What Is the Carcass-Usage Mode of the Collembola? A Case Study of Entomobrya proxima in the Laboratory
by
 ,
,
Lichao Feng
1,2 ,
,
Liang Chang
3,
Shaoqing Zhang
3,
Xinyu Zhu
4,
Sina Adl
5,* and
Donghui Wu
1,3,6,7,* 1
College of Earth Sciences, Jilin University, Changchun 130012, China
2
Department of Plant Sciences, Jilin Agricultural Science and Technology College, Jilin 132101, China
3
Northeast Institute of Geography and Agroecology, Chinese Academy of Sciences, Changchun 130102, China
4
College of Resource and Environmental Sciences, Hebei Normal University, Shijiazhuang 050024, China
5
Department of Soil Science, College of Agricultural and Bioresources, University of Saskatchewan, 51 Campus Drive, Saskatoon, SK S7N5A8, Canada
6
Jilin Provincial Key Laboratory of Animal Resource Conservation and Utilization, Northeast Normal University, Changchun 130117, China
7
Key Laboratory of Vegetation Ecology, Ministry of Education, Northeast Normal University, Changchun 130024, China
*
Authors to whom correspondence should be addressed.
Insects 2019, 10(3), 67; https://doi.org/10.3390/insects10030067
Submission received: 12 February 2019
/
Revised: 1 March 2019
/
Accepted: 4 March 2019
/
Published: 7 March 2019
Abstract
:Collembola display a variety of feeding habits, and prey on many types of food at different trophic levels in the soil. In most cases, their feeding selections are widely varied. In the interest of the food preferences of E. proxima, we attempted to confirm how the Collembola utilize food when feeding on carrion (unusual sources). Four different soil animals (with different stable isotope values and increasing trophic levels) were used to examine whether collembolans can use dead insects as a food resource in specific manners, depending on food preference. Our results demonstrated that the food preference of a collembolan changed significantly after feeding on insects with different feeding habits for 60 days. We found that stable isotope values (δ13C) of Entomobrya proxima approached those of the food sources. A large proportion of the diet (more than 50%) should directly consist of insect body parts, with the remainder consisting of indirectly used, mixed microorganisms naturally growing on animal food, such as fungi (Rhizopus sp., Alternaria sp., Penicillium sp., and Aspergillus sp.) and bacteria (Bacillus sp1. and Bacillus sp2.). Based on this research, the food preference of collembolans is more focused on carcasses (dead insect bodies) than microorganisms during the animal-food decomposition process.
1. Introduction
Collembola are among the most important decomposers [1,2,3,4], promoting the conversion of organic matter into inorganic matter in order to obtain high-quality carbon and nitrogen resources for growth, development and reproduction [5]. The feeding habits of Collembola are highly multifarious and include feeding on plant litter, fungi, bacteria, algae, live plant tissues, some plant pathogens, Protozoa, Nematoda, Rotatoria, Enchytraeidae and, carcasses [6,7,8,9,10,11]; their feeding habits are limited by food availability, abundance, quality and composition to some extent [12,13]. Microorganisms choose invertebrate food sources based on accessibility, digestibility and utilization [14,15]. In nature, dead organic matter (plant and animal residues) can be decomposed by microorganisms (fungi and bacteria) and invertebrates (for example dung beetles, march flies, maggots, mites, collembolans, and earthworms) in the soil [16,17,18], but the decomposition process of most parts is first carried out by microorganisms [19]. Thus, collembolans may feed on the food itself, the microorganisms, or both. As fungal feeders, collembolans are in contact with a large variety of fungi in their environment and experience a seasonal variation in the fungi [20]. The features of the temporal and spatial changes among fungal communities are closely connected with the high flexibility of epigeic Collembola in terms of conditions [16,21].
Thus, food preference is closely related to the range of vertical soil fauna activity [22], and this preference may be attributed to the restriction of feeding preference on utilization [23,24]. In general, detritivores obtain more nutrients or energy when consuming food at higher trophic levels [15], and they must rely on foods of different composition within an identical and environmentally homogeneous space [25]. Variability in the diet is an indication of complex trophic interactions, and members of Collembola benefit by feeding on polytrophic and abundant foods. Food resource availability is dynamic within a habitat, and collembolans thus encounter different foods in time and space. Because of such changes in food resources, the feeding habits and behaviors of collembolans vary with the composition, availability and palatability of a diet [13,26,27]. These habits and behaviors can be seen in fungal diet selection with fungal succession during the litter decaying process [28]. Overall, stable isotopes (δ13C) were used to detect changes in the diets of Collembola, because δ13C values differ largely among animals and life stages [29,30], and the δ13C and δ15N values of soil invertebrates vary over a wide range [31,32]. Moreover, each animal and stage has a unique stable isotopic ratio [33,34,35]. Therefore, δ13C is used to determine food selected in the process of food decomposition [36].
Collembola mouthparts, morphology and ecology. The mouth contain the labrum, a pair of mandibulae, a pair of maxillae, the hypopharynx and the bipartite labium. The frontal labrum, the ventral labium and two lateral oral folds enclose the other mouthparts in the buccal cavity [37]. Thus, Collembola can feed detritus and microorganisms [12]. Collembola inhabit soil, leaf litter, moss, the area under stones, caves, ant nests, etc., which are areas with wet and damp surroundings. They are the primary soil animals, and in forest soils, they can reach densities of 200 to 1800 ind. per dm3. In general, densities are only lower than the soil mites population [37,38]. Their living temperature is −4 °C to 28 °C, and relative humidity is 93% to 100% [39].
In this study, we used insect food to examine the food preference of Entomobrya proxima (an epigeic collembolan) in the spoilage process. We aimed to test whether the collembolan food-use mode is more complex and includes different utilization modes (direct intake or indirect use by microorganism).
2. Materials and Methods
2.1. Insect Food and Collembolan Activity
We selected the following soil invertebrates (insects) as artificial food with different feeding habits and trophic levels from habitats identical to those of E. proxima as food resources: Adults of an omnivorous ant (Formica yessensis Forel), a phytophagous scarab (Popillia quadriguttata Fabr), a saprophagous dung beetle (Onthophagus viduus Harold), and a carnivorous carabid (Anoplogenius cyanescens (Hope)) in a coniferous and broad-leaved mixed forest, located at 43.88° N, 126.57° E, Jilin, China. These insects were starved for 24 h [35] and killed by physical methods; the abdomens were removed and dried at 60 °C for more than 48 h until no further change in weight was observed. The residues were ground using a sterile mortar, and 30 mg was added to the substrate. Because of the presence of saprophytic microorganisms in the air or on the insect themselves, we wanted to check which saprophytic microorganisms would grow on the four food resources, and whether these microorganisms were foraged by the collembolan. First, we used PDA (potato dextrose agar) (Merck Co., Ltd., Kenilworth, NJ, USA) for fungi and NA (nutrient agar) (Merck Co., Ltd.) for bacteria in the air to identify which would grow on the animal food in our tests. Second, after some days of feeding, we inoculated the four insect powders that had been placed on the gypsum-activated carbon substrate onto PDA and NA to check for microorganisms on the insects. Fungi and bacteria were detected by using morphological and physicochemical characteristics and nucleotide sequences [40,41,42,43].
E. proxima was obtained using a suction trap. For rearing E. proxima, food was supplied as dried yeast (Saccharomyces cerevisiae; Angel Yeast Co., Ltd., Yi Chang, China) on a gypsum-activated carbon substrate in a plastic preservation box with a volume of 400 mL (plaster of Paris: activated charcoal at 9:1; Sinopharm Chemical Reagent Co., Ltd., Shanghai, China). Deionized distilled water was applied and maintained at temperatures under 17–19 °C.
2.2. Feeding Strategies (E. proxima)
After seven days of starvation, 20 second-instar E. proxima in the second generation, with identical body sizes and levels of activity (10 days old), were inoculated onto each culture substrate [44] and cultured at 18 ± 1 °C and 85 ± 1% relative humidity with a light:dark cycle of 16:8 h (400–800 Lux) [45,46] while allowing for reproduction. Each day, deionized distilled water was added to maintain constant humidity.
2.3. Stable Isotope Analysis
Isotopes were used to detect the E. proxima diet after 60 days. Although collembolans do not always consume a uniform amount of food in different instars or stages, the diet can be determined by the isotope composition of the consumer [47]. We used a stable isotope (δ13C) analysis to determine food preference and to verify food use. Based on the life cycle and development time, the life expectancy of E. proxima was more than 30 days under normal conditions (22–24 °C). The stable isotope (δ13C) analysis was conducted under a condition of sufficient food. We ground the tissues of the previously dried four soil fauna species to powders, which were then placed in tin capsules for elemental and stable isotope analyses. Fungi were also tested [48]. Samples were analyzed using a continuous-flow isotope ratio mass spectrometer (ThermoScientific Delta V Advantage; Waltham, Middlesex County, Massachusetts, USA). The δ-value is expressed as δM(‰) = [(Rsample/Rstandard) − 1] × 1000‰, where M is the element, 13C, Rsample is the13C/12C ratio of the measured samples, and Rstandard represents the ratio of the heavy to the light isotope in an international standard [49].
2.4. Statistical Analysis
According to the mass conservation of stable isotopes, the number of possible food source combinations and acceptance solutions of food-source proportional contributions to a mixture based on stable isotope analyses were calculated by the IsoSource mixing model (https://isosource.software.informer.com/). This model provided the distribution of the source proportions that are consistent with isotopic mass balance [50,51]. The contribution ratio of different food sources after feeding by E. proxima was analyzed using a nonparametric Kruskal-Wallis test with a median test. The significance level of the statistical tests was p = 0.05. The statistical analyses were performed using R statistical software (‘ggpubr’, ‘digest’ packages) [52].
3. Results
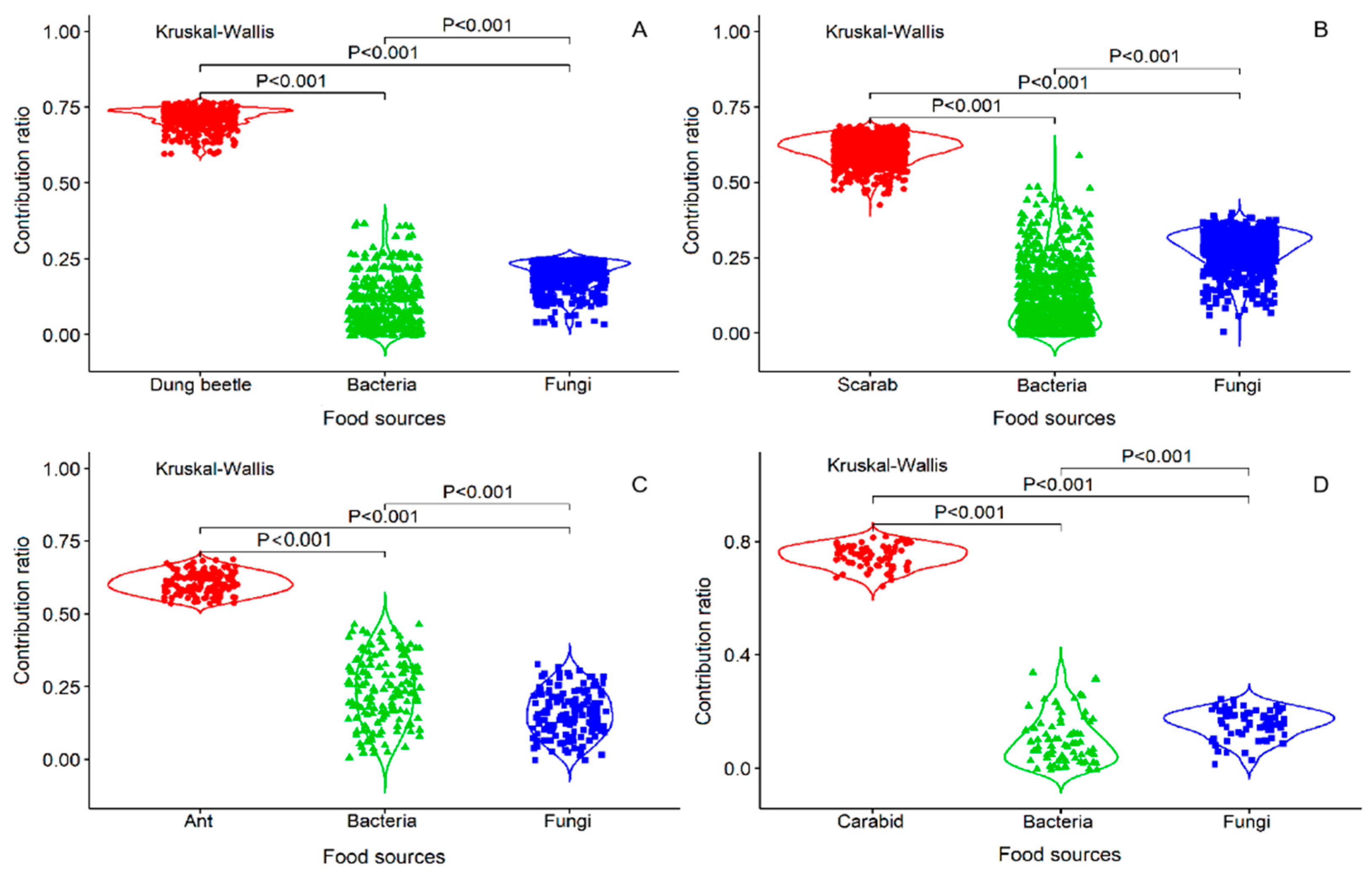
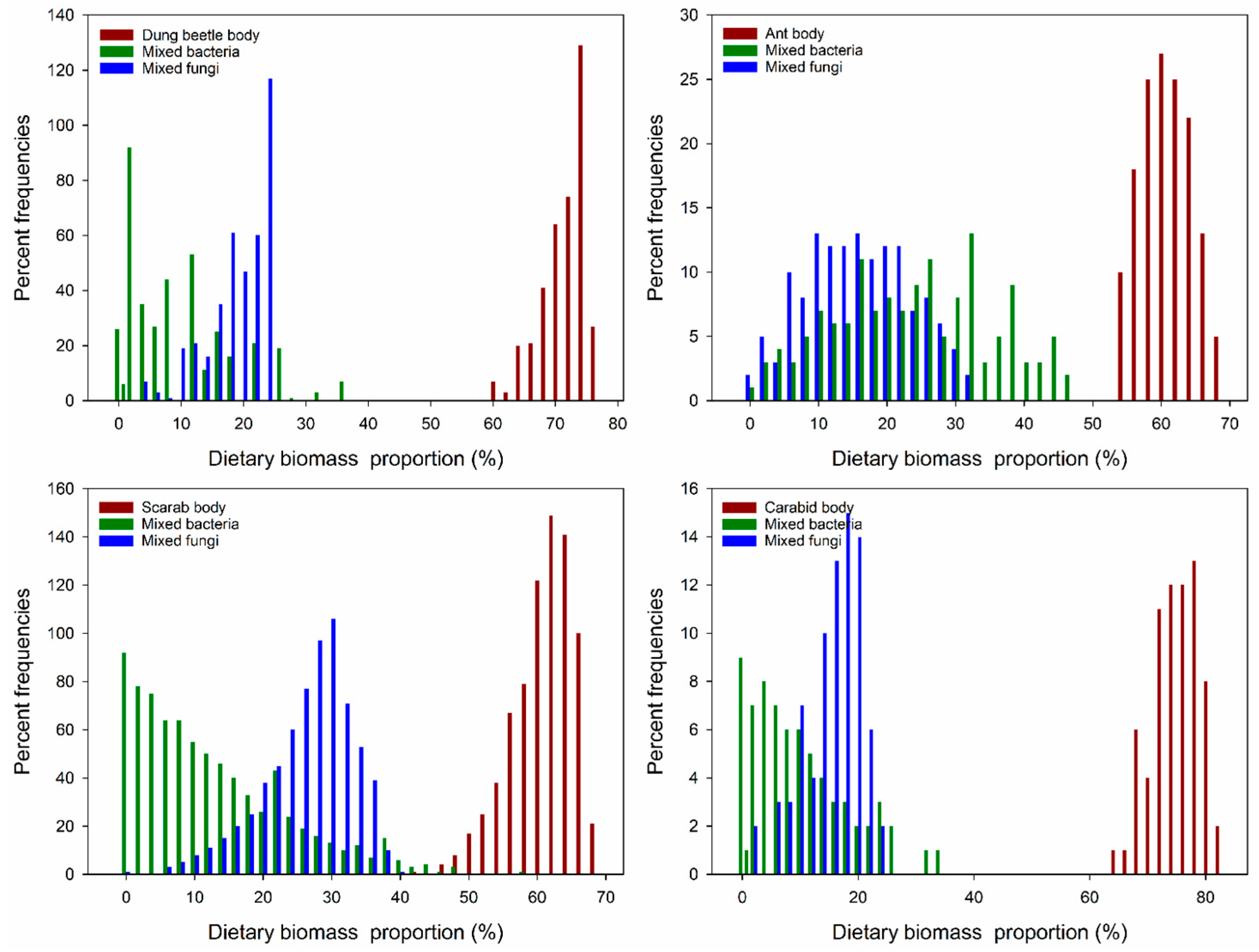
Four fungi (Rhizopus sp., Alternaria sp., Penicillium sp., and Aspergillus sp.) and two bacteria (Bacillus sp1. and Bacillus sp2.) grew on the insect body pieces during the decomposition process. The results of the analysis are listed in Table 1 as the data for calculating the acceptable solutions of food source contributions. By analyzing the food sources and Collembola after feeding, carcasses (insect body: Dung beetle, ant, scarab, and carabid) accounted for the large parts of food and ranged from 60–76%, 54–68%, 42–68%, and 64–82%, respectively. The isotopic values verified that the ratio of the four insect foods was higher than the ratios of the fungi and bacteria, especially for the carabids (p < 0.001, Figure 1A–D). In addition, by comparing the Collembola consumption ratios of the fungi and bacteria, more fungi than bacteria were consumed (p < 0.001, Figure 1A,B,D), except for the fungi growing on ants (p < 0.001, Figure 1C). These results indicate that E. proxima prefers to consume an insect body over microorganisms growing on an insect. The order of food selection was insect body, followed by fungi and then bacteria. For the mean percent frequencies of dietary biomass proportions, most of the carcasses accounted for a higher proportion than the microorganism carcasses, except for carabids. For the microorganism diet, Collembola consumed fungi more frequently (Figure 2).
4. Discussion
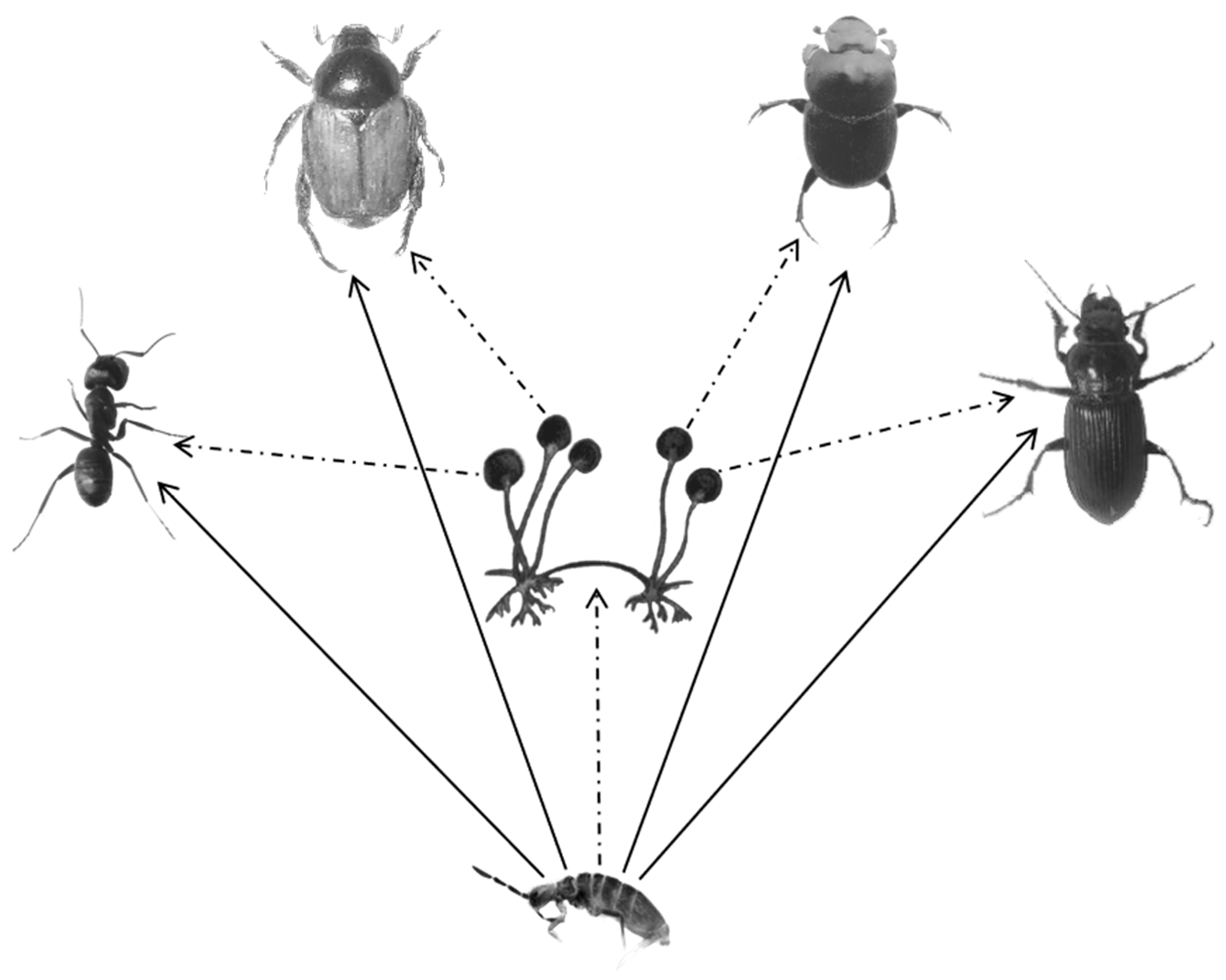
Based on our results, all food types resulted in different stable isotope ratios in the collembolan. Insect carrion was used more frequently than the microorganism by E. proxima during insect carcass decomposition. Direct exploitation (feeding carrion) would be more popular than indirect exploitation (Figure 3). These changes were determined by the eating patterns, and were also dependent on the food resource. For decomposition, microorganisms are typically the pioneer decomposers that decompose dead animals into carrion [19]. Carbon source assimilation explains the stable isotope value changes in the decomposer [53]. For decomposers, different assimilation intensities lead to different utilization and isotope values (δ13C) [54]. In natural habitats, it is difficult or impossible to verify the food source combination, because many food sources are used by the feeders, and the isotope signatures could be the same after being assimilated [51,54]. However, in our test, the aggregated food resources were determined, the dietary contributions were clearly displayed, and the IsoSource model can be applied to detect Collembola feeding habits. As fungal feeders, Collembola choose fungi as food [20,55], which is attributed to the nutrient content available in the habitat [12,56,57]. Although only 9% of animal residues have been found in the gut of most studied Collembola species and the other large parts were plant residues and fungi [58], after pre-decomposition by released microorganism enzymes, the carcasses would be more digestible for the decomposers [59,60].
5. Conclusions
We concluded that as decomposers, Collembola food utilization might be performed in many ways depending on the food sources available. In our study, for carcasses, Collembola were more focused on carrion (dead insect bodies) than microorganisms during the decomposition process.
Author Contributions
Conceptualization, L.F. and D.W.; Methodology, L.F. and L.C.; Software, L.F.; Validation, L.F. and S.A.; Formal Analysis, X.Z. and S.Z.; Investigation, S.Z.; Resources, L.C. and L.F.; Data Curation, L.F.; Writing—Original Draft Preparation, L.F.; Writing—Review & Editing, S.A.; Visualization, D.W. and S.A.; Supervision, D.W. and S.A.; Project Administration, D.W.; Funding Acquisition, D.W.
Funding
This research was supported by the Key Program of National Natural Science of China (41430857), Science and Technology Basic Work of the Ministry of Science and Technology of China (2014FY110600-5), National Natural Sciences Foundation of China (No. 41671259, 31861133006, 41501263, 31200331), the Program of Introducing Talents of Discipline to Universities (No.B16011).
Acknowledgments
We thank the analysis center of IGA-CAS for their help. The authors also wish to thank the editors and anonymous reviewers for their useful comments and suggestions on this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bardgett, R.D.; Whittaker, J.B.; Frankland, J.C. The effect of Collembolan grazing on fungal activity in differently managed upland pastures: A microcosm study. Biol. Fertil. Soils 1993, 16, 255–262. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Chan, K.F. Experimental evidence that soil fauna enhance nutrient mineralization and plant nutrient uptake in montane grassland ecosystems. Soil Biol. Biochem. 1999, 31, 1007–1014. [Google Scholar] [CrossRef]
- Osler, G.H.R.; Sommerkorn, M. Toward a complete soil C and N cycle: Incorporating the soil fauna. Ecology 2007, 88, 1611–1621. [Google Scholar] [CrossRef] [PubMed]
- Schutz, K.; Bonkowski, M.; Scheu, S. Effects of Collembola and fertilizers on plant performance (Triticum aestivum) and aphid reproduction (Rhopalosiphum padi). Basic Appl. Ecol. 2008, 9, 182–188. [Google Scholar] [CrossRef]
- Chamberlain, P.M.; Bull, I.D.; Black, H.I.J.; Ineson, P.; Evershed, R.P. Collembolan trophic preferences determined using fatty acid distributions and compound-specific stable carbon isotope values. Soil Biol. Biochem. 2006, 38, 1275–1281. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Whittaker, J.B.; Frankland, J.C. The diet and food preferences of Onychiurus procampatus (collembola) from upland grassland soils. Biol. Fertil. Soils 1993, 16, 296–298. [Google Scholar] [CrossRef]
- Bokhorst, S.; Ronfort, C.; Huiskes, A.; Convey, P.; Aerts, R. Food choice of antarctic soil arthropods clarified by stable isotope signatures. Polar Biol. 2007, 30, 983–990. [Google Scholar] [CrossRef]
- Buse, T.; Filser, J. Mucilaginous seeds and algal diets attract soil Collembola in preference tests. Eur. J. Soil Biol. 2014, 65, 1–6. [Google Scholar] [CrossRef]
- Rusek, J. Biodiversity of Collembola and their functional role in the ecosystem. Biodivers. Conserv. 1998, 7, 1207–1219. [Google Scholar] [CrossRef]
- Seastedt, T.R.; Mameli, L.; Gridley, K. Arthropod use of invertebrate carrion. Am. Midl. Nat. 1981, 105, 124–129. [Google Scholar] [CrossRef]
- Maynard, E.A. A Monograph of the Collembola or Springtail Insects of New York State. Part I–II; Comstock Publishing Company, Inc.: New York, NY, USA, 1951; p. 339. [Google Scholar]
- Jensen, T.C.; Leinaas, H.P.; Hessen, D.O. Age-dependent shift in response to food element composition in Collembola: Contrasting effects of dietary nitrogen. Oecologia 2006, 149, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Endlweber, K.; Ruess, L.; Scheu, S. Collembola switch diet in presence of plant roots thereby functioning as herbivores. Soil Biol. Biochem. 2009, 41, 1151–1154. [Google Scholar] [CrossRef]
- Heděnec, P.; Radochová, P.; Nováková, A.; Kaneda, S.; Frouz, J. Grazing preference and utilization of soil fungi by Folsomia candida, (Isotomidae: Collembola). Eur. J. Soil Biol. 2013, 55, 66–70. [Google Scholar] [CrossRef]
- Staaden, S.; Milcu, A.; Rohlfs, M.; Scheu, S. Olfactory cues associated with fungal grazing intensity and secondary metabolite pathway modulate Collembola foraging behavior. Soil Biol. Biochem. 2011, 43, 1411–1416. [Google Scholar] [CrossRef]
- Hiol, F.H.; Dixon, R.K.; Curl, E.A. The feeding preference of mycophagous Collembola varies with the ectomycorrhizal symbiont. Mycorrhiza 1994, 5, 99–103. [Google Scholar] [CrossRef]
- Jałoszyński, P. Adults of european ant-like stone beetles (Coleoptera: Staphylinidae: Scydmaeninae) Scydmaenus tarsatus Müller & Kunze and Scydmaenus hellwigii (Herbst) prey on soft-bodied arthropods. Entomol. Sci. 2012, 15, 35–41. [Google Scholar]
- Tantawi, T.I.; El-Kady, E.M.; Greenberg, B.; El-Ghaffar, H.A. Arthropod succession on exposed rabbit carrion in Alexandria, Egypt. J. Med. Entomol. 1996, 33, 566–580. [Google Scholar] [CrossRef] [PubMed]
- Van Veen, J.A.; Kuikman, P.J. Soil structural aspects of decomposition of organic matter by microorganisms. Biogeochemistry 1990, 11, 213–233. [Google Scholar] [CrossRef]
- Schultz, P.A. Grazing preferences of two collembolan species, Folsomia candida and Proisotoma minuta, for ectomycorrhizal fungi. Pedobiologia 1991, 35, 313–325. [Google Scholar]
- Jonas, J.L.; Wilson, G.W.T.; White, P.M.; Joern, A. Consumption of mycorrhizal and saprophytic fungi by Collembola in grassland soils. Soil Biol. Biochem. 2007, 39, 2594–2602. [Google Scholar] [CrossRef]
- Jørgensen, H.B.; Elmholt, S.; Petersen, H. Collembolan dietary specialisation on soil grown fungi. Biol. Fertil. Soils 2003, 39, 9–15. [Google Scholar] [CrossRef]
- Ponge, J.F. Vertical distribution of Collembola (Hexapoda) and their food resources in organic horizons of beech forests. Biol. Fertil. Soils 2000, 32, 508–522. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Snider, R.J.; Snider, R.M. Food preference and effects of food type on the life history of some soil Collembola. Pedobiologia 1995, 39, 496–505. [Google Scholar]
- Bardgett, R.D.; van der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Ladygina, N.; Caruso, T.; Hedlund, K. Dietary switching of Collembola in grassland soil food webs. Soil Biol. Biochem. 2008, 40, 2898–2903. [Google Scholar] [CrossRef]
- Sechi, V.; D’Annibale, A.; Ambus, P.; Sárossy, Z.; Krogh, P.H.; Eriksen, J.; Holmstrup, M. Collembola feeding habits and niche specialization in agricultural grasslands of different composition. Soil Biol. Biochem. 2014, 74, 31–38. [Google Scholar] [CrossRef]
- Klironomos, J.N.; Widden, P.; Deslandes, I. Feeding preferences of the collembolan Folsomia candida in relation to microfungal successions on decaying litter. Soil Biol. Biochem. 1992, 24, 685–692. [Google Scholar] [CrossRef]
- Focken, U.; Becker, K. Metabolic fractionation of stable carbon isotopes: Implications of different proximate compositions for studies of the aquatic food webs using 13C data. Oecologia 1998, 115, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Oelbermann, K.; Scheu, S. Stable isotope enrichment (δ15N and δ13C) in a generalist predator (Pardosa lugubris, Araneae: Lycosidae): Effects of prey quality. Oecologia 2002, 130, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Ponsard, S.; Arditi, R. What can stable isotopes (δ15N and δ13C) tell about the food web of soil macroinvertebrates? Ecology 2000, 81, 852–864. [Google Scholar]
- Okuzaki, Y.; Tayasu, I.; Okuda, N.; Sota, T. Vertical heterogeneity of a forest floor invertebrate food web as indicated by stable-isotope analysis. Ecol. Res. 2009, 24, 1351–1359. [Google Scholar] [CrossRef]
- Gannes, L.Z.; O’Brien, D.M.; Rio, C.M.D. Stable isotopes in animal ecology: Assumptions, caveats, and a call for more laboratory experiments. Ecology 1997, 78, 1271–1276. [Google Scholar] [CrossRef]
- Griffiths, H. Applications of stable isotope technology in physiological ecology. Funct. Ecol. 1991, 5, 254–269. [Google Scholar] [CrossRef]
- McCutchan, J.H.; Lewis, W.M.; Kendall, C.; McGrath, C.C. Variation in trophic shift for stable isotope ratios of carbon, nitrogen, and sulfur. Oikos 2003, 102, 378–390. [Google Scholar] [CrossRef]
- Briones, M.J.I.; Ineson, P.; Sleep, D. Use of δ13C to determine food selection in collembolan species. Soil Biol. Biochem. 1999, 31, 937–940. [Google Scholar] [CrossRef]
- Hopkin, S.P. Biology of the Springtails (Insecta: Collembola); Oxford University Press: Oxford, UK, 1997; pp. 1–330. [Google Scholar]
- Handschin, E. Considérations sur la Position Systématique des Collemboles., Mémoires de la Société Royale d’Entomologie de Belgique; Tome Vingt-Septieme: Brussels, Belgium, 1955; Volume 27, pp. 40–53. [Google Scholar]
- Thibaud, J.M. Biologie et écologie des collemboles Hypogastruidae édaphiques et cavernicoles. Mém. Mus. Natl. d’Hist. Nat. Zool. A 1970, 61, 83–201. [Google Scholar]
- Garrity, G.M.; Bell, J.A.; Lilburn, T.G. Taxonomic outline of the prokaryotes release 5.0. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Springer: New York, NY, USA, 2004; pp. 1–399. [Google Scholar]
- Kirk, P.M.; Cannon, P.E.; Minter, D.W.; Stalpers, J.A. Dictionary of the Fungi, 10th ed.; CABI: London, UK, 2008; pp. 1–784. [Google Scholar]
- Janssen, P.H. Identifying the dominant soil bacterial taxa in libraries of 16S rRNA and 16S rRNA genes. Appl. Environ. Microbiol. 2006, 72, 1719–1728. [Google Scholar] [CrossRef] [PubMed]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavy, D.; Nedved, O.; Verhoef, H.A. Effects of starvation on body composition and cold tolerance in the collembolan Orchesella cincta and the isopod Porcellio scaber. J. Insect Physiol. 1997, 10, 973–978. [Google Scholar] [CrossRef]
- Fountain, M.T.; Hopkin, S.P. Folsomia candida (Collembola): A “standard” soil arthropod. Annu. Rev. Entomol. 2005, 50, 201–222. [Google Scholar] [CrossRef] [PubMed]
- OECD. Guidelines for the Testing of Chemicals. Collembola Reproduction Test. Version 3.4; OECD: Paris, France, 2008; p. 232. [Google Scholar]
- Scheu, S. The soil food web: Structure and perspectives. Eur. J. Soil Biol. 2002, 38, 11–20. [Google Scholar] [CrossRef]
- Anslan, S.; Bahram, M.; Tedersoo, L. Temporal changes in fungal communities associated with guts and appendages of Collembola as based on culturing and high-throughput sequencing. Soil Biol. Biochem. 2016, 96, 152–159. [Google Scholar] [CrossRef]
- Tiunov, A.V. Stable isotopes of carbon and nitrogen in soil ecological studies. Biol. Bull. 2007, 34, 395–407. [Google Scholar] [CrossRef]
- Phillips, D.L.; Gregg, J.W. Source partitioning using stable isotopes: Coping with too many sources. Oecologia 2003, 136, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Phillips, D.L.; Newsome, S.D.; Gregg, J.W. Combining sources in stable isotope mixing models: Alternative methods. Oecologia 2005, 144, 520–527. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria. Available online: http://www.R-project.org/ (accessed on 20 December 2018).
- Potapov, A.M.; Semenina, E.E.; Kurakov, A.V. Large 13C/12C and small 15N/14N isotope fractionation in an experimental detrital foodweb (litter–fungi–collembolans). Ecol. Res. 2013, 28, 1069–1079. [Google Scholar] [CrossRef]
- Martínez del Rio, C.; Wolf, N.; Carleton, S.A.; Gannes, L.Z. Isotopic ecology ten years after a call for more laboratory experiments. Biol. Rev. 2008, 84, 91–111. [Google Scholar] [CrossRef] [PubMed]
- Thimm, T.; Larink, O. Grazing preferences of some Collembola for endomycorrhizal fungi. Biol. Fertil. Soils 1995, 19, 266–268. [Google Scholar] [CrossRef]
- Kitaysky, A.S.; Piatt, J.F.; Hatch, S.A.; Kitaiskaia, E.V.; Benowitz-Fredericks, Z.M.; Shultz, T.; Wingfield, J.C. Food availability and population processes: Severity of nutritional stress during reproduction predicts survival of long-lived seabirds. Funct. Ecol. 2009, 24, 625–637. [Google Scholar] [CrossRef]
- Berg, M.P.; Bengtsson, J. Temporal and spatial variability in soil food web structure. Oikos 2007, 116, 1789–1804. [Google Scholar] [CrossRef]
- Castaño-Meneses, G.; Palacios-Vargas, J.G.; Cutz-Pool, L.Q. Feeding habits of Collembola and their ecological niche. An. Inst. Biol. Ser. Zool. 2004, 75, 135–142. [Google Scholar]
- Maraun, M.; Martens, H.; Migge, S.; Theenhaus, A.; Scheu, S. Adding to ‘the enigma of soil animal diversity’: Fungal feeders and saprophagous soil invertebrates prefer similar food substrates. Eur. J. Soil Biol. 2003, 39, 85–95. [Google Scholar] [CrossRef]
- Shelomi, M.; Matern, L.M.; Dinstell, J.M.; Harris, D.W.; Kimsey, R.B. DEET (N,N-Diethyl-meta-toluamide) induced delay of blowfly landing and oviposition rates on treated pig carrion (Sus scrofa L.). J. Forensic Sci. 2012, 57, 1507–1511. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The bodies of four insects and microorganisms (fungi and bacteria) growing on insect bodies are the three types of food sources. Comparing each of the weighted averages of the number of combinations with the δm, the comparative values for which the mass balance tolerances are between −0.1‰ and 0.1‰ are retained. For the contribution ratio in the figure, each dot represents the comparative values of the acceptable solution for the feeding proportion by E. proxima. Kruskal-Wallis explanatory values are statistically significant at p < 0.001.
Figure 1.
The bodies of four insects and microorganisms (fungi and bacteria) growing on insect bodies are the three types of food sources. Comparing each of the weighted averages of the number of combinations with the δm, the comparative values for which the mass balance tolerances are between −0.1‰ and 0.1‰ are retained. For the contribution ratio in the figure, each dot represents the comparative values of the acceptable solution for the feeding proportion by E. proxima. Kruskal-Wallis explanatory values are statistically significant at p < 0.001.
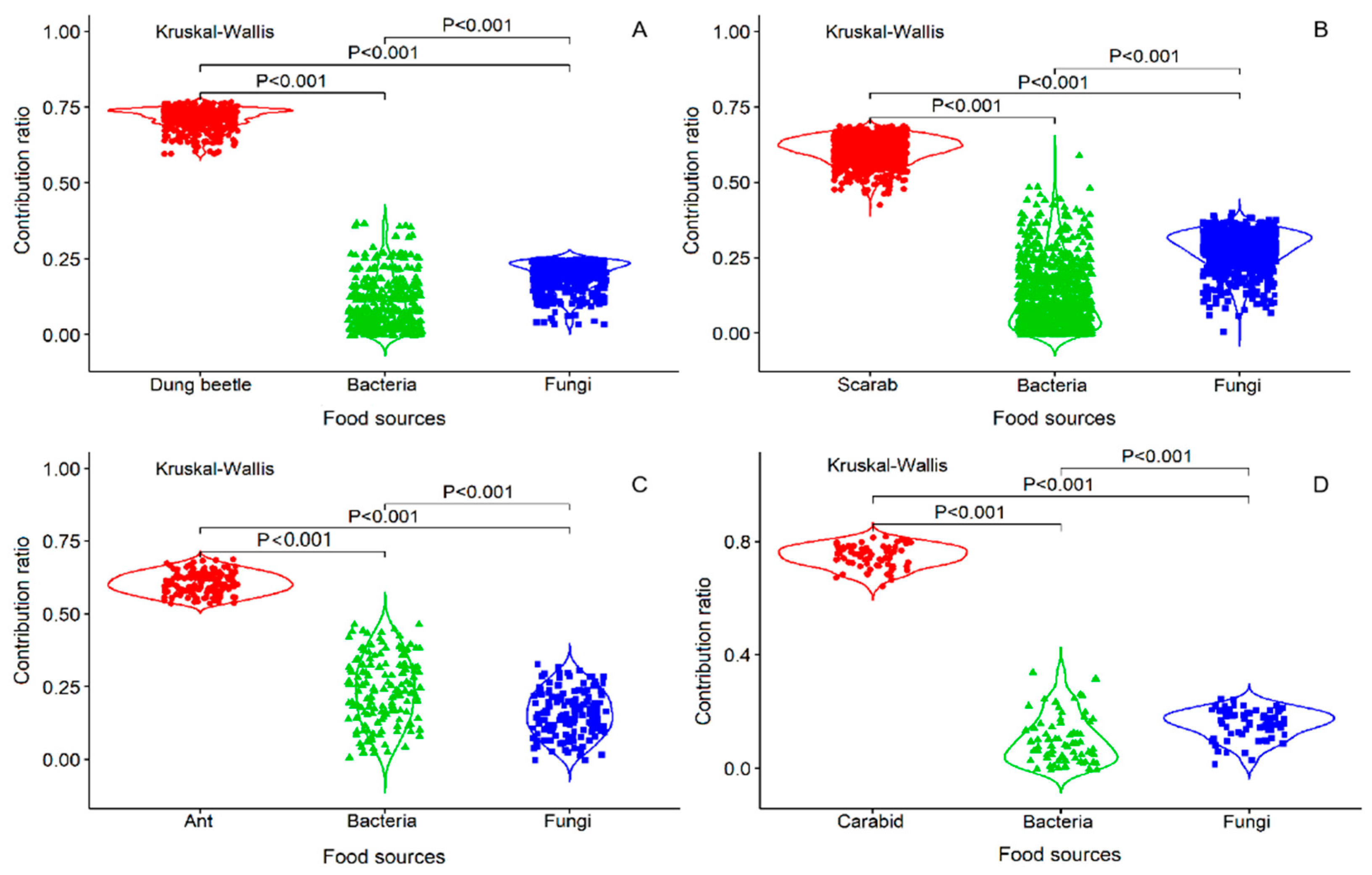
Figure 2.
Dietary biomass proportions of the three aggregate food sources for E. proxima. The aggregate solutions are sums of the biomass contributions for the food sources in each category (insects body, mixed bacteria, and mixed fungi). Increments of 2% were used to calculate the aggregate contributions in the model iterations.
Figure 2.
Dietary biomass proportions of the three aggregate food sources for E. proxima. The aggregate solutions are sums of the biomass contributions for the food sources in each category (insects body, mixed bacteria, and mixed fungi). Increments of 2% were used to calculate the aggregate contributions in the model iterations.

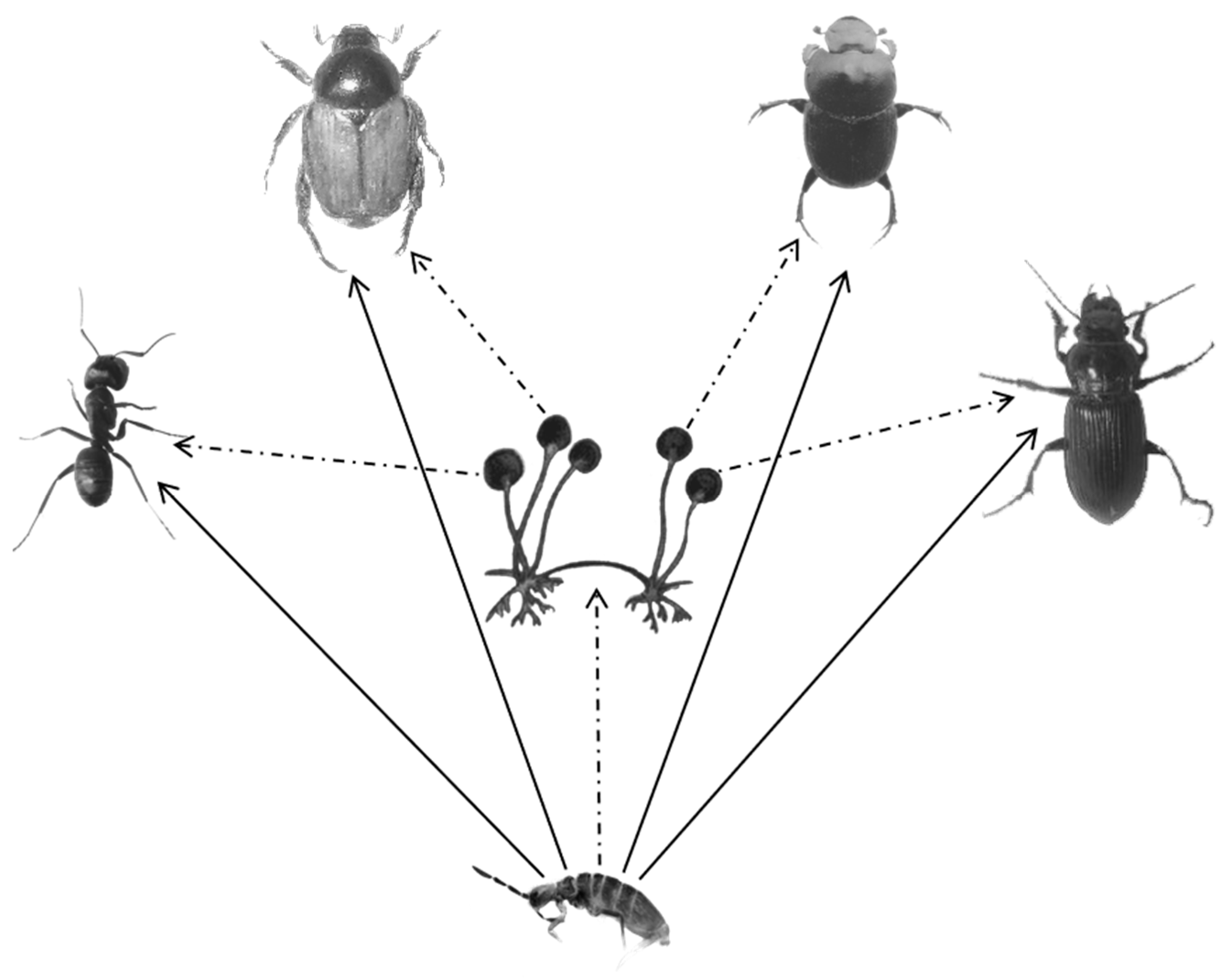
Figure 3.
Each of the insects were a dominant soil organism in the same habitat as an epigeic collembolan. The solid line with an arrow indicates that the collembolan can directly digest the four types of insects, and the dotted line with an arrow indicates that the collembolan may consume microorganisms (such as fungi (Rhizopus sp., Alternaria sp., Penicillium sp., and Aspergillus sp.) and bacteria (Bacillus sp1. and Bacillus sp2.) in an indirect manner.
Figure 3.
Each of the insects were a dominant soil organism in the same habitat as an epigeic collembolan. The solid line with an arrow indicates that the collembolan can directly digest the four types of insects, and the dotted line with an arrow indicates that the collembolan may consume microorganisms (such as fungi (Rhizopus sp., Alternaria sp., Penicillium sp., and Aspergillus sp.) and bacteria (Bacillus sp1. and Bacillus sp2.) in an indirect manner.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Calculations of the stable isotope values and aggregate distributions of each food source to E. proxima after feeding on the insect body, fungi, and bacteria using the IsoSource model.
Table 1.
Calculations of the stable isotope values and aggregate distributions of each food source to E. proxima after feeding on the insect body, fungi, and bacteria using the IsoSource model.
| Insect Carcasses | Food Sources | Stable Isotope Value (δ13C) after Feeding by E. proxima (‰) | δm Value | Median Value of of Each Dietary Aggregate Distribution (%) | Mean Percent Frequencies of Each Dietary Aggregate Distribution |
|---|---|---|---|---|---|
| Dung beetle | Body | −22.0530 | −19.9473 | 72 | 43 |
| Mixed fungi | −13.5415 | 20 | 35 | ||
| Mixed bacteria | −17.1413 | 8 | 26 | ||
| Scarab | Body | −24.7320 | −20.2697 | 62 | 59 |
| Mixed Fungi | −11.6806 | 28 | 36 | ||
| Mixed bacteria | −17.0270 | 10 | 31 | ||
| Ant | Body | −26.3250 | −21.9593 | 60 | 18 |
| Mixed fungi | −12.6193 | 16 | 8 | ||
| Mixed bacteria | −17.0197 | 24 | 6 | ||
| Carabid | Body | −20.7560 | −19.2826 | 75 | 7 |
| Mixed fungi | −13.4896 | 17 | 7 | ||
| Mixed bacteria | −16.7463 | 8 | 5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Feng, L.; Chang, L.; Zhang, S.; Zhu, X.; Adl, S.; Wu, D. What Is the Carcass-Usage Mode of the Collembola? A Case Study of Entomobrya proxima in the Laboratory. Insects 2019, 10, 67. https://doi.org/10.3390/insects10030067
AMA Style
Feng L, Chang L, Zhang S, Zhu X, Adl S, Wu D. What Is the Carcass-Usage Mode of the Collembola? A Case Study of Entomobrya proxima in the Laboratory. Insects. 2019; 10(3):67. https://doi.org/10.3390/insects10030067
Chicago/Turabian StyleFeng, Lichao, Liang Chang, Shaoqing Zhang, Xinyu Zhu, Sina Adl, and Donghui Wu. 2019. "What Is the Carcass-Usage Mode of the Collembola? A Case Study of Entomobrya proxima in the Laboratory" Insects 10, no. 3: 67. https://doi.org/10.3390/insects10030067
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.