
The Complete Mitogenomes of Three Grasshopper Species with Special Notes on the Phylogenetic Positions of Some Related Genera

 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Taxon Sampling
2.2. Sequencing, Assembly and Annotation
2.3. Phylogenetic Analyses
3. Results
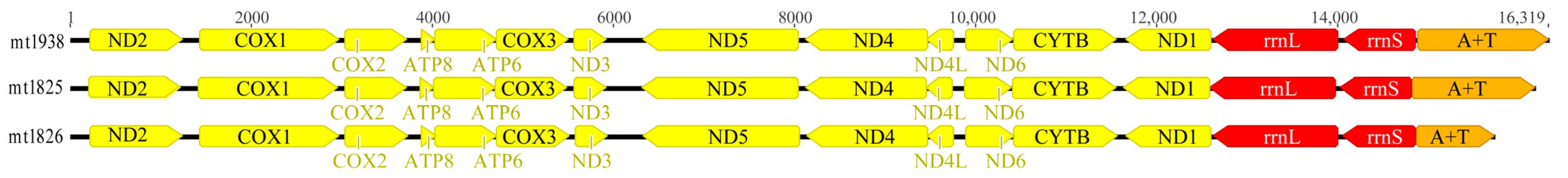
3.1. Characteristics of the Newly Sequenced Mitogenomes
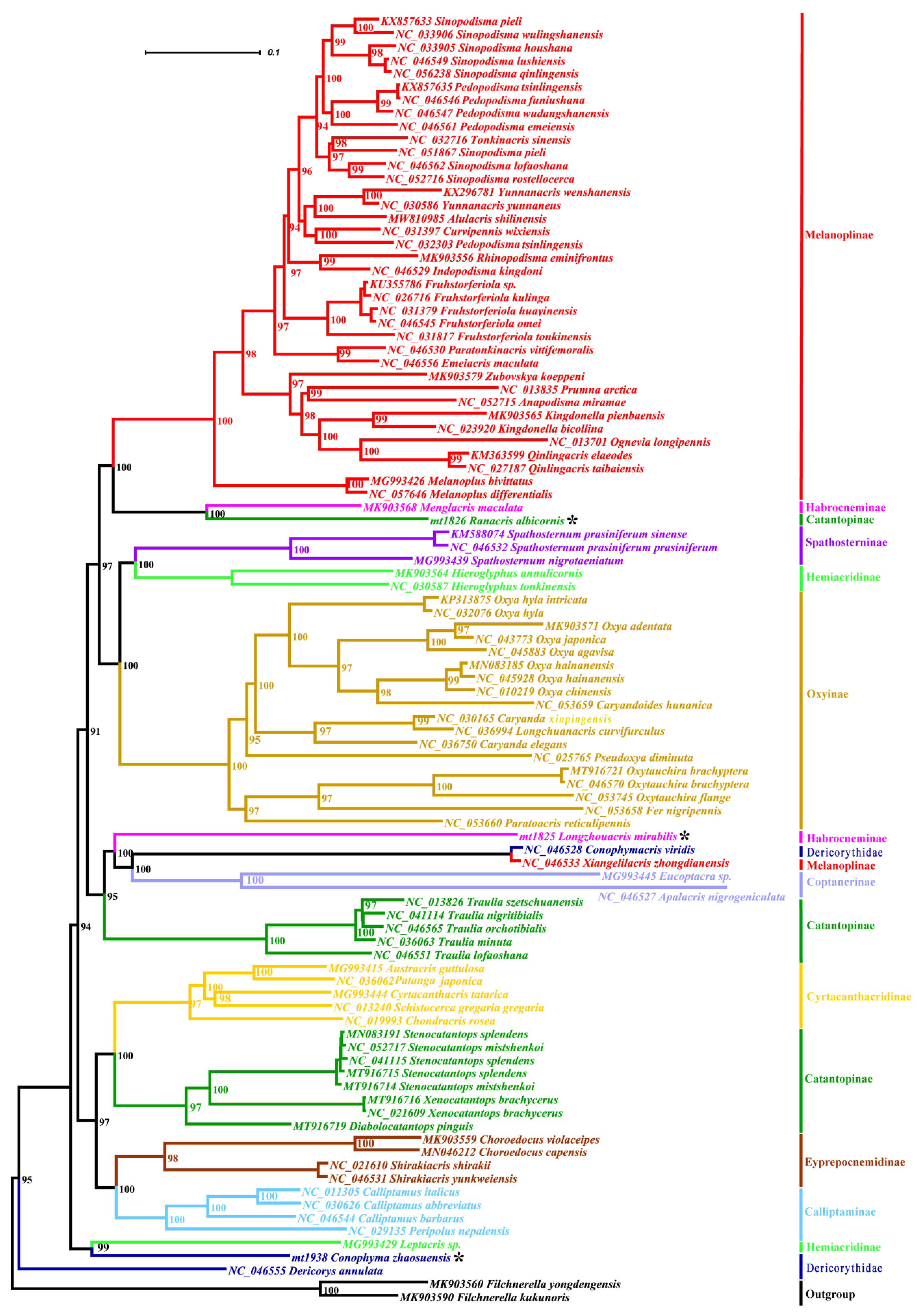
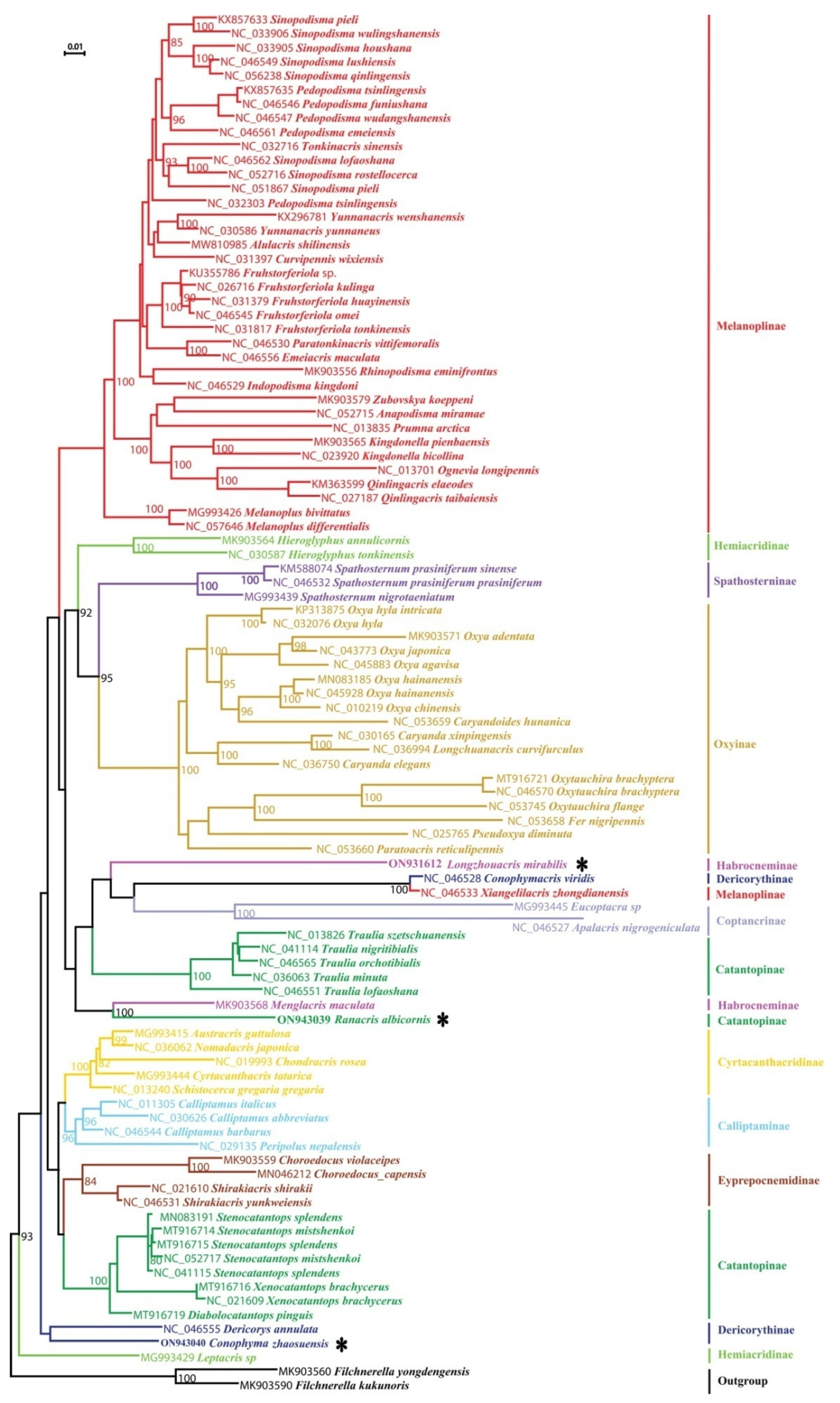
3.2. Phylogeny
4. Discussion
4.1. Phylogenetic Position of the Genus Emeiacris
4.2. Phylogenetic Position of the Genus Choroedocus
4.3. Phylogenetic Relationships among Some Related Genera
4.3.1. Non-Monophyly of the Family Dericorythidae
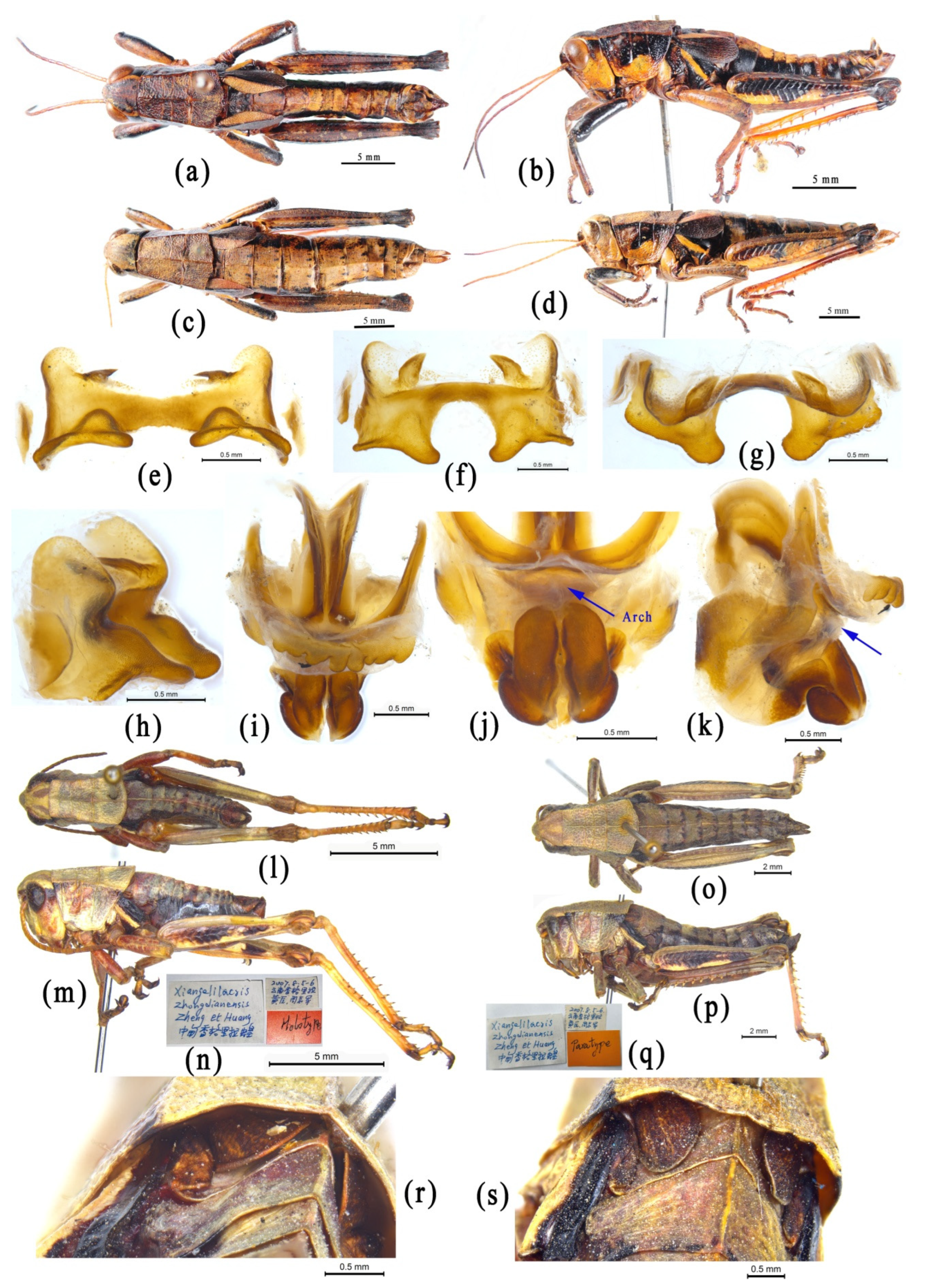
4.3.2. Phylogenetic Relationship between the Genera Conophymacris and Xiangelilacris
4.3.3. Phylogenetic Relationship between the Genera Menglacris, Ranacris and Longzhouacris
4.4. Performance of the Mitochondrial COX1 Gene in Reconstructing Phylogeny
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lecoq, M.; Zhang, L. Encyclopedia of Pest Orthoptera of the World; China Agricultural University Press: Beijing, China, 2019. [Google Scholar]
- Mugleston, J.; Naegle, M.; Song, H.; Bybee, S.M.; Ingley, S.; Suvorov, A.; Whiting, M.F. Reinventing the leaf: Multiple origins of leaf-like wings in katydids (Orthoptera: Tettigoniidae). Invertebr. Syst. 2016, 30, 335–352. [Google Scholar] [CrossRef]
- Li, X.-D.; Jiang, G.-F.; Yan, L.-Y.; Li, R.; Mu, Y.; Deng, W.-A. Positive selection drove the adaptation of mitochondrial genes to the demands of flight and high-altitude environments in grasshoppers. Front. Genet. 2018, 9, 605. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.P.; Zheng, F.Y.; Bai, J.; Wang, J.M.; Lv, C.Y.; Li, X.; Zhi, Y.C.; Li, X.J. Comparative analysis of mitogenomes among six species of grasshoppers (Orthoptera: Acridoidea: Catantopidae) and their phylogenetic implications in wing-type evolution. Int. J. Biol. Macromol. 2020, 159, 1062–1072. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Béthoux, O.; Shin, S.; Donath, A.; Letsch, H.; Liu, S.; McKenna, D.D.; Meng, G.; Misof, B.; Podsiadlowski, L.; et al. Phylogenomic analysis sheds light on the evolutionary pathways towards acoustic communication in Orthoptera. Nat. Commun. 2020, 11, 4939. [Google Scholar] [CrossRef]
- Huang, J.-P.; Hill, J.; Ortego, J.; Knowles, L.L. Paraphyletic species no more—Genomic data resolve a Pleistocene radiation and validate morphological species of the Melanoplus scudderi complex (Insecta: Orthoptera). Syst. Entomol. 2020, 45, 594–605. [Google Scholar] [CrossRef]
- Gu, J.X.; Jiang, B.; Wang, H.J.; Wei, T.; Lin, L.L.; Huang, Y.; Huang, J.H. Phylogeny and species delimitation of the genus Longgenacris and Fruhstorferiola viridifemorata species group (Orthoptera: Acrididae: Melanoplinae) based on molecular evidence. PLoS ONE 2020, 15, e0237882. [Google Scholar] [CrossRef]
- Hemp, C.; Scherer, C.; Brandl, R.; Pinkert, S. The origin of the endemic African grasshopper family Lentulidae (Orthoptera: Acridoidea) and its climate-induced diversification. J. Biogeogr. 2020, 47, 1805–1815. [Google Scholar] [CrossRef]
- Wang, H.; Jiang, B.; Gu, J.X.; Wei, T.; Lin, L.L.; Huang, Y.; Liang, D.; Huang, J.H. Molecular phylogeny and species delimitation of the genus Tonkinacris (Orthoptera, Acrididae, Melanoplinae) from China. PLoS ONE 2021, 16, e0249431. [Google Scholar] [CrossRef]
- Dey, L.-S.; Hochkirch, A.; Moussi, A.; Simões, M.V.P.; Husemann, M. Diversification in and around the Atlas Mountains: Insights into the systematics and biogeography of the genus Thalpomena (Orthoptera: Acrididae: Oedipodinae). Syst. Entomol. 2022, 47, 402–419. [Google Scholar] [CrossRef]
- Fenn, J.D.; Song, H.; Cameron, S.L.; Whiting, M.F. A preliminary mitochondrial genome phylogeny of Orthoptera (Insecta) and approaches to maximizing phylogenetic signal found within mitochondrial genome data. Mol. Phylogenet. Evol. 2008, 49, 59–68. [Google Scholar] [CrossRef]
- Erler, S.; Ferenz, H.J.; Moritz, R.F.A.; Kaatz, H.H. Analysis of the mitochondrial genome of Schistocerca gregaria gregaria (Orthoptera: Acrididae). Biol. J. Linn. Soc. 2010, 99, 296–305. [Google Scholar] [CrossRef]
- Sheffield, N.C.; Hiatt, K.D.; Valentine, M.C.; Song, H.; Whiting, M.F. Mitochondrial genomics in Orthoptera using MOSAS. Mitochondrial DNA 2010, 21, 87–104. [Google Scholar] [CrossRef] [PubMed]
- Leavitt, J.R.; Hiatt, K.D.; Whiting, M.F.; Song, H. Searching for the optimal data partitioning strategy in mitochondrial phylogenomics: A phylogeny of Acridoidea (Insecta: Orthoptera: Caelifera) as a case study. Mol. Phylogenet. Evol. 2013, 67, 494–508. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.L.; Huang, Y.; Lin, L.L.; Wang, X.Y.; Zheng, Z.M. The phylogeny of the Orthoptera (Insecta) as deduced from mitogenomic gene sequences. Zool. Stud. 2013, 52, 37. [Google Scholar] [CrossRef]
- Song, H.; Amédégnato, C.; Cigliano, M.M.; Desutter-Grandcolas, L.; Heads, S.W.; Huang, Y.; Otte, D.; Whiting, M.F. 300 million years of diversification: Elucidating the patterns of orthopteran evolution based on comprehensive taxon and gene sampling. Cladistics 2015, 31, 621–651. [Google Scholar] [CrossRef]
- Song, H.; Mariño-Pérez, R.; Woller, D.A.; Cigliano, M.M. Evolution, diversification, and biogeography of grasshoppers (Orthoptera: Acrididae). Insect Syst. Divers. 2018, 2, 3. [Google Scholar] [CrossRef]
- Liu, H.X.; Yan, L.Y.; Jiang, G.F. The complete mitochondrial genome of the jumping grasshopper Sinopodisma pieli (Orthoptera: Acrididae) and the phylogenetic analysis of Melanoplinae. Zootaxa 2017, 4363, 506–520. [Google Scholar] [CrossRef]
- Zhou, Z.J.; Zhao, L.; Liu, N.; Guo, H.F.; Guan, B.; Di, J.X.; Shi, F.M. Towards a higher-level Ensifera phylogeny inferred from mitogenome sequences. Mol. Phylogenet. Evol. 2017, 108, 22–33. [Google Scholar] [CrossRef]
- Li, R.; Shu, X.; Li, X.; Meng, L.; Li, B. Comparative mitogenome analysis of three species and monophyletic inference of Catantopinae (Orthoptera: Acridoidea). Genomics 2019, 111, 1728–1735. [Google Scholar] [CrossRef]
- Li, R.; Shu, X.; Deng, W.; Meng, L.; Li, B. Complete mitochondrial genome of Atractomorpha sagittaris (Orthoptera: Pyrgomorphidae) and its phylogenetic analysis for Acrididea. Biologia 2020, 75, 1571–1583. [Google Scholar] [CrossRef]
- Li, R.; Wang, Y.; Shu, X.; Meng, L.; Li, B. Complete mitochondrial genomes of three Oxya grasshoppers (Orthoptera) and their implications for phylogenetic reconstruction. Genomics 2020, 112, 289–296. [Google Scholar] [CrossRef]
- Mariño-Pérez, R.; Song, H. On the origin of the New World Pyrgomorphidae (Insecta: Orthoptera). Mol. Phylogenet. Evol. 2019, 139, 106537. [Google Scholar] [CrossRef]
- Chang, H.; Nie, Y.; Zhang, N.; Zhang, X.; Sun, H.; Mao, Y.; Qiu, Z.Y.; Huang, Y. MtOrt: An empirical mitochondrial amino acid substitution model for evolutionary studies of Orthoptera insects. BMC Evol. Biol. 2020, 20, 57. [Google Scholar] [CrossRef]
- Chang, H.; Qiu, Z.; Yuan, H.; Wang, X.; Li, X.; Sun, H.; Guo, X.; Lu, Y.; Feng, X.; Majid, M.; et al. Evolutionary rates of and selective constraints on the mitochondrial genomes of Orthoptera insects with different wingtypes. Mol. Phylogenet. Evol. 2020, 145, 106734. [Google Scholar] [CrossRef]
- Qiu, Z.Y.; Chang, H.H.; Yuan, H.; Huang, Y.; Lu, H.M.; Li, X.; Gou, X.C. Comparative mitochondrial genomes of four species of Sinopodisma and phylogenetic implications (Orthoptera, Melanoplinae). ZooKeys 2020, 969, 23–42. [Google Scholar] [CrossRef]
- Xu, H.Y.; Mao, B.Y.; Storozhenko, S.Y.; Huang, Y.; Chen, Z.L.; Huang, J.H. Phylogenetic position of the genus Alulacris (Orthoptera: Acrididae: Melanoplinae: Podismini) revealed by complete mitogenome evidence. Insects 2021, 12, 918. [Google Scholar] [CrossRef]
- Zahid, S.; Mariño-Pérez, R.; Song, H. Molecular phylogeny of the grasshopper family Pyrgomorphidae (Caelifera, Orthoptera) reveals rampant paraphyly and convergence of traditionally used taxonomic characters. Zootaxa 2021, 4969, 101–118. [Google Scholar] [CrossRef]
- Zeng, X.; Xu, H.Y.; Gu, J.X.; Mao, B.Y.; Chen, Z.L.; Huang, Y.; Huang, J.H. Phylogenetic position of the genera Caryandoides, Paratoacris, Fer and Longchuanacris (Orthoptera: Acrididae) revealed by complete mitogenome sequences. Invertebr. Syst. 2021, 35, 725–741. [Google Scholar] [CrossRef]
- Zheng, F.-Y.; Shi, Q.-Y.; Ling, Y.; Chen, J.-Y.; Zhang, B.-F.; Li, X.-J. Comparative analysis of mitogenomes among five species of Filchnerella (Orthoptera: Acridoidea: Pamphagidae) and their phylogenetic and taxonomic implications. Insects 2021, 12, 605. [Google Scholar] [CrossRef]
- You, Q.J.; Li, T.S.; Bi, D.Y. Descriptions of new genera and species of Catantopidae from Guangxi (Orthoptera: Acridoidea). Entomotaxonomia 1983, 5, 165–181. [Google Scholar]
- Cigliano, M.M.; Braun, H.; Eades, D.C.; Otte, D. Orthoptera Species File. Version 5.0/5.0. 2022. Available online: http://Orthoptera.SpeciesFile.org/HomePage/Orthoptera/HomePage.aspx (accessed on 10 November 2022).
- Li, H.C.; Xia, K.L. Fauna Sinica, Insecta, Vol. 43, Orthoptera, Acridoidea, Catantopidae; Science Press: Beijing, China, 2006. [Google Scholar]
- Mao, B.Y.; Ren, G.D.; Ou, X.H. Fauna, Distribution Pattern and Adaptability on Acridoidea from Yunnan; China Forestry Publishing House: Beijing, China, 2011. [Google Scholar]
- You, Q.J.; Lin, R.Z. Description of a new genus and new species of Catantopidae from Guangxi Province, China. Entomotaxonomia 1983, 5, 255–258. [Google Scholar]
- Zheng, Z.M.; Lin, L.L.; Deng, W.A.; Shi, F.M. Two new species of Catantopidae (Orthoptera: Acridoidea) from China. Entomotaxonomia 2015, 37, 177–181. [Google Scholar] [CrossRef]
- You, Q.J.; Li, T.S.; Zhang, Y.Q.; Lin, R.Z. Economic Insect Iconography of Guangxi, Phytophage Insects; Guangxi Press of Science & Technology: Nanning, China, 1990. [Google Scholar]
- Jiang, G.F.; Zheng, Z.M. Grasshoppers and Locusts from Guangxi; Guangxi Normal University Press: Guilin, China, 1998. [Google Scholar]
- Storozhenko, S.Y. A new species of the genus Mesambria Stål, 1878 with notes on the tribe Mesambriini (Orthoptera: Acrididae, Catantopinae). Zootaxa 2018, 4418, 55–65. [Google Scholar] [CrossRef]
- Otte, D. Orthoptera Species File 4. Grasshoppers (Acridomorpha) D; Orthopterists’ Society & Academy of Natural Sciences of Philadelphia: Philadelphia, PA, USA, 1995. [Google Scholar]
- Eades, D.C. Evolutionary relationships of phallic structures of Acridomorpha (Orthoptera). J. Orthoptera Res. 2000, 9, 181–210. [Google Scholar] [CrossRef]
- Uvarov, B.P. Grasshoppers and Locusts. A Handbook of General Acridology. Vol.1. Anatomy, Physiology, Development, Phase Polymorphism, IntroductiontoTaxonomy; University of Cambridge Press: Cambridge, UK, 1966. [Google Scholar]
- Storozhenko, S.Y.; Kim, T.W.; Jeon, M.J. Monograph of Korean Orthoptera; National Institute of Biological Resources: Incheon, Republic of Korea, 2015. [Google Scholar]
- Woller, D.A.; Song, H. Investigating the functional morphology of genitalia during copulation in the grasshopper Melanoplus rotundipennis (Scudder,1878) via correlative microscopy. J. Morph. 2017, 278, 334–359. [Google Scholar] [CrossRef]
- Han, H.; Wang, N.; Xu, L.; Gao, S.; Liu, A. The complete mitochondrial genome of Calliptamus abbreviatus Ikovnnikov (Orthoptera: Acridoidea). Mitochondrial DNA Part B 2016, 1, 770–771. [Google Scholar] [CrossRef]
- Zhi, Y.; Zhang, N.; Lu, X.; Yin, H.; Zhang, D. The complete mitochondrial genome of Peripolus nepalensis Uvarov, 1942 (Orthoptera: Acridoidea: Catantopidae). Mitochondrial DNA 2016, 27, 26–27. [Google Scholar] [CrossRef]
- Qiu, Z.Y.; Wang, Y.T.; Yuan, H.; Zhao, X.; Ru, N.Y.; Liu, L.; Cui, Y.Y.; Liu, Y.T. The complete mitochondrial genome of Traulia minuta Huang & Xia, 1985 (Orthoptera: Catantopinae). Mitochondrial DNA Part B 2021, 6, 2180–2181. [Google Scholar] [CrossRef]
- Yang, J.; Liu, Y.; Liu, N. The complete mitochondrial genome of the Xenocatantops brachycerus (Orthoptera: Catantopidae). Mitochondrial DNA Part A 2016, 27, 2844–2845. [Google Scholar] [CrossRef]
- Chang, H.; Huang, Y. The complete mitochondrial genome of the Hieroglyphus tonkinensis (Orthoptera: Acrididae). Mitochondrial DNA Part B 2016, 1, 534–535. [Google Scholar] [CrossRef]
- Chen, Z.N.; Xu, S.Q. The complete mitochondrial DNA genome sequence of a terrestrial grasshopper, Curvipennis wixiensis (Acrididae: Podismini). Conserv. Genet. Resour. 2017, 9, 115–118. [Google Scholar] [CrossRef]
- Liu, F.; Qiu, Z. The complete mitochondrial genome of Fruhstorferiola huayinensis (Orthoptera: Catantopidae). Mitochondrial DNA Part B 2016, 1, 273–274. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Guan, D.L.; Xu, S.-Q. Complete mitochondrial genome of the Chinese endemic grasshopper Fruhstorferiola kulinga (Orthoptera: Acrididae: Podismini). Mitochondrial DNA 2016, 27, 3240–3241. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.M.; Lin, L.L. The complete mitochondrial genome of Fruhstorferiola tonkinensis (Orthoptera: Catantopidae). Mitochondrial DNA Part B 2016, 1, 434–435. [Google Scholar] [CrossRef]
- Zhi, Y.; Liu, B.; Han, G.; Yin, H.; Zhang, D.C. The complete mitochondrial genome of Kingdonella bicollina (Orthoptera: Acridoidea: Catantopidae). Mol. Biol. Rep. 2016, 27, 611–619. [Google Scholar] [CrossRef]
- Sun, H.; Zheng, Z.; Huang, Y. Sequence and phylogenetic analysis of complete mitochondrial DNA genomes of two grasshopper species Gomphocerus rufus (Linnaeus, 1758) and Primnoa arctica (Zhang and Jin, 1985) (Orthoptera: Acridoidea). Mitochondrial DNA 2010, 21, 115–131. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Jiang, G.F.; Liang, A.P.; Zhong, X.T.; Liu, Y. Characterization of the mitochondrial genome of the montane grasshopper, Qinlingacris elaeodes (Orthoptera: Catantopidae). Mitochondrial DNA Part A 2016, 27, 1765–1766. [Google Scholar] [CrossRef]
- Zhang, X.; Li, X.; Liu, F.; Yuan, H.; Huang, Y. The complete mitochondrial genome of Tonkinacris sinensis (Orthoptera: Acrididae): A tRNA-like sequence and its implications for phylogeny. Biochem. Syst. Ecol. 2017, 70, 147–154. [Google Scholar] [CrossRef]
- Hu, Z.; Guan, D.L.; Mao, B.Y. Characterization of the complete mitochondrial genome of the Yunnan endemic grasshopper Yunnanacris yunnaneus (Insecta: Orthoptera: Acrididae). Conserv. Genet. Resour. 2016, 8, 267–270. [Google Scholar] [CrossRef]
- Yuan, H.; Qiu, Z.; Yang, C.; Huang, Y. The complete mitochondrial genome sequence of Caryanda elegans (Orthoptera: Acrididae). Mitochondrial DNA Part B 2019, 4, 1580–1581. [Google Scholar] [CrossRef]
- Hu, Z.; Guan, D.-L.; Mao, B.-Y. Description of a new species, Caryanda Stål, 1878 (Acrididae, Orthoptera) from China. Orient. Insects 2017, 51, 124–134. [Google Scholar] [CrossRef]
- Hu, Z.; Han, Y.-P.; Guan, D.-L.; Mao, B.-Y. Characterization of the complete mitochondrial genome of the Yunnan endemic grasshopper Longchuanacris curvifurculus (Insecta: Orthoptera: Catantopidae). Mitochondrial DNA Part B 2018, 3, 670–671. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.Y.; Huang, Y. Complete mitochondrial genome of Oxya chinensis (Orthoptera, Acridoidea). Acta Biochim. Biophys. Sin. 2008, 40, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Song, N.; Li, H.; Song, F.; Cai, W. Molecular phylogeny of Polyneoptera (Insecta) inferred from expanded mitogenomic data. Scient. Rep. 2016, 6, 36175. [Google Scholar] [CrossRef]
- Dong, J.J.; Guan, D.L.; Xu, S.Q. Complete mitogenome of the semiaquatic grasshopper Oxya intricata (Stål.) (Insecta: Orthoptera: Catantopidae). Mitochondrial DNA Part A 2016, 27, 3233–3234. [Google Scholar] [CrossRef]
- Tang, M.; Tan, M.H.; Meng, G.L.; Yang, S.Z.; Su, X.; Liu, S.L.; Song, W.H.; Li, Y.Y.; Wu, Q.; Zhang, A.B.; et al. Multiplex sequencing of pooled mitochondrial genomes—A crucial step toward biodiversity analysis using mito-metagenomics. Nucleic Acids Res. 2014, 42, e166. [Google Scholar] [CrossRef]
- Zhou, F.; Huang, Y. The complete mitochondrial genome of Spathosternum prasiniferum sinense Uvarov, 1931 (Orthoptera: Acridoidea: Acrididae). Mitochondrial DNA 2016, 27, 1932–1933. [Google Scholar] [CrossRef]
- Hahn, C.; Bachmann, L.; Chevreux, B. Reconstructing mitochondrial genomes directly from genomic next-generation sequencing reads—A baiting and iterative mapping approach. Nucleic Acids Res. 2013, 41, e129. [Google Scholar] [CrossRef]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol. Phylogenet. Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Darty, K.; Denise, A.; Ponty, Y. VARNA: Interactive drawing and editing of the RNA secondary structure. Bioinformatics 2009, 25, 1974–1975. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Ranwez, V.; Douzery, E.J.P.; Cambon, C.; Chantret, N.; Delsuc, F. MACSE v2: Tool kit for the alignment of coding sequences accounting for frame shifts and stop codons. Mol. Biol. Evol. 2018, 35, 2582–2584. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Res. 2020, 20, 34–355. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.-T.; Schmidt, H.A.; Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Trifinopoulos, J.; Nguyen, L.-T.; Haeseler, A.; Minh, B.Q. WIQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef]
- Hillis, D.M.; Bull, J.J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Zheng, Z.M. New genera and new species of grasshoppers from Yunnan, Guizhou and Sichuan, China. Acta Zootax. Sin. 1981, 6, 60–68. [Google Scholar]
- Zheng, Z.M. Grasshoppers from the Areas of Yunnan, Guizhou, Sichuan, Shanxi and Ningxia; Science Press: Beijing, China, 1985. [Google Scholar]
- Bolívar, I. Estudios Entomológicos Segunda Parte. I. El grupo de los Euprepocnemes. Trab. Mus. Nac. Cienc. Nat. Zool. 1914, 20, 1–40. [Google Scholar]
- Stål, C. Systema Acridiodeorum. Essai d’une systématisation des Acridiodées. Bih. K. Sven. Vet.-Akad. Handl. 1878, 5, 1–100. [Google Scholar]
- Brunner von Wattenwyl, C. Révision du système des orthoptères et description des espèces rapportées par M. Leonardo Fea de Birmanie. Ann. Mus. Civ. Stor. Nat. Genova Ser. 2 1893, 13, 1–230. [Google Scholar]
- Kirby, W.F. List of a small collection of orthopterous insects formed by Sir Harry Johnston in British East Africa and Uganda in 1899 and 1900, with descriptions of five new species. P. Gener. Meet. Scient. Bus. Zool. Soc. Lond. 1902, 1, 93–101. [Google Scholar]
- Willemse, C. Bijdrage tot de kennis der Orthoptera s.s. van den Nederlandsch Indischen Archipel en omliggende gebieden. Zool. Meded. 1921, 6, 1–44. [Google Scholar]
- Uvarov, B.P. Notes on the Orthoptera in the British Museum. 1. The group of Euprepocnemini. Trans. Entomol. Soc. Lond. 1921, 69, 106–144. [Google Scholar] [CrossRef]
- Tinkham, E.R. Taxonomic and biological studies of the Cyrtacanthacrinae of South China. Lingnan Sci. J. 1940, 19, 269–382. [Google Scholar]
- Xia, K.L. Synopsis of the Classification on the Chinese Acrididae [Taxonomic Essentials (Principal classification) of Acrididae from China]; Science Press: Beijing, China, 1958. [Google Scholar]
- Yin, X.C. Grasshoppers and Locusts from Qinghai-Xizang Plateau of China; Science Press: Beijing, China, 1984. [Google Scholar]
- Liu, J.P. (Ed.) Studies on Acridoidea of Hainan Island; Tianze Publishing House: Yangling, China, 1995. [Google Scholar]
- Owen, C.L.; Miller, G.L. Phylogenomics of the Aphididae: Deep relationships between subfamilies clouded by gene tree discordance, introgression and the gene tree anomaly zone. Syst. Entomol. 2022, 47, 470–486. [Google Scholar] [CrossRef]
- Lu, L.; Dietrich, C.H.; Cao, Y.H.; Zhang, Y.L. A multi-gene phylogenetic analysis of the leafhopper subfamily Typhlocybinae (Hemiptera: Cicadellidae) challenges the traditional view of the evolution of wing venation. Mol. Phylogenet. Evol. 2021, 165, 107299. [Google Scholar] [CrossRef]
- Huber, B.A.; Astrin, J.J. Increased sampling blurs morphological and molecular species limits: Revision of the Hispaniolan endemic spider genus Tainonia (Araneae: Pholcidae). Invert. Syst. 2009, 23, 281–300. [Google Scholar] [CrossRef]
- Vahtera, V.; Edgecombe, G.D.; Giribet, G. Phylogenetics of scolopendromorph centipedes: Can denser taxon sampling improve an artificial classification? Invert. Syst. 2013, 27, 578–602. [Google Scholar] [CrossRef]
- Cao, Y.; Dietrich, C.H.; Zahniser, J.N.; Dmitriev, D.A. Dense sampling of taxa and characters improves phylogenetic resolution among deltocephaline leafhoppers (Hemiptera: Cicadellidae: Deltocephalinae). Syst. Entomol. 2022, 47, 430–444. [Google Scholar] [CrossRef]
- Willemse, C. On a small collection of Orthoptera from the Chungking District, S.E. China. Nat. Maandbl. 1933, 22, 15–18. [Google Scholar]
- Mistshenko, L.L. Fauna of the USSR, Orthoptera 4 Catantopinae; Bey-Bienko, G.J., Mistshenko, L.L., Eds.; Zoological Institute Akademia Nauk SSSR (NS): St. Petersburg, Russia, 1952; Volume 54, pp. 1–610. [Google Scholar]
- Zheng, Z.M.; Huang, Y.; Zhou, Z.J. New genus and new species of grasshoppers from Hengduanshan Region, China (Orthoptera, Acridoidea). Acta Zootax. Sin. 2008, 33, 363–367. [Google Scholar]
- Jiang, G.-F.; Zheng, Z.-M. A new genus and species of Catantopidae (Orthoptera: Acridoidea) from Yunnan Province, China. Acta Entomol. Sin. 1994, 37, 463–467. [Google Scholar]
- Huang, J.H.; Zhang, A.B.; Mao, S.L.; Huang, Y. DNA barcoding and species boundary delimitation of selected species of Chinese Acridoidea (Orthoptera: Caelifera). PLoS ONE 2013, 8, e82400. [Google Scholar] [CrossRef]
- Bugrov, A.G.; Smyshlyaev, G.A.; Blinov, A.G. Phylogeny of grasshoppers (Orthoptera, Acrididae) based on the analysis of DNA sequences in COI mitochondrial gene. Euroasian Entomol. J. 2012, 11, 493–502. [Google Scholar]
- Saad, Y.M.; El-Sadek, H.E.-S.A. The Efficiency of cytochrome oxidase subunit 1 gene (cox1) in reconstruction of phylogenetic relations among some crustacean species. Int. J. Anim. Vet. Sci. 2017, 11, 515–520. [Google Scholar] [CrossRef]
- Hendrich, L.; Pons, J.; Ribera, I.; Balke, M. Mitochondrial cox1 sequence data reliably uncover patterns of insect diversity but suffer from high lineage-idiosyncratic error rates. PLoS ONE 2010, 5, e14448. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
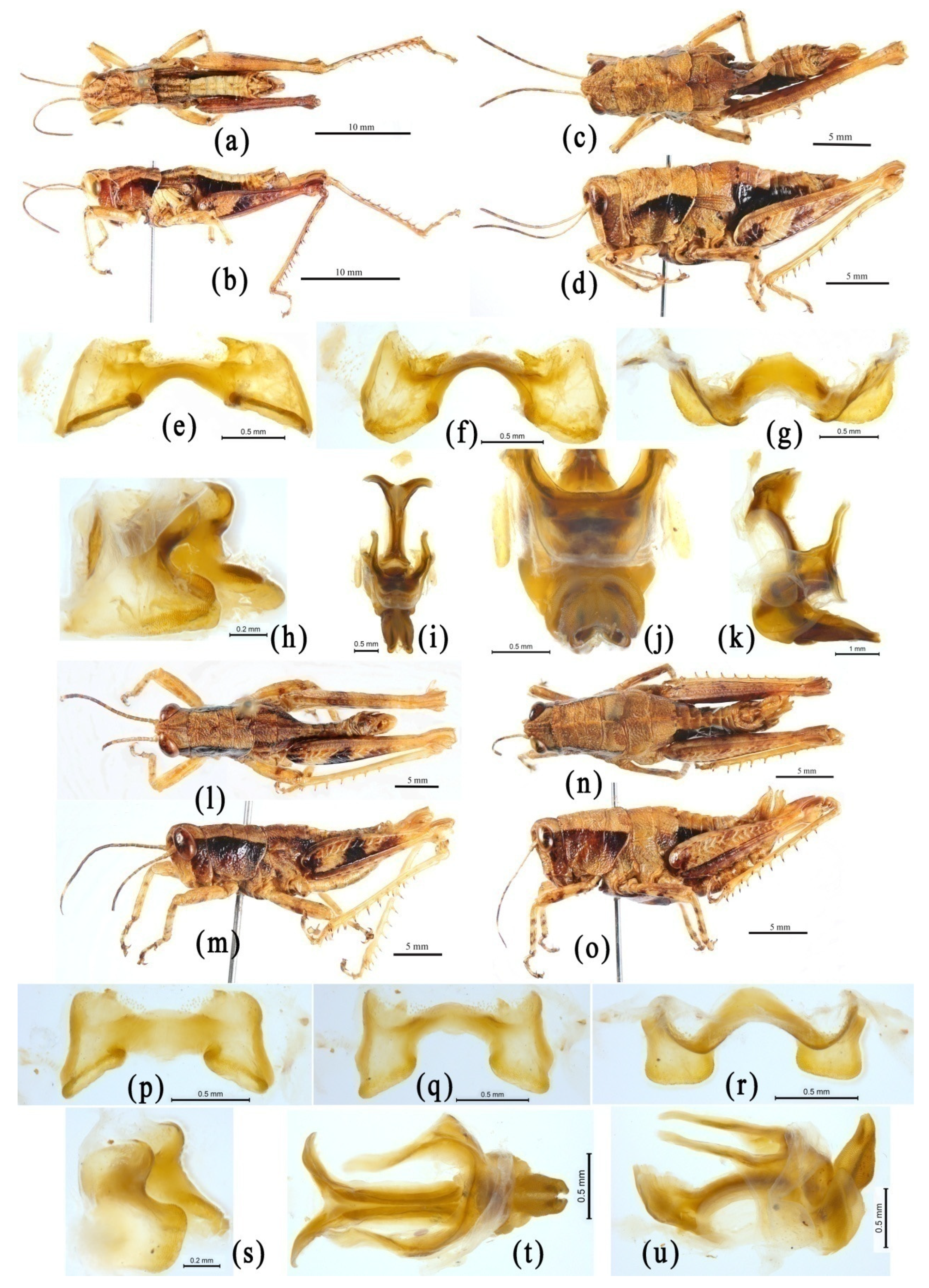{kind=link}
{kind=link}
| Species | Accession Number | Reference |
|---|---|---|
| Caelifera, Acridoidea, Acrididae | ||
| Calliptaminae | ||
| Calliptamus abbreviates | NC_030626 | Han et al. (2016) [45] |
| Calliptamus barbarous | NC_046544 | Chang et al. (2020) [24] |
| Calliptamus italicus | NC_011305 | Fenn et al. (2008) [11] |
| Peripolus nepalensis | NC_029135 | Zhi et al. (2016) [46] |
| Catantopinae | ||
| Diabolocatantops pinguis | MT916719 | Zeng et al. (2021) [29] |
| Ranacris albicornis | ON943039 | This study (mt1826) |
| Stenocatantops mistshenkoi | NC_052717 | Chen et al. (2020) [4] |
| MT916714 | Zeng et al. (2021) [29] | |
| Stenocatantops splendens | MN083191 | Chang et al. (2020) [24] |
| MT916715 | Zeng et al. (2021) [29] | |
| NC_041115 | Li et al. (2019) [20] | |
| Traulia lofaoshana | NC_046551 | Chang et al. (2020) [24] |
| Traulia minuta | NC_036063 | Qiu et al. (2021) [47] |
| Traulia nigritibialis | NC_041114 | Li et al. (2019) [20] |
| Traulia orchotibialis | NC_046565 | Chang et al. (2020) [24] |
| Traulia szetschuanensis | NC_013826 | Direct Submission |
| Xenocatantops brachycerus | MT916716 | Zeng et al. (2021) [29] |
| NC_021609 | Yang J. et al. (2016) [48] | |
| Coptancrinae | ||
| Apalacris nigrogeniculata | NC_046527 | Chang et al. (2020) [24] |
| Eucoptacra sp. | MG993445 | Song et al. (2018) [17] |
| Cyrtacanthacridinae | ||
| Austracris guttulosa | MG993415 | Song et al. (2018) [17] |
| Chondracris rosea | NC_019993 | Direct Submission |
| Cyrtacanthacris tatarica | MG993444 | Song et al. (2018) [17] |
| Patanga japonica | NC_036062 | Direct Submission |
| Schistocerca gregaria gregaria | NC_013240 | Erler et al. (2010) [12] |
| Eyprepocnemidinae | ||
| Choroedocus capensis | MN046212 | Chang et al. (2020) [24] |
| Choroedocus violaceipes | MK903559 | Chang et al. (2020) [24] |
| Shirakiacris shirakii | NC_021610 | Direct Submission |
| Shirakiacris yunkweiensis | NC_046531 | Chang et al. (2020) [24] |
| Habrocneminae | ||
| Longzhouacris mirabilis | ON931612 | This study (mt1825) |
| Menglacris maculata | MK903568 | Chang et al. (2020) [25] |
| Hemiacridinae | ||
| Hieroglyphus annulicornis | MK903564 | Chang et al. (2020) [25] |
| Hieroglyphus tonkinensis | NC_030587 | Chang and Huang (2016) [49] |
| Leptacris sp. | MG993429 | Song et al. (2018) [17] |
| Melanoplinae | ||
| Alulacris shilinensis | MW810985 | Xu et al. (2021) [27] |
| Anapodisma miramae | NC_052715 | Chen et al. (2020) [4] |
| Curvipennis wixiensis | NC_031397 | Chen and Xu (2017) [50] |
| Emeiacris maculate | NC_046556 | Chang et al. (2020) [24] |
| Fruhstorferiola huayinensis | NC_031379 | Liu and Qiu (2016) [51] |
| Fruhstorferiola kulinga | NC_026716 | Yang R. et al. (2016) [52] |
| Fruhstorferiola omei | NC_046545 | Chang et al. (2020) [24] |
| Fruhstorferiola sp. | KU355786 | Direct submission |
| Fruhstorferiola tonkinensis | NC_031817 | Zhang and Lin (2016) [53] |
| Indopodisma kingdoni | NC_046529 | Chang et al. (2020) [24] |
| Kingdonella bicollina | NC_023920 | Zhi et al. (2016) [54] |
| Kingdonella pienbaensis | MK903565 | Chang et al. (2020) [25] |
| Melanoplus bivittatus | MG993426 | Song et al. (2018) [17] |
| Melanoplus differentialis | NC_057646 | Direct Submission |
| Ognevia longipennis | NC_013701 | Direct Submission |
| Paratonkinacris vittifemoralis | NC_046530 | Chang et al. (2020) [24] |
| Pedopodisma emeiensis | NC_046561 | Chang et al. (2020) [24] |
| Pedopodisma funiushana | NC_046546 | Chang et al. (2020) [24] |
| Pedopodisma tsinlingensis | KX857635 | Qiu et al. (2020) [26] |
| NC_032303 | Direct Submission | |
| Pedopodisma wudangshanensis | NC_046547 | Chang et al. (2020) [24] |
| Prumna arctica | NC_013835 | Sun et al. (2010) [55] |
| Qinlingacris elaeodes | KM363599 | Li et al. (2016) [56] |
| Qinlingacris taibaiensis | NC_027187 | Direct Submission |
| Rhinopodisma eminifrontus | MK903556 | Chang et al. (2020) [25] |
| Sinopodisma houshana | NC_033905 | Qiu et al. (2020) [26] |
| Sinopodisma lofaoshana | NC_046562 | Chang et al. (2020) [24] |
| Sinopodisma lushiensis | NC_046549 | Chang et al. (2020) [24] |
| Sinopodisma pieli | KX857633 | Qiu et al. (2020) [26] |
| NC_051867 | Liu et al. (2017) [18] | |
| Sinopodisma qinlingensis | NC_056238 | Qiu et al. (2020) [26] |
| Sinopodisma rostellocerca | NC_052716 | Chen et al. (2020) [4] |
| Sinopodisma wulingshanensis | NC_033906 | Qiu et al. (2020) [26] |
| Tonkinacris sinensis | NC_032716 | Zhang et al. (2017) [57] |
| Xiangelilacris zhongdianensis | NC_046533 | Chang et al. (2020) [24] |
| Yunnanacris wenshanensis | KX296781 | Direct Submission |
| Yunnanacris yunnaneus | NC_030586 | Hu et al. (2016) [58] |
| Zubovskya koeppeni | MK903579 | Chang et al. (2020) [25] |
| Oxyinae | ||
| Caryanda elegans | NC_036750 | Yuan et al. (2019) [59] |
| Caryanda xinpingensis | NC_030165 | Hu et al. (2017) [60] |
| Caryandoides hunanica | NC_053659 | Zeng et al. (2021) [29] |
| Fer nigripennis | NC_053658 | Zeng et al. (2021) [29] |
| Longchuanacris curvifurculus | NC_036994 | Hu et al. (2018) [61] |
| Oxya adentata | MK903571 | Chang et al. (2020) [25] |
| Oxya agavisa | NC_045883 | Li et al. (2020) [22] |
| Oxya chinensis | NC_010219 | Zhang and Huang (2008) [62] |
| Oxya hainanensis | MN083185 | Chang et al. (2020) [24] |
| NC_045928 | Li et al. (2020) [22] | |
| Oxya hyla | NC_032076 | Song et al. (2016) [63] |
| Oxya hyla intricate | KP313875 | Dong et al. (2016) [64] |
| Oxya japonica | NC_043773 | Li et al. (2020) [22] |
| Oxytauchira brachyptera | MT916721 | Zeng et al. (2021) [29] |
| NC_046570 | Chang et al. (2020a) [24] | |
| Oxytauchira flange | NC_053745 | Zeng et al. (2021) [29] |
| Paratoacris reticulipennis | NC_053660 | Zeng et al. (2021) [29] |
| Pseudoxya diminuta | NC_025765 | Tang et al. (2014) [65] |
| Spathosterninae | ||
| Spathosternum nigrotaeniatum | MG993439 | Song et al. (2018) [17] |
| Spathosternum prasiniferum sinense | KM588074 | Zhou et al. (2016) [66] |
| Spathosternum prasiniferum prasiniferum | NC_046532 | Chang et al. (2020) [24] |
| Dericorythidae | ||
| Conophyminae | ||
| Conophyma zhaosuensis | ON943040 | This study (mt1938) |
| Conophymacris viridis | NC_046528 | Chang et al. (2020) [24] |
| Dericorythinae | ||
| Dericorys annulata | NC_046555 | Chang et al. (2020) [24] |
| Pamphagidae | ||
| Pamphaginae | ||
| Filchnerella kukunoris | MK903590 | Chang et al. (2020) [25] |
| Filchnerella yongdengensis | MK903560 | Chang et al. (2020) [25] |
| PCGs | Initiation Codons | Termination Codons | ||||
|---|---|---|---|---|---|---|
| mt1825 | mt1826 | mt1938 | mt1825 | mt1826 | mt1938 | |
| ND2 | ATG | ATG | ATG | TAA | TAA | T |
| COX1 | ACC | ACC | CAA | T | T | T |
| COX2 | ATG | ATG | ATG | TAA | TAA | TAA |
| ATP8 | ATT | ATT | ATC | TAA | TAA | TAA |
| APT6 | GTG | ATG | ATG | TAA | TAA | TAA |
| COX3 | ATG | ATG | ATG | TAA | TAA | TAA |
| ND3 | ATG | ATT | ATT | TAA | TAA | TAA |
| ND5 | ATT | ATT | ATT | TAA | TAA | T |
| ND4 | ATG | ATG | ATG | T | TAA | T |
| ND4L | ATG | ATG | ATG | TAA | TAA | TAA |
| ND6 | ATG | ATG | ATG | TAA | TAA | TA |
| CYTB | ATG | ATG | ATG | TA | TAA | TAA |
| ND1 | ATA | ATA | ATG | TAG | TAG | TAG |
| Species | A–A | A–G | A–C | G–U | C–U | U–U |
|---|---|---|---|---|---|---|
| mt1825 | 1(trnD) | 1(trnW) | 0 | 28 | 0 | 4(trnQ,trnD,trnH,trnS2) |
| mt1826 | 1(trnD) | 1(trnW) | 0 | 22 | 0 | 4(trnQ,trnE) |
| mt1938 | 1(trnD) | 1(trnW) | 0 | 16 | 0 | 3(trnI, trnQ, trnH) |
| Transfer RNA | mt1825 | mt1826 | mt1938 | Transfer RNA | mt1825 | mt1826 | mt1938 |
|---|---|---|---|---|---|---|---|
| trnI | 1 | 0 | 0 | trnR | 1 | 1 | 1 |
| trnQ | 1 | 1 | 1 | trnN | 0 | 0 | 0 |
| trnM | 0 | 0 | 0 | trnS1 | 1 | 1 | 2 |
| trnW | 0 | 0 | 0 | trnE | 1 | 0 | 1 |
| trnC | 2 | 1 | 0 | trnF | 5 | 2 | 2 |
| trnY | 2 | 2 | 2 | trnH | 1 | 3 | 0 |
| trnL2 | 1 | 1 | 0 | trnT | 1 | 1 | 0 |
| trnD | 0 | 0 | 0 | trnP | 2 | 2 | 2 |
| trnK | 1 | 1 | 0 | trnS2 | 1 | 0 | 0 |
| trnG | 1 | 1 | 2 | trnL1 | 1 | 1 | 0 |
| trnA | 3 | 2 | 2 | trnV | 2 | 2 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Mao, B.; Wang, H.; Dai, L.; Huang, Y.; Chen, Z.; Huang, J. The Complete Mitogenomes of Three Grasshopper Species with Special Notes on the Phylogenetic Positions of Some Related Genera. Insects 2023, 14, 85. https://doi.org/10.3390/insects14010085
Zhang C, Mao B, Wang H, Dai L, Huang Y, Chen Z, Huang J. The Complete Mitogenomes of Three Grasshopper Species with Special Notes on the Phylogenetic Positions of Some Related Genera. Insects. 2023; 14(1):85. https://doi.org/10.3390/insects14010085
Chicago/Turabian StyleZhang, Chulin, Benyong Mao, Hanqiang Wang, Li Dai, Yuan Huang, Zhilin Chen, and Jianhua Huang. 2023. "The Complete Mitogenomes of Three Grasshopper Species with Special Notes on the Phylogenetic Positions of Some Related Genera" Insects 14, no. 1: 85. https://doi.org/10.3390/insects14010085
APA StyleZhang, C., Mao, B., Wang, H., Dai, L., Huang, Y., Chen, Z., & Huang, J. (2023). The Complete Mitogenomes of Three Grasshopper Species with Special Notes on the Phylogenetic Positions of Some Related Genera. Insects, 14(1), 85. https://doi.org/10.3390/insects14010085