3.3. Taxonomy
Key to genera Gymnaetoides, Pseudotachycines, and Homotachycines
- -
The apical area of the dorsal median lobe of the male genitalia does not divide; the dorsal lateral lobe of the male genitalia is much or little longer than the dorsal median lobe, in which the apical area is weakly sclerotized…………………………….……...2
- 2.
The male paraproct is prolonged; the dorsal lateral lobe of the male genitalia is much longer than the dorsal median lobe, in which the apical area is almost as wide as the basal area………………………………………..………………...…...Pseudotachycines
- -
The male paraproct is normally developed; the dorsal lateral lobe of the male genitalia is little longer than the dorsal median lobe, in which the apical area is much narrower than the basal area……………………………………………... Homotachycines
Genus Gymnaetoides Qin, Liu & Li, 2017
Gymnaetoides Qin, Liu & Li, 2017: 186; Feng, Huang & Luo, 2020: 447.
Type species. Gymnaetoides testaceus Qin, Liu & Li, 2017, by original designation.
Diagnosis. Body is smaller than other genera in Aemodogryllini. All of the femora are unarmed on ventral surfaces or only the hind femur with one inner spine on ventral surface. The male epiproct and paraproct are normally developed. The male genitalia has an unpaired dorsal sclerite; the dorsal median lobe of the male genitalia has paired lateral sclerites at the basal area, and the apical area divides into two lobes; the dorsal lateral lobe of the male genitalia is broad, nearly equal in length to dorsal median lobe, and the apical area is sclerotized.
Gymnaetoides testaceus Qin, Liu & Li, 2017
Gymnaetoides testaceus Qin, Liu & Li, 2017: 188.
Material examined. Holotype: ‘3♂1♀’ (Qin et al., 2017a), China, Zhejiang, Kaihua, Gutian Mountain, 14~16.Ⅹ.2012, coll. X.W. Liu and H.Q. Wang, dep. SEMCAS. Paratypes: 1♂1♀, same data as for holotype. Other material: 13♂11♀, China, Zhejiang, Kaihua, Gutian Mountain, 8.Ⅹ.2018, coll. T. Wang, dep. HBU; 3♂17♀, China, Zhejiang, Kaihua, Gutian Mountain, 10.Ⅹ.2018, coll. T. Wang, dep. HBU; 1♂, China, Zhejiang, Linan, Tianmu Mountain, 12.Ⅹ.2018, coll. T. Wang, dep. HBU.
Distribution. China (Zhejiang).
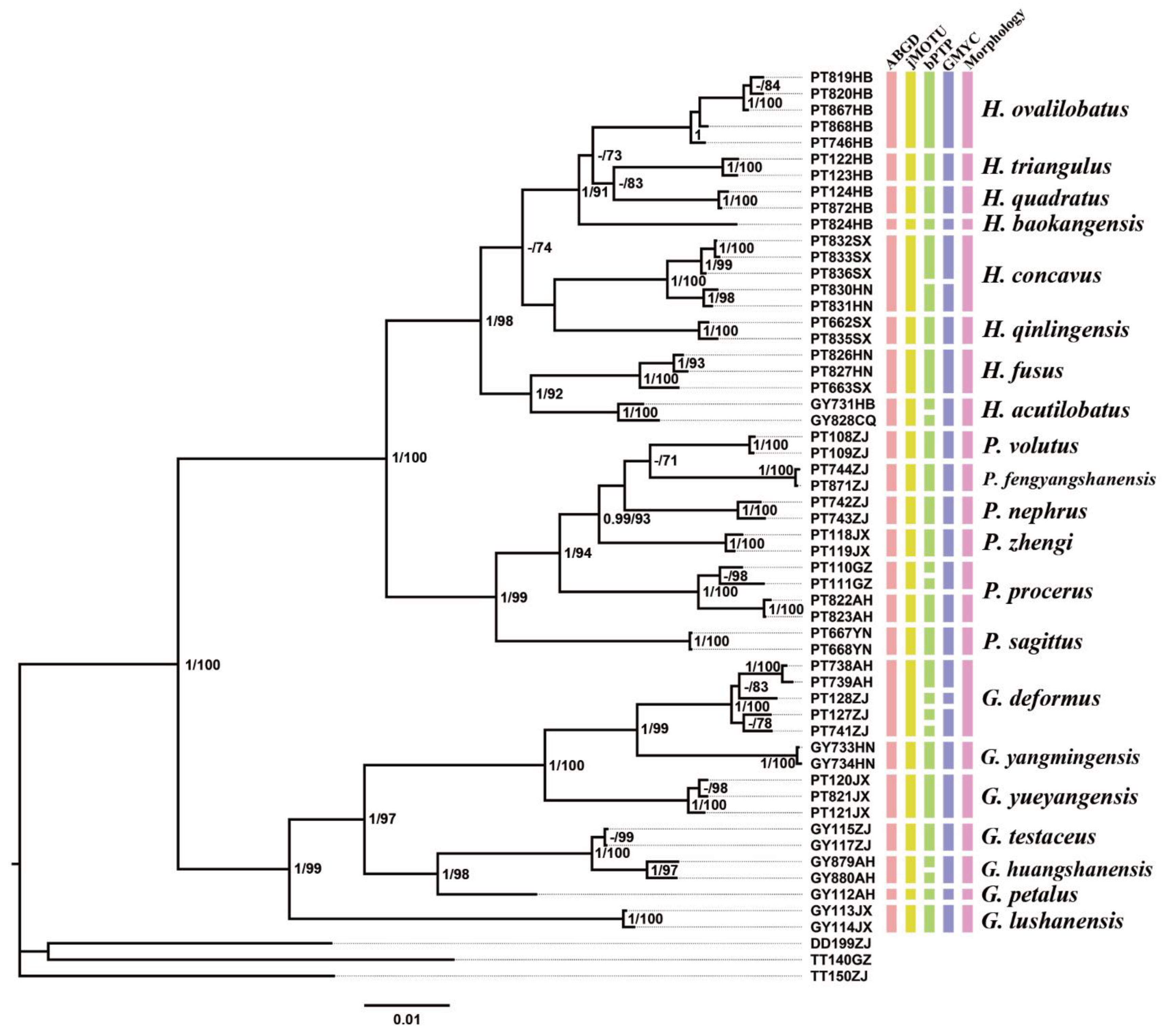
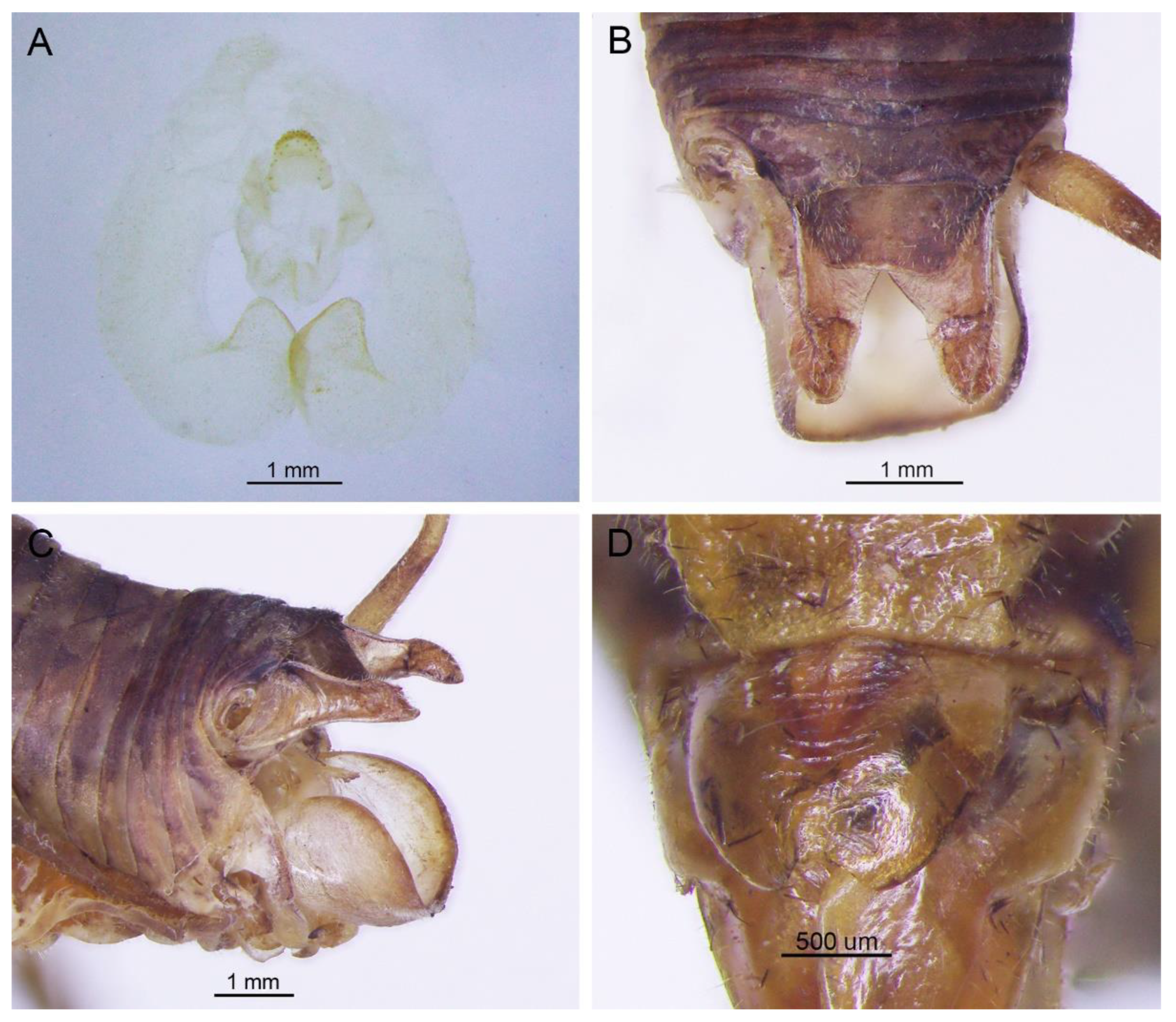
Remarks. The species was recorded in Zhejiang (Gutian Mountain, Tianmu Mountain) and Anhui (Huang Mountain). In the original description, 65–68 inner and 64–69 outer spines on dorsal surface of the hind tibia were reported. However, after examining the type and topotype specimens, 30–39 inner and 37–40 outer spines on dorsal surface of the hind tibia in specimens from Zhejiang were observed, while 60–69 inner spines and 64–69 outer spines were observed in specimens collected from Anhui. This was also indicated by Qin [
16]. Moreover, all of the four species delimitation approaches suggest that the specimens from Zhejiang and Anhui are two separate species (
Figure 1). Therefore, we consider the population from Anhui as a new species,
Gymnaetoides huangshanensis Zhu & Shi sp. nov. (described below).
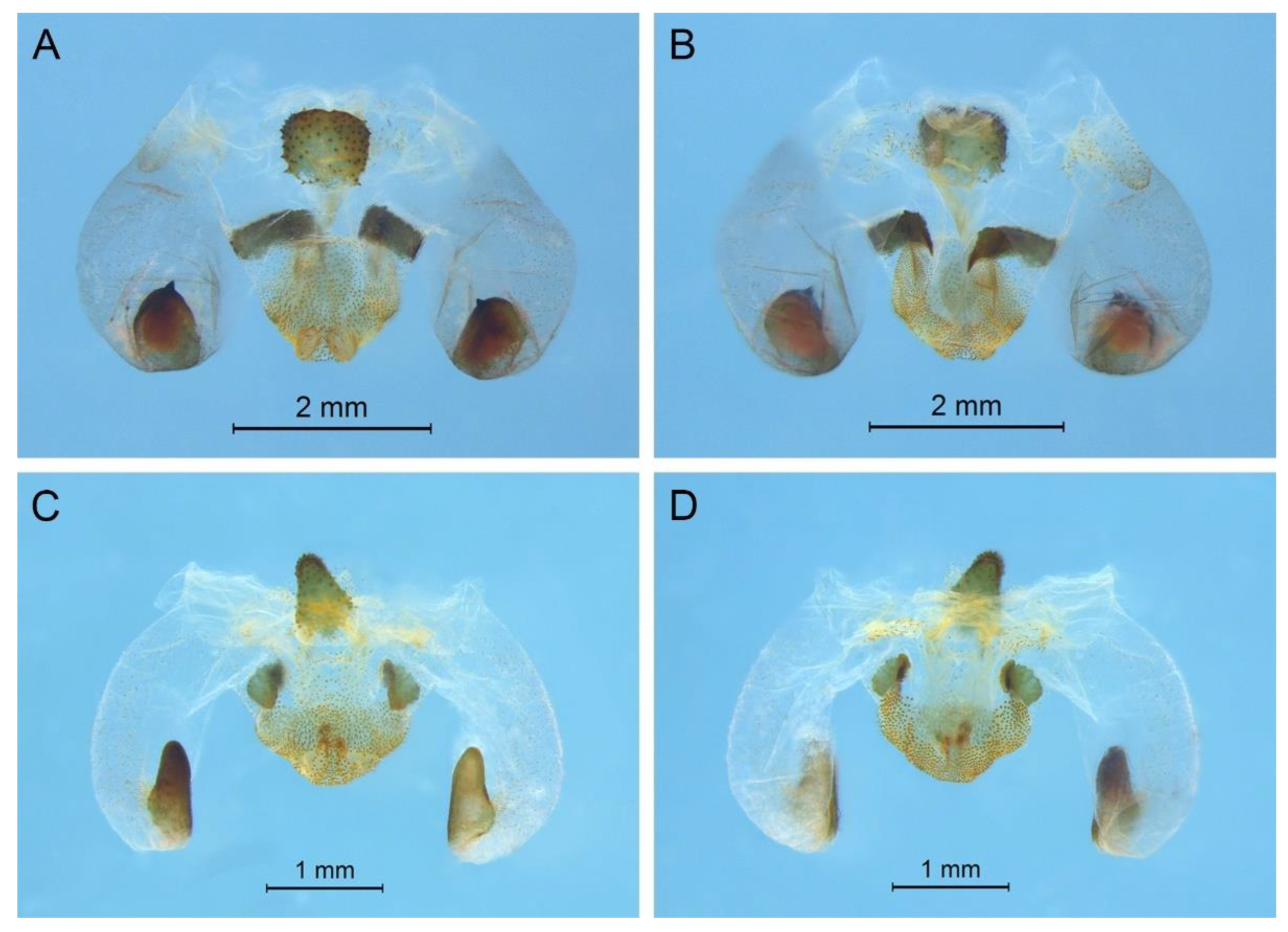
G. testaceus is currently found only in Zhejiang and the description of spines on hind tibia should be changed to “hind tibia with 30–39 inner spines and 37–40 outer spines on dorsal surface”. In addition, the line drawings of the male genitalia did not adequately represent the characters (Figures 3 and 4 in [
13]). The photograph of type specimen is shown in
Figure 2A.
Gymnaetoides huangshanensis Zhu & Shi sp. nov.
urn:lsid:zoobank.org:act:A80F697E-9710-4268-839E-44603F1B7D5B
Material examined. Holotype: ♂, China, Anhui, Huang Mountain, Tangkou, 15.Ⅸ.2019, coll. T. Wang, dep. HBU. Paratypes: 3♂4♀, same data as for holotype. Other material: 3♂, same data as for holotype.
Etymology. The name of the new species derives from the type locality.
Diagnosis. The new species is similar to G. testaceus Qin, Liu & Li, 2017, but it can be distinguished by the shape of the male genitalia and by more dorsal spines on the hind tibia. The number of dorsal spines on the hind tibia of G. testaceus is 30–39 inner spines and 37–40 outer spines, while that of G. huangshanensis is 60–69 inner spines and 64–69 outer spines. The dorsal sclerite of the male genitalia is quadrangular, the basal area broad with concavity, the apical area slightly concave; lateral sclerites of dorsal median lobe are nearly semicircular.
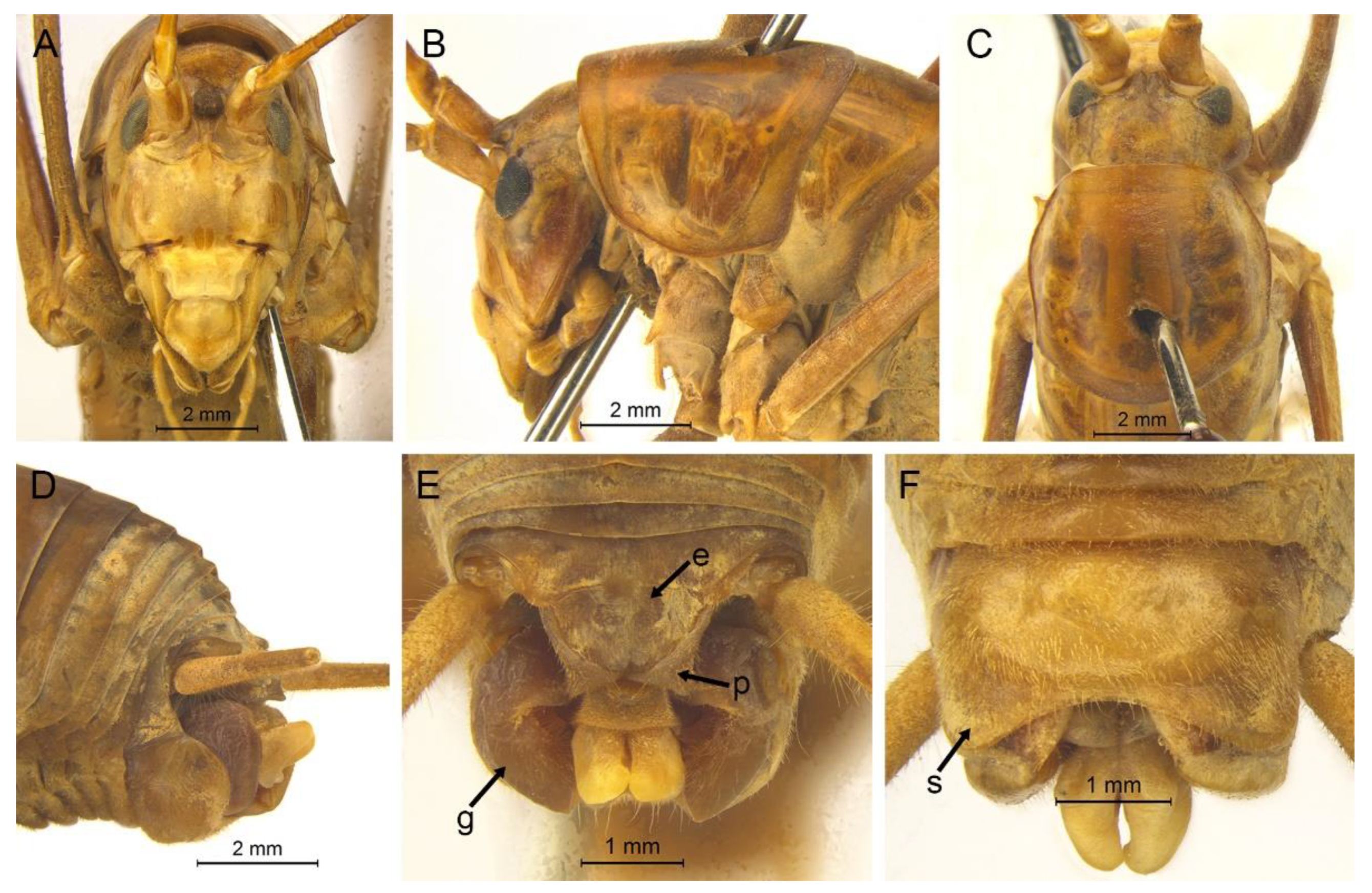
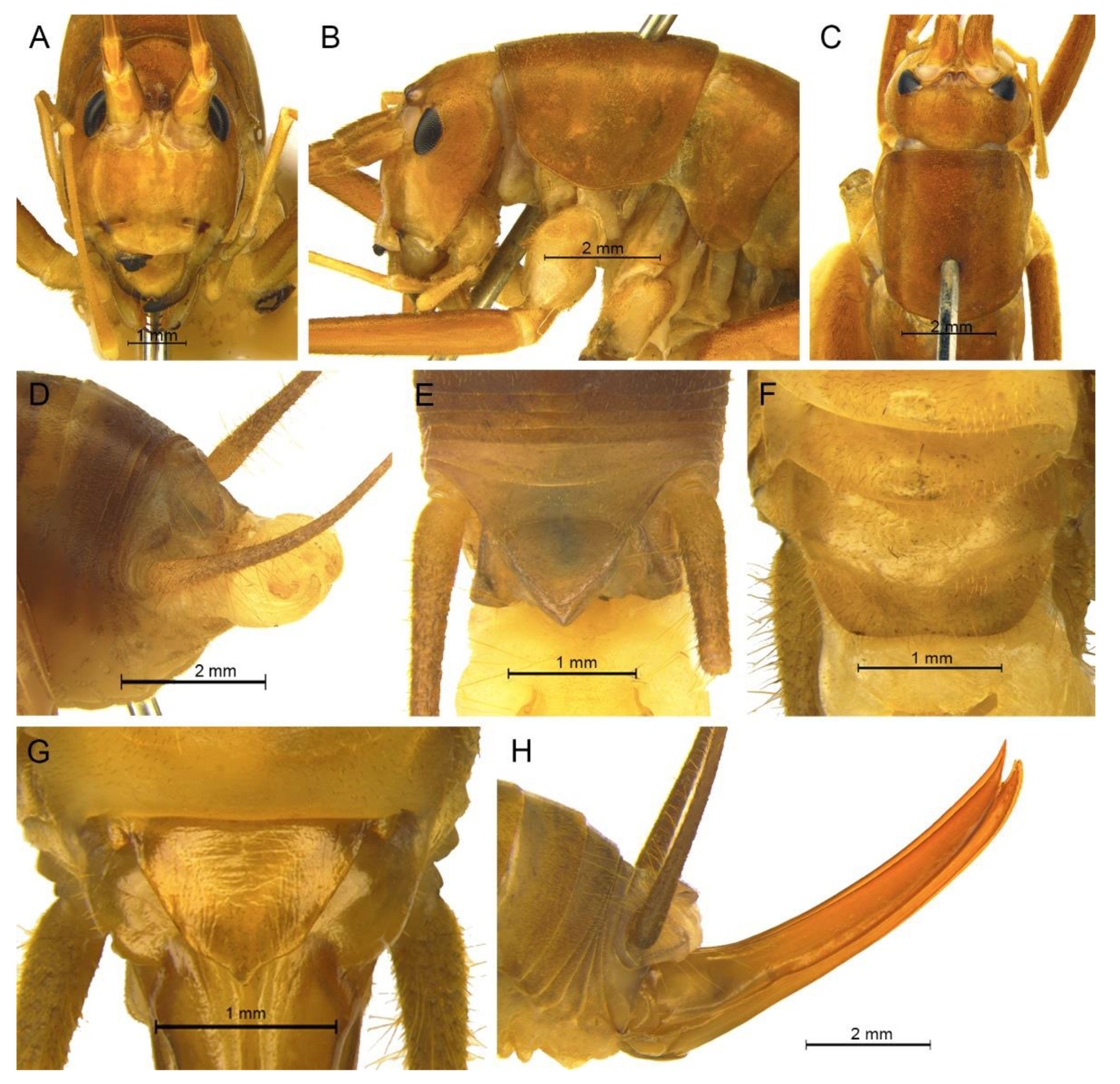
Description. Male: Body medium sized in Gymnaetoides. Fastigium verticis with two conical tubercles, apices rounded, directing forward. Eyes ovoid, protruding forward; median ocellus oval, located between antennal sockets; lateral ocelli circular, situated on lateral margins of basal fastigium verticis. Apical segment of maxillary palpus much longer than subapical segment, apex inflated, globular.
Pronotum long, anterior margin of disc straight, posterior margin arcuate; lateral lobes longer than high, ventral margins arc shaped. Mesonotum and metanotum short, posterior margin of mesonotum arcuate, posterior margin of metanotum straight. Fore coxa with one small spine; femur unarmed on ventral surface, internal genicular lobe with one small spine, external genicular lobe with one long spine; tibia with one inner spine on ventral surface, apical area with one extero-dorsal spine and a pair of ventral spines, between the paired ventral spines with one small spine. Mid femur unarmed on ventral surface, internal and external genicular lobes each with one long spine; tibia with one inner spine and one outer spine on ventral surface, apical area with a pair of dorsal spines and a pair of ventral spines, between the paired ventral spines with one small spine. Hind femur unarmed on ventral surface, internal and external genicular lobes unarmed; tibia with 60–69 inner spines and 64–68 outer spines on dorsal surface, with a pair of dorsal spines in subapical area, and at apex with a pair of dorsal spines and two pairs of ventral spines, intero-dorsal spine shorter than hind basitarsus. Hind basitarsus with one dorsal apical spine.
Posterior margins of all abdominal tergites straight. Epiproct ligulate, paraproct triangular in lateral view. Cercus slender, conical, apex acute. Dorsal sclerite of genitalia quadrangular, basal area broad, with concavity, apical area slightly concave; basal area of dorsal median lobe with paired lateral sclerites, nearly semicircular, apical area divided into two lobes; dorsal lateral lobes broad, nearly equal in length to dorsal median lobe, apical area sclerotized, spinous; ventral lateral lobes tiny, much shorter than dorsal lateral lobes; ventral median lobe short. Subgenital plate quadrangular, transverse and wide, posterior margin slightly concave.
Female: Appearance is similar to male. Ovipositor longer than half the length of hind femur, slightly curved upward, dorsal margin smooth, apical area of ventral margin denticulate. Subgenital plate triangular, apex obtuse.
Coloration. Body yellowish brown, uniform. Eyes black, ocelli yellow.
Measurements (mm). Body: ♂10.9–13.8, ♀9.5–11.6; pronotum: ♂4.1–4.5, ♀3.5–3.8; fore femur: ♂9.3–10.0, ♀8.0–8.4; hind femur: ♂16.5–18.6, ♀13.8–14.6; hind tibia: ♂17.7–19.2, ♀14.8–16.0; hind basitarsus: ♂4.3–4.4, ♀3.3–3.7; ovipositor: 8.6–9.5.
Distribution. China (Anhui).
Gymnaetoides indivisus (Zhang & Liu, 2009)
Diestrammena (Gymnaeta) indivisa Zhang & Liu, 2009: 26.
Gymnaetoides indivisus: Qin, Liu & Li, 2017: 188.
Material examined. Holotype: ♂, China, Jiangxi, Wannian County, 6.Ⅸ.2007, coll. Jiao et al., dep. SEMCAS. Paratype: 1♂, same data as for holotype.
Distribution. China (Jiangxi).
Remarks. The characters of this species were very similar to those of
G. testaceus and thus it was regarded as a synonym of
G. testaceus [
16]. However, the line drawings of
G. indivisus were too abstract and the type specimen was not well preserved. It is difficult to judge whether they are the same species. Therefore, we regard it as a suspect species temporarily, and more evidence is needed for further study.
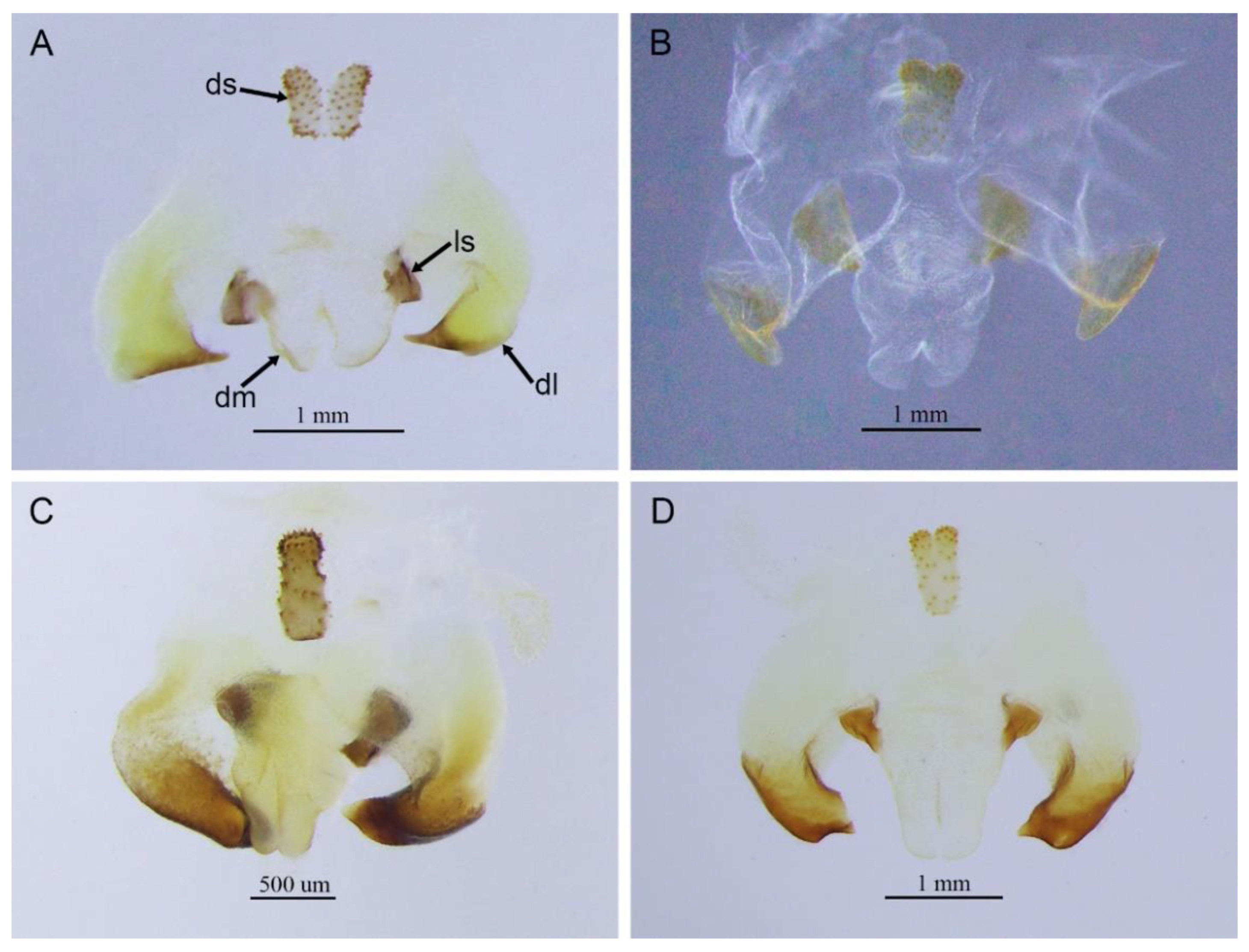
Gymnaetoides deformus (Qin, Liu & Li, 2017) comb. nov.
Pseudotachycines deformis Qin, Liu & Li, 2017: 483.
Material examined. Holotype: ♂, China, Zhejiang, Linan, Tianmu Mountain, 14~15.Ⅹ.2009, coll. X.W. Liu, dep. SEMCAS. Paratypes: 9♂16♀, same data as for holotype. Other material: 1♂2♀, China, Anhui, Huang Mountain, Tangkou, 15.Ⅸ.2019, coll. Y.Q. Li, dep. HBU; 5♂3♀, China, Zhejiang, Linan, Qiangliangfeng, 4.Ⅹ.2019, coll. Q.D. Zhu, dep. HBU; 5♂4♀, China, Zhejiang, Linan, Tianmu Mountain, 30.Ⅸ.2019, coll. Q.D. Zhu, dep. HBU; 4♂8♀, China, Zhejiang, Linan, Tianmu Mountain, 11.Ⅹ.2018, coll. T. Wang, dep. HBU; 2♂13♀, Tianmu Mountain, Linan, Zhejiang, China, 12.Ⅹ.2018, coll. T. Wang, dep. HBU.
Distribution. China (Anhui, Zhejiang).
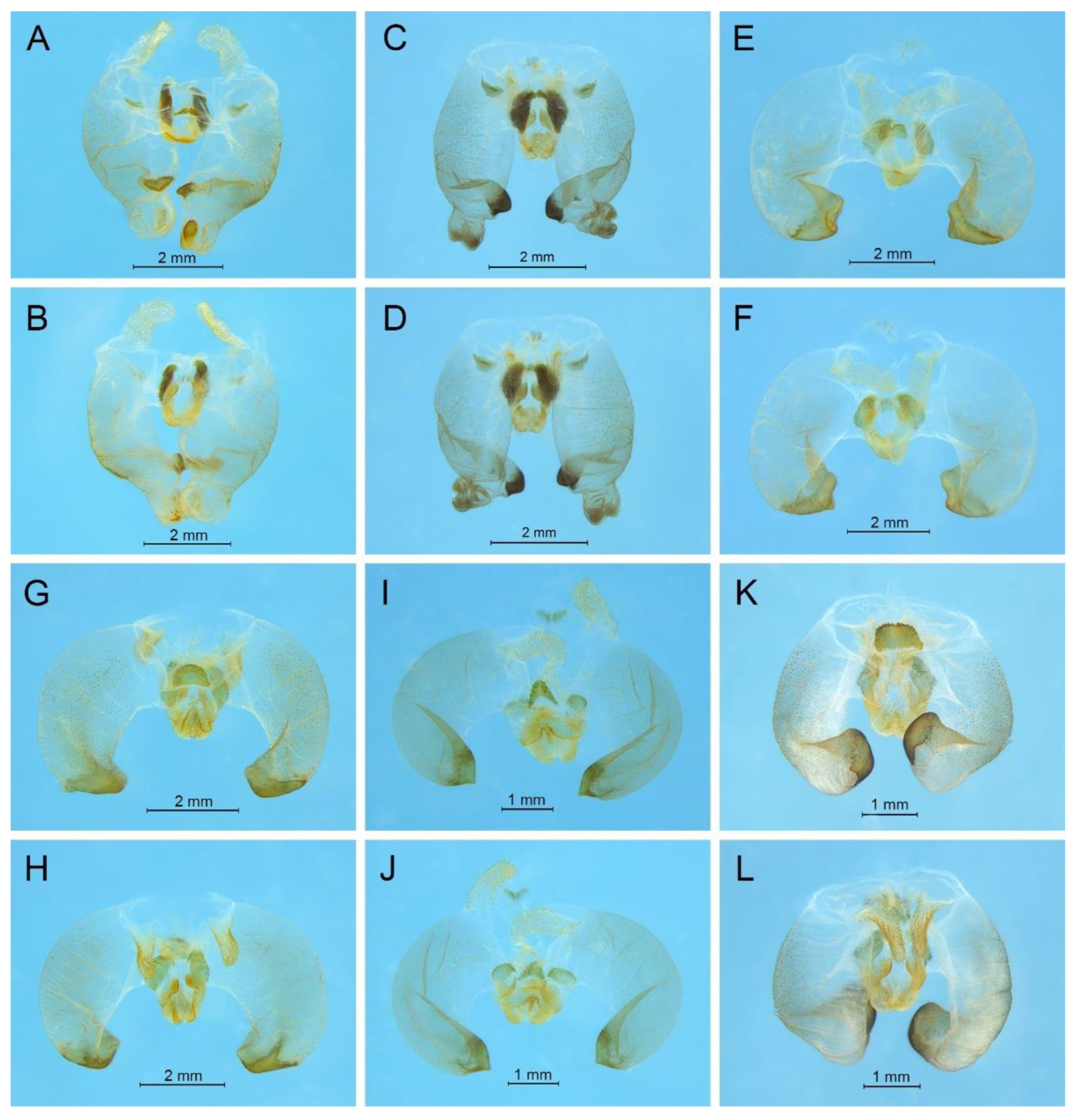
Remarks. This species was designated as the type species of
Pseudotachycines, which was characterized by the prolonged male paraproct [
14]. However, after examining the type specimen, we did not find the specialized paraproct. Moreover, the line drawings of the male genitalia did not adequately represent the characters (Figures 4 and 5 in [
14]). The photograph of the type specimen is shown in
Figure 2B. The result of phylogenetic analyses also clusters this species in genus
Gymnaetoides (
Figure 1). It should be transferred into the genus
Gymnaetoides according to the characters of the male genitalia: the apical area of the dorsal median lobe divided into two lobes, the dorsal lateral lobes broad, nearly equal in length to dorsal median lobe, the apical area sclerotized.
Gymnaetoides inermus (Qin, Liu & Li, 2017) comb. nov.
Pseudotachycines inermis Qin, Liu & Li, 2017: 483.
Material examined. Holotype: ♂, China, Hubei, Xianning, Jiugong Mountain, 30.Ⅶ.2016, coll. Hu and Liu, dep. SEMCAS.
Distribution. China (Hubei).
Remarks. The species was classified into
Pseudotachycines based on the prolonged male paraproct [
14]. However, after examining the type specimen, this feature was not distinct. The line drawings of the male genitalia did not adequately represent the characters (Figures 8 and 9 in [
14]). The photograph of the type specimen is shown in
Figure 2C. The dorsal sclerite of the male genitalia is rectangular, much longer than wide; the lateral sclerite of the dorsal median lobe is nearly rectangular, with the apical area divided into two lobes; the dorsal lateral lobe is broad, nearly equal in length to the dorsal median lobe, with the apical area sclerotized; the ventral lateral lobe and the ventral median lobe are short. According to the characters of the male genitalia, it should be transferred into the genus
Gymnaetoides.
Gymnaetoides yueyangensis (Qin, Liu & Li, 2017) comb. nov.
Pseudotachycines yueyangensis Qin, Liu & Li, 2017: 485.
Material examined. Holotype: ♂, China, Hunan, Yueyang, Forest Park of Fushou Mountain, 18~26.Ⅹ.2016, coll. Jiang et al., dep. SEMCAS. Paratypes: 6♂6♀, same data as for holotype. Other material: 6♂3♀, China, Jiangxi, Lu Mountain, Guling, 7.Ⅷ.2018, coll. T. Wang, dep. HBU.
Distribution. China (Hunan, Jiangxi).
Remarks. After examining the type specimen, we found that the line drawings of the male genitalia did not adequately represent the characters (Figures 10 and 11 in [
14]). The photograph of the type specimen is shown in
Figure 2D. The apical area of the dorsal median lobe of the male genitalia divides into two lobes; the dorsal lateral lobe is broad, nearly equal in length to the dorsal median lobe, with the apical area sclerotized. According to the characters of the male genitalia and the phylogenetic analyses, we transfer it into genus
Gymnaetoides.
Gymnaetoides petalus Zhu & Shi sp. nov.
urn:lsid:zoobank.org:act:6395D039-02AF-464D-A89A-5B33A3E7DBD0
Material examined. Holotype: ♂, China, Anhui, Chizhou City, Shitai County, Gongxi Village, Du hamlet, Penglaixian Cave, 17.Ⅶ.2014, coll. M.Y. Tian, dep. HBU. Paratype: 1♂, same data as for holotype.
Etymology. The name of the new species derives from the Greek word ‘petal’ (petaloid), referring to the petaloid dorsal sclerite of the male genitalia.
Diagnosis. The new species can easily be distinguished from known congeneric species by the shape of the male genitalia: the dorsal sclerite of the male genitalia is petaloid and the lateral sclerite of the dorsal median lobe is slender.
Description. Male: Body medium sized in Gymnaetoides. Fastigium verticis with two conical tubercles, apices divided, directing forward. Eyes ovoid, protruding forward; median ocellus oval, located between antennal sockets; lateral ocelli circular, situated on lateral margins of basal fastigium verticis. Apical segment of maxillary palpus little longer than subapical segment, apex truncate.
Anterior margin of pronotum straight, posterior margin arcuate; lateral lobes longer than high, ventral margins arc shaped. Mesonotum and metanotum short, posterior margin of mesonotum arcuate, posterior margin of metanotum straight. Fore coxa with one small spine; femur unarmed on ventral surface, internal genicular lobe with one small spine, external genicular lobe with one long spine; tibia with two outer spines on ventral surface, apical area with one extero-dorsal spine and a pair of ventral spines, between the paired ventral spines with one small spine. Mid femur unarmed on ventral surface, internal and external genicular lobes each with one long spine; tibia with one inner spine and one outer spine on ventral surface, apical area with a pair of dorsal spines and a pair of ventral spines, between the paired ventral spines with one small spine. Hind femur unarmed on ventral surface; tibia with 87–92 inner spines and 84–86 outer spines on dorsal surface, with a pair of dorsal spines in subapical area, and at apex with a pair of dorsal spines and two pairs of ventral spines, intero-dorsal spine shorter than hind basitarsus. Hind basitarsus with one dorsal apical spine.
All abdominal tergites without process. Epiproct ligulate, paraproct slightly longer than epiproct. Cercus slender, conical, apex acute. Dorsal sclerite of genitalia petaloid; basal area of dorsal median lobe with paired lateral sclerites, slender, apical area divided into two lobes; dorsal lateral lobes broad, nearly equal in length to dorsal median lobe, apical area sclerotized, spinous; ventral lateral lobes cylindrical, much shorter than dorsal lateral lobes; ventral median lobe short. Subgenital plate transverse and wide, posterior margin widely concave.
Female: Unknown.
Coloration. Body yellowish brown, uniform.
Measurements (mm). Body: ♂12.6–13.9; pronotum: ♂6.6–6.8; fore femur: ♂9.7–11.8; hind femur: ♂18.2–18.4; hind tibia: ♂19.9–20.4; hind basitarsus: ♂4.4–4.9.
Distribution. China (Anhui).
Gymnaetoides yangmingensis Zhu & Shi sp. nov.
urn:lsid:zoobank.org:act:AEFABE7E-E962-47A7-B2EB-37CF8E3D6EE4
Material examined. Holotype: ♂, China, Hunan, Shuangpai, Yangming Mountain, 21.Ⅷ.2019, coll. Y.R. Xin, dep. HBU. Paratypes: 2♂4♀, same data as for holotype. Other material: 1♂8♀, same data as for holotype.
Etymology. The name of the new species refers to the type locality.
Diagnosis. The new species is similar to Gymnaetoides yueyangensis (Qin, Liu & Li, 2017), but it differs from the latter one by the shape of the male genitalia. The dorsal sclerite of the male genitalia is quadrangular, with a concavity on the basal area; the lateral sclerite of the dorsal median lobe is semicircular; the apical area of the dorsal lateral lobe is spinous.
Description. Male: Body medium sized in Gymnaetoides. Fastigium verticis with two conical tubercles, apices divided, directing forward. Eyes ovoid, protruding forward; median ocellus oval, located between antennal sockets; lateral ocelli circular, situated on lateral margins of basal fastigium verticis. Apical segment of maxillary palpus little longer than subapical segment, apex inflated, globular.
Pronotum long, anterior margin of disc straight, posterior margin arcuate; lateral lobes longer than high, ventral margins arc shaped. Mesonotum and metanotum short, posterior margin of mesonotum arcuate, posterior margin of metanotum straight. Fore coxa with one small spine; femur unarmed on ventral surface, internal genicular lobe with one small spine, external genicular lobe with one long spine; tibia with two outer spines on ventral surface, apical area with one extero-dorsal spine and a pair of ventral spines, between the paired ventral spines with one small spine. Mid femur unarmed on ventral surface, internal and external genicular lobes each with one long spine; tibia with one inner spine and one outer spine on ventral surface, apical area with a pair of dorsal spines and a pair of ventral spines, between the paired ventral spines with one small spine. Hind femur unarmed or with one spine on ventral surface, internal genicular lobe with one small spine; tibia with 50–56 inner spines and 51–53 outer spines on dorsal surface, with a pair of dorsal spines in subapical area, and at apex with a pair of dorsal spines and two pairs of ventral spines, intero-dorsal spine nearly equal in length to hind basitarsus. Hind basitarsus with one dorsal apical spine.
All abdominal tergites without process. Epiproct ligulate, paraproct triangular in lateral view. Cercus slender, conical, apex acute. Dorsal sclerite of genitalia quadrangular, basal area wide, with concavity; basal area of dorsal median lobe with paired lateral sclerites, semicircular, apical area divided into two lobes; dorsal lateral lobes broad, nearly equal in length to dorsal median lobe, apical area spinous; ventral lateral lobes much shorter than dorsal lateral lobes; ventral median lobe short. Subgenital plate transverse and wide, posterior margin straight.
Female: Appearance is similar to male. Ovipositor longer than half the length of hind femur, slightly curved upward, dorsal margin smooth, apical area of ventral margin denticulate. Subgenital plate trapezoidal, posterior margin slightly protruding.
Coloration. Body light brown, uniform.
Measurements (mm). Body: ♂10.3–11.7, ♀11.9–12.7; pronotum: ♂4.2–4.5, ♀3.8–4.6; fore femur: ♂6.3–7.4, ♀7.1–7.6; hind femur: ♂14.1–14.7, ♀14.7–15.4; hind tibia: ♂14.3–15.2, ♀15.4–16.4; hind basitarsus: ♂3.3–4.1, ♀3.3–3.6; ovipositor: 8.1–8.3.
Distribution. China (Hunan).
Gymnaetoides lushanensis Zhu & Shi sp. nov.
urn:lsid:zoobank.org:act:0DC9C41D-1105-4D8A-882D-FBFA70F04889
Material examined. Holotype: ♂, China, Jiangxi, Lu Mountain, Guling, 7.Ⅷ.2018, coll. T. Wang, dep. HBU. Paratypes: 1♀, same data as for holotype; 1♂1♀, China, Jiangxi, Lu Mountain, Guling, 6.Ⅷ.2018, coll. T. Wang, dep. HBU. Other material: 2♂, China, Jiangxi, Lu Mountain, Dayueshan, 3.Ⅷ.2018, coll. T. Wang, dep. HBU; 2♂2♀, China, Jiangxi, Lu Mountain, Guling, 6.Ⅷ.2018, coll. T. Wang, dep. HBU; 5♂2♀, China, Jiangxi, Lu Mountain, Guling, 7.Ⅷ.2018, coll. T. Wang, dep. HBU.
Etymology. The name of the new species derives from the type locality.
Diagnosis. The new species is similar to Gymnaetoides huangshanensis Zhu & Shi sp. nov., but it can be distinguished by the shapes of lateral sclerite of the dorsal median lobe and the dorsal lateral lobe of the male genitalia. The lateral sclerite of the dorsal median lobe is spoon shaped and the apical area of the dorsal lateral lobe is slightly sclerotized.
Description. Male: Body medium sized in Gymnaetoides. Fastigium verticis with two conical tubercles, apices divided, directing forward. Eyes ovoid, protruding forward; median ocellus oval, located between antennal sockets; lateral ocelli circular, situated on lateral margins of basal fastigium verticis. Apical segment of maxillary palpus much longer than subapical segment, apex inflated, globular.
Pronotum long, anterior margin of disc straight, posterior margin arcuate; lateral lobes longer than high, ventral margins arc shaped. Mesonotum and metanotum short, posterior margin of mesonotum arcuate, posterior margin of metanotum straight. Fore coxa with one small spine; femur unarmed on ventral surface, internal genicular lobe with one small spine, external genicular lobe with one long spine; tibia with two outer spines on ventral surface, apical area with one extero-dorsal spine and a pair of ventral spines, between the paired ventral spines with one small spine. Mid femur unarmed on ventral surface, internal and external genicular lobes each with one long spine; tibia with one inner spine and two outer spines on ventral surface, apical area with a pair of dorsal spines and a pair of ventral spines, between the paired ventral spines with one small spine. Hind femur unarmed on ventral surface; tibia with 65–75 inner spines and 63–71 outer spines on dorsal surface, with a pair of dorsal spines in subapical area, and at apex with a pair of dorsal spines and two pairs of ventral spines, intero-dorsal spine slightly shorter than hind basitarsus. Hind basitarsus with one dorsal apical spine.
Posterior margins of all abdominal tergites straight. Epiproct ligulate, paraproct simple, triangular in lateral view. Cercus slender, conical, apex acute. Dorsal sclerite of genitalia quadrangular, basal area wide, with a concavity, apex slightly concave; basal area of dorsal median lobe with paired lateral sclerites, spoon-shaped, apical area divided into two lobes; dorsal lateral lobes broad, nearly equal in length to dorsal median lobe, apical area slightly sclerotized; ventral lateral lobes cylindrical, nearly equal in length to dorsal lateral lobes; ventral median lobe short. Subgenital plate transverse and wide, posterior margin straight.
Female: Appearance is similar to male. Ovipositor longer than half the length of hind femur, dorsal margin smooth, apical area of ventral margin denticulate. Subgenital plate triangular, apex acute.
Coloration. Body yellow. Pronotum and apex of abdomen yellowish brown.
Measurements (mm). Body: ♂10.0–11.5, ♀11.0–12.5; pronotum: ♂4.2–5.5, ♀4.5–5.4; fore femur: ♂9.0–9.5, ♀7.0–8.0; hind femur: ♂15.2–16.0, ♀14.5–15.8; hind tibia: ♂15.2–16.3, ♀15.5–17.0; hind basitarsus: ♂4.0–4.4, ♀3.5–4.2; ovipositor: 8.5–9.0.
Distribution. China (Jiangxi).
Genus Pseudotachycines Qin, Liu & Li, 2017
Pseudotachycines Qin, Liu & Li, 2017: 482.
Type species. Pseudotachycines volutus Qin, Liu & Li, 2017, here redesignation.
Diagnosis. Body is medium sized in Aemodogryllini. The male paraproct are specialized, slightly or much longer than epiproct. The male genitalia have unpaired dorsal sclerite, smaller than that of the other genera in Aemodogryllini; the dorsal median lobe of the male genitalia has paired lateral sclerites at the basal area, the apical area does not divide; the dorsal lateral lobes of the male genitalia are broad and folded, much longer than dorsal median lobe, with the apical area slightly sclerotized, almost as wide as the basal area.
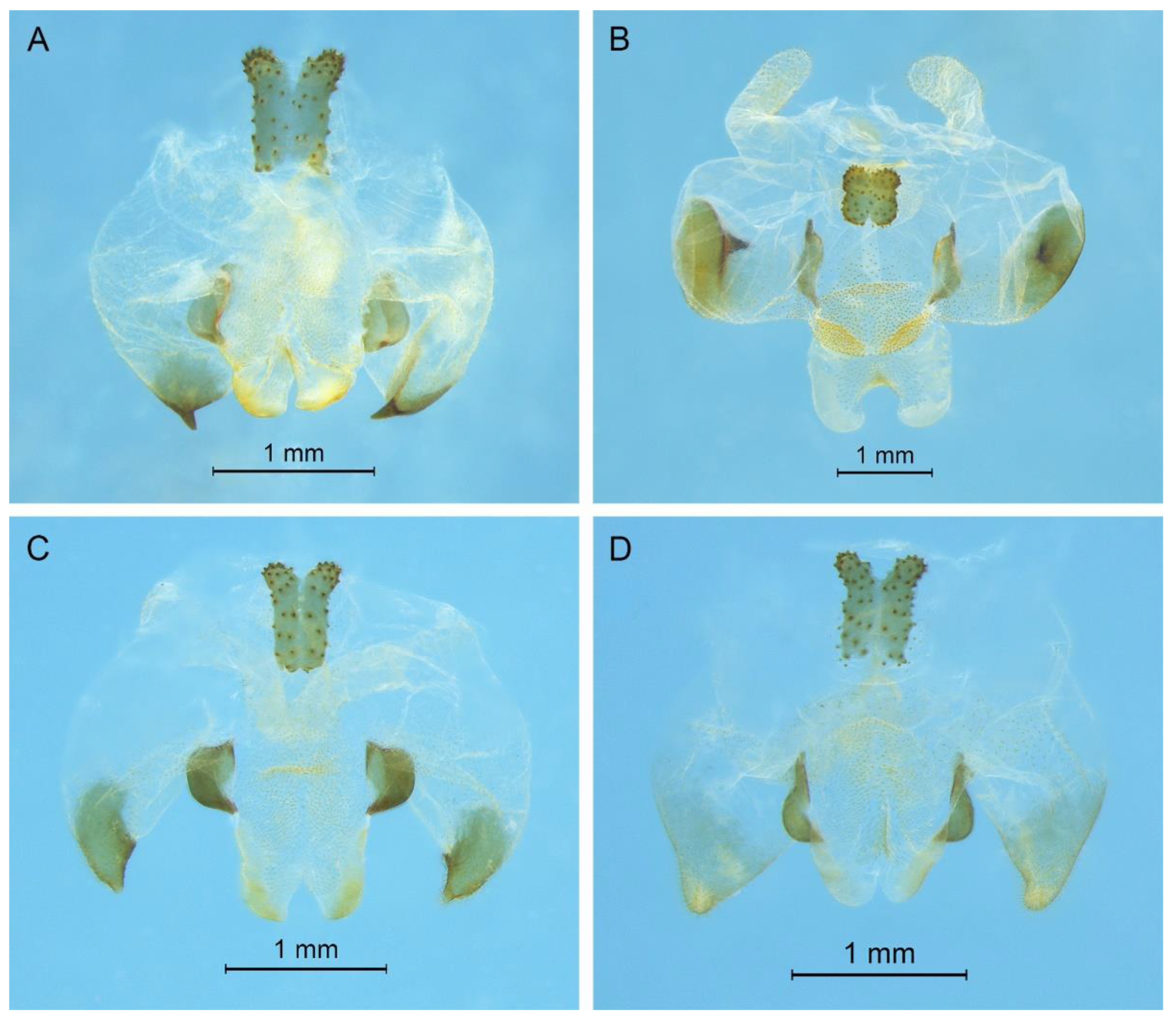
Pseudotachycines volutus Qin, Liu & Li, 2017
Pseudotachycines volutus Qin, Liu & Li, 2017: 486.
Material examined. Holotype: ♂, China, Hunan, Yueyang, Forest Park of Fushoushan, 18~26.Ⅸ.2016, coll. Jiang et al., dep. SEMCAS. Paratypes: 1♂3♀, same data as for holotype; 2♂, China, Zhejiang, Tianmu Mountain, 31.Ⅷ.2012, coll. X.W. Liu et al., dep. SEMCAS. Other material: 1♂, China, Zhejiang, Linan, Tianmu Mountain, 15.Ⅷ.2018, coll. Y.J. Dou and Y.X. Zhen, dep. HBU; 1♀, China, Zhejiang, Linan, Tianmu Mountain, 12.Ⅷ.2018, coll. Y.J. Dou and Y.X. Zhen, dep. HBU.
Distribution. China (Hunan, Zhejiang).
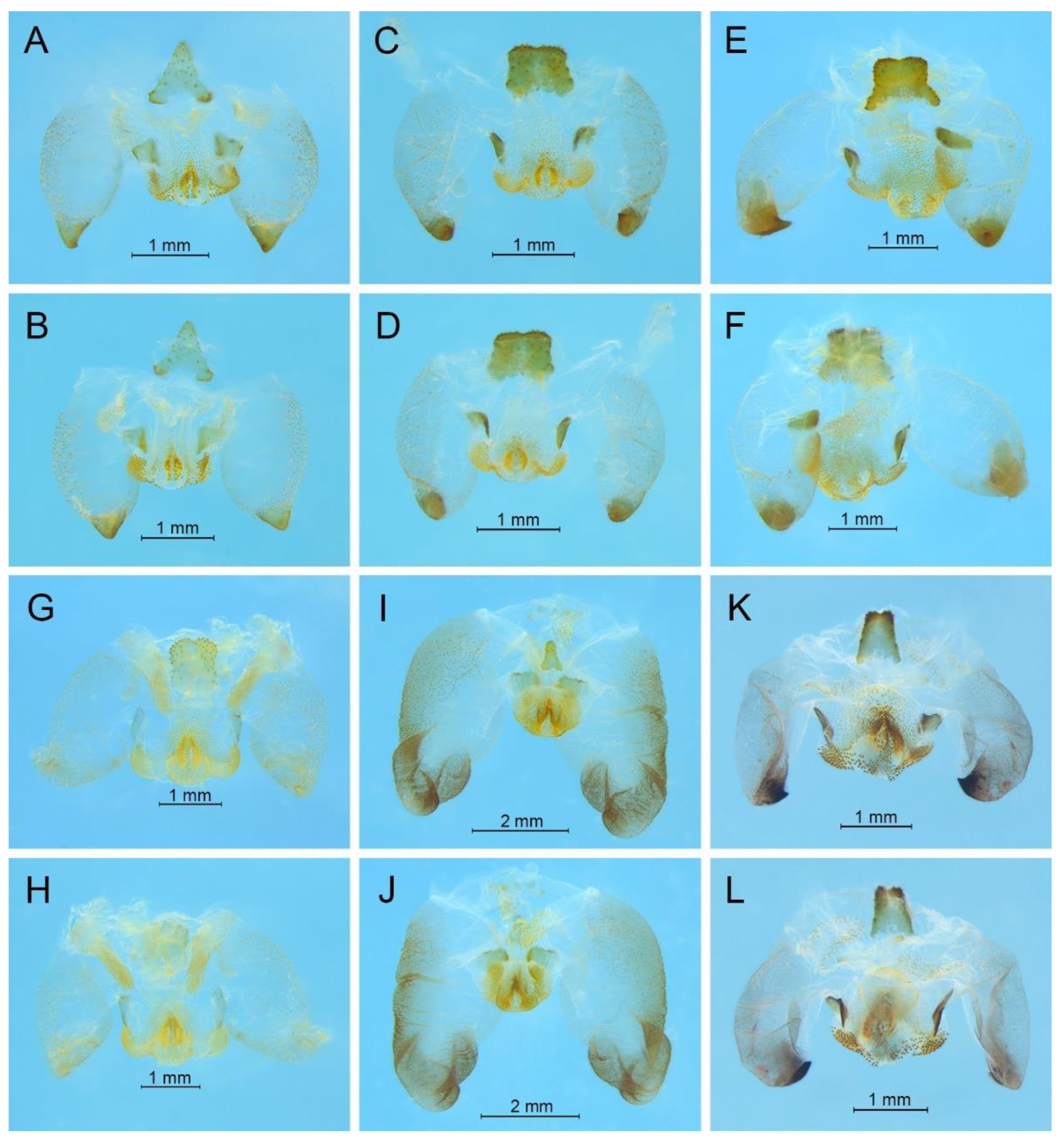
Remarks. Because the type species of genus
Pseudotachycines is moved to genus
Gymnaetoides,
Pseudotachycines volutus is redesignated as the type species of the genus
Pseudotachycines. After examining the type specimen, we found that the line drawings of the male genitalia did not adequately represent the characters (Figures 14–16 in [
14]). The photographs of the type specimens are shown in
Figure 8.
Pseudotachycines procerus Zhu & Shi sp. nov.
urn:lsid:zoobank.org:act:9BC06DD7-1F67-4F05-89F1-366023B49965
Material examined. Holotype: ♂, China, Anhui, Yuexi, Yaoluoping, 18.Ⅸ.2019, coll. Y.Q. Li, dep. HBU. Paratypes: 3♂1♀, same data as for holotype.
Etymology. The name of the new species derives from the Latin word ‘procer’ (‘process’), referring to the subapical area of the dorsal lateral lobe of the male genitalia with a process on the dorsal surface.
Diagnosis. The new species can easily be distinguished from known congeneric species by the shape of the male genitalia and paraproct. The dorsal sclerite of the male genitalia is lunular and the lateral sclerite of the dorsal median lobe is oblong. The subapical area of the dorsal lateral lobe of the male genitalia has a process on the dorsal surface. The male paraproct is oblong in the lateral view.
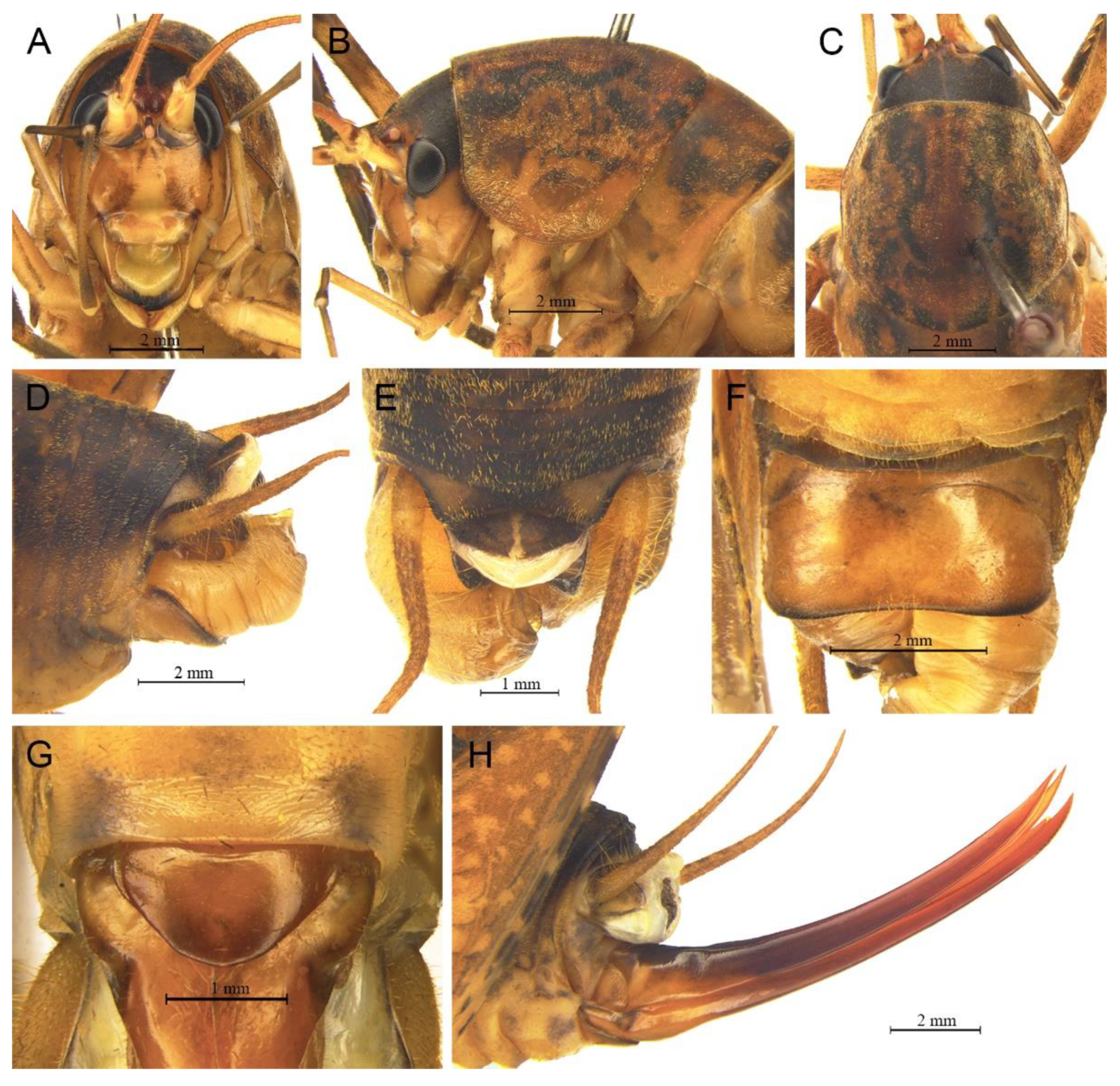
Description. Male: Body medium sized in Pseudotachycines. Fastigium verticis with two conical tubercles, apices rounded, directing forward. Eyes ovoid, protruding forward; median ocellus oval, located between antennal sockets; lateral ocelli circular, situated on lateral margins of basal fastigium verticis. Apical segment of maxillary palpus much longer than subapical segment, apex inflated, globular.
Anterior margin of pronotum straight, posterior margin arcuate; lateral lobes longer than high, ventral margins arc shaped. Mesonotum and metanotum short, posterior margin of mesonotum arcuate, posterior margin of metanotum straight. Fore coxa with one small spine; femur unarmed on ventral surface, internal genicular lobe with one small spine, external genicular lobe with one long spine; tibia with two to three inner spines and two outer spines on ventral surface, apical area with one extero-dorsal spine and a pair of ventral spines, between the paired ventral spines with one small spine. Mid femur unarmed on ventral surface, internal and external genicular lobes each with one long spine; tibia with one inner spine and two outer spines on ventral surface, apical area with a pair of dorsal spines and a pair of ventral spines, between the paired ventral spines with one small spine. Hind femur with three to five inner spines on ventral surface; tibia with 57–60 inner spines and 55–56 outer spines on dorsal surface, with a pair of dorsal spines in subapical area, and at apex with a pair of dorsal spines and two pairs of ventral spines, intero-dorsal spine much longer than hind basitarsus. Hind basitarsus with one dorsal apical spine.
All abdominal tergites without process. Epiproct ligulate, paraproct extended backward, slightly longer than epiproct, oblong in lateral view. Cercus slender, conical, apex acute. Dorsal sclerite of genitalia small and lunular; basal area of dorsal median lobe with paired lateral sclerites, oblong, apical area not divided; dorsal lateral lobes broad and folded, much longer than dorsal median lobe, subapical area with a process on dorsal surface, apical area slightly sclerotized, almost as wide as basal area; ventral lateral lobes and ventral median lobe short. Subgenital plate quadrangular, basal half broad, apical half narrow, posterior margin slightly concave.
Female: Appearance is similar to male. Ovipositor longer than half the length of hind femur, curved upward, dorsal margin smooth, apical area of ventral margin denticulate. Subgenital plate quadrangular, basal area broad, gradually narrowing to apex, posterior margin blunt.
Coloration. Dorsal surface of body light brown, ventral surface yellowish brown. Face with four longitudinal black stripes.
Measurements (mm). Body: ♂15.3–17.2, ♀18.5; pronotum: ♂4.8–5.2, ♀6.4; fore femur: ♂6.8–7.0, ♀7.3; hind femur: ♂15.5–16.0, ♀17.7; hind tibia: ♂14.7–15.0, ♀16.8; hind basitarsus: ♂2.8–3.1, ♀2.8; ovipositor: 12.6.
Distribution. China (Anhui).
Pseudotachycines procerus guizhouensis Zhu & Shi ssp. nov.
Material examined. Holotype: ♂, China, Guizhou, Suiyang, Kuankuoshui, 13.Ⅷ.2017, coll. Q.D. Zhu and Y.Q. Li, dep. HBU. Paratype: 1♂, China, Guizhou, Daozhen, Dashahe, 7.Ⅷ.2017, coll. Q.D. Zhu and Y.Q. Li, dep. HBU.
Etymology. The name of the new species derives from the type locality.
Diagnosis. The species is very similar to Pseudotachycines procerus Zhu & Shi sp. nov., the slight differences are as follows: the lateral sclerite of the dorsal median lobe of the male genitalia is semicircular, and the hind femur has one to three inner spines on the ventral surface, which is less than three to five inner spines of the latter one.
Description. Male: Body medium sized in Pseudotachycines. Fastigium verticis with two conical tubercles, apices divided, directing forward. Eyes ovoid, protruding forward; median ocellus oval, located between antennal sockets; lateral ocelli circular, situated on lateral margins of basal fastigium verticis. Apical segment of maxillary palpus much longer than subapical segment, apex inflated, globular.
Pronotum long, anterior margin of disc straight, posterior margin arcuate; lateral lobes longer than high, ventral margins arc shaped. Mesonotum and metanotum short, posterior margin of mesonotum arcuate, posterior margin of metanotum straight. Fore coxa with one small spine; femur unarmed on ventral surface, internal genicular lobe with one small spine, external genicular lobe with one long spine; tibia with two inner spines and two outer spines on ventral surface, apical area with one extero-dorsal spine and a pair of ventral spines, between the paired ventral spines with one small spine. Mid femur unarmed on ventral surface, internal and external genicular lobes each with one long spine; tibia with one inner spine and two outer spines on ventral surface, apical area with a pair of dorsal spines and a pair of ventral spines, between the paired ventral spines with one small spine. Hind femur with one to three inner spines on ventral surface; tibia with 53–56 inner spines and 50–54 outer spines on dorsal surface, with a pair of dorsal spines in subapical area, and at apex with a pair of dorsal spines and two pairs of ventral spines, intero-dorsal spine much longer than hind basitarsus. Hind basitarsus with one dorsal apical spine.
All abdominal tergites without process. Epiproct ligulate, paraproct extended backward, slightly longer than epiproct, oblong in lateral view. Cercus slender, conical, apex acute. Dorsal sclerite of genitalia small and lunular, basal area slightly concave; basal area of dorsal median lobe with paired lateral sclerites, semicircular, apical area not divided; dorsal lateral lobes broad and folded, much longer than dorsal median lobe, subapical area with a process on dorsal surface, apical area sclerotized, almost as wide as basal area; ventral lateral lobes and ventral median lobe short. Subgenital plate quadrangular, basal half broad, apical half narrow, posterior margin slightly concave.
Female: Unknown.
Coloration. Dorsal surface of body light brown, ventral surface yellowish brown. Face with four longitudinal black stripes.
Measurements (mm). Body: ♂16.3–16.7; pronotum: ♂5.0–5.6; fore femur: ♂6.9–7.2; hind femur: ♂15.1–15.9; hind tibia: ♂15.1–15.5; hind basitarsus: ♂3.0–3.1.
Distribution. China (Guizhou).
Remarks. The species is very similar to Pseudotachycines procerus Zhu & Shi sp. nov. and there are only slight differences between them. However, all of the four species delimitation methods divide them into two MOTUs. Therefore, we regard it as a subspecies of Pseudotachycines procerus Zhu & Shi sp. nov.
Pseudotachycines zhengi Zhu & Shi sp. nov.
urn:lsid:zoobank.org:act:E355BD1A-AE33-42BF-BE49-8BFFC44654CF
Material examined. Holotype: ♂, China, Jiangxi, Lushan, Guling, 7.Ⅷ.2018, coll. T. Wang, dep. HBU. Paratypes: 3♂2♀, same data as for holotype. Other material: 5♂2♀, same data as for holotype; 1♂, China, Jiangxi, Lushan, Dayueshan, 3.Ⅷ.2018, coll. T. Wang, dep. HBU; 2♂, China, Zhejiang, Longquan, Fengyangshan, 18.Ⅸ.2019, coll. Y.X. Zhen, dep. HBU.
Etymology. The new species is named after Mr. Zhemin Zheng to commemorate his great contribution to the study of Chinese Orthoptera.
Diagnosis. The new species can easily be distinguished from known congeneric species by the shape of the male genitalia and paraproct. The dorsal sclerite of the male genitalia is nearly semicircular, with a concavity at the basal area and the apical area broadly concave. The lateral sclerite of the dorsal median lobe of the male genitalia is oblong. The male paraproct is semicircular in the lateral view.
Description. Male: Body medium sized in Pseudotachycines. Fastigium verticis with two conical tubercles, apices rounded, directing forward. Eyes ovoid, protruding forward; median ocellus oval, located between antennal sockets; lateral ocelli circular, situated on lateral margins of basal fastigium verticis. Apical segment of maxillary palpus much longer than subapical segment, apex inflated, globular.
Pronotum long, anterior margin straight, posterior margin arcuate; lateral lobes longer than high, ventral margins arc shaped. Mesonotum and metanotum short, posterior margin of mesonotum arcuate, posterior margin of metanotum straight. Fore coxa with one small spine; femur unarmed on ventral surface, internal genicular lobe with one small spine, external genicular lobe with one long spine; tibia with two inner spines and two outer spines on ventral surface, apical area with one extero-dorsal spine and a pair of ventral spines, between the paired ventral spines with one small spine. Mid femur unarmed on ventral surface, internal and external genicular lobes each with 1 long spine; tibia with one inner spine and two to three outer spines on ventral surface, apical area with a pair of dorsal spines and a pair of ventral spines, between the paired ventral spines with one small spine. Hind femur with two inner spines on ventral surface; tibia with 56–59 inner spines and 59–62 outer spines on dorsal surface, with a pair of dorsal spines in subapical area, and at apex with a pair of dorsal spines and two pairs of ventral spines, intero-dorsal spine much longer than hind basitarsus. Hind basitarsus with one dorsal apical spine.
All abdominal tergites without process. Epiproct lingulate; paraproct slightly longer than epiproct, semicircular in lateral view. Cercus slender, conical, apex acute. Dorsal sclerite of genitalia nearly semicircular, basal area with a concavity, apical area broadly concave; basal area of dorsal median lobe with paired lateral sclerites, oblong, apical area not divided; dorsal lateral lobes broad and folded, much longer than dorsal median lobe, apical area weakly sclerotized, almost as wide as basal area; ventral median lobe and ventral lateral lobes short. Subgenital plate quadrangular, wide, posterior margin straight.
Female: Appearance is similar to male. Ovipositor longer than half the length of hind femur, dorsal margin smooth, apical area of ventral margin denticulate. Subgenital plate semicircular, posterior margin blunt.
Coloration. Dorsal surface of body brown, ventral surface yellowish brown. Face with four longitudinal black stripes.
Measurements (mm). Body: ♂15.2–18.0, ♀17.8–20.1; pronotum: ♂5.8–7.1, ♀6.0–7.0; fore femur: ♂7.3–8.8, ♀7.0–8.3; hind femur: ♂17.0–19.0, ♀18.1–19.0; hind tibia: ♂15.0–16.9, ♀17.8–19.0; hind basitarsus: ♂4.0–4.8, ♀3.2–4.0; ovipositor: 12.9–15.1.
Distribution. China (Jiangxi, Zhejiang).
Pseudotachycines nephrus Zhu & Shi sp. nov.
urn:lsid:zoobank.org:act:31599921-80F7-4902-9FEC-0FCDCA3B848F
Material examined. Holotype: ♂, China, Zhejiang, Linan, Qingliangfeng, 4.Ⅹ.2019, coll. Q.D. Zhu, dep. HBU. Paratypes: 2♀, same data as for holotype.
Etymology. The name of the new species is the Greek word ‘nephr’ (nephroid), referring to the nephroid lateral sclerites of the dorsal median lobe of the male genitalia.
Diagnosis. The new species is similar to Pseudotachycines zhengi Zhu & Shi sp. nov., but it can easily be distinguished from the latter one by the shape of the male genitalia and female subgenital plate. The dorsal sclerite of the male genitalia is semicircular and the lateral sclerite of the dorsal median lobe is nephroid. The female subgenital plate is trapezoid, with slightly concave posterior margin.
Description. Male: Body medium sized in Pseudotachycines. Fastigium verticis with two conical tubercles, apices rounded, directing forward. Eyes ovoid, protruding forward; median ocellus oval, located between antennal sockets; lateral ocelli circular, situated on lateral margins of basal fastigium verticis. Apical segment of maxillary palpus much longer than subapical segment, apex truncated.
Pronotum long, anterior margin straight, posterior margin arcuate; lateral lobes longer than high, ventral margins arc shaped. Mesonotum and metanotum short, posterior margin of mesonotum arcuate, posterior margin of metanotum straight. Fore coxa with one small spine; femur unarmed on ventral surface, internal genicular lobe with one small spine, external genicular lobe with one long spine; tibia with two inner spines and two outer spines on ventral surface, apical area with one extero-dorsal spine and a pair of ventral spines, between the paired ventral spines with one small spine. Mid femur unarmed on ventral surface, internal and external genicular lobes each with one long spine; tibia with one inner spine and two outer spines on ventral surface, apical area with a pair of dorsal spines and a pair of ventral spines, between the paired ventral spines with one small spine. Hind femur with three to five inner spines on ventral surface, internal genicular lobe with one small spine, external genicular lobe unarmed; tibia with 55–57 inner spines and 55–57 outer spines on dorsal surface, with a pair of dorsal spines in subapical area, and at apex with a pair of dorsal spines and two pairs of ventral spines, intero-dorsal spine much longer than hind basitarsus. Hind basitarsus with one dorsal apical spine.
Posterior margins of all abdominal tergites straight. Epiproct semicircular; paraproct much longer than epiproct, triangular in lateral view. Cercus slender, conical, apex acute. Dorsal sclerite of genitalia semicircular; basal area of dorsal median lobe with paired lateral sclerites, nephroid, apical area not divided; dorsal lateral lobes broad and folded, much longer than dorsal median lobe, apical area weakly sclerotized, almost as wide as basal area; ventral median lobe and ventral lateral lobes short. Subgenital plate quadrangular, posterior margin straight.
Female: Appearance is similar to male. Ovipositor longer than half the length of hind femur, curved upward, dorsal margin smooth, apical area of ventral margin denticulate. Subgenital plate trapezoid, posterior margin slightly concave.
Coloration. Dorsal surface of body light brown, with yellow spots, ventral surface yellowish brown. Eyes black.
Measurements (mm). Body: ♂14.1, ♀17.4–18.3; pronotum: ♂6.6, ♀7.3–8.0; fore femur: ♂8.4, ♀9.0–11.2; hind femur: ♂19.5, ♀21.6–23.2; hind tibia: ♂18.8, ♀21.4–22.5; hind basitarsus: ♂3.3, ♀4.1–4.6; ovipositor: 13.0–15.9.
Distribution. China (Zhejiang).
Pseudotachycines sagittus Zhu & Shi sp. nov.
urn:lsid:zoobank.org:act:65264F0A-DAC2-4CD0-BE06-0097493456ED
Material examined. Holotype: ♂, China, Yunnan, Yiliang, Chaotianma, 11.Ⅷ.2020, coll. T. Wang, dep. HBU. Paratypes: 2♀, same data as for holotype.
Etymology. The name of the new species derives from the Latin word ‘sagitt’, referring to the sagittate-shaped dorsal sclerite of the male genitalia.
Diagnosis. The new species can easily be distinguished from known congeneric species by the shape of the male genitalia and epiproct. The dorsal sclerite of the male genitalia is sagittate and the lateral sclerite of the dorsal median lobe of the male genitalia is oval. The male epiproct is nearly triangular.
Description. Male: Body medium sized in Pseudotachycines. Fastigium verticis short, with two conical tubercles, apices rounded, directing forward. Eyes ovoid, protruding forward; median ocellus oval, located between antennal sockets; lateral ocelli circular, situated on lateral margins of basal fastigium verticis. Apical segment of maxillary palpus much longer than subapical segment, apex truncated.
Pronotum long, anterior margin straight, posterior margin arcuate; lateral lobes longer than high, ventral margins arc shaped. Mesonotum and metanotum short, posterior margin of mesonotum arcuate, posterior margin of metanotum straight. Fore coxa with one small spine; femur unarmed on ventral surface, internal genicular lobe unarmed, external genicular lobe with one long spine; tibia with two inner spines and two outer spines on ventral surface, apical area with one extero-dorsal spine and a pair of ventral spines, between the paired ventral spines with one small spine. Mid femur unarmed on ventral surface, internal and external genicular lobes each with one long spine; tibia with one inner spine and two outer spines on ventral surface, apical area with a pair of dorsal spines and a pair of ventral spines, between the paired ventral spines with one small spine. Hind femur unarmed on ventral surface, internal genicular lobe with one small spine, external genicular lobe unarmed; tibia with 47–52 inner spines and 49–50 outer spines on dorsal surface, with a pair of dorsal spines in subapical area, and at apex with a pair of dorsal spines and two pairs of ventral spines, intero-dorsal spine nearly equal in length to hind basitarsus. Hind basitarsus with one dorsal apical spine.
All abdominal tergites without process. Epiproct nearly triangular, apex blunt; paraproct triangular in lateral view. Cercus slender, conical, apex acute. Dorsal sclerite of genitalia sagittate; basal area of dorsal median lobe with paired lateral sclerites, oval, apical area not divided; dorsal lateral lobes broad and folded, much longer than dorsal median lobe, apical area weakly sclerotized, almost as wide as basal area; ventral median lobe and ventral lateral lobes short. Subgenital plate quadrangular, posterior margin arcuate.
Female: Appearance is similar to male. Ovipositor longer than half the length of hind femur, curved upward, dorsal margin smooth, apical area of ventral margin denticulate. Subgenital plate semicircular, posterior margin slightly prominent.
Coloration. Body light brown, face with four longitudinal brown stripes, eyes black.
Measurements (mm). Body: ♂13.7, ♀15.6–15.8; pronotum: ♂4.6, ♀5.0–5.7; fore femur: ♂7.2, ♀7.0–7.5; hind femur: ♂15.2, ♀15.6–17.4; hind tibia: ♂15.3, ♀15.5–16.9; hind basitarsus: ♂3.0, ♀2.6–2.7; ovipositor: 9.7–9.8.
Distribution. China (Yunnan).
Pseudotachycines fengyangshanensis Zhu & Shi sp. nov.
urn:lsid:zoobank.org:act:1AD154B4-D04F-47F7-988D-8C568B94F515
Material examined. Holotype: ♂, China, Zhejiang, Longquan, Fengyangshan, 18.Ⅸ.2019, coll. Y.X. Zhen, dep. HBU. Paratype: 1♂, same data as for holotype.
Etymology. The name of the new species derives from the type locality.
Diagnosis. The new species is most similar to Pseudotachycines nephrus Zhu & Shi sp. nov., but it can be distinguished by the dorsal sclerite of the male genitalia semicircular, with the basal area slightly concave and the apex truncate, and the lateral sclerite of the dorsal median lobe of the male genitalia oblong.
Description. Male: Body medium sized in Pseudotachycines. Fastigium verticis with two conical tubercles, apices divided, directing forward. Eyes ovoid, protruding forward; median ocellus oval, located between antennal sockets; lateral ocelli circular, situated on lateral margins of basal fastigium verticis. Apical segment of maxillary palpus much longer than subapical segment, apex inflated, globular.
Pronotum long, anterior margin of disc straight, posterior margin arcuate; lateral lobes longer than high, ventral margins arc shaped. Mesonotum and metanotum short, posterior margin of mesonotum arcuate, posterior margin of metanotum straight. Fore coxa with one small spine; femur unarmed on ventral surface, internal genicular lobe with one small spine, external genicular lobe with one long spine; tibia with two inner spines and two outer spines on ventral surface, apical area with one extero-dorsal spine and a pair of ventral spines, between the paired ventral spines with one small spine. Mid femur unarmed on ventral surface, internal and external genicular lobes each with one long spine; tibia with one inner spine and two outer spines on ventral surface, apical area with a pair of dorsal spines and a pair of ventral spines, between the paired ventral spines with one small spine. Hind femur with zero to four spines, internal genicular lobe with one small spine; tibia with 51 inner spines and 58 outer spines on dorsal surface, with a pair of dorsal spines in subapical area, and at apex with a pair of dorsal spines and two pairs of ventral spines, intero-dorsal spine much longer than hind basitarsus. Hind basitarsus with one dorsal apical spine.
All abdominal tergites without process. Epiproct nearly semicircular; paraproct much longer than epiproct, oblong in lateral view. Cercus slender, conical, apex acute. Dorsal sclerite of genitalia semicircular, basal area slightly concave, apex truncate; basal area of dorsal median lobe with paired lateral sclerites, oblong, apical area not divided; dorsal lateral lobes broad and folded, much longer than dorsal median lobe, apical area weakly sclerotized, almost as wide as basal area; ventral median lobe and ventral lateral lobes short. Subgenital plate quadrangular, transverse and wide, posterior margin straight.
Female: Unknown.
Coloration. Body brown on dorsal surface and yellowish brown on ventral surface. Face with four longitudinal black stripes. Basal half of the hind femur with pinnate black stripes.
Measurements (mm). Body: ♂15.3; pronotum: ♂6.1; fore femur: ♂7.8; hind femur: ♂17.1; hind tibia: ♂16.9; hind basitarsus: ♂3.3.
Distribution. China (Zhejiang).
Genus Homotachycines Zhu & Shi gen. nov.
urn:lsid:zoobank.org:act:D01BD682-64B3-4466-A591-DA784748B724
Type species.Homotachycines quadratus Zhu & Shi sp. nov., here designated.
Etymology. The name of the new genus refers to the Latin word ‘homo’ and the genus Tachycines.
Diagnosis. Body is medium sized in Aemodogryllini. The male epiproct and paraproct are normally developed. The male genitalia have unpaired dorsal sclerite; the dorsal median lobe of the male genitalia has paired lateral sclerites at the basal area, the apical area does not divide; the dorsal lateral lobes of the male genitalia are little longer than the dorsal median lobe, the basal area broad, gradually narrowing to apices, the apical area sclerotized.
Homotachycines acutilobatus (Gorochov, 2010) comb. nov.
Diestrammena (Gymnaeta) acutilobata Gorochov, 2010a: 10; Gorochov, 2010b: 13.
Gymnaetoides acutilobatus: Qin, Liu & Li, 2017: 189.
Material examined. 1♂, China, Chongqing, Wushan, Guanyang, 22.Ⅶ.2021, coll. L.Y. Wang, dep. HBU; 4♂, China, Hubei, Shennongjia, Huangbaiqian, 17.Ⅷ.2019, coll. P. Wang, dep. HBU.
Distribution. China (Chongqing, Hubei).
Remarks. The species was transferred to genus
Gymnaetoides according to the sclerotized dorsal lateral lobes of the male genitalia [
13]. However, some characters of the species are inconsistent with
Gymnaetoides: the apical area of the dorsal median lobe of the male genitalia does not divide, and the dorsal lateral lobe of the male genitalia is a little longer than the dorsal median lobe. Therefore, we transfer it into the genus
Homotachycines based on morphological and molecular evidence.
Homotachycines ovalilobatus (Gorochov, 2010) comb. nov.
Diestrammena (Gymnaeta) ovalilobata Gorochov, 2010a: 11.
Pseudotachycines ovalilobatus: Qin, Liu & Li, 2017: 488.
Material examined. 2♂2♀, China, Hubei, Baokang, Houping, 8.Ⅷ.2020, coll. G.L. Xie, dep. HBU; 2♂2♀, China, Hubei, Shennongjia, Huangbaiqian, 17.Ⅷ.2019, coll. P. Wang, dep. HBU.
Distribution. China (Hubei).
Remarks. The species was moved to the genus
Pseudotachycines according to the generic diagnosis [
14]. However, there are some characters that differ from
Pseudotachycines: the male paraproct is normally developed; the dorsal lateral lobes of the male genitalia are little longer than dorsal median lobe, with the basal area broad, gradually narrowing to apices, the apical area sclerotized, and oval lobate. Therefore, it should be transferred into
Homotachycines based on morphological and molecular evidence.
Homotachycines trapezialis (Qin, Liu & Li, 2017) comb. nov.
Pseudotachycines trapezialis Qin, Liu & Li, 2017: 487.
Material examined. Holotype: ♂, China, Henan, Funiu Mountain, 2.Ⅷ.2009, coll. W.B. Zhu, dep. SEMCAS. Paratype: 1♀, same data as for holotype.
Distribution. China (Henan).
Remarks. The line drawing of the male genitalia of this species did not adequately represent the characters (Figure 19 in [
14]). The photographs of the type specimens are shown in
Figure 19. The male paraproct is normally developed. The apical area of the dorsal median lobe of the male genitalia does not divide. The dorsal lateral lobes of the male genitalia are longer than the dorsal median lobe, the basal area is broad, gradually narrowing to apices, and the apical area is sclerotized. According to these characters, this species should be moved to genus
Homotachycines. The species is similar to
Homotachycines ovalilobatus comb. nov. in the shape of the male genitalia, but the female subgenital plate is quite different.
Homotachycines triangulus Zhu & Shi sp. nov.
urn:lsid:zoobank.org:act:85851298-45FA-4F51-9858-85EC8A4E7F3A
Material examined. Holotype: ♂, China, Hubei, Shennongjia, Maohu Protection Station, 15.Ⅷ.2018, coll. P. Wang, dep. HBU. Paratype: 1♂, China, Hubei, Shennongjia, Chaoshuihe, 8.Ⅷ.2018, coll. P. Wang, dep. HBU.
Etymology. The name of the new species derives from the Latin word ‘tri’ (triple) and ‘angul’ (angle), referring to the triangular dorsal sclerite of the male genitalia.
Diagnosis. The new species differs from other congeneric species by the shapes of the dorsal sclerite of the male genitalia and the lateral sclerite of the dorsal median lobe of the male genitalia. The dorsal sclerite of the male genitalia is triangular, with a broadly concave apical area, and the lateral sclerite of the dorsal median lobe is quadrangular.
Description. Male: Body medium sized in Homotachycines. Fastigium verticis with two conical tubercles, apices rounded, directing forward. Eyes ovoid, protruding forward; median ocellus oval, located between antennal sockets; lateral ocelli circular, situated on lateral margins of basal fastigium verticis. Apical segment of maxillary palpus much longer than subapical segment, apex truncated.
Pronotum long, anterior margin straight, posterior margin arcuate; lateral lobes longer than high, ventral margins arc shaped. Mesonotum and metanotum short, posterior margin of mesonotum arcuate, posterior margin of metanotum straight. Fore coxa with one small spine; femur unarmed on ventral surface, internal genicular lobe with one small spine, external genicular lobe with one long spine; tibia with two inner spines and two outer spines on ventral surface, apical area with one extero-dorsal spine and a pair of ventral spines, between the paired ventral spines with one small spine. Mid femur unarmed on ventral surface, internal and external genicular lobes each with one long spine; tibia with one inner spine and two outer spines on ventral surface, apical area with a pair of dorsal spines and a pair of ventral spines, between the paired ventral spines with one small spine. Hind femur with zero to one inner spine on ventral surface; tibia with 52–60 inner spines and 53–57 outer spines on dorsal surface, with a pair of dorsal spines in subapical area, and at apex with a pair of dorsal spines and two pairs of ventral spines, intero-dorsal spine distinctly longer than hind basitarsus. Hind basitarsus with one dorsal apical spine.
Posterior margins of all abdominal tergites straight. Epiproct semicircular; paraproct simple, triangular in lateral view. Cercus slender, conical, apex acute. Dorsal sclerite of genitalia triangular, apical area broadly concave; basal area of dorsal median lobe with paired lateral sclerites, quadrangular, apical area not divided; dorsal lateral lobes little longer than dorsal median lobe, basal area broad, gradually narrowing to apices, apical area sclerotized, hooked; ventral median lobe and ventral lateral lobes very short. Subgenital plate quadrangular, wide, posterior margin straight.
Female: Unknown.
Coloration. Body light brown. Eyes black, ocelli yellow. Face with four longitudinal black stripes, apical areas of all femora black.
Measurements (mm). Body: ♂15.5–17.8; pronotum: ♂5.8–6.0; fore femur: ♂6.2–7.0; hind femur: ♂15.3–16.2; hind tibia: ♂14.5–16.0; hind basitarsus: ♂3.0–3.2.
Distribution. China (Hubei).
Homotachycines quadratus Zhu & Shi sp. nov.
urn:lsid:zoobank.org:act:2E728F0F-915C-4C36-98E3-B1CC9E531A7D
Material examined. Holotype: ♂, China, Hubei, Shennongjia, Banbiyan, 14.Ⅷ.2018, coll. P. Wang, dep. HBU. Paratypes: 2♀, same data as for holotype. Other material: 1♀, same data as for holotype.
Etymology. The name of the new species is the Latin word ‘quadrat’ (quadrate), referring to the dorsal sclerite of the male genitalia having a quadrate shape.
Diagnosis. The new species is similar to Homotachycines acutilobatus comb. nov., but it can be distinguished from the latter one by the apical area of the dorsal sclerite of the male genitalia slightly concave and the lateral sclerite of the dorsal median lobe of the male genitalia crescent shaped.
Description. Male: Body medium sized in Homotachycines. Fastigium verticis short, with two conical tubercles, directing forward. Eyes ovoid, protruding forward; median ocellus oval, located between antennal sockets; lateral ocelli circular, situated on lateral margins of basal fastigium verticis. Apical segment of maxillary palpus much longer than subapical segment, apex truncated.
Pronotum long, anterior margin straight, posterior margin arcuate; lateral lobes longer than high, ventral margins arc shaped. Mesonotum and metanotum short, posterior margin of mesonotum arcuate, posterior margin of metanotum straight. Fore coxa with one small spine; femur unarmed on ventral surface, internal genicular lobe unarmed, external genicular lobe with 1 long spine; tibia with two inner spines and two outer spines on ventral surface, apical area with one extero-dorsal spine and a pair of ventral spines, between the paired ventral spines with one small spine. Mid femur unarmed on ventral surface, internal and external genicular lobes each with one long spine; tibia with one inner spine and one outer spine on ventral surface, apical area with a pair of dorsal spines and a pair of ventral spines, between the paired ventral spines with one small spine. Hind femur unarmed on ventral surface; tibia with 68–75 inner spines and 70–74 outer spines on dorsal surface, with a pair of dorsal spines in subapical area, and at apex with a pair of dorsal spines and two pairs of ventral spines, intero-dorsal spine slightly shorter than hind basitarsus. Hind basitarsus with one dorsal apical spine.
Posterior margins of all abdominal tergites straight. Epiproct semicircular; paraproct simple, triangular in lateral view. Cercus slender, conical, apex acute. Dorsal sclerite of genitalia quadrate, apical area slightly concave; basal area of dorsal median lobe with paired lateral sclerites, crescent-shaped, apical area not divided; dorsal lateral lobes little longer than dorsal median lobe, basal area broad, gradually narrowing to apices, apical area sclerotized, hooked; ventral median lobe and ventral lateral lobes short. Subgenital plate trapezoidal, posterior margin straight.
Female: Appearance is similar to male. Ovipositor short, curved upward, dorsal margin smooth, apical area of ventral margin denticulate. Subgenital plate trapezoid, posterior margin straight.
Coloration. Dorsal surface of body brown, ventral surface yellowish brown. Legs with black stripes.
Measurements (mm). Body: ♂17.0, ♀17.0–17.5; pronotum: ♂4.8, ♀5.0–6.0; fore femur: ♂8.0, ♀8.8–9.0; hind femur: ♂15.6, ♀16.0–18.2; hind tibia: ♂16.2, ♀16.2–18.0; hind basitarsus: ♂3.2, ♀3.5–4.0; ovipositor: 9.0–10.4.
Distribution. China (Hubei).
Homotachycines baokangensis Zhu & Shi sp. nov.
urn:lsid:zoobank.org:act:CE6C4893-12E1-4C96-A2DD-5D67369ED27B
Material examined. Holotype: ♂, China, Hubei, Baokang, Houping, 8.Ⅷ.2020, coll. G.L. Xie, dep. HBU.
Etymology. The name of the new species refers to the type locality.
Diagnosis. The new species can be distinguished from known congeneric species by the shape of the male genitalia. The dorsal sclerite of the male genitalia is quadrangular, with the basal half rectangular and the apical half wide and slightly concave. The lateral sclerite of the dorsal median lobe of the male genitalia is oblong.
Description. Male: Body medium sized in Homotachycines. Fastigium verticis with two conical tubercles, apices rounded, directing forward. Eyes ovoid, protruding forward; median ocellus oval, located between antennal sockets; lateral ocelli circular, situated on lateral margins of basal fastigium verticis. Apical segment of maxillary palpus much longer than subapical segment, apex inflated, globular.
Pronotum long, anterior margin straight, posterior margin arcuate; lateral lobes longer than high, ventral margins arc shaped. Mesonotum and metanotum short, posterior margin of mesonotum arcuate, posterior margin of metanotum straight. Fore coxa with one small spine; femur unarmed on ventral surface, internal genicular lobe with 1 small spine, external genicular lobe with one long spine; tibia with two inner spines and two outer spines on ventral surface, apical area with one extero-dorsal spine and a pair of ventral spines, between the paired ventral spines with one small spine. Mid femur unarmed on ventral surface, internal and external genicular lobes each with one long spine; tibia with one inner spine and two outer spines on ventral surface, apical area with a pair of dorsal spines and a pair of ventral spines, between the paired ventral spines with one small spine. Hind femur with two inner spines on ventral surface, internal genicular lobe unarmed; tibia with 79 inner spines and 74 outer spines on dorsal surface, at apex with a pair of dorsal spines and two pairs of ventral spines, intero-dorsal spine nearly equal in length to hind basitarsus. Hind basitarsus with one dorsal apical spine.
Posterior margins of all abdominal tergites straight. Epiproct semicircular; paraproct triangular in lateral view. Cercus slender, conical, apex acute. Dorsal sclerite of genitalia quadrangular, basal half rectangular, apical half wide, slightly concave; basal area of dorsal median lobe with paired lateral sclerites, oblong, apical area not divided; dorsal lateral lobes little longer than dorsal median lobe, basal area broad, gradually narrowing to apices, apical area sclerotized, hooked; ventral median lobe and ventral lateral lobes short. Subgenital plate transverse and wide, posterior margin straight.
Female: Unknown.
Coloration. Body light brown, with yellow spots. Eyes black. Apices of all femora with ring-like black stripes.
Measurements (mm). Body: ♂16.6; pronotum: ♂5.4; fore femur: ♂10.3; hind femur: ♂19.8; hind tibia: ♂20.8; hind basitarsus: ♂4.0.
Distribution. China (Hubei).
Homotachycines fusus Zhu & Shi sp. nov.
urn:lsid:zoobank.org:act:85F797F6-1C52-4667-9639-B740D5F53059
Material examined. Holotype: ♂, China, Henan, Luoyang, Longyuwan, 8.Ⅶ.2021, coll. G.L. Hu, dep. HBU. Paratype: 1♂, same data as for holotype. Other material: 1♀ (nymph), same data as for holotype.
Etymology. The name of the new species derives from the Latin word ‘fus’ (fusiform), referring to the lateral sclerite of dorsal median lobe of the male genitalia fusiform.
Diagnosis. The new species is distinguished from known congeneric species by the structure of the male genitalia. The dorsal sclerite of the male genitalia is quadrate, with the basal and lateral margins slightly concave. The lateral sclerite of the dorsal median lobe of the male genitalia is fusiform.
Description. Male: Body medium sized in Homotachycines. Fastigium verticis with two conical tubercles, apices rounded, directing forward. Eyes ovoid, protruding forward; median ocellus oval, located between antennal sockets; lateral ocelli circular, situated on lateral margins of basal fastigium verticis. Apical segment of maxillary palpus much longer than subapical segment, apex inflated, globular.
Pronotum long, anterior margin straight, posterior margin arcuate; lateral lobes longer than high, ventral margins arc shaped. Mesonotum and metanotum short, posterior margin of mesonotum arcuate, posterior margin of metanotum straight. Fore coxa with one small spine; femur unarmed on ventral surface, internal genicular lobe unarmed, external genicular lobe with one long spine; tibia with two inner spines and two outer spines on ventral surface, apical area with one extero-dorsal spine and a pair of ventral spines, between the paired ventral spines with one small spine. Mid femur unarmed on ventral surface, internal and external genicular lobes each with one long spine; tibia with one inner spine and two outer spines on ventral surface, apical area with a pair of dorsal spines and a pair of ventral spines, between the paired ventral spines with one small spine. Hind femur with three to five inner spines on ventral surface, internal genicular lobe unarmed; tibia with 51–58 inner spines and 54–59 outer spines on dorsal surface, with a pair of dorsal spines in subapical area, and at apex with a pair of dorsal spines and two pairs of ventral spines, intero-dorsal spine longer than hind basitarsus. Hind basitarsus with one dorsal apical spine.
Posterior margins of all abdominal tergites straight. Epiproct semicircular; paraproct triangular in lateral view. Cercus slender, conical, apex acute. Dorsal sclerite of genitalia quadrate, basal and lateral margins slightly concave; basal area of dorsal median lobe with paired lateral sclerites, fusiform, apical area not divided; dorsal lateral lobes little longer than dorsal median lobe, basal area broad, gradually narrowing to apices, apical area sclerotized, hooked; ventral median lobe and ventral lateral lobes short. Subgenital plate transverse and wide, posterior margin straight.
Female (nymph): Appearance is similar to male. Ovipositor longer than half the length of hind femur, dorsal and ventral margins smooth. Subgenital plate trapezoid, posterior margin with a concavity.
Coloration. Body yellowish brown, apex of abdomen black. Face with four longitudinal black stripes, eyes black.
Measurements (mm). Body: ♂12.4–13.3, ♀15.4; pronotum: ♂6.0–6.4, ♀6.2; fore femur: ♂9.1–9.5, ♀8.4; hind femur: ♂18.7–19.1, ♀17.6; hind tibia: ♂19.8–20.3, ♀18.1; hind basitarsus: ♂3.5–4.1, ♀3.4; ovipositor: 13.8.
Distribution. China (Henan).
Homotachycines concavus Zhu & Shi sp. nov.
urn:lsid:zoobank.org:act:04A2480F-C810-4169-9B5F-4B02C2ECD16C
Material examined. Holotype: ♂, China, Henan, Luoyang, Longyuwan, 8.Ⅶ.2021, coll. G.L. Hu, dep. HBU. Other material: 1♀ (nymph), same data as for holotype; 3♂, China, Shaanxi, Feng County, Hekou, 6.Ⅶ.2021, coll. J. Li, dep. HBU; 1♂, China, Shaanxi, Liuba, Zhangliangmiao, 4.Ⅷ.2017, coll. Z.X. Li, dep. HBU; 2♂, China, Shaanxi, Liuba, Zhangliangmiao, 11.Ⅶ.2021, coll. J. Li, dep. HBU; 2♂, China, Shaanxi, Mian County, Miaoping, 22.Ⅶ.2021, coll. J. Li, dep. HBU.
Etymology. The name of the new species is the Latin words ‘concav’ (concave), referring to the concave posterior margin of male subgenital plate.
Diagnosis. The new species can easily be distinguished from known congeneric species by the structure of the male genitalia. The basal two-thirds of the dorsal sclerite of the male genitalia are narrow and obtuse, the apical one-third is wide with the apex concave. The lateral sclerite of the dorsal median lobe of the male genitalia is nearly rectangular.
Description. Male: Body medium sized in Homotachycines. Fastigium verticis with two conical tubercles, apices rounded, directing forward. Eyes ovoid, protruding forward; median ocellus oval, located between antennal sockets; lateral ocelli circular, situated on lateral margins of basal fastigium verticis. Apical segment of maxillary palpus much longer than subapical segment, apex inflated, globular.
Pronotum long, anterior margin of disc straight, posterior margin arcuate; lateral lobes longer than high, ventral margins arc shaped. Mesonotum and metanotum short, posterior margin of mesonotum arcuate, posterior margin of metanotum straight. Fore coxa with one small spine; femur unarmed on ventral surface, internal genicular lobe with one small spine, external genicular lobe with one long spine; tibia with two inner spines and two outer spines on ventral surface, apical area with one extero-dorsal spine and a pair of ventral spines, between the paired ventral spines with one small spine. Mid femur unarmed on ventral surface, internal and external genicular lobes each with one long spine; tibia with one inner spine and one outer spine on ventral surface, apical area with a pair of dorsal spines and a pair of ventral spines, between the paired ventral spines with one small spine. Hind femur with three to eleven inner spines on ventral surface, internal genicular lobe unarmed; tibia with 60 inner spines and 54 outer spines on dorsal surface, with a pair of dorsal spines in subapical area, and at apex with a pair of dorsal spines and two pairs of ventral spines, intero-dorsal spine longer than hind basitarsus. Hind basitarsus with one dorsal apical spine.
All abdominal tergites without process. Epiproct semicircular; paraproct triangular in lateral view. Cercus slender, conical, apex acute. Basal two thirds of dorsal sclerite of genitalia narrow, obtuse, apical one third wide, apex concave; basal area of dorsal median lobe with paired lateral sclerites, nearly rectangular, apical area not divided; dorsal lateral lobes longer than dorsal median lobe, basal area broad, gradually narrowing to apices, apical area weakly sclerotized; ventral median lobe and ventral lateral lobes short. Subgenital plate transverse and wide, posterior margin concave.
Female (nymph): Appearance is similar to male. Ovipositor longer than half the length of hind femur, with dorsal and ventral margins smooth. Subgenital plate trapezoid, posterior margin straight.
Coloration. Body light brown, face with two longitudinal black stripes, eyes black.
Measurements (mm). Body: ♂15.1–15.5, ♀13.6; pronotum: ♂5.0–5.6, ♀5.3; fore femur: ♂7.4–7.7, ♀7.6; hind femur: ♂15.9–16.2, ♀15.7; hind tibia: ♂15.6–16.1, ♀15.5; hind basitarsus: ♂3.1–3.2, ♀3.2; ovipositor: 12.5.
Distribution. China (Henan, Shaanxi).
Homotachycines qinlingensis Zhu & Shi sp. nov.
urn:lsid:zoobank.org:act:0755A42B-F2A2-4B61-92A6-01C28FE56EFA
Material examined. Holotype: ♂, China, Shaanxi, Liuba, Huoshaodian, 4.Ⅹ.2020, coll. J. Li, dep. HBU. Paratype: 1♂, China, Shaanxi, Lueyang, Wulongdong, 11.Ⅹ.2020, coll. J. Li, dep. HBU. Other material: 3♂, China, Shaanxi, Yang County, Huayang town, 6.Ⅷ.2017, coll. H.Y. Liu, dep. HBU.
Etymology. The name of the new species derives from the type locality, which is located in the Qinling Mountains.
Diagnosis. The new species can be distinguished from known congeneric species by the dorsal sclerite of the male genitalia trapezoidal, the basal area with a concavity, and the apical area that is widely concave; the lateral sclerite of the dorsal median lobe of the male genitalia is oval.
Description. Male: Body medium sized in Homotachycines. Fastigium verticis with two conical tubercles, apices rounded, directing forward. Eyes ovoid, protruding forward; median ocellus oval, located between antennal sockets; lateral ocelli circular, situated on lateral margins of basal fastigium verticis. Apical segment of maxillary palpus much longer than subapical segment, apex inflated, globular.
Pronotum long, anterior margin of disc straight, posterior margin arcuate; lateral lobes longer than high, ventral margins arc shaped. Mesonotum and metanotum short, posterior margin of mesonotum arcuate, posterior margin of metanotum straight. Fore coxa with one small spine; femur with 19–25 spines on ventral surface, internal genicular lobe with one small spine, external genicular lobe with one long spine; tibia with two inner spines and two outer spines on ventral surface, apical area with one extero-dorsal spine and a pair of ventral spines, between the paired ventral spines with one small spine. Mid femur unarmed on ventral surface, internal and external genicular lobes each with one long spine; tibia with one inner spine and two outer spines on ventral surface, apical area with a pair of dorsal spines and a pair of ventral spines, between the paired ventral spines with one small spine. Hind femur with two to four inner spines on ventral surface, internal genicular lobe with one small spine; tibia with 53–57 inner spines and 55–59 outer spines on dorsal surface, with a pair of dorsal spines in subapical area, and at apex with a pair of dorsal spines and two pairs of ventral spines, intero-dorsal spine slightly longer than hind basitarsus. Hind basitarsus with one dorsal apical spine.
All abdominal tergites without process. Epiproct semicircular; paraproct triangular in lateral view. Cercus slender, conical, apex acute. Dorsal sclerite of genitalia trapezoidal, basal area with a concavity, apical area widely concave; basal area of dorsal median lobe with paired lateral sclerites, oval, apical area not divided; dorsal lateral lobes longer than dorsal median lobe, basal area broad, gradually narrowing to apices, apical area sclerotized, apex acute; ventral median lobe and ventral lateral lobes short, cylindrical. Subgenital plate transverse and wide, posterior margin slightly concave.
Female: Unknown.
Coloration. Body light brown. Face with two longitudinal black stripes, eyes black, ocelli yellow. All femora with ring black stripes, basal half of hind femur with pinnate black stripes.
Measurements (mm). Body: ♂14.3–16.6; pronotum: ♂5.3–5.4; fore femur: ♂7.4–7.5; hind femur: ♂15.2–16.5; hind tibia: ♂14.8–17.2; hind basitarsus: ♂3.1–3.2.
Distribution. China (Shaanxi).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

























