 1962, 53, 139–171. [Google Scholar] [CrossRef]
- Ahmad, I.; Afzal, M.; Khan, A.M.M. Aspects of larval morphology and larval and adult key to the rice stem borers (Insecta: Lepidoptera) and a new record of a rice stem borer Niphadoses gilviberbis (Zell) from Pakistan. Pak. J. Sci. Ind. Res. 1979, 23, 40–45. [Google Scholar]
- Na, B.H.M.P. Etude Morphologique et Biologique de Scirpophaga Melanoclista-(Lep., Pyralidae, Schoenobiinae), un Ravageur du Rizirrigué en Côte d’Ivoire. Morphologie et Biologie d’un Parasite d’Œuf: Telenomus Thestor NIXON (Hym., Scelionidae). Ph.D. Thesis, l’Universite Pierre et Marie Curie, Paris, France, 1978. [Google Scholar]
- Monnet, C. The control of rice insects in the Ivory Coast. In Proceedings of the Congress on the Control of Insects in the Tropical Environment, Part I, Tropical Crops, Marseilles, France, 13–16 March 1979; Chamber of Commerce and Industry of Marseilles: Marseilles, France, 1979; pp. 559–567. [Google Scholar]
- Emosairue, S.; Shiyam, J. A compendium of insect pests and natural enemies associated with lowland rice in south eastern Nigeria. Glob. J. Pure Appl. Sci. 2000, 6, 385–388. [Google Scholar]
- Hafez, M.; Salama, H.S.; Tolba, R.A. Investigations on the biology of the corn borer Sesamia cretica Led. (Lepidoptera—Agrotidae). Z. Angew. Entomol. 1971, 67, 38–44. [Google Scholar] [CrossRef]
- Camargo, A.M.; Arias-Martin, M.; Castanera, P.; Farinos, G.P. Performance of Sesamia nonagrioides on cultivated and wild host plants: Implications for Bt maize resistance management. Pest Manag. Sci. 2020, 76, 3657–3666. [Google Scholar] [CrossRef] [PubMed]
- El-Hawary, I.S.; El-Naggar, M.A.; Henishen, E.Z. Rice stem borer, Chilo agamemnon, population fluctuation and rice varietal resistance. J. Plant Prot. Pathol. Mansoura Univ. 2015, 6, 1519–1525. [Google Scholar] [CrossRef]
- Nye, I.W.B. The Insect Pests of Graminaceous Crops in East Africa. Report of a Survey Carried out Between March 1956 and April 1958, Colonial Research Studies Number 31; Colonial Office: London, UK, 1960.
- John, V.T.; Alum, M.S.; Thottappilly, G. Diseases and insect pests of wetland rice in tropical Africa. In The Wetlands and Rice in SubSaharan Africa, Proceedings of an International Conference on Wetland Utilization for Rice Production in SubSaharan Africa, 4–8 November 1985; Juo, A.S.R., Lowe, J.A., Eds.; International Institute of Tropical Agriculture: Ibadan, Nigeria, 1986; pp. 141–150. [Google Scholar]
- Heinrichs, E.A.; Barrion, A.T. Rice-Feeding Insects and Selected Natural Enemies in West Africa: Biology, Ecology, Identification; International Rice Research Institute: Los Baños, Philippines, 2004. [Google Scholar]
- Pollet, A. Pests on rice in Ivory Coast. 5. Interrelationship between Maliarpha separatella and Pyricularia Oryzae. J. Appl. Entomol. 1978, 85, 324–327. [Google Scholar]
- Descamps, M. Contributions to the study of Diptera: Diopsidae pests of rice in North Cameroon. J. Agric. Trop. Bot. Appl. 1957, 4, 83–93. [Google Scholar]
- Bernard, V. Diopsis thoracica WEST (Dipt. Diopsidae) important ravageur du riz en Afrique de l’Ouest: Données bio-écologiques et application à la lutte intégrée. L’Agron. Trop. 1975, 37, 89–98. [Google Scholar]
- Feijen, H.R.; Feijen, C. A new species of Diopsis L. (Diptera: Diopsidae) from South Africa and Swaziland, and brief review of African species with a large apical wing spot. Afr. Invertebr. 2012, 53, 125–142. [Google Scholar] [CrossRef]
- Gautam, R.D.; Suroshe, S.; Mahapatro, G.K. Unique record of chloropid pest in rice (Oryza sativa). Indian J. Agric. Sci. 2009, 79, 841–843. [Google Scholar]
- Taneja, S.; Nwanze, K.F. Mass rearing of Chilo spp. on artificial diets and its use in resistance screening. Int. J. Trop. Insect Sci. 1990, 11, 605–616. [Google Scholar] [CrossRef]
- Chakravorty, S. Damage to rice grains by stem borer attack. Int. Rice Res. Newsl. 1979, 4, 1. [Google Scholar]
- Ooi, A.C. A padi stem-borer survey in the Muda scheme, Kedah. Malays. Agric. J. 1974, 49, 525–531. [Google Scholar]
- Razali, R.; Yaakop, S.; Abdullah, M.; Ghazali, S.Z.; Zuki, A.A. Insect species composition under SRI management in Tanjung Karang, Selangor, Malaysia. Malays. Appl. Biol. 2015, 44, 59–66. [Google Scholar]
- Jiang, M.X.; Cheng, J.A. Interactions between the striped stem borer Chilo suppressalis (Walk.) (Lep., Pyralidae) larvae and rice plants in response to nitrogen fertilization. J. Pest Sci. 2003, 76, 124–128. [Google Scholar] [CrossRef]
- Koch, L. Rice stem borers at the Kimberley Research Station. J. Dep. Agric. 1960, 1, 1061–1063. [Google Scholar]
- Saini, V.; Singh, S.; Rawal, R.; Venkatesh, Y. Species diversity and distribution of Cnaphalocrocis and Scirpophaga (Lepidoptera: Crambidae) species complex in rice in Tamil Nadu, India. J. Entomol. Zool. Stud. 2017, 5, 1308–1313. [Google Scholar]
- Kumar, A.; Ram, L.; Singh, R.; Singh, B. Biology and behavior of white stem borer (Scirpophaga fusciflua) on rice (Oryza sativa) in India. Indian J. Agric. Sci. 2018, 88, 1937–1940. [Google Scholar] [CrossRef]
- Tandon, V.; Srivastava, A. Seasonal incidence of white stemborer, Scirpophaga fasciflua Hampson on paddy in Himachal Pradesh. Oryza 2018, 55, 349–352. [Google Scholar] [CrossRef]
- Tandon, V.; Srivastava, A. Assessment of yield losses due to White Stem Borer Scirpophaga fusciflua (Hampson) in rice. Indian J. Entomol. 2022, 84, 634–636. [Google Scholar] [CrossRef]
- Litsinger, J.A.; Alviola, A.L.; Dela Cruz, C.G.; Canapi, B.L.; Batay-An, E.H.; Barrion, A.T. Rice white stemborer Scirpophaga innotata (Walker) in southern Mindanao, Philippines. II. Synchrony of planting and natural enemies. Int. J. Pest Manag. 2006, 52, 23–37. [Google Scholar] [CrossRef]
- Saini, V.; Ramaraju, K.; Chitra, N. Occurrence of new stem borer species, Scirpophaga virginia Lepidoptera: Pyraloidea: Crambidae from Tamil Nadu, India and its taxonomic re-description. Ecol. Environ. Conserv. 2017, 23, 325–328. [Google Scholar]
- Joyce, A.L.; Chicas, M.S.; Cervantes, L.S.; Paniagua, M.; Scheffer, S.J.; Solis, M.A. Host-plant associated genetic divergence of two Diatraea spp. (Lepidoptera: Crambidae) stemborers on novel crop plants. Ecol. Evol. 2016, 6, 8632–8644. [Google Scholar] [CrossRef]
- Wilson, B.E.; Villegas, J.M.; Stout, M.J.; Landry, K.J. Relative yield loss from stem borers (Lepidoptera: Crambidae) and Rice Water Weevil (Coleoptera: Curculionidae) in rice. J. Econ. Entomol. 2021, 114, 1159–1165. [Google Scholar] [CrossRef]
- Pantoja, A.; Matta, J.; Correa, F. Stem borer, Rupela albinella (Cramer), and stem-rot, Sclerotium oryza, in southwestren Colombia rice fields. J Agric. Univ. Puerto Rico 1994, 78, 17–22. [Google Scholar]
- Rebelles, R. Principle pests of upland rice. Inf. Agropecuário (Belo Horiz.) 1989, 14, 44–58. [Google Scholar]
- Ferreira, E.; Barrigossi, J.A.F. A field technique for infesting rice with Elasmopalpus lignosellus (Zeller)(Lepidoptera: Pyralidae) and evaluating insecticide treatments. Neotrop. Entomol. 2003, 32, 367–371. [Google Scholar] [CrossRef]
- Rothschild, G. The biology and ecology of rice-stem borers in Sarawak (Malaysian Borneo). J. Appl. Ecol. 1971, 8, 287–322. [Google Scholar] [CrossRef]
- Cook, M. Revision of the genus Maliarpha (Lepidoptera: Pyralidae), based on adult morphology with description of three new species. Bull. Entomol. Res. 1997, 87, 25–36. [Google Scholar] [CrossRef]
- Li, C.S. Sugarcane insect pests with special reference to the moth borers in the Markham Valley, Papua New Guinea. Mushi 1985, 50, 13–18. [Google Scholar]
- Sandhu, G.; Chander, R. Occurrence of green striped borer, Maliarpha separatella Ragonot on sorghum in the Punjab. J. Bombay Nat. Hist. Soc. 1976, 72, 872–873. [Google Scholar]
- Delfosse, E. Les mouches de la famille des Diopsidae (Insecta: Diptera). Bull. D’arthropoda 2006, 27, 31–38. [Google Scholar]
- Gounou, S.; Schulthess, F. Effect of traditional rice/maize intercropping on population densities, crop damage and parasitism of stem-borers in the Ivory Coast. Afr. Plant Prot. 2006, 12, 93–102. [Google Scholar]
- Zheng, X.; Xu, H.; Chen, G.; Wu, J.; Lu, Z. Potential function of Sudan grass and vetiver grass as trap crops for suppressing population of Stripped Stem Borer, Chilo suppressalis in rice. Chin. J. Biol. Control 2009, 25, 299–303. [Google Scholar]
- van den Berg, J. Vetiver grass (Vetiveria zizanioides (L.) Nash) as trap plant for Chilo partellus (Swinhoe) (Lepidoptera: Pyralidae) and Busseola fusca (Fuller) (Lepidoptera: Noctuidae). Ann. Soc. Entomol. Fr. 2006, 42, 449–454. [Google Scholar] [CrossRef]
- Soliman, A. Effect of plant trap, Echinochloa crus-galli on stem borer, Chilo agamemnon Bles infestation in rice fields. Egypt. J. Agric. Res. 1997, 75, 1–9. [Google Scholar]
- Ranganath, H.R.; Prasad, G.S.; Sheeba. The sedge Eleocharis dulcis (Burm. f.) Henschel (Cyperaceae) traps rice white stem borer Scirpophaga innotata (Walker) (Lepidoptera: Pyralidae) in Andamans. Curr. Sci. 2002, 83, 371–372. [Google Scholar]
- Rajesh, K.M.; Sinu, P.A.; Nasser, M. Eleocharis dulcis (Burm.f) as a promising trap plant for the biocontrol of rice white stem borer, Scirpophaga innotata (Walker). Biol. Control 2021, 160, 104676. [Google Scholar] [CrossRef]
- Lu, Y.H.; Zheng, X.S.; Lu, Z.X. Application of vetiver grass Vetiveria zizanioides: Poaceae (L.) as a trap plant for rice stem borer Chilo suppressalis: Crambidae (Walker) in the paddy fields. J. Integr. Agric. 2019, 18, 797–804. [Google Scholar] [CrossRef]
- Butterfly House. Lepidoptera. Available online: http://lepidoptera.butterflyhouse.com.au/acro/truncata.html (accessed on 15 June 2023).
- Mohyuddin, A.I. Biological control of Chilo spp. in maize, sorghum and millet. Int. J. Trop. Insect Sci. 1990, 11, 719–732. [Google Scholar] [CrossRef]
- Moyal, P.; Tran, M. Chilo aleniellus (Lepidoptera: Pyralidae), a stem borer of maize in Côte d’Ivoire. Bull. Entomol. Res. 1992, 82, 67–72. [Google Scholar] [CrossRef]
- Akinsola, E. Management of Chilo spp. in rice in Africa. Int. J. Trop. Insect Sci. 1990, 11, 815–823. [Google Scholar] [CrossRef]
- Kfir, R.; Overholt, W.A.; Khan, Z.R.; Polaszek, A. Biology and management of economically important lepidopteran cereal stem borers in Africa. Annu. Rev. Entomol. 2002, 47, 701–731. [Google Scholar] [CrossRef]
- Catindig, J.; Barrion, A.; Litsinger, J. Life history, alternate hosts, and natural enemies of the gold fringed borer, Chilo auricilius Dudgeon (Lepidoptera: Pyralidae): A new pest of upland rice (Oryza sativa) in the Philippines. In Proceedings of the 17th Anniversary and Annual Convention of the Pest Control Council of the Philippines, Iloilo City, Philippines, 8–10 May 1986. [Google Scholar]
- Harris, K. Lepidopterous stem borers of sorghum. In Proceedings of the International Sorghum Entomology Workshop, Texas A and M University, College Station, TX, USA, 15–21 July 1984; International Crops Research Institute for the Semi-Arid Tropics: Patancheru, Andhra Pradesh, India, 1985; pp. 161–167. [Google Scholar]
- Chamberlain, K.; Khan, Z.R.; Pickett, J.A.; Toshova, T.; Wadhams, L.J. Diel periodicity in the production of green leaf volatiles by wild and cultivated host plants of stemborer moths, Chilo partellus and Busseola fusca. J. Chem. Ecol. 2006, 32, 565–577. [Google Scholar] [CrossRef]
- Midega, C.A.O.; Khan, Z.R.; Pickett, J.A.; Nylin, S. Host plant selection behaviour of Chilo partellus and its implication for effectiveness of a trap crop. Entomol. Exp. Appl. 2011, 138, 40–47. [Google Scholar] [CrossRef]
- Mohamed, H.; Khan, Z.; Overholt, W.; Elizabeth, D. Behaviour and biology of Chilo partellus (Lepidoptera: Pyralidae) on maize and wild gramineous plants. Int. J. Trop. Insect Sci. 2004, 24, 287–297. [Google Scholar] [CrossRef]
- Rebe, M.; Van den Berg, J.; McGeoch, M. Growth and development of Chilo partellus (Swinhoe)(Lepidoptera: Crambidae) on cultivated and indigenous graminaceous host plants. Afr. Entomol. 2004, 12, 253–258. [Google Scholar]
- Chaudhary, R.; Khush, G.; Heinrichs, E. Varietal resistance to rice stem-borers in Asia. Int. J. Trop. Insect Sci. 1984, 5, 447–463. [Google Scholar] [CrossRef]
- Cuong, N.L.; Cohen, M.B. Field survey and greenhouse evaluation of non-rice host plants of the Striped Stem Borer, Chilo suppressalis (Lepidoptera: Pyralidae), as refuges for resistance management of rice transformed with Bacillus thuringiensis toxin genes. Bull. Entomol. Res. 2002, 92, 265–268. [Google Scholar] [CrossRef] [PubMed]
- Hou, M.L.; Hao, L.X.; Han, Y.Q.; Liao, X.L. Host status of wheat and corn for Chilo suppressalis (Lepidoptera: Crambidae). Environ. Entomol. 2010, 39, 1929–1935. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Hu, B.; Li, J.H.; Wang, Y.M. Water-oats harbors two strains of the Striped Stem Borer Chilo suppressalis (Lepidoptera: Crambidae) with temporal divergence in mating behavior. Appl. Entomol. Zool. 2016, 51, 457–463. [Google Scholar] [CrossRef]
- Jiang, W.H.; Li, H.D.; Cheng, X.F.; Ye, J.R.; Feng, Y.B.; Han, Z.J. Study on host plants for reproduction of Chilo suppressalis. J. Asia-Pac. Entomol. 2015, 18, 591–595. [Google Scholar] [CrossRef]
- Kung, K.-S. Ecological studies on the rice stem borer (Chilo suppressalis Walker) in Taiwan (II): Host plant survey. In Proceedings of the Symposium on Tropical Agriculture Researches, Tokyo, Japan, 19–24 July 1971; p. 27. [Google Scholar]
- Morrill, W.L.; Arida, G.S. Oviposition preference and survival of selected rice insects on wheat. J. Econ. Entomol. 1991, 84, 656–658. [Google Scholar] [CrossRef]
- Quan, W.L.; Liu, W.; Zhou, R.Q.; Chen, R.; Ma, W.H.; Lei, C.L.; Wang, X.P. Difference in diel mating time contributes to assortative mating between host plant-associated populations of Chilo suppressalis. Sci. Rep. 2017, 7, 45265. [Google Scholar] [CrossRef]
- Beuzelin, J.M.; Meszaros, A.; Reagan, T.E.; Wilson, L.T.; Way, M.O.; Blouin, D.C.; Showler, A.T. Seasonal infestations of two stem borers (Lepidoptera: Crambidae) in noncrop grasses of Gulf Coast rice agroecosystems. Environ. Entomol. 2011, 40, 1036–1050. [Google Scholar] [CrossRef] [PubMed]
- Sampson, M.; Kumar, R. Alternative host plants of sugar-cane stem-borers in southern Ghana. Int. J. Trop. Insect Sci. 1986, 7, 539–541. [Google Scholar] [CrossRef]
- White, W.; Richard, R.; Hale, A. Erianthus: A sugarcane relative with potential as a source of resistance to the stem borer Diatraea saccharalis (F.)(Lepidoptera: Crambidae). In Proceedings of the International Society of Sugar Cane Technologists: Proceedings of the XXVIIIth Congress, São Paulo, Brazil, 24–27 June 2013; pp. 825–834. [Google Scholar]
- Atachi, P.; Sekloka, E.; Schulthess, F. Study on some bioecological aspects of Eldana saccharina Walker (Lep., Pyralidae) on Zea mays L. and alternative host plants. J. Appl. Entomol. 2005, 129, 447–455. [Google Scholar] [CrossRef]
- Beuzelin, J.M.; Meszaros, A.; Way, M.O.; Reagan, T.E. Rice harvest cutting height and ratoon crop effects on late season and overwintering stem borer (Lepidoptera: Crambidae) infestations. Crop Prot. 2012, 34, 47–55. [Google Scholar] [CrossRef]
- Beuzelin, J.M.; Reagan, T.E.; Way, M.O.; Meszaros, A.; Akbar, W.; Wilson, L.T. Potential impact of Mexican Rice Borer non-crop hosts on sugarcane IPM. Int. Sugar J. 2011, 113, 660–665. [Google Scholar]
- Rodríguez-del-Bosque, L.A.; Palomo Salas, J.; Mendez Rodriguez, A. Susceptibility of bermudagrass cultivars to Eoreuma loftini (Lepidoptera: Pyralidae) in subtropical Mexico. Fla. Entomol. 1996, 79, 188–193. [Google Scholar] [CrossRef]
- Showler, A.T.; Reagan, T.E. New findings on factors affecting Mexican rRice Borer, Eoreuma loftini Dyar, infestation of sugarcane: Leaf characteristics, drought stress, soil fertility, and alternate host plants. Int. Sugar J. 2011, 113, 744. [Google Scholar]
- Showler, A.T.; Beuzelin, J.M.; Reagan, T.E. Alternate crop and weed host plant oviposition preferences by the Mexican Rice Borer (Lepidoptera: Crambidae). Crop Prot. 2011, 30, 895–901. [Google Scholar] [CrossRef]
- Showler, A.T.; Reagan, T.E. Mexican Rice Borer, Eoreuma loftini (Dyar)(Lepidoptera: Crambidae): Range expansion, biology, ecology, control tactics, and new resistance factors in United States sugarcane. Am. Entomol. 2017, 63, 36–51. [Google Scholar] [CrossRef]
- Showler, A.T.; Wilson, B.E.; Reagan, T.E. Mexican Rice Borer (Lepidoptera: Crambidae) injury to corn greater than to sorghum and sugarcane under field conditions. J. Econ. Entomol. 2012, 105, 1597–1602. [Google Scholar] [CrossRef]
- Arvind, A. Host plants for yellow rice borer (YSB) Scirpophaga incertulas and white stem borer (WSB) Scirpophaga innotata. Int. Rice Res. Newsl. 1987, 12, 3. [Google Scholar]
- Catindig, J.; Barrion, A.; Litsinger, J. Host range of yellow rice borer, brown and whitebacked planthoppers. In Proceedings of the 19th Pest Control Council of the Philippines, Cebu City, Philippines, 3–7 May 1988. [Google Scholar]
- Chen, Y.H.; Romena, A. Feeding patterns of Scirpophaga incertulas (Lepidoptera: Crambidae) on wild and cultivated rice during the booting stage. Environ. Entomol. 2006, 35, 1094–1102. [Google Scholar] [CrossRef]
- Goftishu, M. Ecology and Genetic Diversity of Cereal Stem Borers in Cultivated and Natural Habitats of Ethiopia. Ph.D. Thesis, University of Fort Hare, Alice, South Africa, 2018. [Google Scholar]
- Khaliq, A.; Ashfaq, M.; Akram, W.; Choi, J.K.; Lee, J.J. Effect of plant factors, sugar contents, and control methods on the Top Borer (Scirpophaga nivella F.) Infestation in selected varieties of sugarcane. Entomol. Res. 2005, 35, 153–160. [Google Scholar] [CrossRef]
- Liu, Z.R.; Gao, Y.L.; Luo, J.; Lai, F.X.; Li, Y.H.; Fu, Q.; Peng, Y.F. Evaluating the non-rice host plant species of Sesamia inferens (Lepidoptera: Noctuidae) as natural refuges: Resistance management of Bt rice. Environ. Entomol. 2011, 40, 749–754. [Google Scholar] [CrossRef]
- Ndemah, R.; Gounou, S.; Schulthess, F. The role of wild grasses in the management of lepidopterous stem-borers on maize in the humid tropics of western Africa. Bull. Entomol. Res. 2007, 92, 507–519. [Google Scholar] [CrossRef] [PubMed]
- Padhi, G.; Prakasa Rao, P.S. Preliminary studies on reaction of some wild rices to infestation by the yellow rice borer Tryporyza incertulas Wlk. Oryza 1978, 15, 99–101. [Google Scholar]
- Passoa, S.; Habeck, D.H. A description of the larva and pupa of Rupela albinella, a pest of rice in Latin America (Lepidoptera: Pyralidae: Schoenobiinae). Fla. Entomol. 1987, 70, 368–375. [Google Scholar] [CrossRef]
- Srivastava, A.; Sarao, P.S.; Ram, L.; Salaia, R.; Singh, A. New record of Scirpophaga fusciflua Hampson from North India. New Facet. 21 St. Century Plant Breed 2012, 5, 62. [Google Scholar]
- Dutta, S.; Roy, N. Population dynamics and host preference of a major pest, Scirpophaga incertulas Walker (Pyralidae: Lepidoptera). Int. J. Adv. Sci. Res. 2018, 3, 120–127. [Google Scholar]
- Sutrisno, H. New host record of the Yellow Stem Borer moths, Scirpophaga incertulas Shall (Lepidoptera: Pyralidae). Indones. J. Biol. 2007, 4, 80500. [Google Scholar]
- Zaheruddeen, S.M.; Rao, P.S. Host status of weed flora of rice ecosystem to the yellow rice borer Scirpophaga incertulas Walker. Oryza 1983, 20, 177–179. [Google Scholar]
- Zaheruddeen, S.M.; Rao, P.S. Record of some wild species of rice as potential hosts of the yellow rice borer. Sci. Cult. 1983, 49, 207–208. [Google Scholar]
- Zhang, Z.Y.; Soulemane, B.; Guo, X.P.; Zhu, C.L.; Xu, H.L. Grain yield and weed-mediated pest incidence in paddy rice crops under controllable irrigation regimes. J. Food Agric. Environ. 2012, 10, 626–631. [Google Scholar]
- Yasumatsu, K.; Nishida, T.; Bess, H.A. On the extinction of Asiatic rice borer Chilo suppressalis in Hawaii. Proc. Hawaii. Entomol. Soc. 1968, 20, 239–245. [Google Scholar]
- Sezonlin, M.; Dupas, S.; Le RÜ, B.; Le Gall, P.; Moyal, P.; Calatayud, P.A.; Giffard, I.; Faure, N.; Silvain, J.F. Phylogeography and population genetics of the maize stalk borer Busseola fusca (Lepidoptera, Noctuidae) in sub-Saharan Africa. Mol. Ecol. 2006, 15, 407–420. [Google Scholar] [CrossRef]
- Koutroubas, S.D.; Ntanos, D. Plant traits affecting rice resistance to Pink Stem Borer (Sesamia nonagrioides Lefebre). In Rice Genetic Resources and Breeding for Europe and Other Temperate Areas, Proceedings of Eurorice 2001 Symposium, Krasnodar, Russia, 3–8 September 2001; Centre de Coopération International en Recherche Agronomique Pour le Développement (CIRAD IRD): Monpellier, France, 2002; pp. 504–509. [Google Scholar]
- Esfandiari, M.; Mossadegh, M.S.; Shishehbor, P. Noctuidae sl (Lepidoptera) from sugarcane fields of SW Iran. Fragm. Faun. 2011, 54, 137–147. [Google Scholar] [CrossRef]
- Esfandiari, M.; Mehravar, N.; Motamedi, H.; Memari, H.R.; Rajaei, H. Phylogeography of Sesamia nonagrioides (Lepidoptera: Noctuidae) in Iran inferred from mitochondrial DNA. Acta Zool. Acad. Sci. Hung. 2015, 61, 341–359. [Google Scholar] [CrossRef]
- Sharma, A.N.; Sharma, V.K. Studies on the economic injury level in maize, Zea mays L. to stem borer, Chilo partellus (Swinhoe) (Pyralidae: Lepidoptera) in India. Trop. Pest Manag. 1987, 33, 44–51. [Google Scholar] [CrossRef]
- Mahadevan, N.R.; Chelliah, S. Influence of intercropping legumes with sorghum on the infestation of the stem borer, Chilo partellus (Swinhoe) in Tamil Nadu, India. Trop. Pest Manag. 1986, 32, 162–163. [Google Scholar] [CrossRef]
- Ebert, G. The rice stem borer Chilo suppressalis Walker (Lep./Pyral.), a new pest for Iran. Entomol. Phytopathol. Appliquées 1973, 35, 4. [Google Scholar]
- Poltavsky, A.N.; Artokhin, K.S. Invasions by Chilo Zincken, 1817 to the south of European Russia (Lepidoptera: Crambidae). Shilap-Rev. Lepidopterol. 2015, 43, 461–465. [Google Scholar]
- Moche, K.; Lordon, C.D.; Tadu, Z.; Mokam, D.G.; AlainNana, P.; Fokam, Z.; Woin, N. Diversity and agronomic impact of rice stem borer at Nkolbisson, Yaoundé-Cameroon (Central Africa). Int. J. Agron. Agric. Res. 2015, 6, 181–189. [Google Scholar]
- Nwilene, F.; Onasanya, A.; Togola, A.; Oyetunji, O.; Semon, M.; Tamo, M.; Bright, E.; Ofodile, S. Effect of intercropping maize and cassava with upland NERICA rice varieties on stemborer attack in southwest Nigeria. J. Entomol. 2011, 8, 417–428. [Google Scholar] [CrossRef]
- Vera Piguabe, K.E. Incidencia de Rupela albinella y Diatraea saccharalis en Siete Líneas Avanzadas F4 de Arroz, Derivadas de Cruces Interespecíficos Entre Oryza sativa L. ssp japonica y Oryza rufipogon G. en el Cantón Babahoyo, Los Ríos. Bachelor’s Thesis, Universidad Técnica de Babahoya, Babahoya, Ecuador, 2019. [Google Scholar]
- Das, Y.T. Cross resistance to stemborers in rice varieties. J. Econ. Entomol. 1976, 69, 41–46. [Google Scholar] [CrossRef]
- Zhu, Z.R.; Romena, A.M.; Cohen, M.B. Comparison of stem borer damage and resistance in semidwarf Indica rice varieties and prototype lines of a new plant type. Field Crops Res. 2002, 75, 37–45. [Google Scholar] [CrossRef]
- Neupane, F.P.; Coppel, H.C.; Chapman, R.K. Bionomics of the maize borer, Chilo partellus (Swinhoe), in Nepal. Int. J. Trop. Insect Sci. 2011, 6, 547–553. [Google Scholar] [CrossRef]
- Rahaman, M.M.; Islam, K.S.; Jahan, M.; Mamun, M.A.A. Relative abundance of stem borer species and natural enemies in rice ecosystem at Madhupur, Tangail, Bangladesh. J. Bangladesh Agril. Univ. 2014, 12, 267–272. [Google Scholar] [CrossRef]
- Ooi, A.C.P. Assessment of incidence of rice stem-borers in Tanjong Karang, Malaysia. Malays. Agric. J. 1976, 50, 314–321. [Google Scholar]
- Wijaya, I.N.; Adiartayasa, W.; Yuliadhi, D.K.A. Komposisi spesies penggerek batang padi dan parasitoid telurnya pada Ketinggian tempat yang berbeda. J. Agric. Sci. 2021, 11, 1–9. [Google Scholar] [CrossRef]
- Kumbhar, C.R.; Singh, S.P.N. Species compositions of rice stem borers in different rice cultivation system under north Bihar condition. J. Entomol. Zool. Stud. 2020, 6, 1906–1909. [Google Scholar]
- Horgan, F.G.; Crisol-Martinez, E.; Almazan, M.L.P.; Romena, A.; Ramal, A.F.; Ferrater, J.B.; Bernal, C.C. Susceptibility and tolerance in hybrid and pure-line rice varieties to herbivore attack: Biomass partitioning and resource-based compensation in response to damage. Ann. Appl. Biol. 2016, 169, 200–213. [Google Scholar] [CrossRef]
- Hummel, N.A.; Hardy, T.; Reagan, T.E.; Pollet, D.; Carlton, C.; Stout, M.J.; Beuzelin, J.M.; Akbar, W.; White, W. Monitoring and first discovery of the Mexican Rice Borer Eoreuma loftini (Lepidoptera: Crambidae) in Louisiana. Fla. Entomol. 2010, 93, 123–124. [Google Scholar] [CrossRef]
- Reay-Jones, F.P.F.; Wilson, L.T.; Way, M.O.; Reagan, T.E.; Carlton, C.E. Movement of Mexican Rice Borer (Lepidoptera: Crambidae) through the Texas rice belt. J. Econ. Entomol. 2007, 100, 54–60. [Google Scholar] [CrossRef] [PubMed]
- da Silva Martins, J.F.; da Silveira Pinheiro, B.; Lowe, J.A. Nitrogênio e infestação da broca-do-colmo em arroz irrigado. Pesqui. Agropecuária Bras. 1978, 13, 23–25. [Google Scholar]
- Ho, D.T.; Kibuka, J.G. Effect of nitrogen and plant density on rice stemborer infestation in Western Kenya. Int. Rice Res. Newsl. 1983, 8, 17–18. [Google Scholar]
- Litsinger, J.A.; Canapi, B.L.; Bandong, J.P. Cultural practices mitigate irrigated rice insect pest losses in the Philippines. Philipp. J. Sci. 2011, 140, 179–194. [Google Scholar]
- Zhu, Y.; Chen, J.; Zou, Y.; Huong, X.; Jiong, T.; Wyckhuys, K.A.G.; van der Werf, W.; Xiao, H. Response of the rice stem borer Chilo suppressalis (Walker) and its parasitoid assemblage to landscape composition. Agric. Ecosyst. Environ. 2023, 343, 108259. [Google Scholar] [CrossRef]
- Sarwar, M. Effects of potassium fertilization on population build up of rice stem borers (lepidopteron pests) and rice (Oryza sativa L.) yield. J. Cereals Oilseeds 2012, 3, 6–9. [Google Scholar]
- Van Weelden, M.T.; Wilson, B.E.; Beuzelin, J.M.; Reagan, T.E.; Way, M.O. Impact of nitrogen fertilization on Mexican Rice Borer (Lepidoptera: Crambidae) injury and yield in bioenergy sorghum. Crop Prot. 2016, 84, 37–43. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhang, X.; Liu, X.; Qin, N.; Xu, K.; Zeng, R.; Liu, J.; Song, Y. Nitrogen supply alters rice defense against the striped stem borer Chilo suppressalis. Front. Plant Sci. 2021, 12, 691292. [Google Scholar] [CrossRef]
- Boucher, A.; Silvie, P.; Menozzi, P.; Adda, C.; Auzoux, S.; Jean, J.; Huat, J. Landscape changes in a lowland in Benin: Ecological impact on pests and natural enemies. Commun. Agric. Appl. Biol. Sci. 2015, 80, 79–89. [Google Scholar]
- Kega, V.M.; Kasina, M.; Olubayo, F.M.; Nderitu, J.H. A study of the African White Rice Stem Borer (Maliarpha separatella Rag.) population density fluctuations at Mwea Irrigation Scheme in central Kenya. Univ. J. Agric. Res. 2017, 5, 52–56. [Google Scholar]
- Babendreier, D.; Tang, R.; Horgan, F.G. Prospects for Integrating augmentative and conservation biological control of leaffolders and stemborers in rice. Agronomy 2022, 12, 2958. [Google Scholar] [CrossRef]
- Ndemah, R.; Schulthess, F.; Le Rü, B.; Bame, I. Lepidopteran cereal stemborers and associated natural enemies on maize and wild grass hosts in Cameroon. J. Appl. Entomol. 2007, 131, 658–668. [Google Scholar] [CrossRef]
- Ogah, E.; Nwilene, F. Incidence of insect pests on rice in Nigeria: A review. J. Entomol. 2017, 14, 58–72. [Google Scholar] [CrossRef]
- Sitepu, M.B.; Tobing, M.C.L.; Bakti, D. The role of refugia plants to the eggs yellow rice stem borer (Scirpophaga incertulas Walker) parasitoids parasitation rate. J. Phys. Conf. Ser. 2018, 1116, 052064. [Google Scholar] [CrossRef]
- Zhu, P.Y.; Wang, G.W.; Zheng, X.S.; Tian, J.C.; Lu, Z.X.; Heong, K.L.; Xu, H.X.; Chen, G.H.; Yang, Y.J.; Gurr, G.M. Selective enhancement of parasitoids of rice Lepidoptera pests by sesame (Sesamum indicum) flowers. Biocontrol 2015, 60, 157–167. [Google Scholar] [CrossRef]
- Joyce, A.L.; White, W.H.; Nuessly, G.S.; Solis, M.A.; Scheffer, S.J.; Lewis, M.L.; Medina, R.F. Geographic population structure of the Sugarcane Borer, Diatraea saccharalis (F.) (Lepidoptera: Crambidae), in the Southern United States. PLoS ONE 2014, 9, e110036. [Google Scholar] [CrossRef] [PubMed]
- Ben-Yakir, D.; Chen, M.; Sinev, S.; Seplyarsky, V. Chilo partellus (Swinhoe)(Lepidoptera: Pyralidae) a new invasive species in Israel. J. Appl. Entomol. 2013, 137, 398–400. [Google Scholar] [CrossRef]
- Zahedi, T.S.; Hajizadeh, J.; Azimi, R. First report of the White Stem Borer, Scirpophaga innotata (Lep.: Crambidae: Schoenobiinae), from Iran. J. Entomol. Soc. Iran 2014, 34, 3–4. [Google Scholar]
- Esfandiari, M.; Mossadegh, M.S.; Shishehbor, P. Sesamia botanephaga Tams & Bowden, 1953 auct. in Iran, read Sesamia nonagrioides (Lefebvre, 1827) (Lepidoptera: Noctuidae). Munis Entomol. Zool. 2011, 6, 400–403. [Google Scholar]
- Stiling, P.D.; Strong, D.R. Weak competition among Spartina stem borers, by means of murder. Ecology 1983, 64, 770–778. [Google Scholar] [CrossRef]
- Wan, N.F.; Cavalieri, A.; Siemann, E.; Dainese, M.; Li, W.W.; Jiang, J.X. Spatial aggregation of herbivores and predators enhances tri-trophic cascades in paddy fields: Rice monoculture versus rice-fish co-culture. J. Appl. Ecol. 2022, 59, 2036–2045. [Google Scholar] [CrossRef]
- Hartley, S.; Shorrocks, B. A general framework for the aggregation model of coexistence. J. Anim. Ecol. 2002, 71, 651–662. [Google Scholar] [CrossRef]
- Oyediran, I.O.; Heinrichs, E.A.; Johnson, D.E. Abundance of rice arthropods and weeds on the continuum toposequence in a West African inland valley. Int. J. Trop. Insect Sci. 2011, 19, 109–119. [Google Scholar] [CrossRef]
- Girling, D.J. Distribution and biology of Eldana saccharina Walker (Lepidoptera-Pyralidae) and its relationship to other stem-borers in Uganda. Bull. Entomol. Res. 1978, 68, 471–488. [Google Scholar] [CrossRef]
- Sokame, B.M.; Rebaudo, F.; Malusi, P.; Subramanian, S.; Kilalo, D.C.; Juma, G.; Calatayud, P.-A. Influence of temperature on the interaction for resource utilization between Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), and a community of lepidopteran maize stemborers larvae. Insects 2020, 11, 73. [Google Scholar] [CrossRef]
- Divya, J.; Kalleshwaraswamy, C.; Mallikarjuna, H.; Deshmukh, S. Does recently invaded Fall Armyworm, Spodoptera frugiperda displace native lepidopteran pests of maize in India. Curr. Sci 2021, 120, 1358–1367. [Google Scholar] [CrossRef]
- Mutua, J.M.; Mutyambai, D.M.; Asudi, G.O.; Khamis, F.; Niassy, S.; Jalloh, A.A.; Salifu, D.; Magara, H.J.O.; Calatayud, P.-A.; Subramanian, S. Competitive plant-mediated and intraguild predation interactions of the invasive Spodoptera frugiperda and resident stemborers Busseola fusca and Chilo partellus in maize cropping systems in Kenya. Insects 2022, 13, 790. [Google Scholar] [CrossRef]
- Sokame, B.M.; Malusi, P.; Subramanian, S.; Kilalo, D.C.; Juma, G.; Calatayud, P.-A. Do the invasive Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), and the maize lepidopteran stemborers compete when sharing the same food? Phytoparasitica 2022, 50, 21–34. [Google Scholar] [CrossRef]
- Kalleshwaraswamy, C.; Divya, J.; Mallikarjuna, H.; Deshmukh, S.S.; Sunil, C. Cannibalistic nature and time of habitat occupancy of invasive maize Fall Armyworm, Spodoptera frugiperda are the key factors for competitive displacement of native stem borer, Sesamia inferens in India. Curr. Sci. 2023, 124, 348–354. [Google Scholar]
- Sokame, B.M.; Musyoka, B.; Mohammed, S.A.; Tamiru, A.; Bruce, A.; Anderson, P.; Karlsson Green, K.; Calatayud, P.-A. Cannibalism and intraguild predation involved in the intra-and inter-specific interactions of the invasive Fall Armyworm, Spodoptera frugiperda, and lepidopteran maize stemborers. J. Pest Sci. 2023, 96, 1455–1464. [Google Scholar] [CrossRef]
- Bordat, D.; Pichot, M. Chilo zacconius Blez. technique d’élevage en milieu artificial et observations sur sa biologie en laboratoire. Agron. Trop. 1978, 33, 337–343. [Google Scholar]
- Shi, P.J.; Wang, B.; Ayres, M.P.; Ge, F.; Zhong, L.; Li, B.L. Influence of temperature on the northern distribution limits of Scirpophaga incertulas Walker (Lepidoptera: Pyralidae) in China. J. Therm. Biol. 2012, 37, 130–137. [Google Scholar] [CrossRef]
- Yonow, T.; Kriticos, D.J.; Ota, N.; Van Den Berg, J.; Hutchison, W.D. The potential global distribution of Chilo partellus, including consideration of irrigation and cropping patterns. J. Pest Sci. 2017, 90, 459–477. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, S.; Chander, S. Simulation of climatic change impact on crop-pest interactions: A case study of rice Pink Stem Borer Sesamia inferens (Walker). Clim. Change 2015, 131, 259–272. [Google Scholar] [CrossRef]
- Ntiri, E.S.; Calatayud, P.-A.; Van den Berg, J.; Le Ru, B.P. Density dependence and temporal plasticity of competitive interactions during utilisation of resources by a community of lepidopteran stemborer species. Entomol. Exp. Appl. 2017, 162, 272–283. [Google Scholar] [CrossRef]
- Ntiri, E.S.; Calatayud, P.-A.; Van den Berg, J.; Le Ru, B.P. Spatio-temporal interactions between maize lepidopteran stemborer communities and possible implications from the recent invasion of Spodoptera frugiperda (Lepidoptera: Noctuidae) in Sub-Saharan Africa. Environ. Entomol. 2019, 48, 573–582. [Google Scholar] [CrossRef]
- Mutamiswa, R.; Chidawanyika, F.; Nyamukondiwa, C. Dominance of Spotted Stemborer Chilo partellus Swinhoe (Lepidoptera: Crambidae) over indigenous stemborer species in Africa’s changing climates: Ecological and thermal biology perspectives. Agric. For. Entomol. 2017, 19, 344–356. [Google Scholar] [CrossRef]
- Ogah, E. Evaluating the impact of new rice for Africa (NERICA) in the management of rice stem borers. Sci. Int. 2013, 1, 160–166. [Google Scholar] [CrossRef]
- Sokame, B.M.; Ntiri, E.S.; Ahuya, P.; Torto, B.; Le Ru, B.P.; Kilalo, D.C.; Juma, G.; Calatayud, P.-A. Caterpillar-induced plant volatiles attract conspecific and heterospecific adults for oviposition within a community of lepidopteran stemborers on maize plant. Chemoecology 2019, 29, 89–101. [Google Scholar] [CrossRef]
- Potting, R.P.J.; Vet, L.E.M.; Dicke, M. Host microhabitat location by stem-borer parasitoid Cotesia flavipes: The role of herbivore volatiles and locally and systemically induced plant volatiles. J. Chem. Ecol. 1995, 21, 525–539. [Google Scholar] [CrossRef] [PubMed]
- Ngi-Song, A.J.; Njagi, P.G.N.; Torto, B.; Overholt, W.A. Identification of behaviourally active components from maize volatiles for the stemborer parasitoid Cotesia flavipes Cameron (Hymenoptera: Braconidae). Int. J. Trop. Insect Sci. 2000, 20, 181–189. [Google Scholar] [CrossRef]
- Gohole, L.S.; Overholt, W.A.; Khan, Z.R.; Vet, L.E.M. Role of volatiles emitted by host and non-host plants in the foraging behaviour of Dentichasmias busseolae, a pupal parasitoid of the Spotted Stemborer Chilo partellus. Entomol. Exp. Appl. 2003, 107, 1–9. [Google Scholar] [CrossRef]
- Horgan, F.G.; Crisol-Martínez, E.; Stuart, A.M.; Villegas, J.M.; Peñalver-Cruz, A.; Mundaca, E.A.; Perez, M.O.; Bernal, C.C.; Almazan, M.L.P.; Ramal, A.F. Direct and indirect effects of planting density, nitrogenous fertilizer and host plant resistance on rice herbivores and their natural enemies. Agriculture 2022, 12, 2053. [Google Scholar] [CrossRef]
- Bianchi, G. Comportement et Nuisibilité de Maliarpha separatella Ragonot (Lep. Phycitinae) Dans les Rizières Malgaches du lac Alaotra et Modélisation de la Dynamique de Ses Populations; ETH: Zurich, Switzerland, 1989. [Google Scholar]
- Bianchi, G.; Baumgartner, J.; Delucchi, V.; Rahalivavololona, N.; Skillman, S.; Zahner, P. Sampling egg batches of Maliarpha separatella Rag (Lep, Pyralidae) in Madagascan rice fields. Trop. Pest Manag. 1989, 35, 420–424. [Google Scholar] [CrossRef]
- Kishore Kumar, V.; Sharma, H.C.; Dharma Reddy, K. Antibiosis mechanism of resistance to Spotted Stem Borer, Chilo partellus in sorghum, Sorghum biocolor. Crop Prot. 2006, 25, 66–72. [Google Scholar] [CrossRef]
- Ngomane, N.C. Optimization of Formulated Artificial Diets with the Addition of Sterols and Cryoprotectants for Effective Rearing and Fitness of Eldana saccharina Walker (Lepidoptera: Pyralidae). Master’s Thesis, Stellenbosch University, Stellenbosch, South Africa, 2021. [Google Scholar]
- Ofomata, V.; Overholt, W.; Lux, S.; Van Huis, A.; Egwuatu, R. Comparative studies on the fecundity, egg survival, larval feeding, and development of Chilo partellus and Chilo orichalcociliellus (Lepidoptera: Crambidae) on five grasses. Ann. Entomol. Soc. Am. 2000, 93, 492–499. [Google Scholar] [CrossRef]
- Tamiru, A.; Getu, E.; Jembere, B.; Bruce, T. Effect of temperature and relative humidity on the development and fecundity of Chilo partellus (Swinhoe)(Lepidoptera: Crambidae). Bull. Entomol. Res. 2012, 102, 9–15. [Google Scholar] [CrossRef]
- Fantinou, A.A.; Perdikis, D.C.; Chatzoglou, C.S. Development of immature stages of Sesamia nonagrioides (Lepidoptera: Noctuidae) under alternating and constant temperatures. Environ. Entomol. 2003, 32, 1337–1342. [Google Scholar] [CrossRef]
- Fantinou, A.A.; Perdikis, D.C.; Stamogiannis, N. Effect of larval crowding on the life history traits of Sesamia nonagrioides (Lepidoptera: Noctuidae). Eur. J. Entomol. 2008, 105, 625–630. [Google Scholar] [CrossRef]
- Fantinou, A.A.; Perdikis, D.C.H.; Zota, K.F. Reproductive responses to photoperiod and temperature by diapausing and nondiapausing populations of Sesamia nonagrioides Lef. (Lepidoptera−Noctuidae). Physiol. Entomol. 2004, 29, 169–175. [Google Scholar] [CrossRef]
- López, C.; Sans, A.; Asin, L.; EizaGuirre, M. Phenological model for Sesamia nonagrioides (Lepidoptera: Noctuidae). Environ. Entomol. 2001, 30, 23–30. [Google Scholar] [CrossRef]
- Sandhu, H.S.; Nuessly, G.S.; Webb, S.E.; Cherry, R.H.; Gilbert, R.A. Temperature-dependent development of Elasmopalpus lignosellus (Lepidoptera: Pyralidae) on sugarcane under laboratory conditions. Environ. Entomol. 2010, 39, 1012–1020. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, H.S.; Nuessly, G.S.; Webb, S.E.; Cherry, R.H.; Gilbert, R.A. Temperature-dependent reproductive and life table parameters of Elasmopalpus lignosellus (Lepidoptera: Pyralidae) on sugarcane. Fla. Entomol. 2013, 2013, 380–390. [Google Scholar] [CrossRef]
- Castro Borbor, Á.R. Altura del Daño Ocasionado por Larvas de Rupela albinella (Cramer), en Tres Variedades de Arroz, Bajo Tres Sistemas de Manejo de Agua. Bachelor’s Thesis, Universidad Tecnica de Babahoyo, Babahoyo, Ecuador, 2011. [Google Scholar]
- Jarvis, E. Notes on Insects Damaging Sugar Cane in Queensland; Bureau of Sugar Experiment Stations: Brisbane, Australia, 1916; Volume 3, p. 47. [Google Scholar]
- Kiritani, K. The biology and life cycle of Chilo suppressalis (Walker) and Tryporyza (Shoenobius) incertulas (Walker) in temperate-climate areas. In Proceedings of the Symposium: The Major Insect Pests of the Rice Plant, International Rice Research Institute (IRRI), Los Baños, Philippines, July 1964; IRRI: Los Baños, Philippines, 1967. [Google Scholar]
- Ramoneda, J.; Roig, J. Characteristics of the larval development of the rice stem borer, Chilo suppressalis WALKER (Lepidoptera:Crambidae) in the Ebro Delta (Northeastern Spain). Appl. Entomol. Zool. 1993, 28, 267–273. [Google Scholar] [CrossRef]
- Rao, Y.; Rao, Y. Bionomics of the crambid borer, Chilo auricilius Dudgeon infesting rice. J. Entomol. Res. 1980, 4, 68–72. [Google Scholar]
- Huang, X.L.; Xiao, L.; He, H.M.; Xue, F.S. Effect of rearing conditions on the correlation between larval development time and pupal weight of the rice stem borer, Chilo suppressalis. Ecol. Evol. 2018, 8, 12694–12701. [Google Scholar] [CrossRef] [PubMed]
- Islam, Z.; Catling, H. Biology and behaviour of rice Yellow Stem Borer in deepwater rice. J. Plant Prot. Trop. 1991, 8, 85–96. [Google Scholar]
- Kumar, A.; Ram, L.; Yadav, S.S.; Mehra, S.; Jat, M.K. Biological parameters and comparative morphometrics studies of Scirpophaga spp. in rice. J. Agric. Ecol. 2023, 15, 62–68. [Google Scholar] [CrossRef]
- Manikandan, N.; Kennedy, J.S.; Geethalakshmi, V. Effect of elevated temperature on life-history parameters of rice Yellow Stem Borer (Scirpophaga incertulas Walker). Curr. Sci. 2016, 110, 851–857. [Google Scholar]
- Padmakumari, A.; Katti, G.; Sailaja, V.; Padmavathi, C.; Lakshmi, V.J.; Prabhakar, M.; Prasad, Y. Delineation of larval instars in field populations of rice Yellow Stem Borer, Scirpophaga incertulas (Walk.). ORYZA-Int. J. Rice 2013, 50, 259–267. [Google Scholar]
- Xiao, H.J.; Chen, J.H.; Chen, L.Y.; Chen, C.; Wu, S.H. Exposure to mild temperatures decreases overwintering larval survival and post-diapause reproductive potential in the rice stem borer Chilo suppressalis. J. Pest Sci. 2017, 90, 117–125. [Google Scholar] [CrossRef]
- Ruhela, S.K.; Yadav, P.; Kumar, A. Biology of Sugarcane Top Borer, Scirpophaga nivella (Lepidoptera). Ann. Entomol. 2017, 35, 47–51. [Google Scholar]
- Viswajyothi, K.; Aggarwal, N.; Jindal, J. The biology of Sesamia inferens (Walker)(Lepidoptera: Noctuidae) on maize in the north western plains of India. Acta Phytopathol. Et Entomol. Hung. 2019, 54, 69–84. [Google Scholar] [CrossRef]
- Horgan, F.G.; Arida, A.; Ardestani, G.; Almazan, M.L.P. Intraspecific competition counters the effects of elevated and optimal temperatures on phloem-feeding insects in tropical and temperate rice. PLoS ONE 2020, 15, 0240130. [Google Scholar] [CrossRef]
- Horgan, F.G.; Arida, A.; Ardestani, G.; Almazan, M.L.P. Positive and negative interspecific interactions between coexisting rice planthoppers neutralise the effects of elevated temperatures. Funct. Ecol. 2021, 35, 181–192. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Approximate distributions of economically important rice stemborer moths in Africa and the Mediterranean (blue font), and in Asia and Oceania (green font). Shaded polygons indicate distinct stemborer assemblages occurring in rice. Distribution ranges were estimated using published field data and regional/national records as indicated with the Supplementary Information. The known distributions of the economically important rice diopsids based on information in Khan et al. (1991) [9] are indicated in the inset figure.
Figure 1.
Approximate distributions of economically important rice stemborer moths in Africa and the Mediterranean (blue font), and in Asia and Oceania (green font). Shaded polygons indicate distinct stemborer assemblages occurring in rice. Distribution ranges were estimated using published field data and regional/national records as indicated with the Supplementary Information. The known distributions of the economically important rice diopsids based on information in Khan et al. (1991) [9] are indicated in the inset figure.

Figure 2.
Examples of stemborer assemblage composition based on dissected rice stems from farmers’ fields and experimental field plots in Africa (A–F), the Americas (G–J), South Asia (K–O). and Southeast Asia (P–Y). Pie charts represent the proportional occurrence of each stemborer species based on numbers of individuals and are not indicative of related damage. The pie charts are based on data presented by Ba et al. (2008) [96] (A,B); Adewoye et al. (2021) [49] (C–E); January et al. (2021) [48] (F); Van Dither (1971) [91] (G); Vera Piguabe (2019) [202] (H); Roldan et al. (2020) [82] (I); Beuzelin et al. (2012) [169] (J); Katti et al. (2011) [67] (K–O); Khari and Hamid (2022) ([23]) (P–R); Das et al. (1976) [203] (S); Horgan et al. (2021) [73] (T); Horgan (unpublished data) (U); and Zhu et al. (2002) [204] (V–Y). Stemborer groups or species are indicated in the legends. DS = dry season, WS = wet season.
Figure 2.
Examples of stemborer assemblage composition based on dissected rice stems from farmers’ fields and experimental field plots in Africa (A–F), the Americas (G–J), South Asia (K–O). and Southeast Asia (P–Y). Pie charts represent the proportional occurrence of each stemborer species based on numbers of individuals and are not indicative of related damage. The pie charts are based on data presented by Ba et al. (2008) [96] (A,B); Adewoye et al. (2021) [49] (C–E); January et al. (2021) [48] (F); Van Dither (1971) [91] (G); Vera Piguabe (2019) [202] (H); Roldan et al. (2020) [82] (I); Beuzelin et al. (2012) [169] (J); Katti et al. (2011) [67] (K–O); Khari and Hamid (2022) ([23]) (P–R); Das et al. (1976) [203] (S); Horgan et al. (2021) [73] (T); Horgan (unpublished data) (U); and Zhu et al. (2002) [204] (V–Y). Stemborer groups or species are indicated in the legends. DS = dry season, WS = wet season.

Figure 3.
Approximate distribution of lepidopteran stemborers that attack rice and other crops in the Americas. Colored polygons indicate distinct rice stemborer assemblages.
Figure 3.
Approximate distribution of lepidopteran stemborers that attack rice and other crops in the Americas. Colored polygons indicate distinct rice stemborer assemblages.

Figure 4.
Approximate use of three resources by stemborers based on published information (darker shading indicates peak occupancy). ‘Rice main’ indicates that rice is the main host; ‘other main’ indicates that other crops are the main hosts.
Figure 4.
Approximate use of three resources by stemborers based on published information (darker shading indicates peak occupancy). ‘Rice main’ indicates that rice is the main host; ‘other main’ indicates that other crops are the main hosts.

Figure 5.
Biplot of reported minimum stemborer female pupal weights versus minimum female wingspans for African (brown symbols), Asian (blue symbols), and American (gray symbols) species. Some of the data may refer to species reared on other crops and not rice. See Table S4 for further details on species bionomics and source information [10,21,55,62,64,66,67,72,73,76,83,89,90,94,101,106,107,111,117,128,134,245,255,256,257,258,259,260,261,262,263,264,265,266,267,268,269,270,271,272,273,274,275,276,277,278,279].
Figure 5.
Biplot of reported minimum stemborer female pupal weights versus minimum female wingspans for African (brown symbols), Asian (blue symbols), and American (gray symbols) species. Some of the data may refer to species reared on other crops and not rice. See Table S4 for further details on species bionomics and source information [10,21,55,62,64,66,67,72,73,76,83,89,90,94,101,106,107,111,117,128,134,245,255,256,257,258,259,260,261,262,263,264,265,266,267,268,269,270,271,272,273,274,275,276,277,278,279].


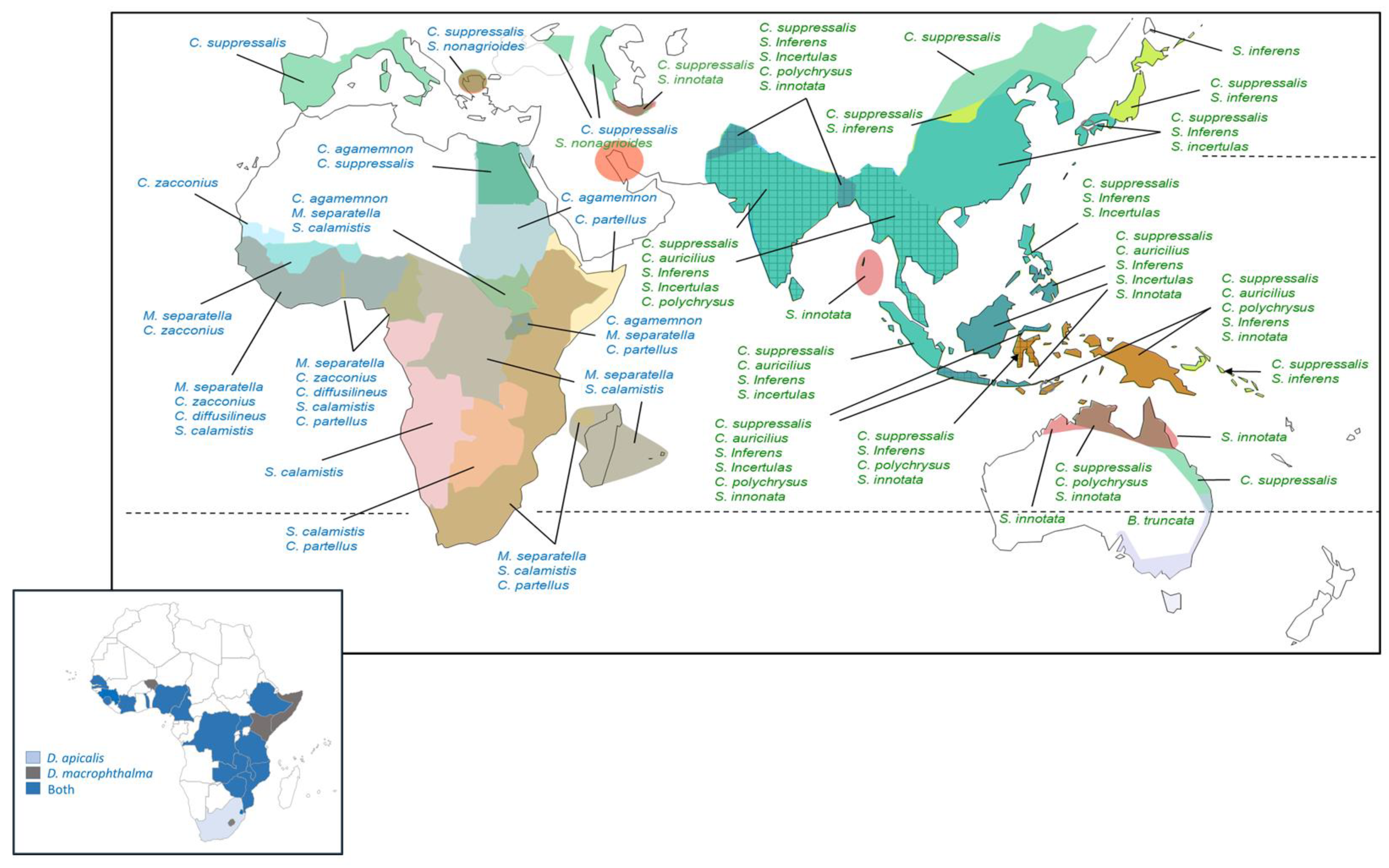{kind=link}
{kind=link}
{kind=link}
{kind=link}
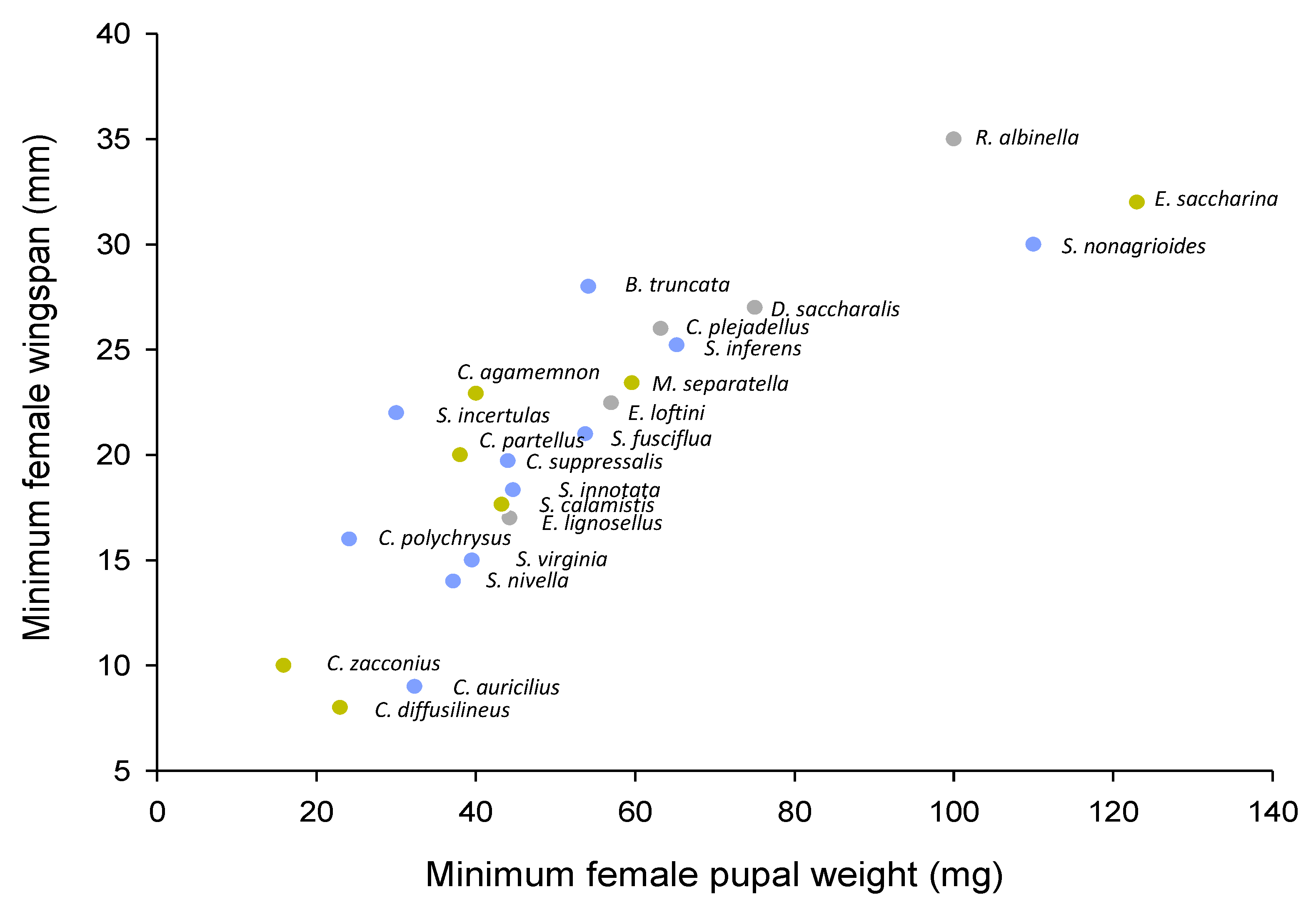{kind=link}
Table 1.
Stemborer species identified as primary or secondary pests of rice. For further details and a full list of possible rice-associated species, including occasional rice stemborers, see Table S1. Species of relatively localized occurrence are reported in Section 3.3.
Table 1.
Stemborer species identified as primary or secondary pests of rice. For further details and a full list of possible rice-associated species, including occasional rice stemborers, see Table S1. Species of relatively localized occurrence are reported in Section 3.3.
| Species | Status in Rice 1 | Mode of Attack in Rice |
|---|---|---|
| Africa and the Mediterranean | ||
| Crambidae | ||
| Chilo agamemnon Bleszynski | Primary species [40,41,42] | Rice age at planting has no effect on damage [43]; some resistance as plants age [44] |
| Chilo diffusilineus (J. de Joannis) | Some early reports as primary species; mainly on upland rice [45,46], more recently appears as a secondary species | NA |
| Chilo partellus (Swinhoe) | Primary species in upland rice in some regions, secondary species in irrigated rice [10,47,48]; maize and sorghum are the main crop hosts | Early instars feed on leaf sheaths before entering the stem; late instars located in upper internodes [38] |
| Chilo zacconius Bleszynski | Primary species [49] | Mainly affects mid- to late tillering plants, some resistance as plants age [50] |
| Noctuidae | ||
| Sesamia calamistis (Hampson) | Secondary species [49,51] | Occurs at the onset of flowering towards harvest [47] |
| Sesamia nonagrioides (Lefèbvre) | Primary species in southern Europe [52]; maize is the main crop host | Highest densities occur at about the time of head emergence [52] |
| S. nonagrioides botanephaga 2 Tams and Bowden | Subspecies that occurs as a secondary rice pest in West Africa [32]; sugarcane is the main crop host | NA |
| Pyralidae | ||
| Maliarpha separatella Ragonot | Primary species [30,53,54] | Feeds on green tissues of the leaf sheath for about 5 days before entering the stem. Remains in a single stem, feeding in one or two internodes. Rarely cause whiteheads [38,55] |
| Diopsidae 3 | ||
| Diopsis apicalis Dalman | Secondary diopsid species [56,57] | Young plants [56,58] |
| Diopsis macrophthalma Dalman | Primary diopsid species; mainly in lowland, irrigated systems [56,59,60] | Young plants (<40 days after transplanting), feeds on meristems causing death of leaves and stems [38,58] |
| Asia and Oceania | ||
| Crambidae | ||
| Chilo auricilius Dudgeon | Some early reports as primary species [61]; more recently regarded as a secondary species [22,62]; shows little preference among crop hosts [63]; but is more commonly regarded as a sugarcane pest [64] | Prevalent in rice that is adjacent to other crop hosts, with the 2nd generation attacking rice [65,66] |
| Chilo polychrysus (Meyrick) | Primary species in some parts of Malaysia; mainly occurs as a secondary species [23,24,67,68] | Occurs in deepwater rice before flooding and at about harvest [69]; more prevalent at tillering in irrigated rice [70] |
| Chilo suppressalis (Walker) | Primary species [7,8,9,12,22,69,71] | Occurs at mid- to late tillering, poor development on mature plants; prevalent on thick-stemmed plants [72,73] |
| Scirpophaga incertulas (Walker) | Primary species; also prevalent in deepwater rice [7,8,9,22] | Occur at early to late tillering; prefers high-tillering plants [72,73]; eggs and pupae capable of surviving submergence [22,71] |
| Scirpophaga innotata (Walker) | Primary species; secondary species in some regions [22,62,74] | Occurs at early to late tillering [75]; capable of surviving prolonged drought through diapause [22,76] |
| Noctuidae | ||
| Bathytricha truncata (Walker) | Primary species [77] | NA |
| Sesamia inferens (Walker) | Secondary species [22,67,73]; maize is the main crop host | Occurs at late crop stages occupying internodes near the plant base; prevalent in long-duration varieties [22,73] |
| North America, South America, and the Caribbean | ||
| Crambidae | ||
| Chilo plejadellus Zincken | Primary species [26,78,79] | Ealy larvae damage leaves and leaf sheaths, older larvae bore into the stems [27,79] |
| Diatraea saccharalis (Fabricius) | Occurs as the only stemborer in some parts of its range; prevalent in upland rice in some parts of northern Brazil [80], secondary species in the humid tropics [81,82,83,84]; sugarcane is the main crop host | Prevalent during stem elongation and panicle initiation; oviposition preference for older plants [84,85] |
| Eoreuma loftini (Dyar) | Primary species [86,87]; sugarcane is preferred for oviposition [88] | Prefers older vegetative plants for oviposition; larvae occur about 20 cm above the plant base [89,90] |
| Rupela albinella (Cramer) | Primary species in irrigated rice [83,91] | Prefers older vegetative plants for oviposition, will oviposit on younger plants but plants 40–60 days of age have optimal lumen space; can occur in internodes below water [83,92] |
| Pyralidae | ||
| Elasmopalpus lignosellus (Zeller) | Primary species in upland rice [93] | Eggs are deposited below the soil surface, larvae bore into the stem and tunnel upwards [93,94]; can attack and kill seedlings before tillering [93] |
1: Status in rice is based on reports of prominence; for details on occasional species see Table S1 and associated references [9,10,22,23,30,33,34,35,40,41,45,46,47,49,51,52,57,58,59,61,69,70,72,77,78,82,83,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112,113,114,115,116,117,118,119,120,121,122,123,124,125,126,127,128,129,130,131,132,133,134,135,136,137,138,139]; for information on occurrence in other crops see Table S2. 2: Sesamia nonagrioides botanephaga Tams and Bowden is considered a subspecies. 3: Diopsidae are a family of flies (Diptera), all other species on this table are Lepidoptera.
Table 2.
Host ranges of the 21 main rice stemborer species based on reported associations. Numbers indicate the total number of stemborer-associated plants as listed in Table S2 1.
Table 2.
Host ranges of the 21 main rice stemborer species based on reported associations. Numbers indicate the total number of stemborer-associated plants as listed in Table S2 1.
| Stemborer Species | Poaceae (Grasses) 2 | Other Monocots (e.g., Sedges and Reeds) 2 | Eudicots 2 | Crop Species | Total |
|---|---|---|---|---|---|
| Africa | |||||
| C. agamemnon | 8 | 1 | 5 | 9 | |
| C. diffusilineus | 8 | 6 | 8 | ||
| C. partellus | 34 | 8 | 34 | ||
| C. zacconius | 13 | 4 | 13 | ||
| D. apicalis | 5 | 2 | 3 | 7 | |
| D. macrophthalma | 24 | 1 | 3 | 25 | |
| M. separatella | 10 | 4 | 10 | ||
| S. calamistis | 54 | 3 | 7 | 57 | |
| S. nonagrioides | 20 | 9 | 6 | 29 | |
| S. nonagrioides botanephaga | 13 | 1 | 5 | 14 | |
| Asia and Oceania | |||||
| B. truncata | 10 | 4 | 10 | ||
| C. auricilius | 16 | 1 | 5 | 17 | |
| C. polychrysus | 30 | 2 | 7 | 32 | |
| C. suppressalis | 53 | 4 | 6 | 15 | 63 |
| S. incertulas | 47 | 12 | 1 | 6 | 60 |
| S. inferens | 74 | 11 | 2 | 15 | 87 |
| S. innotata | 11 | 3 | 4 | 14 | |
| Americas | |||||
| C. plejadellus | 6 | 3 | 6 | ||
| D. saccharalis | 75 | 8 | 8 | 83 | |
| E. lignosellus | 21 | 2 | 12 | 20 | 35 |
| E. loftini | 31 | 3 | 8 | 35 | |
| R. albinella | 3 | 1 | 3 |
1: See Tables S2 and S3 for full information on potential host plants and related references [9,10,33,37,38,41,45,46,55,56,58,63,64,65,66,67,83,88,89,90,94,95,96,100,101,107,123,128,129,140,141,143,144,145,146,147,148,149,150,151,152,153,154,155,156,157,158,159,160,161,162,163,164,165,166,167,168,169,170,171,172,173,174,175,176,177,178,179,180,181,182,183,184,185,186,187,188,189,190]; 2: Includes crop species.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Horgan, F.G. The Structure of Rice Stemborer Assemblages: A Review of Species’ Distributions, Host Ranges, and Interspecific Interactions. Insects 2023, 14, 921. https://doi.org/10.3390/insects14120921
AMA Style
Horgan FG. The Structure of Rice Stemborer Assemblages: A Review of Species’ Distributions, Host Ranges, and Interspecific Interactions. Insects. 2023; 14(12):921. https://doi.org/10.3390/insects14120921
Chicago/Turabian StyleHorgan, Finbarr G. 2023. "The Structure of Rice Stemborer Assemblages: A Review of Species’ Distributions, Host Ranges, and Interspecific Interactions" Insects 14, no. 12: 921. https://doi.org/10.3390/insects14120921
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.