Morphology and Morphometry of the Midgut in the Stingless Bee Friesella schrottkyi (Hymenoptera: Apidae)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
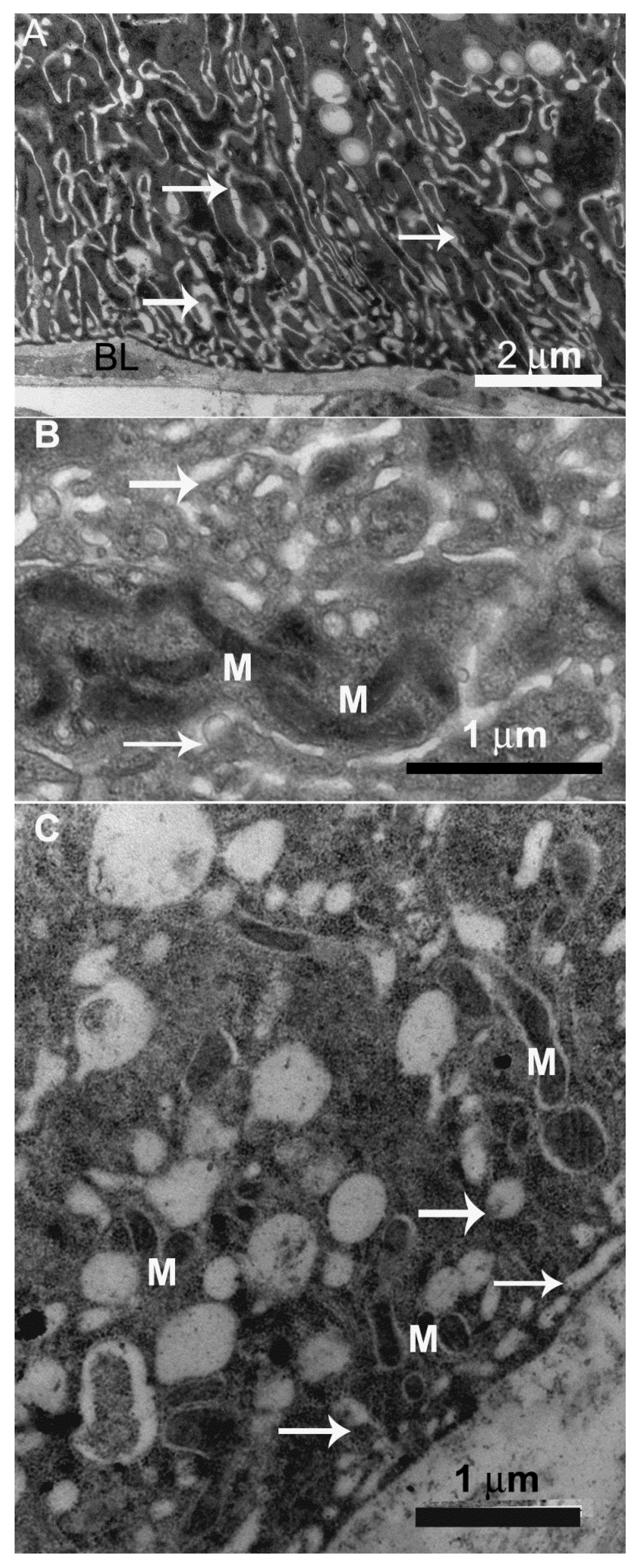{kind=link}
{kind=link}
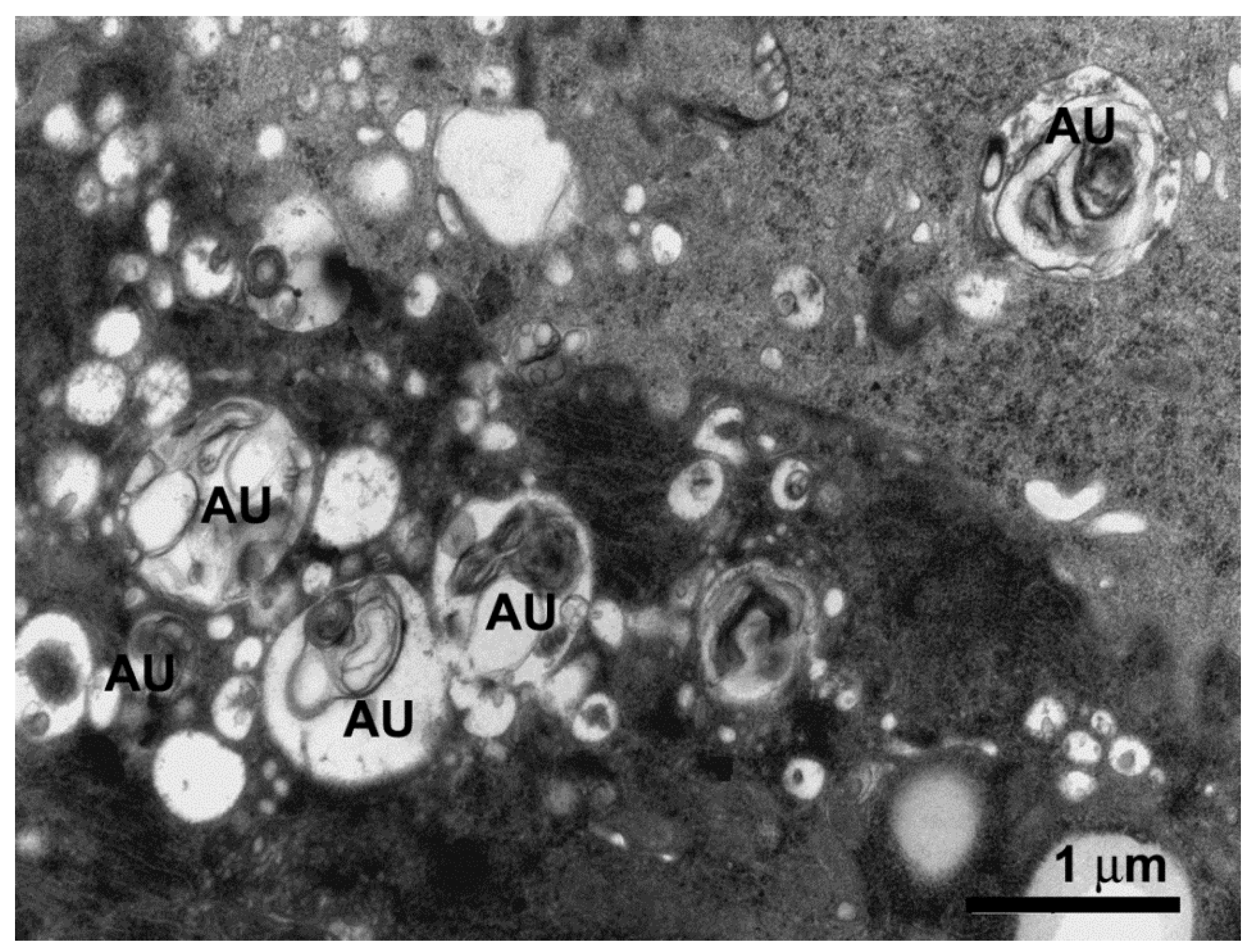{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Bees
2.2. Histology and Histochemistry
2.3. Immunofluorescence
2.4. Transmission Electron Microscope
2.5. Morphometry
2.6. Statistics
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schwarz, H.F. The stingless bees of the Western Hemisphere. Bull. Am. Mus. Nat. Hist. 1948, 90, 1–546. [Google Scholar]
- Vit, P.; Pedro, S.R.M.; Roubik, D.W. Pot-Honey. A Legacy of Stingless Bees; Springer: New York, NY, USA, 2013. [Google Scholar]
- Boti, J.B. Comportamento de oviposição apresentado por Friesella schrottkyi Friese, 1900 (Hymenoptera, Apidae: Meliponinae). Natureza 2017, 15, 27–31. [Google Scholar]
- Oliveira, F.P.M.; Absy, M.L.; Miranda, I.S. Recurso polínico coletado por abelhas sem ferrão (Apidae, Meliponinae) em um fragmento de floresta na região de Manaus–Amazonas. Acta Amazon. 2009, 39, 505–518. [Google Scholar] [CrossRef]
- Catae, A.F.; Roat, T.C.; Oliveira, R.A.; Ferreira Nocelli, R.C.; Malaspina, O. Cytotoxic effects of thiamethoxam in the midgut and Malpighian tubules of Africanized Apis mellifera (Hymenoptera: Apidae). Microsc. Res. Tech. 2014, 77, 274–281. [Google Scholar] [CrossRef]
- Catae, A.F.; Roat, T.C.; Pratavieira, M.; da Silva Menegasso, A.R.; Palma, M.S.; Malaspina, O. Exposure to a sublethal concentration of imidacloprid and the side effects on target and nontarget organs of Apis mellifera (Hymenoptera, Apidae). Ecotoxicology 2018, 27, 109–121. [Google Scholar] [CrossRef]
- Gregorc, A.; Ellis, J.D. Cell death localization in situ in laboratory reared honey bee (Apis mellifera L.) larvae treated with pesticides. Pestic. Biochem. Physiol. 2011, 99, 200–207. [Google Scholar] [CrossRef]
- Santos, H.P.; Gutierrez, Y.; Oliveira, E.E.; Serrão, J.E. Sublethal dose of deltamethrin damage the midgut cells of the mayfly Callibaetis radiatus (Ephemeroptera: Baetidae). Environ. Sci. Pollut. Res. 2018, 25, 1418–1427. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.A.P.; Martins, G.F.; Oliveira, E.E.; Guedes, R.N.C. Agrochemical-induced stress in stingless bees: Peculiarities, underlying basis, and challenges. J. Comp. Physiol. A 2016, 202, 733–747. [Google Scholar] [CrossRef] [PubMed]
- Fialho, M.C.Q.; Terra, W.R.; Moreira, N.R.; Zanuncio, J.C.; Serrão, J.E. Ultrastructure and immunolocalization of digesive enzymes in the midgut of Podisus nigrispinus (Heteroptera: Pentatomidae). Arthropod Struct. Dev. 2013, 42, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Serrão, J.E.; Cruz-Landim, C. Ultrastructure of digestive cells in stingless bees of various ages (Hymenoptera, Apidae,Meliponinae). Cytobios 1996, 88, 161–171. [Google Scholar]
- Cruz, L.C.; Araújo, V.A.; Dolder, H.; Araújo, A.P.A.; Serrão, J.E.; Neves, C.A. Morphometry of the midgut of Melipona quadrifasciata anthidioides (Lepeletier) (Hymenoptera: Apidae) during metamorphosis. Neotrop. Entomol. 2011, 40, 677–681. [Google Scholar] [CrossRef] [PubMed]
- Neves, C.A.; Bhering, L.L.; Serrão, J.E.; Gitirana, L.B. FMRFamide like midgut endocrine cells during the metamorphosis in Melipona quadrifasciata anthidioides (Hymenoptera. Apidae). Micron 2002, 33, 453–460. [Google Scholar] [CrossRef]
- Cruz, L.C.; Araújo, V.A.; Fialho, M.C.Q.; Serrão, J.E.; Neves, C.A. Proliferation and cell death in the midgut of the stingless bee Melipona quadrifasciata anthidioides (Apidae: Meliponini) during metamorphosis. Apidologie 2013, 44, 458–466. [Google Scholar] [CrossRef]
- Fernandes, K.M.; Martins, G.F.; Serrão, J.E.; Neves, C.A. Digestive and regenerative cells in the midgut of haploid and diploid males of the stingless bee Melipona quadrifasciata anthidioides (Hymenoptera: Apidae). Zoologia 2012, 29, 488–492. [Google Scholar] [CrossRef]
- Martins, G.F.; Neves, C.A.; Campos, L.A.; Serrão, J.E. The regenerative cells during the metamorphosis in the midgut of bees. Micron 2006, 37, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, D.R.; Gilliam, M. Age-related changes in midgut ultrastructure and trypsin activity in the honey bee, Apis mellifera. Apidologie 1989, 20, 287–303. [Google Scholar] [CrossRef]
- Jimenez, D.R.; Gilliam, M. Ultrastructure of the ventriculus of the honeybee, Apis mellifera (L.): Cytochemical localization of acid phosphatase, alkaline phosphatase, and nonspecific esterase. Cell Tissue Res. 1990, 261, 431–443. [Google Scholar] [CrossRef]
- Serrão, J.E.; Cruz-Landim, C. Ultrastructure of midgut endocrine cells in workers of stingless bees (Hymenoptera, Apidae, Meliponinae). Iheringia 1996, 81, 151–156. [Google Scholar]
- Serrão, J.E.; Cruz-Landim, C. Ultrastructure of the midgut epithelium of Meliponinae larvae with different developmental stages and diets. J. Apic. Res. 2000, 39, 9–17. [Google Scholar] [CrossRef]
- Lo, N.; Gloag, R.S.; Anderson, D.L.; Oldroyd, B.P. A molecular phylogeny of the genus Apis suggests that giant honey bee of the Phillipines, A. breviligula Maa, and plains honey bee of southern India, A. indica Fabricius, are valid species. Syst. Entomol. 2010, 35, 226–233. [Google Scholar] [CrossRef]
- Camargo, J.M.F.; Pedro, S.R.M. Systematics, phylogeny and biogeography of the Meliponinae (Hymenoptera, Apidae) A mini-review. Apidologie 1992, 23, 509–523. [Google Scholar] [CrossRef]
- Stefanini, M.; De Martino, C.E.S.A.R.E.; Zamboni, L. Fixation of ejaculated spermatozoa for electron microscopy. Nature 1967, 216, 173–174. [Google Scholar] [CrossRef] [PubMed]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques, 5th ed.; Churchill Livingstone Elsevier: Philadelphia, PA, USA, 2002; p. 19. [Google Scholar]
- Gajger, I.; Kozaric, Z.; Berta, D.; Nejedli, S.; Petrinec, Z. Effect of the herbal preparation Nozevict on the midgut structure of honeybees (Apis melıfera) infected with Nosema sp. Spores. Vet. Med. 2011, 56, 344–351. [Google Scholar] [CrossRef]
- Gonçalves, W.G.; Fernandes, K.M.; Gonçalves, M.P.; Neves, C.A.; Martins, G.F.; Zanuncio, J.C.; Serrão, J.E. The midgut of the parasitoid Campoletis flavicincta (Hymenoptera: Ichneumonidae). Fla. Entomol. 2013, 96, 1016–1022. [Google Scholar] [CrossRef]
- Gonçalves, W.G.; Fernandes, K.M.; Barcellos, M.S.; Silva, F.P.; Magalhães-Junior, M.J.; Zanuncio, J.C.; Martins, G.F.; Serrão, J.E. Ultrastructure and immunofluorescence of the midgut of Bombus morio (Hymenoptera: Apidae: Bombini). C. R. Biol. 2014, 337, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Pipan, N.; Rakovec, V. Cell death in the midgut epithelium of the worker honey bee (Apis mellifera carnica) during metamorphosis. Zoomorphologie 1980, 94, 217–224. [Google Scholar] [CrossRef]
- Bution, M.L.; Caetano, F.H.; Fabio, B.B.; Giovana, A.T.G.; Zara, F.J. Histology and histochemistry of the ventriculus Dolichoderus (=Monacis) bispinosus (Olivier, 1792) (Hymenoptera: Formicidae). Micron 2006, 37, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Marques-Silva, S.; Serrão, J.E.; Mezêncio, J.M.S. Peritrophic membrane protein in the larval stingless bee Melipona quadrifasciata anthidioides: Immunolocalization of secretory sites. Acta Histochem. 2005, 107, 23–30. [Google Scholar] [CrossRef]
- Teixeira, A.D.; Marques-Araujo, S.; Zanuncio, J.C.; Serrão, J.E. Peritrophic membrane origin in adult bees (Hymenoptera): Immunolocalization. Micron 2015, 68, 91–97. [Google Scholar] [CrossRef]
- O’Donnell, M.J.; Maddrell, S.H.; Skaer, H.B.; Harrison, J.B. Elaborations of the basal surfaces of the cells of the Malpighian tubules of an insect. Tissue Cell 1985, 17, 865–881. [Google Scholar] [CrossRef]
- Serrão, J.E.; Ronnau, M.; Neves, C.A.; Campos, L.A.O.; Zanuncio, J.C. Ultrastructure of anterior midgut region of corbiculate bees (Hymenoptera:Apidae). Ann. Entomol. Soc. Am. 2008, 101, 915–921. [Google Scholar] [CrossRef]
- Grogan, D.; Hunt, J.H. Chymotrypsin-Like activity in the honeybee midgut: Patterns in the three-year study. J. Apic. Res. 1984, 23, 61–63. [Google Scholar] [CrossRef]
- Chan, D.C. Mitochondria: Dynamic organelles in disease, aging, and development. Cell 2006, 125, 1241–1252. [Google Scholar] [CrossRef] [PubMed]
- Kleizen, B.; Braakman, I. Protein folding and quality control in the endoplasmic reticulum. Curr. Opin. Cell Biol. 2004, 16, 343–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cnop, M.; Foufelle, F.; Velloso, L.A. Endoplasmic reticulum stress, obesity and diabetes. Trends Mol. Biol. 2012, 18, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Serrão, J.E.; Cruz-Landim, C. The striated border of digestive cells in adult stingles bees (Hymenoptera, Apidae, Meliponinae). Cytobios 1995, 83, 229–235. [Google Scholar]
- Cruz-Landim, C. Abelhas: Morfologia e Função de Sistemas; Editora UNESP: Rio Claro, Colombia, 2009. [Google Scholar]
- Fialho, M.C.Q.; Zanuncio, J.C.; Neves, C.A.; Ramalho, F.S.; Serrão, J.E. Ultrastructure of the digestive cells in the midgut of the predator Brontocoris tabidus (Heteroptera: Pentatomidae) after different feeding periods on prey and plants. Ann. Entomol. Soc. Am. 2009, 102, 119–127. [Google Scholar] [CrossRef]
- Moure, J.S.; Sakagami, S.F. As mamangabas sociais do Brasil (Bombus Latr.) (Hymenoptera, Apoidea). Stud. Entomol. 1962, 51, 65–194. [Google Scholar]
- Martins, G.F.; Serrão, J.E. A comparative study of the ovaries in some Brazilian bees (Hymenoptera; Apoidea). Pap. Avulsos Zool. 2004, 44, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Terra, W.R. Physiology and biochemistry of insect digestion. An evolutionary perspective. Braz. J. Med. Biol. Res. 1988, 21, 675–734. [Google Scholar]
- Schumaker, T.T.S.; Cristofoletti, P.T.; Terra, W.R. Properties and compartmentalization of digestive carbohydrases and proteases in Scaptotrigona bipunctata (Apidae, Meliponinae) larvae. Apidologie 1993, 24, 3–17. [Google Scholar] [CrossRef]
- Billinglsley, P.F. The midgut ultrastructure of hematophagous insects. Annu. Rev. Entomol. 1990, 35, 219–248. [Google Scholar] [CrossRef]
- Douglas, A.E. Phloem-sap feeding by animals: Problems and solutions. J. Exp. Bot. 2006, 57, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Serrão, J.E.; Fialho, M.C.Q.; Azevedo, D.O.; Zanuncio, J.C. Aquaporins in the honeybee crop—A new function for an old organ. Protoplasma 2014, 251, 1441–1447. [Google Scholar] [CrossRef] [PubMed]
- Neves, C.A.; Gitirana, L.B.; Serrao, J.E. Ultrastructural study of the metamorphosis in the midgut of Melipona quadrifasciata athidioides (Apidae: Meliponini) worker. Sociobiology 2003, 41, 443–459. [Google Scholar]
- Souza, D.L.L.; Zanuncio, J.C.; Serrão, J.E. FMRFamide-cells in the midgut of Scaptotrigona xanthotricha (Apidae: Meliponini) of different ages and fed different diets. J. Apic. Res. 2016, 55, 428–432. [Google Scholar] [CrossRef]
- Santos, D.E.; Zanuncio, J.C.; de Oliveira, A.A.G.; Serrão, J.E. Endocrine cells in the midgut of bees (Hymenoptera: Apoidea) with different levels of sociability. J. Apic. Res. 2015, 54, 394–398. [Google Scholar] [CrossRef]
- Lajeunesse, D.R.; Johnson, B.; Presnell, J.S.; Catignas, K.K.; Zapotoczny, G. Peristalsis in the junction region of the Drosophila larval midgut is modulated by DH31 expressing enteroendocrine cells. BMC Physiol. 2010, 10, 14. [Google Scholar] [CrossRef] [PubMed]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, A.H.; Gonçalves, W.G.; Fernandes, K.M.; Barcellos, M.S.; Sampaio, W.M.S.; Lopes, M.P.; Martins, G.F.; Serrão, J.E. Morphology and Morphometry of the Midgut in the Stingless Bee Friesella schrottkyi (Hymenoptera: Apidae). Insects 2019, 10, 73. https://doi.org/10.3390/insects10030073
Oliveira AH, Gonçalves WG, Fernandes KM, Barcellos MS, Sampaio WMS, Lopes MP, Martins GF, Serrão JE. Morphology and Morphometry of the Midgut in the Stingless Bee Friesella schrottkyi (Hymenoptera: Apidae). Insects. 2019; 10(3):73. https://doi.org/10.3390/insects10030073
Chicago/Turabian StyleOliveira, André H., Wagner G. Gonçalves, Kenner M. Fernandes, Marcelo S. Barcellos, Wagner M. S. Sampaio, Marcos P. Lopes, Gustavo F. Martins, and José E. Serrão. 2019. "Morphology and Morphometry of the Midgut in the Stingless Bee Friesella schrottkyi (Hymenoptera: Apidae)" Insects 10, no. 3: 73. https://doi.org/10.3390/insects10030073