Sanitation Improves Stored Product Insect Pest Management

 ,
, 
Abstract
:
1. Sanitation Historically and Its Contemporary Significance
2. Materials and Methods
3. Sanitation and Chemical Control
4. Sanitation and Physical/Cultural Control Measures
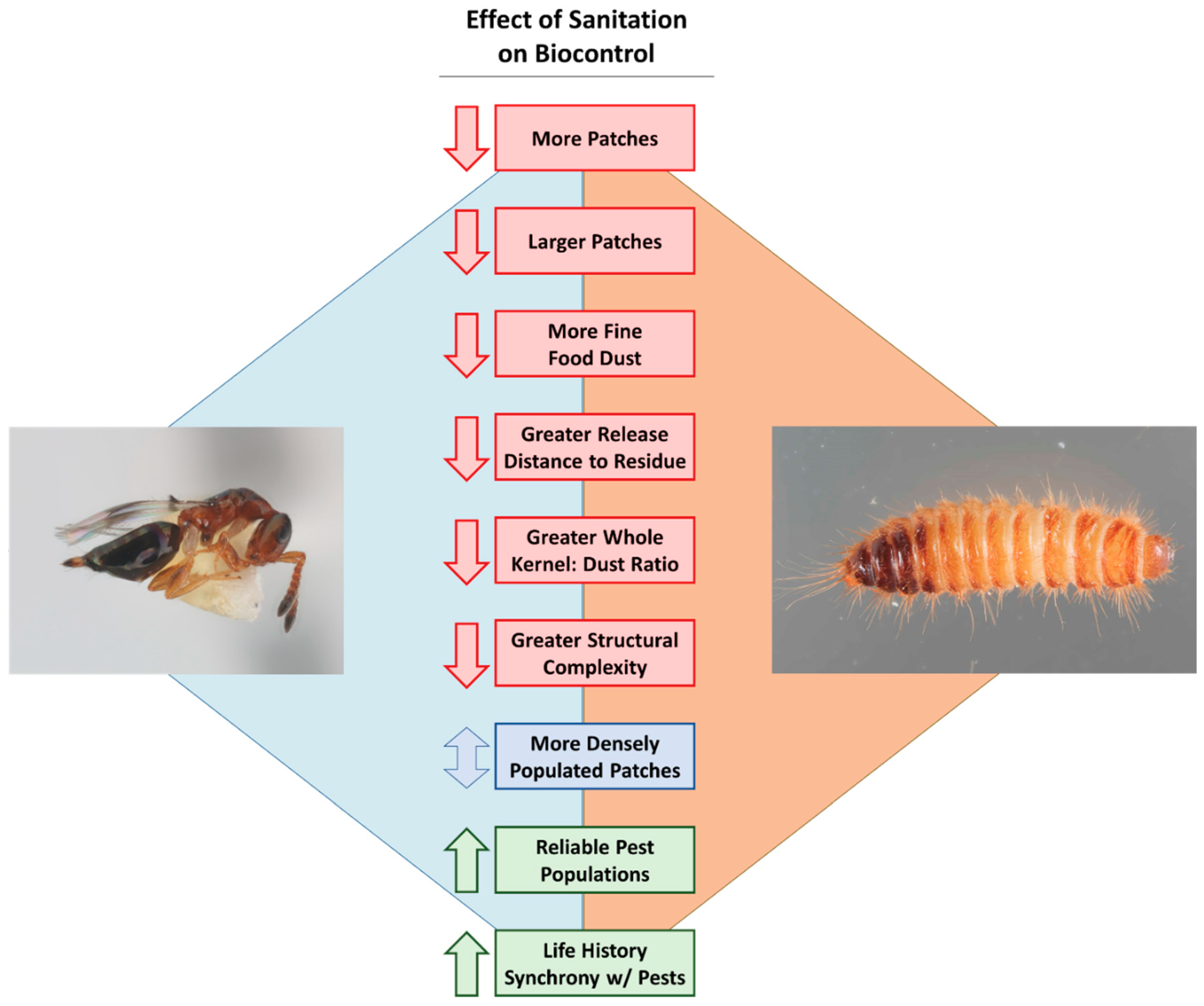
5. Sanitation and Biological Control
6. Sanitation and Behaviorally-Based Management
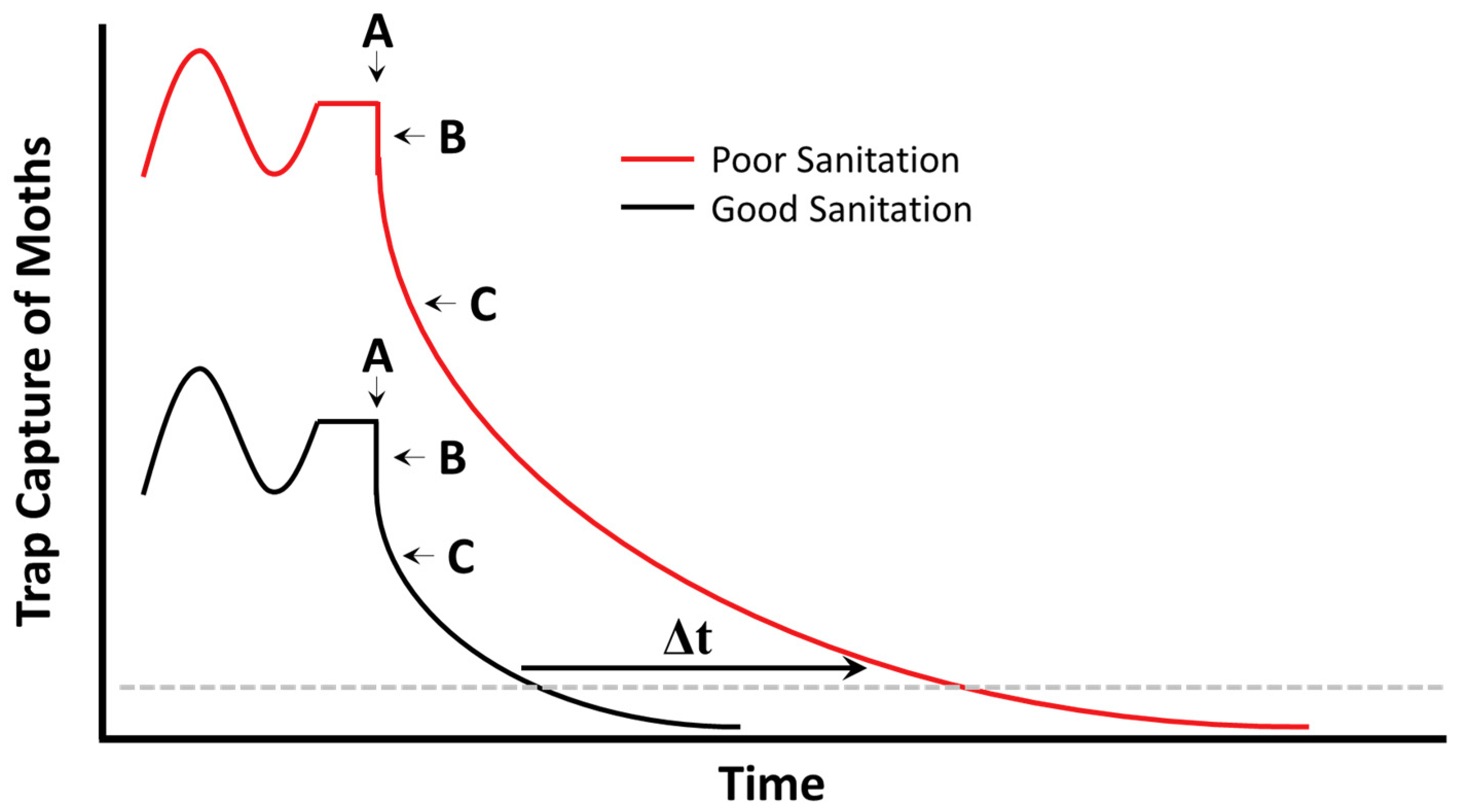
7. Sanitation and Monitoring Techniques
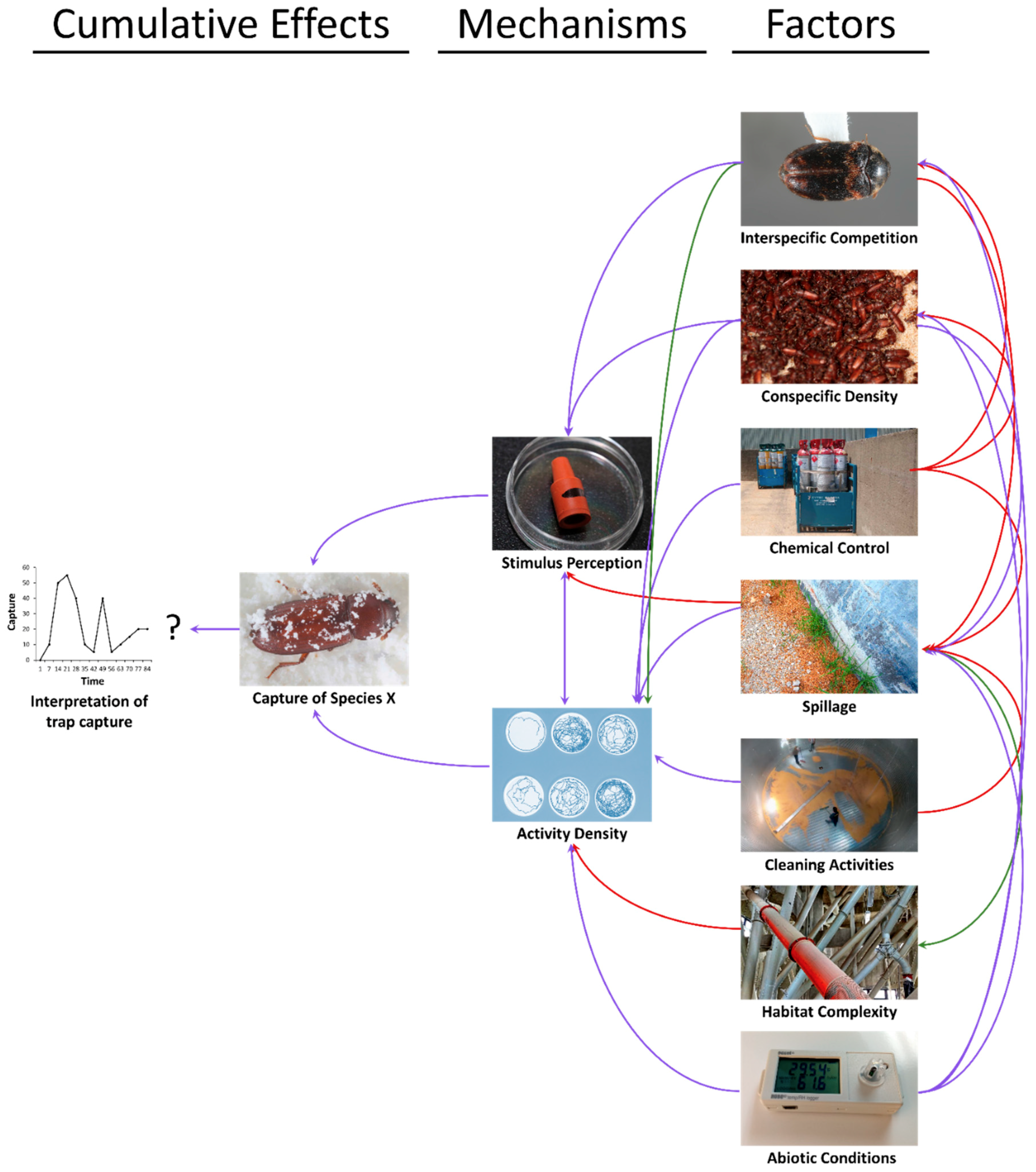
8. Sanitation and Other Factors
9. Gaps in Knowledge and Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nayak, M.K.; Daglish, G.J. Importance of Stored Product Insects. In Recent Advances in Stored Product Protection; Athanassiou, C.G., Arthur, F.H., Eds.; Springer: Berlin, Germany, 2018; pp. 1–17. [Google Scholar]
- Hagstrum, D.W.; Phillips, T.W. Evolution of stored-product entomology: Protecting the world food supply. Annu. Rev. Entomol. 2017, 62, 379–397. [Google Scholar] [CrossRef] [PubMed]
- Buckland, P.C. The early dispersal of insect pests of stored products as indicated by the archaeological records. J. Stored Prod. Res. 1981, 17, 1–12. [Google Scholar] [CrossRef]
- Mason, L. Insects and Mites. In Food Plant Sanitation; Hui, Y.H., Bruinsma, L.B., Gorham, J.R., Nip, W.-K., Tong, P.S., Ventresca, P., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 2003; pp. 293–316. [Google Scholar]
- Davis, R. Introduction to Food-Pest Ecology and Management. In Ecology and Management of Food-Industry Pests1; Gorham, J.R., Ed.; Assocation of Official Analytical Chemists: Arlington, VA, USA, 1991; pp. 3–4. [Google Scholar]
- Wacker, F. Food waste and food losses—Importance of international partnerships and research. In Proceedings of the 12th International Working Conference on Stored Product Protection, Berlin, Germany, 7–11 October 2018; Adler, C., Blank, C., Fuerstenau, B., Kern, P., Mueller-Blenkle, C., Eds.; Julius-Kuhn Institut: Berlin, Germany, 2018; p. 31. [Google Scholar]
- Semeao, A.A.; Campbell, J.F.; Hutchinson, J.M.S.; Whitworth, R.J.; Sloderbeck, P.E. Spatio-temporal distribution of stored-product insects around food processing and storage facilities. Agric. Ecosyst. Environ. 2013, 165, 151–162. [Google Scholar] [CrossRef]
- Tilley, D.R.; Casada, M.E.; Subramanyam, B.; Arthur, F.H. Temporal changes in stored-product insect populations associated with boot, pit, and load-out areas of grain elevators and feed mills. J. Stored Prod. Res. 2017, 73, 62–73. [Google Scholar] [CrossRef]
- Tilley, D.R.; Casada, M.E.; Langemeier, M.R.; Subramanyam, B.; Arthur, F.H. Economic analysis for commingling effects of insect activity in the elevator boot area. J. Econ. Entomol. 2015, 108, 2800–2807. [Google Scholar] [CrossRef]
- Stejskal, V.; Hubert, J.; Aulicky, R.; Kucerova, Z. Overview of present and past and pest-associated risks in stored food and feed products: European perspective. J. Stored Prod. Res. 2015, 64, 122–132. [Google Scholar] [CrossRef]
- Arthur, F.H.; Hagstrum, D.W.; Flinn, P.W.; Reed, C.R.; Phillips, T.W. Insect populations in grain residues associated with commercial Kansas grain elevators. J. Stored Prod. Res. 2006, 42, 226–239. [Google Scholar] [CrossRef]
- Edde, P.A. A review of the biology and control of Rhyzopertha dominica (F.) the lesser grain borer. J. Stored Prod. Res. 2012, 48, 1–18. [Google Scholar] [CrossRef]
- Mahroof, R.M.; Edde, P.A.; Robertson, B.; Puckette, J.A.; Phillips, T.W. Dispersal of Rhyzopertha dominica (Coleoptera: Bostrichidae) in different habitats. Environ. Entomol. 2010, 39, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.F.; Mullen, M.A. Distribution and dispersal behavior of Trogoderma variabile and Plodia interpunctella outside a food processing plant. J. Econ. Entomol. 2004, 97, 1455–1464. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.F.; Arbogast, R.T. Stored-product insects in a flour mill: Population dynamics and response to fumigation treatments. Entomol. Exp. Appl. 2004, 112, 217–225. [Google Scholar] [CrossRef]
- Toews, M.D.; Campbell, J.F.; Arthur, F.H.; Ramaswamy, S.B. Outdoor flight activity and immigration of Rhyzopertha dominica into seed wheat warehouses. Entomol. Exp. Appl. 2006, 121, 73–85. [Google Scholar] [CrossRef]
- McKay, T.; White, A.L.; Starkus, L.A.; Arthur, F.H.; Campbell, J.F. Seasonal patterns of stored-product insects at a rice mill. J. Econ. Entomol. 2017, 110, 1366–1376. [Google Scholar] [CrossRef]
- Campbell, J.F.; Ching’oma, G.P.; Toews, M.D.; Ramaswamy, S.B. Spatial distribution and movement patterns of stored-product insects. In Proceedings of the 9th International Working Conference on Stored Product Protection, São Paulo, Brazil, 15–18 October 2006; pp. 15–18. [Google Scholar]
- Morrison, W.R.; Wilkins, R.V.; Gerken, A.R.; Scheff, D.S.; Zhu, K.Y.; Arthur, F.H.; Campbell, J.F. Mobility of adult Tribolium castaneum (Coleoptera: Tenebrionidae) and Rhyzopertha dominica (Coleoptera: Bostrichidae) after exposure to long-lasting insecticide-incorporated netting. J. Econ. Entomol. 2018, 111, 2443–2453. [Google Scholar] [CrossRef]
- Roesli, R.; Subramanyam, B.; Campbell, J.F.; Kemp, K. Stored-product insects associated with a retail pet store chain in Kansas. J. Econ. Entomol. 2003, 96, 1958–1966. [Google Scholar] [CrossRef]
- Beck, S.D. Growth and retrogression in larvae of Trogoderma glabrum (Coleoptera: Dermestidae). 3. Ecdysis and form determination. Ann. Entomol. Soc. Am. 1972, 64, 149–155. [Google Scholar] [CrossRef]
- Beck, S.D.; Bharadwaj, R.K. Reversed development and cellular aging in an insect. Science 1972, 178, 1210–1211. [Google Scholar] [CrossRef]
- Tuda, M. Evolutionary diversification of bruchine beetles: Climate-dependent traits and development associated with pest status. Bull. Entomol. Res. 2011, 101, 415–422. [Google Scholar] [CrossRef]
- Campbell, J.F.; Hagstrum, D.W. Patch exploitation by Tribolium castaneum: Movement patterns, distribution, and oviposition. J. Stored Prod. Res. 2002, 38, 55–68. [Google Scholar] [CrossRef]
- Flinn, P.W.; Campbell, J.F. Effects of flour conditioning on cannibalism of T. castaneum eggs and pupae. Environ. Entomol. 2012, 41, 1501–1504. [Google Scholar] [CrossRef] [PubMed]
- Ghent, A.W. Studies of behavior of the Tribolium flour beetles I. Contrasting responses of T. castaneum and T. confusum to fresh and conditioned flours. Ecology 1963, 44, 269–283. [Google Scholar] [CrossRef]
- Mueller, D.K. Reducing Customer Complaints in Stored Products; Beckett-Highland Publishing: Carmel, IN, USA, 2010. [Google Scholar]
- Chowaniec, T. Sanitation Cost—The Bottom Line; Association of Operative Millers Bulletin: Kansas City, MO, USA, December 1986; pp. 4867–4871. [Google Scholar]
- Rolston, L.H.; Rouse, P. Insects in stored rice can be controlled by sanitation. Arkansas Farm Res. 1957, 6, 3. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 15 December 2016).
- Haritos, V.S.; Damcevski, K.A.; Dojchinov, G. Improved efficacy of ethyl formate against stored grain insects by combination with carbon dioxide in a “dynamic” application. Pest Manag. Sci. 2006, 62, 325–333. [Google Scholar] [CrossRef]
- Leesch, J.G. Carbon dioxide on the penetration and distribution of phosphine through wheat. J. Econ. Entomol. 1992, 85, 157–161. [Google Scholar] [CrossRef]
- Ren, Y.; Lee, B.; Padovan, B. Penetration of methyl bromide, sulfuryl fluoride, ethanedinitrile and phosphine into timber blocks and the sorption rate of the fumigants. J. Stored Prod. Res. 2011, 47, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Mason, L.; Tsai, W.; Ileleji, K. Influence of sanitation on post-fumigation pest rebound. In Proceedings of the 10th International Working Conference on Stored Product Protection, Estoril, Portugal, 27 June–2 July 2010; Carvalho, M.O., Fields, P.G., Adler, C.S., Arthur, F.H., Athanassiou, C.G., Campbell, J.F., Fleurat-Lessard, F., Flinn, P.W., Hodges, R.J., Isikber, A.A., et al., Eds.; 2010; p. 983. [Google Scholar]
- Athanassiou, C.G.; Papagregoriou, A.S.; Buchelos, C.T. Insecticidal and residual effect of three pyrethroids against Sitophilus oryzae (L.) (Coleoptera: Curculionidae) on stored wheat. J. Stored Prod. Res. 2004, 40, 289–297. [Google Scholar] [CrossRef]
- Kavallieratos, N.G.; Athanassiou, C.G.; Hatzikonstantinou, A.N.N.N.; Kavallieratou, H.N. Abiotic and biotic factors affect efficacy of chlorfenapyr for control of stored-product insect pests. J. Food Prot. 2011, 74, 1288–1299. [Google Scholar] [CrossRef]
- Collins, P.J.; Lambkin, T.M.; Bridgeman, B.W.; Pulvirenti, C. Resistance to grain-protectant insecticides in coleopterous pests of stored cereals in Queensland, Australia. J. Econ. Entomol. 1993, 86, 239–245. [Google Scholar] [CrossRef]
- Arthur, F.H.; Campbell, J.F. Distribution and efficacy of pyrethrin aerosol to control Tribolium confusum (Coleoptera: Tenebrionidae) in food storage facilities. J. Stored Prod. Res. 2008, 44, 58–64. [Google Scholar] [CrossRef]
- Toews, M.D.; Campbell, J.F.; Arthur, F.H. The presence of flour affects the efficacy of aerosolized insecticides used to treat the red flour beetle, Tribolium castaneum. J. Insect Sci. 2010, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Arthur, F.H.; Campbell, J.F.; Ducatte, G.R. Susceptibility of Tribolium confusum (Coleoptera: Tenebrionidae) to pyrethrin aerosol: Effects of aerosol particle size, concentration, and exposure conditions. J. Econ. Entomol. 2014, 107, 2239–2251. [Google Scholar] [CrossRef]
- Arthur, F.H.; Campbell, J.F.; Fontenot, E.A.; Toews, M.D. Assessing effects of esfenvalerate aerosol applications on resident populations of Tribolium castaneum (Herbst), the red flour beetle, through direct and indirect sampling. J. Stored Prod. Res. 2013, 53, 1–6. [Google Scholar] [CrossRef]
- Arthur, F.H. Impact of accumulated food on survival of Tribolium castaneum on concrete treated with cyfluthrin wettable powder. J. Stored Prod. Res. 2000, 36, 15–23. [Google Scholar] [CrossRef]
- Arthur, F.H. Efficacy of chlorfenapyr against adult Tribolium castaneum exposed on concrete: Effects of exposure interval, concentration and the presence of a food source after exposure. Insect Sci. 2009, 16, 157–163. [Google Scholar] [CrossRef]
- Athanassiou, C.G.; Arthur, H.; Throne, J.E. Efficacy of layer treatment with methoprene for control of Rhyzopertha dominica (Coleoptera: Bostrychidae) on wheat, rice and maize. Pest Manag. Sci. 2011, 67, 380–384. [Google Scholar] [CrossRef]
- Salama, H.S.; Abdel-Razek, A. Effect of different kinds of food on susceptibility of some stored products insects to Bacillus thuringiensis. J. Appl. Entomol. 1992, 113, 107–110. [Google Scholar] [CrossRef]
- Athanassiou, C.G.; Kavallieratos, N.G.; Arthur, F.H.; Throne, J.E. Efficacy of a combination of beta-cyfluthrin and imidacloprid and beta-cyfluthrin alone for control of stored-product insects on concrete. J. Econ. Entomol. 2013, 106, 1064–1070. [Google Scholar] [CrossRef]
- Isikber, A.A.; Athanassiou, C.G. The use of ozone gas for the control of insects and micro-organisms in stored products. J. Stored Prod. Res. 2015, 64, 139–145. [Google Scholar] [CrossRef]
- Isikber, A.A.; Oztekin, S. Comparison of susceptibility of two stored-product insects, Ephestia kuehniella Zeller and Tribolium confusum du Val to gaseous ozone. J. Stored Prod. Res. 2009, 45, 159–164. [Google Scholar] [CrossRef]
- Athanassiou, C.; Milonas, D.N.; Saitanis, C.J. Insecticidal effect of ozone against Rhyzopertha dominica (F.) (Coleoptera: Bostrichidae), Sitophilus oryzae Du Val (Coleoptera: Curculionidae) and Tribolium confusum Du Val (Coleoptera: Tenebrionidae): Influence of commodity. In Proceedings of the 8th International Conference on Controlled Atmosphere and Fumigation in Stored Products, Chengdu, China, 21–26 September 2008; Daolin, G., Ed.; Sichuan Publishing House of Science and Technology: Chengdu, China, 2008. [Google Scholar]
- Mendez, F.; Maier, D.E.; Mason, L.J.; Woloshuk, C.P. Penetration of ozone into columns of stored grains and effects on chemical composition and processing performance. J. Stored Prod. Res. 2003, 39, 33–44. [Google Scholar] [CrossRef]
- Navarro, S.; Amos, T.G.; Williams, P. The effect of oxygen and carbon dioxide gradients on the vertical dispersion of grain insects in wheat. J. Stored Prod. Res. 1981, 17, 101–107. [Google Scholar] [CrossRef]
- Navarro, S. Modified atmospheres for the control of stored-product insects and mites. In Insect Management for Food Storage and Processing; Heaps, J.W., Ed.; AACC International: St. Paul, MN, USA, 2006; pp. 105–146. [Google Scholar]
- Navarro, S. The use of modified and controlled atmospheres for the disinfestation of stored products. J. Pest Sci. 2012, 85, 301–322. [Google Scholar] [CrossRef]
- Williams, P.; Minett, W.; Navarro, S.; Amos, T.G. Sealing a farm silo for insect control by nitrogen swamping for fumigation. Aust. J. Exp. Anim. Husb. 1980, 20, 108–114. [Google Scholar] [CrossRef]
- White, N.D.G.; Jayas, D.S. Controlled atmosphere storage of grain. In Cereals, Fruits, Vegetables, Tea, and Spices; Chakraverty, A., Majumdar, A.S., Raghavan, G.S.W., Ramaswamy, H.S., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 2003; pp. 235–251. [Google Scholar]
- Toews, M.D.; Subramanyam, B. Contribution of contact toxicity and wheat condition to mortality of stored-product insects exposed to spinosad. Pest Manag. Sci. 2003, 59, 538–544. [Google Scholar] [CrossRef]
- Marinotti, O.; Nguyen, Q.K.; Calvo, E.; James, A.A.; Ribeiro, J.M.C. Microarray analysis of genes showing variable expression following a blood meal in Anopheles gambiae. Insect Mol. Biol. 2005, 14, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Vincent, C.; Weintraub, P.G.; Hallman, G.J. Insect management with physical methods in pre- and post-harvest situations. In Integrated Pest Management; Radcliffe, E.B., Hutchison, W.D., Cancelado, R.E., Eds.; Cambridge University Press: Cambridge, MA, USA, 2009; pp. 309–323. [Google Scholar]
- Subramanyam, B.; Mahroof, R.; Brijwani, M. Heat treatment of grain-processing facilities for insect management: A historical overview and recent advances. Stewart Postharvest Rev. 2011, 3, 1–11. [Google Scholar]
- Brijwani, M.; Subramanyam, B.; Flinn, P.W. Impact of varying levels of sanitation on mortality of Tribolium castaneum eggs and adults during heat treatment of a pilot flour mill. J. Econ. Entomol. 2012, 105, 703–708. [Google Scholar] [CrossRef]
- Opit, G.P.; Arthur, F.H.; Bonjour, E.L.; Jones, C.L.; Phillips, T.W. Efficacy of heat treatment for disinfestation of concrete grain silos. J. Econ. Entomol. 2011, 104, 1415–1422. [Google Scholar] [CrossRef] [PubMed]
- Fields, P.G. Control of insects post-harvest: Low temperature. In Physical Control Methods in Plant Protection; Vincent, C., Panneton, B., Fleurat-Lessard, F., Eds.; Springer: Berlin, Germany, 2001; pp. 95–107. [Google Scholar]
- Arthur, F.H.; Casada, M.E. Temperature stratification and insect pest populations in stored wheat with suction versus pressure aeration. Appl. Eng. Agric. 2016, 32, 849–860. [Google Scholar]
- Arthur, F.H.; Casada, M.E. Directional flow of summer aeration to manage insect pests in stored wheat. Appl. Eng. Agric. 2010, 26, 115–122. [Google Scholar] [CrossRef]
- Quiros, A.M. Evaluation of Ambient and Chilled Aeration Strategies to Maintain the Quality of Stored Grain in Tropical Climates and During Summer in Temperate Climates. Master’s Thesis, Kansas State University, Manhattan, KS, USA, 2017. [Google Scholar]
- Maier, D.E.; Adams, W.H.; Throne, J.E.; Mason, L.J. Temperature management of the maize weevil, Sitophilus zeamais Motsch. (Coleoptera: Curculionidae), in three locations in the United States. J. Stored Prod. Res. 1996, 32, 255–273. [Google Scholar] [CrossRef]
- Flinn, P.W.; Hagstrum, D.W.; Muir, W.E. Effects of time of aeration, bin size, and latitude on insect populations in stored wheat: A simulation study. J. Econ. Entomol. 1997, 90, 646–651. [Google Scholar] [CrossRef]
- Arthur, F.H. Structural pest management for stored product insects. In Recent Advances in Stored Product Protection; Athanassiou, C.G., Arthur, F.H., Eds.; Springer: Heidelberg, Germany, 2018; pp. 65–82. [Google Scholar]
- De Groote, H.; Kimenju, S.C.; Likhayo, P.; Kanampiu, F.; Tefera, T.; Hellin, J. Effectiveness of hermetic systems in controlling maize storage pests in Kenya. J. Stored Prod. Res. 2013, 53, 27–36. [Google Scholar] [CrossRef]
- Cline, L.D. Penetration of seven common flexible packaging materials by larvae and adults of eleven species of stored-product insects. J. Econ. Entomol. 1978, 71, 726–729. [Google Scholar] [CrossRef]
- Athanassiou, C.; Riudavets, J.; Kavallieratos, N. Preventing stored-product insect infestations in packaged-food products. Stewart Postharvest Rev. 2011, 3, 1–5. [Google Scholar]
- Scheff, D.S.; Subramanyam, B.; Arthur, F.H.; Dogan, H. Plodia interpunctella and Trogoderma variabile larval penetration and invasion of untreated and methoprene-treated foil packaging. J. Stored Prod. Res. 2018, 78, 74–82. [Google Scholar] [CrossRef]
- Mullen, M.A.; Mowery, S.V. Insect-resistant packaging. In Insect Management for Food Storage and Processing; Heaps, J.W., Ed.; AACC International: St. Paul, MN, USA, 2006; pp. 564–570. [Google Scholar]
- Arthur, F.H. Impact of food source on survival of red flour beetles and confused flour beetles (Coleoptera: Tenebrionidae) exposed to diatomaceous earth. J. Econ. Entomol. 2000, 93, 1347–1356. [Google Scholar] [CrossRef]
- Arthur, F.H.; Puterka, G.J. Evaluation of kaolinite-based particle films to control Tribolium species (Coleoptera: Tenebrionidae). J. Stored Prod. Res. 2002, 38, 341–348. [Google Scholar] [CrossRef]
- Scholler, M.; Prozell, S.; Al-Kirshi, A.G.; Reichmuth, C. Towards biological control as a major component of integrated pest management in stored product protection. J. Stored Prod. Res. 1997, 33, 81–97. [Google Scholar] [CrossRef]
- Scholler, M.; Prozell, S. Natural enemies to control stored-product pests in grain stores and retail stores. In Proceedings of the International Workshop “Implementation of Biocontrol in Practice in Temperate Regions—Present and Near Future”; DIAS Report No. 119. Research Centre Flakkebjerg: Flakkebjerg, Denmark, 2006; pp. 85–106. [Google Scholar]
- Zdarkova, E. Control of stored-food mites by non-chemical control. In Proceedings of the Proceedings International Forum, Strasbourg, France, 7–8 November; Council of Europe Publishers: Strasbourg, France, 1995; pp. 165–169. [Google Scholar]
- Sanon, A.; Ouedraogo, A.P.; Tricault, Y.; Credland, P.F.; Huignard, J. Biological Control of Bruchids in Cowpea Stores by Release of Dinarmus basalis (Hymenoptera: Pteromalidae) Adults. Environ. Entomol. 1998, 27, 717–725. [Google Scholar] [CrossRef]
- Cline, L.D.; Press, J.W.; Flaherty, B.R. Preventing the spread of the almond moth (Lepidoptera: Pyralidae) from infested food debris to adjacent uninfested packages, using the parasite Bracon hebetor (Hymenoptera: Braconidae). J. Econ. Entomol. 1984, 77, 331–333. [Google Scholar] [CrossRef]
- Grieshop, M.J.; Flinn, P.W.; Nechols, J.R. Biological control of indianmeal noth (Lepidoptera: Pyralidae) on finished stored products using egg and larval parasitoids. J. Econ. Entomol. 2006, 99, 1080–1084. [Google Scholar] [CrossRef] [PubMed]
- Godfray, H.C.J. Parasitoids: Behavioral and Evolutionary Ecology; Princeton University Press: Princeton, NJ, USA, 1994. [Google Scholar]
- Sieminska, E.; Ryne, C.; Löfstedt, C.; Anderbrant, O. Long-term pheromone-mediated mating disruption of the Mediterranean flour moth, Ephestia kuehniella, in a flourmill. Entomol. Exp. Appl. 2009, 131, 294–299. [Google Scholar] [CrossRef]
- Trematerra, P.; Athanassiou, C.; Stejskal, V.; Sciarretta, A.; Kavallieratos, N.; Palyvos, N. Large-scale mating disruption of Ephestia spp. and Plodia interpunctella in Czech Republic, Greece and Italy. J. Appl. Entomol. 2011, 135, 749–762. [Google Scholar] [CrossRef]
- Burks, C.S.; Mclaughlin, J.R.; Miller, J.R.; Brandl, D.G. Mating disruption for control of Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae) in dried beans. J. Stored Prod. Res. 2011, 47, 216–221. [Google Scholar] [CrossRef]
- Ryne, C.; Svensson, G.P.; Anderbrant, O.; Löfstedt, C.L. Evaluation of long-term mating disruption of Ephestia kuehniella and Plodia interpunctella (Lepidoptera: Pyralidae) in indoor storage facilities by pheromone traps and monitoring of relative aerial concentrations of pheromone. J. Econ. Entomol. 2007, 100, 1017–1025. [Google Scholar] [CrossRef]
- El-Sayed, A.M.; Suckling, D.M.; Byers, J.A.; Jang, E.B.; Wearing, C.H. Potential of “lure and kill” in long-term pest management and eradication of invasive species. J. Econ. Entomol. 2009, 102, 815–835. [Google Scholar] [CrossRef]
- El-Sayed, A.M.; Suckling, D.M.; Wearing, C.H.; Byers, J.A. Potential of mass trapping for long-lerm pest management and eradication of invasive species. J. Econ. Entomol. 2006, 99, 1550–1564. [Google Scholar] [CrossRef]
- Nansen, C.; Subramanyam, B.; Roesli, R. Characterizing spatial distribution of trap captures of beetles in retail pet stores using SADIE software. J. Stored Prod. Res. 2004, 40, 471–483. [Google Scholar] [CrossRef]
- Toews, M.D.; Campbell, J.F.; Arthur, F.H.; West, M. Monitoring Tribolium castaneum (Coleoptera: Tenebrionidae) in pilot-scale warehouses treated with residual applications of (S)-hydroprene and cyfluthrin. J. Econ. Entomol. 2005, 98, 1391–1398. [Google Scholar] [CrossRef]
- Campbell, J.F. Influence of landscape pattern in flour residue amount and distribution on Tribolium castaneum (Herbst) response to traps baited with pheromone and kairomone. J. Stored Prod. Res. 2013, 52, 112–117. [Google Scholar] [CrossRef]
- Romero, S.; Campbell, J.F.; Nechols, J.R.; With, K.A. Movement behavior in response to landscape structure: The role of functional grain. Landsc. Ecol. 2009, 24, 39–51. [Google Scholar] [CrossRef]
- Toews, M.D.; Arthur, F.H.; Campbell, J.F. Role of food and structural complexity on capture of Tribolium castaneum (Herbst) (Coleoptera: Tenebrionidae) in simulated warehouses. Environ. Entomol. 2005, 34, 164–169. [Google Scholar] [CrossRef]
- Toews, M.D.; Arthur, F.H.; Campbell, J.F. Monitoring Tribolium castaneum (Herbst) in pilot-scale warehouses treated with b-cyfluthrin: Are residual insecticides and trapping compatible? Bull. Entomol. Res. 2009, 99, 121–129. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Control | Tactic | Decreased Sanitation Impact on Efficacy | |||||
|---|---|---|---|---|---|---|---|
| Direct Effects | Indirect Effects | ||||||
| Effect Direction | n | Avg. Fold-Δ 1 | ± | SE | Effect Direction 2 | ||
| Chemical Control | − | 1240 | 8.5 | ± | 0.39 | − | |
| Fumigation | − | 67 | 15.7 | ± | 2.04 | − | |
| Grain Protectants | − | 100 | 1.3 | ± | 0.18 | − | |
| Aerosols | − | 389 | 2.8 | ± | 0.19 | − | |
| Residual Treatments | − | 656 | 10.5 | ± | 0.56 | − | |
| Modified Atmospheres | − | 28 | 1.1 | ± | 0.11 | − | |
| Physical/Cultural | − | 294 | 17.1 | ± | 1.76 | − | |
| Heat Treatment | − | 73 | 13.6 | ± | 4.35 | − | |
| Grain Chilling/Aeration | + | 132 | 31.5 | ± | 2.48 | − | |
| Passive Methods (packaging) | − | 64 | 1.6 | ± | 0.07 | − | |
| Active Methods (DE, inert dusts) | − | 25 | 3.5 | ± | 1.33 | − | |
| Biological Control | + | 113 | 6.8 | ± | 1.8 | −/+ | |
| Behaviorally-Based | 0/− | na 3 | na | 0/− | |||
| Mating Disruption | 0 | na | na | − | |||
| Stimuli-Based Monitoring | −/+ | na | na | −/+ | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morrison, W.R., III; Bruce, A.; Wilkins, R.V.; Albin, C.E.; Arthur, F.H. Sanitation Improves Stored Product Insect Pest Management. Insects 2019, 10, 77. https://doi.org/10.3390/insects10030077
Morrison WR III, Bruce A, Wilkins RV, Albin CE, Arthur FH. Sanitation Improves Stored Product Insect Pest Management. Insects. 2019; 10(3):77. https://doi.org/10.3390/insects10030077
Chicago/Turabian StyleMorrison, William R., III, Alexander Bruce, Rachel V. Wilkins, Chloe E. Albin, and Frank H. Arthur. 2019. "Sanitation Improves Stored Product Insect Pest Management" Insects 10, no. 3: 77. https://doi.org/10.3390/insects10030077