1. Introduction
Fruit and vegetable production are important source of income for East African growers. Production of these crops is, however, constrained by insect pests and diseases resulting in yield loss and poor quality. The false codling moth (FCM),
Thaumatotibia leucotreta (Meyrick) (Lepidoptera: Tortricidae), one of the pests of these crops, is native to sub-Saharan Africa [
1] and has been recorded on 24 cultivated and 50 wild species in different plant families [
2]. It is a key pest of citrus (Rutaceae) [
3,
4], avocado,
Persea americana (Mill) (Lauraceae) [
5] macadamias,
Macadamia spp. (Proteaceae) [
6] and cotton,
Gossypium spp. (Malvaceae) [
1].
Thaumatotibia leucotreta is a multivoltine pest [
7] which does not enter diapause leading to year-round overlapping generations on host plants [
8]. The female moths lay eggs on fruit, often near the stylar end [
9]. The hatched larvae penetrate and feed inside the fruit resulting in fruit dropping. Damage symptoms caused by
T. leucotreta vary with the host plant. For example, scull on avocado [
5] and a yellowish-brown rind around a penetration hole on citrus orange [
4] have been documented. Larval incidence on orange can be up to 75% [
10]. In addition to direct losses,
T. leucotreta infestations also cause financial losses due to quarantine restrictions imposed on exporting countries and detection of a single larva can result in rejection of an entire consignment [
9].
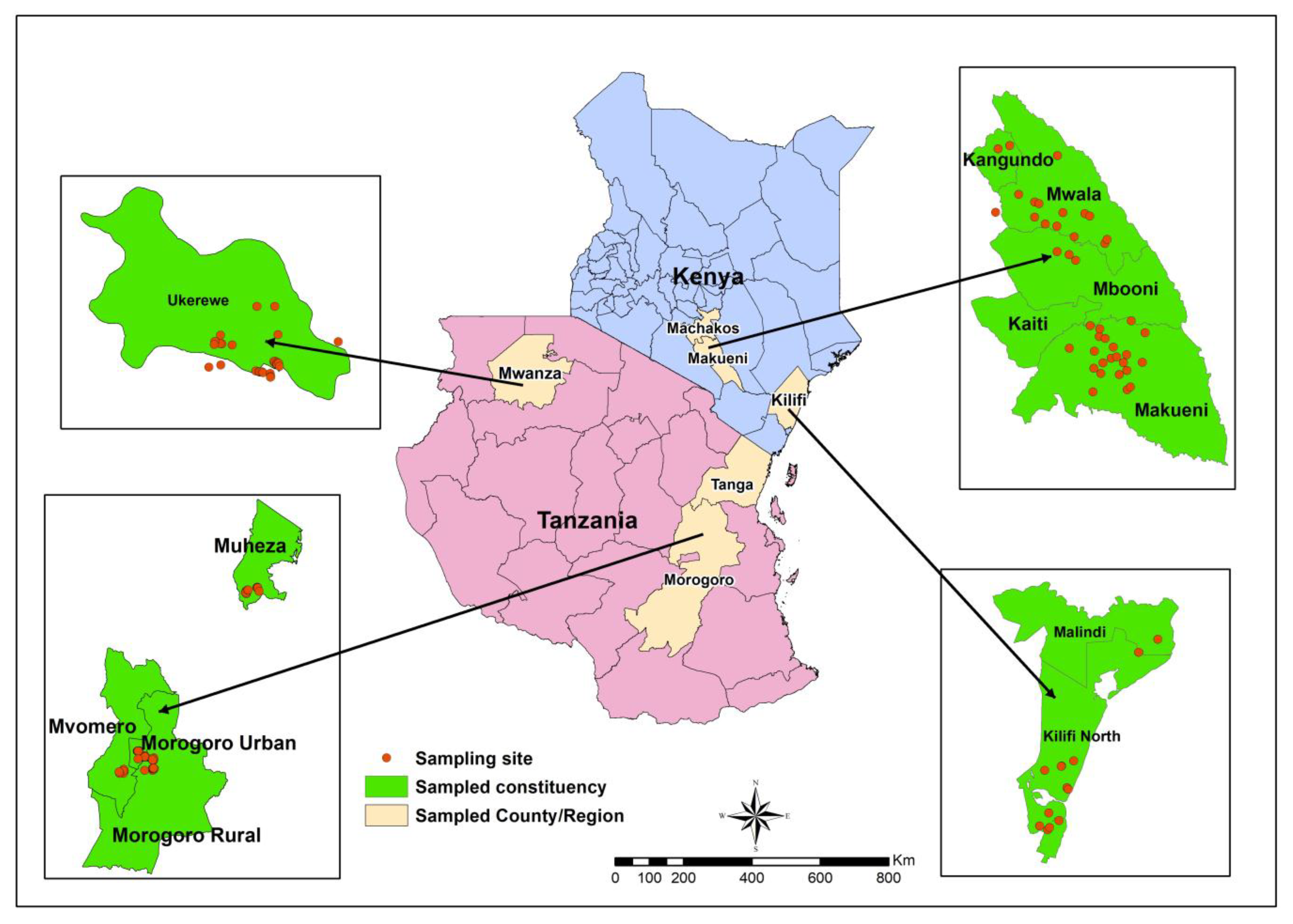
Although South Africa and Egypt are the largest citrus producing countries in Africa, Tanzania and Kenya are considered as the leading countries in citrus production in East Africa [
11]. Citrus production in Tanzania is largely concentrated on the North East Coast. The main production areas are in Tanga and Coast region, followed by Morogoro, Mwanza and Ruvuma. In Kenya, citrus production is concentrated in Coast, Eastern and Rift valley provinces [
12]. Although
T. leucotreta has been reported in Kenya and Tanzania [
13], little is known about the larval incidence of the pest especially during the citrus orange fruit harvesting period. Orange is produced from low to high altitudes in these countries. Altitudinal gradients and vegetation had been reported to influence distribution and abundance of moths [
14,
15,
16], which are highly diverse and ecologically important herbivorous insects [
17]. Odanga et al. [
13] reported similar
T. leucotreta infestations on avocado grown at different altitudinal gradients in Kenya and Tanzania. Knowledge on the effect of altitude on
T. leucotreta infestation on orange will contribute to management of the pest. The incidence of the pest on other crops which may serve as alternative host crops between successive orange fruiting seasons is not well known.
The ovipositional preference and offspring performance of
T. leucotreta on orange in a laboratory study was reported by Love et al. [
18]. The ovipositional preference of the pest on orange and vegetables has not been determined. According to Thompson and Pellmyr [
19], the plant selection made by egg laying females may often provide the initial basis for divergence of insect populations onto different plant species and it may drive the evolution of some plant defences. Developmental biology and adult life parameters of
T. leucotreta reared on artificial diet have been reported [
8,
20,
21,
22] and to a limited extend on orange, grapes and apple [
23]. However, no detailed study on the offspring performance of
T. leucotreta on other key host plants has been reported. The field dynamics of
T. leucotreta in a mixed cropping system, a common practice in sub-Saharan Africa, need to be investigated to develop better management strategies. The aims of this study were therefore to determine
T. leucotreta larval incidence on ripe orange as well as on mature vegetables of okra (
Abelmoschus esculentus (L.) Moench var. Clemson), African eggplant (
Solanum aethiopicum L., var. Tengeru white), chili pepper (
Capsicum anuum L., var. Jalapeno) and sweet pepper (
Capsicum anuum L., var. California Wonder). These vegetables are mainly grown near or within orange orchards. The developmental performance and life table parameters of the pest on three solanaceous vegetables
viz. chili, sweet pepper and African eggplant were also determined and compared to that of orange, the most common host.
4. Discussion
Thaumatotibia leucotreta occurred in all surveyed sites of Kenya and Tanzania from low to high altitudes. The higher percentage of damaged fruit recorded in Kenya compared to Tanzania could be attributed to the different farming practices within each country. In Kenya, most citrus trees are intercropped with peppers and maize which are also recorded as crop hosts of
T. leucotreta [
1,
6] while such practices are not common in Tanzania. Pests and diseases are generally controlled by means of pesticide application, which may result in a reduction of the natural enemy populations in the citrus orchards in Tanzania. According to Hofmeyr and Pringle [
30], excessive use of broad spectrum chemical pesticides causes pest outbreaks due to disruption of natural enemies and it could also lead to the development of insecticide resistance. The fruit sampled from the ground had a higher percentage of
T. leucotreta damage symptoms than those sampled from trees (with or without branch shaking). Higher
T. leucotreta larval incidence was, however, recorded from fruit sampled after shaking of the tree branches. This could be ascribed to
T. leucotreta laying eggs mainly on fruit while they are still on the trees [
31] and the larvae completing their development while the fruit is still on the tree [
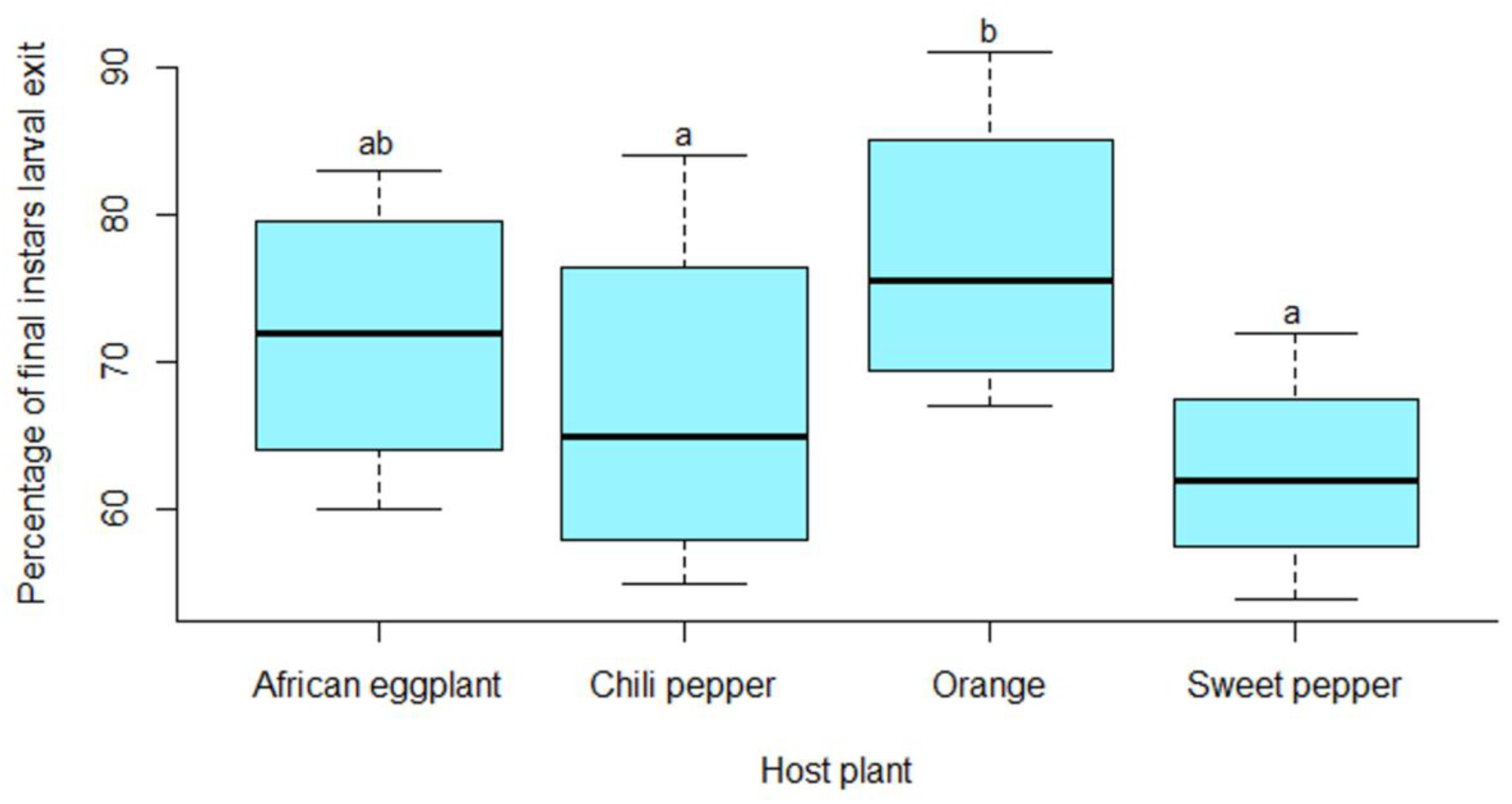
9]. When the fruit drop to the ground, the larvae would have already completed their development and exited the fruit to pupate in the soil. Lower larval incidence was therefore recorded in the fruit sampled on the ground. Although.
T. leucotreta infested fruit do often not show damage symptoms, final instar larvae may be present in the fruit mainly in the centre cores. This may have implications for
T. leucotreta management especially in terms of orchard sanitation and post-harvest treatment. Proper orange orchard sanitation and harvesting should therefore also take in consideration infested fruit that is still on the tree by shaking of the branches. Higher
T. leucotreta larval incidence on orange fruit was recorded in high than low altitudes of both countries. In contrast, Odanga et al. [
13] reported a similar population density and number of infested avocado fruit in different altitudes of Mount Kilimanjaro in Tanzania and Taita Hills in Kenya. This difference could possibly be ascribed to the difference in crop and altitudinal ranges.
Amongst the vegetables sampled, the highest percentage of
T. leucotreta infested fruit was on African eggplant compared to okra, sweet and chili peppers. This could be due to the lower moisture content of the fruit of this host plant, since infested African eggplant produced very little fluid under field and laboratory conditions. According to Jaenike [
32], the suitability of a plant for larval development is a function of many variables, including its chemical and physical properties, microhabitat and degree of infestation. During dissection of fruit, final instar larvae were also mainly found in the central core of orange and placenta of peppers where very little fluid was produced due to insect feeding and decay. African eggplant, okra, sweet and chili pepper serve as reservoirs for
T. leucotreta when grown in the vicinity or in mixed cropping with orange orchards. These vegetables are also grown throughout the year in irrigated fields which provides a continuous availability of host plants for this pest.
Although vegetables can serve as host plants for
T. leucotreta, oranges were the most preferred host plant for oviposition in both choice and no-choice tests. The
T. leucotreta moths do prefer certain orange varieties above others for oviposition. For example, Love et al. [
18] reported that Fischer Navels were the least preferred early maturing variety for oviposition. Navel oranges were reported to be more preferred for oviposition than Valencia oranges and guava [
33]. Most of the available literature report on the oviposition preference of
T. leucotreta for different citrus varieties and guava under laboratory and field conditions [
18,
33]. This study provides additional information on
T. leucotreta ovipositional preference for and performance on solanaceous vegetables, with chili and sweet pepper more preferred for oviposition than African eggplant.
In general, insects are reported to oviposit on hosts that maximize the performance of their progeny, a hypothesis referred to as “mother knows best” [
19,
32,
34,
35]. In this study, oviposition preference by
T. leucotreta did not mirror host suitability for development of the immatures and survival on these host plants. The sweet and chili peppers which were more preferred than African eggplant for oviposition were less suitable in terms of developmental time, larval mass and survival of immature stages (larvae and pupae) as well as female fecundity and net reproductive rate. The mismatch between preferred vegetable hosts for oviposition and host suitability of peppers could be explored for use as trap crop. A similar finding was reported for the butterfly,
Anthocharis cardamines (L) (Lepidoptera: Pieridae). The oviposition preference of females did not correlate with the fitness parameter of the offspring reared on 51 populations of two ploidy types of the perennial herb
Cardamine pratensis L. (Brassicales: Brassicaceae) [
36]. The suitability of the African eggplant for development of the immature stages of
T. leucotreta could be explained by the higher protein and lower moisture content of African eggplant compared to sweet and chili pepper [
37,
38,
39]. Rearing host quality has also been reported to influence the developmental duration of other lepidopteran species. For example, Traore et al. [
40] reported the larvae of
Maruca vitrata (Fabricius) (Lepidoptera: Pyralidae) reared on cowpea flowers to have a short developmental time compared to those reared on the other plant parts. The effect of host plants on the life table parameters is dependent on the quality of certain components, such as carbon, nitrogen and defensive metabolites [
41].
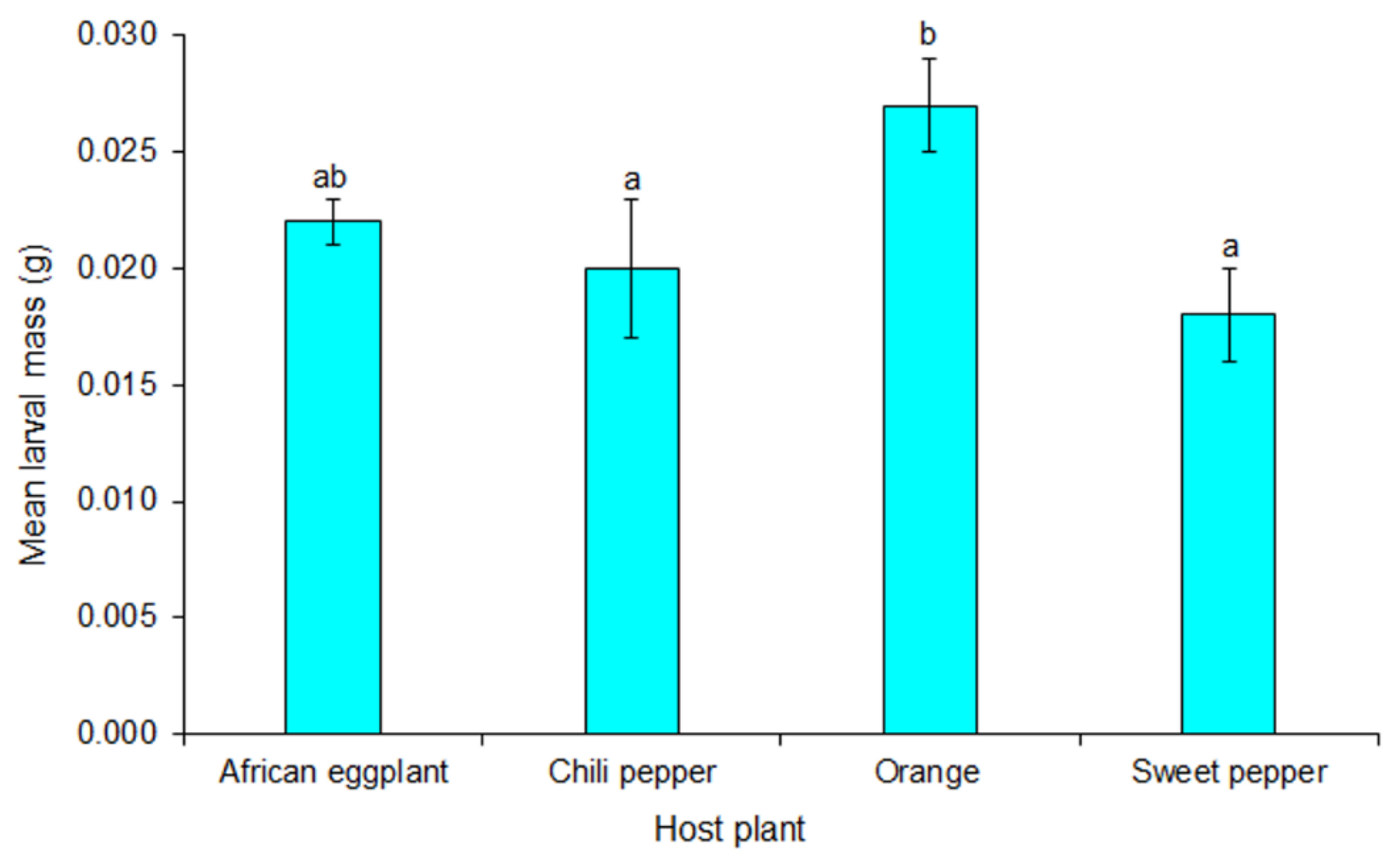
In this study, adult life table parameters for moths from larvae reared on the respective host plants are similar except for fecundity and net reproductive rate. Fecundity of
T. leucotreta ranged from a total of 341 to 415 eggs per female for those reared on sweet pepper and orange, respectively. Both these values are within the range reported for fecundity of
T. leucotreta from larvae reared on artificial diet [
8,
25]. Females reared on solanaceous vegetables laid fewer eggs than moths from larvae reared on orange. Daily egg laying peaked at day three and four after eclosion, for females reared on solanaceous vegetables and orange, respectively.
Thaumatotibia leucotreta oviposition from larvae reared on artificial diet reached a peak, 2 and 3 days earlier than for the females reared on solanaceous vegetables and orange, respectively [
8]. Generally, the highest daily fecundity was recorded for females reared from orange while the lowest was for females reared from sweet pepper. Oviposition peaked one day earlier (day three) for those of which larvae were reared on solanaceous vegetables than those reared on orange (day four) after moths emerged.
The intrinsic rate of natural increase, mean generation time, doubling time and finite rate of increase are important biotic parameters of insect performance [
42,
43]. Considering these parameters, survival of
T. leucotreta is equal on orange, sweet and chili pepper as well as African eggplant. This has implications for management strategies of the pest. According to Birch [
44] the intrinsic rate of increase is a basic parameter for insect population development, which is an indicator of the species developmental speed, longevity and fecundity [
28].
Although oranges sampled from the ground had more T. leucotreta damage symptoms compared to fruit sampled from the trees, the latter hosted a higher number of T. leucotreta larvae. It is therefore recommended that citrus trees should be subjected to gentle shaking prior to fruit harvesting to reduce citrus orange post-harvest fruit decay and minimize indirect losses due to quarantine measures. Infested orange fruit from both ground and tree after branch shaking should be collected and removed using sustainable sanitation measures such as the use of augumentorio which sequesters the moth while conserving any parasitoids. This control tactic should be integrated with other T. leucotreta control measures such as the use of biopesticides viz. Cryptophlebia leucotreta granulovirus (CrleGV), entomopathogenic nematode, Heterorhabditis bacteriophora, entomopathogenic fungi, Metarhizium anisopliae, Beauveria bassiana, attract and kill through the use of Lastcall® and the Sterile Insect Technique. The potential of solanaceous vegetables to act as T. leucotreta reservoirs between successive citrus orange seasons for rainfed orange production, the common cropping systems in Kenya and Tanzania were determined. The strong preference of T. leucotreta to chili and sweet peppers for oviposition and relatively higher mortality of the larvae on these crops could be explored for management of this pest using attract and kill approaches. Although there is currently a very potent male moth attractant commercially available, a need to identify semio-chemicals that can attract female T. leucotreta moths for both monitoring and suppression of the pest, is needed.

 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}