The Ser/Thr Kinase PknH Is Essential for Maintaining Heterocyst Pattern in the Cyanobacterium Anabaena sp. Strain PCC 7120


Department of Biological Sciences, Graduate School of Science, Tokyo Metropolitan University, 1-1 Minami-Osawa, Hachioji, Tokyo 192-0397, Japan
*
Author to whom correspondence should be addressed.
Life 2018, 8(3), 34; https://doi.org/10.3390/life8030034
Submission received: 6 July 2018
/
Revised: 21 August 2018
/
Accepted: 22 August 2018
/
Published: 24 August 2018
(This article belongs to the Special Issue Developmental Biology in Cyanobacteria)
Abstract
:In the filamentous cyanobacterium Anabaena sp. strain, PCC 7120, heterocysts (which are nitrogen-fixing cells) are formed in the absence of combined nitrogen in the medium. Heterocysts are separated from one another by 10 to 15 vegetative cells along the filaments, which consist of a few hundred of cells. hetR is necessary for heterocyst differentiation; and patS and hetN, expressed in heterocysts, play important roles in heterocyst pattern formation by laterally inhibiting the expression of hetR in adjacent cells. The results of this study indicated that pknH, which encodes a Ser/Thr kinase, was also involved in heterocyst pattern formation. In the pknH mutant, the heterocyst pattern was normal within 24 h after nitrogen deprivation, but multiple contiguous heterocysts were formed from 24 to 48 h. A time-lapse analysis of reporter strains harboring a fusion between gfp and the hetR promoter indicated that pknH was required to suppress hetR expression in cells adjacent to the preexisting heterocysts. These results indicated that pknH was necessary for the lateral inhibition of heterocyst differentiation to maintain the heterocyst pattern.
1. Introduction
Anabaena (Nostoc) sp. strain PCC 7120 (hereafter Anabaena) of the phylum Cyanobacteria is a model organism for observing prokaryotic cellular differentiation. Anabaena is a filamentous cyanobacterium, that forms unbranched multicellular filaments. In the presence of combined nitrogen, the filaments consist of only vegetative cells, which perform oxygenic photosynthesis. After depriving the cells of combined nitrogen, specific vegetative cells differentiate into heterocysts, which are cells that are specialized for nitrogen fixation, with a semiregular spacing of one heterocyst surrounded by approximately 10 vegetative cells [1,2]. When the vegetative cells between the heterocysts proliferate and increase, one vegetative cell in the middle of a string of vegetative cells differentiates into a heterocyst to maintain the spatial pattern.
The HetR protein is a master regulator of heterocyst differentiation and is necessary and sufficient for this process [3,4]. In response to nitrogen step-down, hetR expression increases in specific vegetative cells, and then the HetR-induced cells differentiate into heterocysts [5]. patS is induced at the early stages of heterocyst differentiation and plays a key role in heterocyst pattern formation [6,7]. The patS mutant exhibits the phenotype of multiple contiguous heterocysts (Mchs). The C-terminal pentapeptide or hexapeptide of PatS ([E]RGSGR) binds to HetR and inhibits its DNA-binding activity [8,9,10]. The RGSGR-containing peptides are transferred from the proheterocysts to adjacent vegetative cells, where heterocyst differentiation is suppressed [11]. The Mch phenotype is also observed in the mutants of the hetN gene, which is highly expressed in heterocysts at the late stages of differentiation [12,13]. The RGSGR motif is located in the central part of HetN and is necessary for the suppression of the Mch phenotype [14,15]. In the hetN mutant, the initial spatial pattern of the heterocysts is normal, but the Mch phenotype is observed after a prolonged incubation period under diazotrophic growth conditions. Therefore, PatS is involved in initial pattern formation after removal of combined nitrogen, and HetN is required for maintenance of the heterocyst pattern during diazotrophic growth.
pknH, which encodes a Ser/Thr kinase, is exclusively expressed in the heterocysts [16]. In a previous report, it was proposed that pknH is involved in stabilizing cell junctions, particularly those between heterocysts and vegetative cells [16]. The pknH mutant can form heterocysts with nitrogenase activity. However, most heterocysts are detached from the filaments, resulting in filament fragmentation. Thus, the pknH mutant exhibits a growth defect under diazotrophic growth conditions. In the present study, we found that the pknH mutant exhibits the Mch phenotype. At 24 h after nitrogen deprivation, the heterocyst pattern of the pknH mutant was normal, but contiguous heterocysts were formed during the subsequent 24 h. We conducted time-lapse analysis of heterocyst development from 24 to 48 h after nitrogen deprivation and found that the vegetative cells adjacent to the preexisting heterocysts differentiated into heterocysts, resulting in the Mch phenotype. We also report here the dynamics of hetR expression in the heterocysts’ neighboring cells in the wild-type (WT) strain and the pknH mutant.
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
Anabaena and its derivatives were grown in BG-11 medium (containing NaNO3 as a nitrogen source), as previously described [17]. Liquid cultures were infused with air containing 1.0% (v/v) CO2. For nitrogen-deprivation experiments, cells grown in BG-11 medium up to the mid-logarithmic phase (OD750 of 0.4–0.5) were washed three times with nitrogen-free medium (BG-110) and then resuspended in BG-110 medium containing 5 mM NaHCO3. Spectinomycin or neomycin was added to the medium at a final concentration of 10 or 30 µg mL−1, respectively, as required.
2.2. Construction of Mutant Strains
To construct complementation strains, after substituting Asn for Asp at position 184, pknH and pknHD184N were inserted into its original locus on the chromosome of the pknH disruptant DR1336S [16], as follows. A DNA fragment containing the pknH gene was amplified by PCR using the primer pair 1336-5F and PknH-R (Table 1) and cloned between the SacI and SalI sites of the suicide vector pSU101 [18] to construct pSpknH. The plasmid pSpknHD184N, which contains the pknHD184N allele, was generated by site-directed mutagenesis using the PrimeSTAR Mutagenesis Basal Kit (TaKaRa Bio, Inc., Otsu, Japan) and pSpknH as templates. The resultant plasmids were transferred into DR1336S according to the method of Elhai et al. [19], and single recombinants were selected on a BG-11 plate containing spectinomycin and neomycin.
To construct green fluorescent protein (GFP) reporter strains, the plasmid pRhetRG [5], which harbors a transcriptional-fusion of gfp and the hetR promoter, was transferred into the WT strain and DR1336G, and single recombinants were selected.
2.3. RNA Analysis
Total RNA was purified from whole filaments according to the method of Pinto et al. [20], and residual genomic DNA was removed by treating with DNase I (TaKaRa Bio, Inc., Shiga, Japan). Quantitative reverse transcription qRT-PCR was performed as previously described [21] using the primer pairs listed in Table 1.
2.4. Microscopic Analysis of Development of Heterocyst Spatial Patterns
Filaments of Anabaena suspended in 5 µL of BG-110 medium were applied to a thin (0.5 mm) pad of BG-110 medium containing 1% agarose on a chambered cover glass (AGC Techno Glass, Shizuoka, Japan) prepared by following the method described by Aldea et al. [22]. The filaments on the agarose pad were incubated at 30 °C with continuous illumination at 30 µmol photons m−2 s−1, and fluorescent images of GFPs and phycobilisomes were taken using the AXIO Imager A2 fluorescence microscope (Carl Zeiss AG, Oberkochen, Germany) with high-efficiency filter sets 38 and 50, respectively. Small black dots were painted on the cover glass as positional markers to track the same microscopic field during sequential observations. The images were captured with a DP73 digital camera (Olympus Corporation, Tokyo, Japan). To quantify the fluorescence intensity, images were converted to grayscale, and the noise was removed using a median filter (5 by 5 cross-shaped window). The fluorescence intensity of each cell was measured using ImageJ v1.46r (National Institutes of Health, Bethesda, MD, USA). GFP fluorescence levels for each cell were normalized using the following formula:
where is the intensity value in cell position i at time t, and nv is the normalized GFP fluorescence level.
3. Results
3.1. Spatial Pattern of Heterocysts in the pknH Mutant
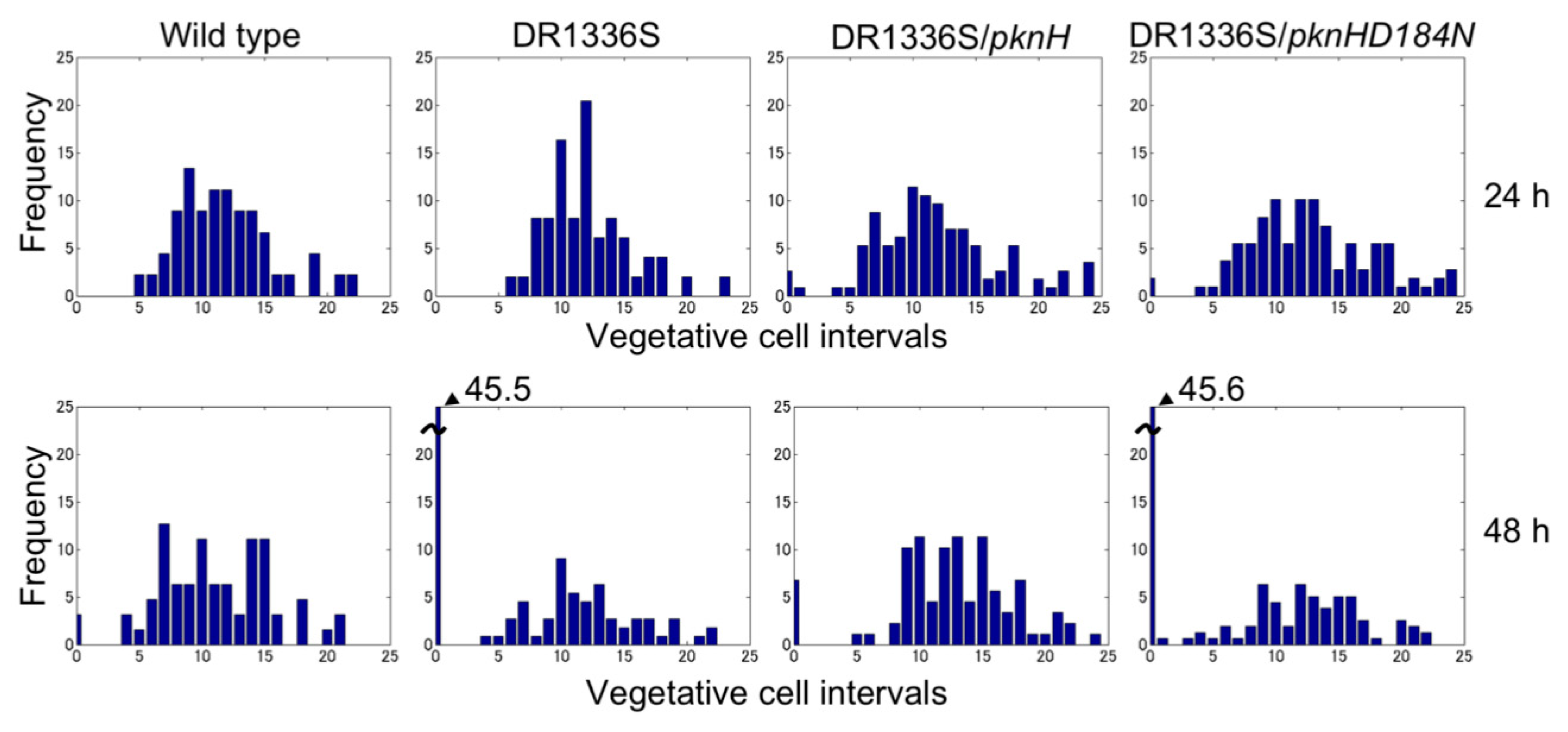
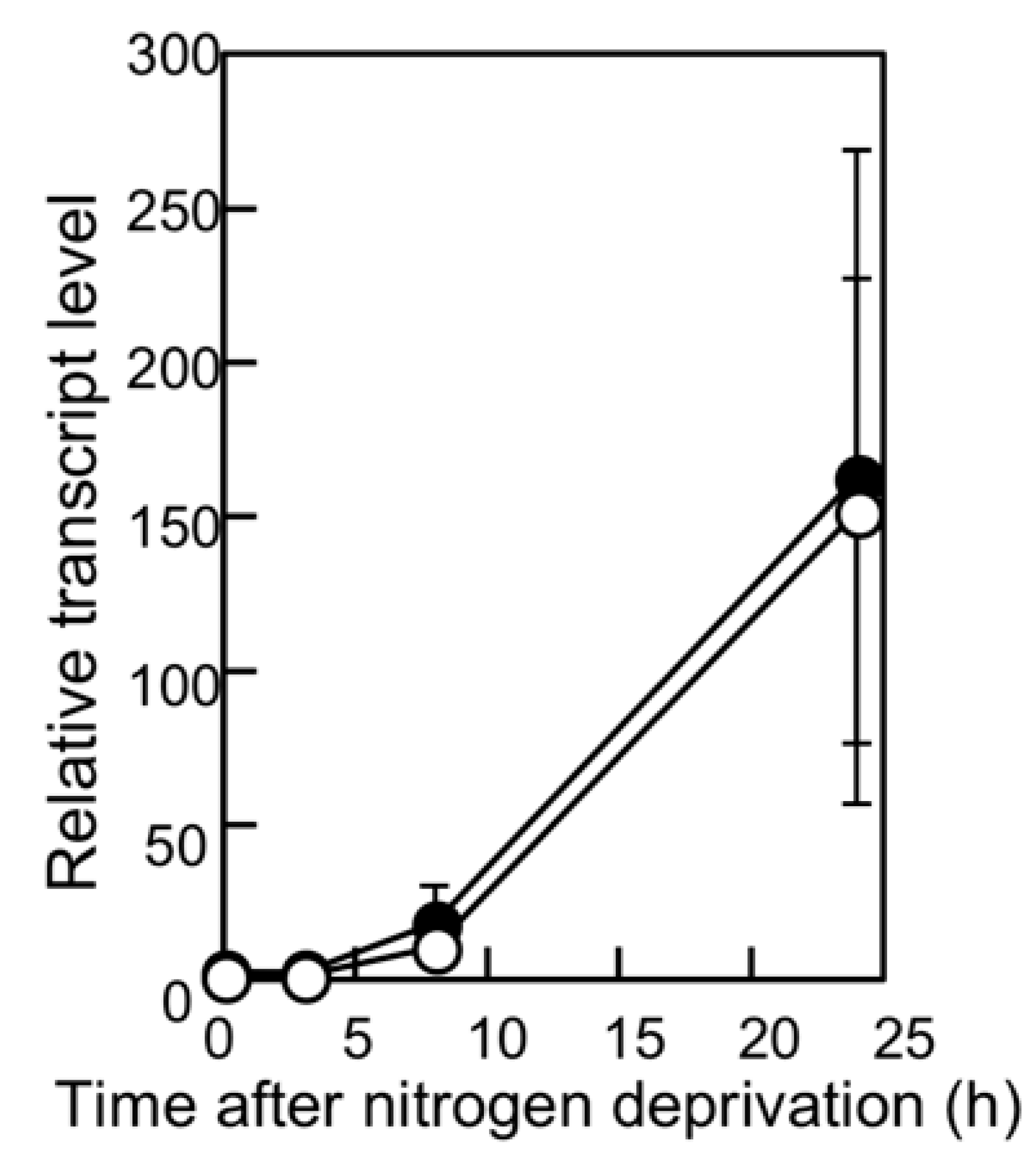
We have previously reported that filaments in the pknH-deleted mutant were fragmented during incubation in the absence of combined nitrogen, resulting in many heterocysts being detached from the filaments. Hence, we proposed that pknH is involved in maintaining connections between heterocysts and vegetative cells [16]. We attempted to maintain the connections between the heterocysts and filaments in the pknH mutant. To prevent the heterocyst detachment that is caused by bubbling with air containing 1% (w/w) CO2, heterocyst differentiation was induced in the nitrogen-free medium containing 5 mM NaHCO3 with neither aeration nor agitation. We found pairs of heterocysts and, at times, three or four contiguous heterocysts in the filaments of the pknH mutant (Figure 1). Figure 2 shows the number and distribution of the vegetative cells separating two heterocysts. After 24 h of nitrogen deprivation, most heterocysts were separated by 8–15 vegetative cells, and few contiguous heterocysts were formed in either the WT strain or the pknH mutant. However, after 48 h, the percentage of contiguous heterocysts was greatly increased in the pknH mutant as compared to the WT strain. Therefore, the Mch phenotype in the pknH mutant appeared from 24 to 48 h after nitrogen deprivation. Because the phenotype of the pknH mutant closely resembles that of the hetN mutant [13], the expression of hetN after nitrogen deprivation in the pknH mutant was determined (Figure 3). Changes in the transcript level of hetN in response to nitrogen deprivation were comparable in the pknH mutant and the WT strain.
The PknH protein has a Ser/Thr protein kinase domain with 12 amino acid residues that are conserved among the catalytic domains of Ser/Thr kinases. An Asp residue at position 184 of PknH corresponds to the Asp residue of the highly conserved Asp-Phe-Gly motif of the activation loop, which is necessary for phosphorylation activity [23,24]. A pknH gene encoding a PknH protein with a substitution of Asn for Asp at position 184 (pknHD184N) was introduced into the genome of the pknH mutant. Although the heterocyst pattern was recovered by complementation with the original pknH, the strain having pknHD184N showed a Mch phenotype that was similar to that of the pknH mutant (Figure 2), which indicated that Asp-184 of PknH is necessary for suppression of the Mch phenotype.
3.2. Time-Lapse Analysis of Heterocyst Development in the pknH Mutant
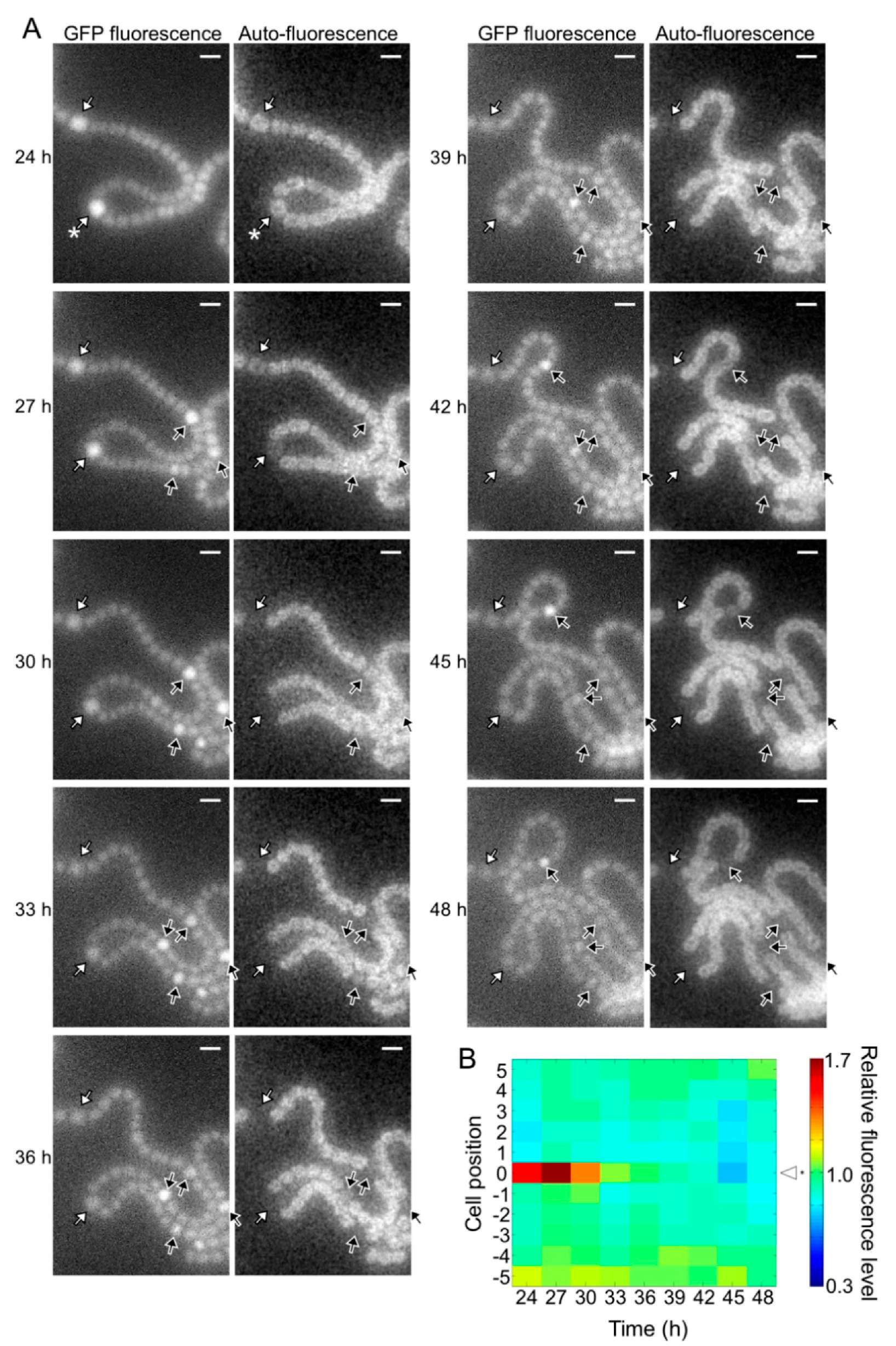
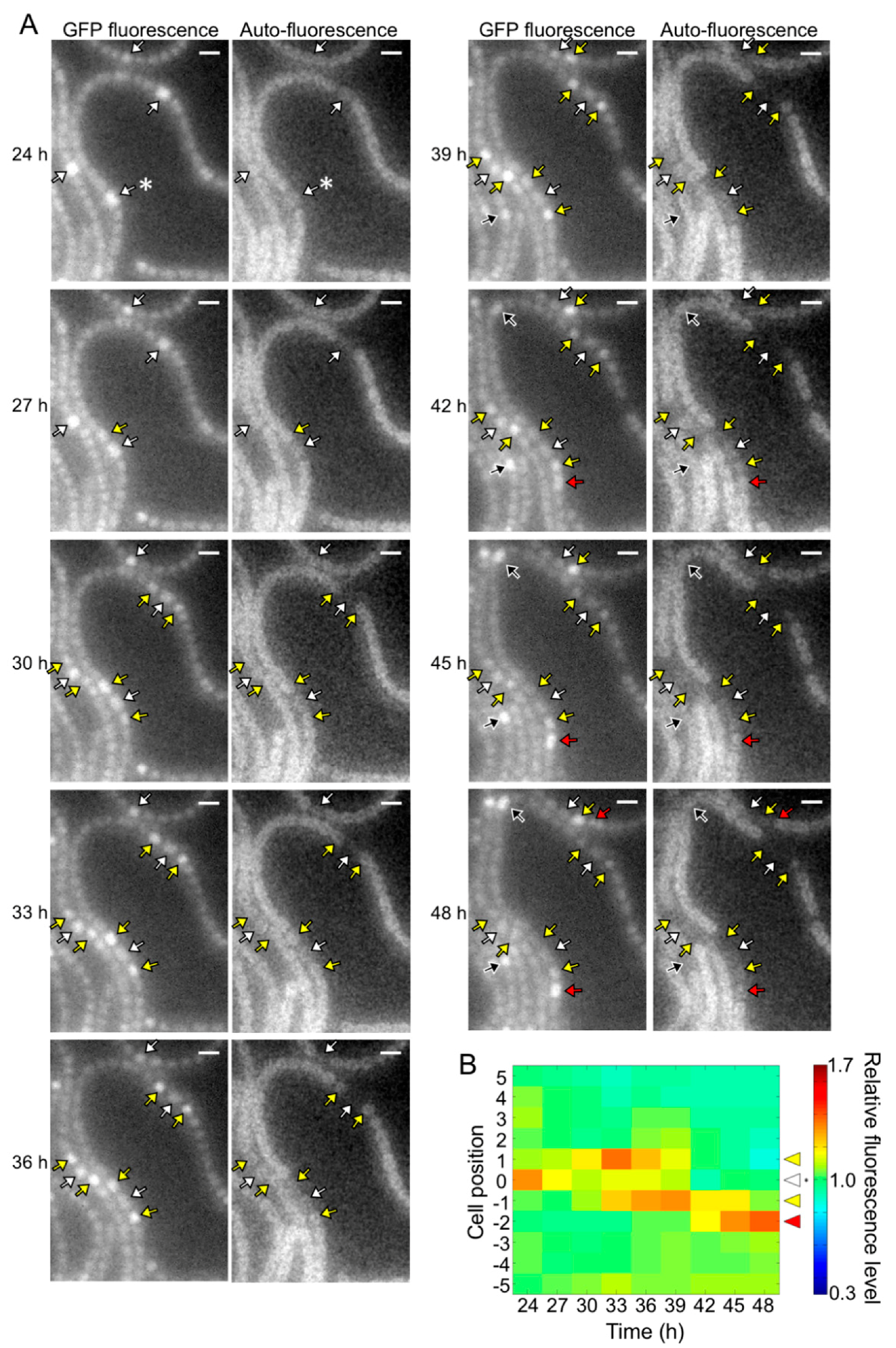
The pknH mutant showed a normal heterocyst pattern 24 h after nitrogen deprivation, but Mchs were observed after 48 h (Figure 2). We conducted a time-lapse analysis of heterocyst development during the 24 to 48 h period. Heterocyst differentiation was induced by settling filaments on agar medium containing no combined nitrogen sources. Heterocyst formation was monitored by GFP fluorescence that was expressed from the hetR promoter and auto-fluorescence from photosynthetic pigments. At 24 h, high GFP fluorescence was observed in some cells and auto-fluorescence of these cells decreased during subsequent incubation, indicating that these cells were heterocysts (white arrows in Figure 4A). In the WT background, GFP fluorescence of cells located in the center of a string of vegetative cells increased at 27 h with the proliferation of vegetative cells, and then auto-fluorescence of these cells decreased (black arrows in Figure 4A). Thus, the heterocyst distribution was maintained from 24 to 48 h. Meanwhile, in the pknH mutant, GFP fluorescence increased in cells adjacent to preexisting heterocysts (yellow arrows in Figure 5A). These cells with high GFP fluorescence differentiated into heterocysts, as indicated by a decrease in auto-fluorescence, resulting in the formation of Mchs. Moreover, further differentiation was initiated at 42 h at a cell adjacent to Mchs (red arrows in Figure 5A). On the other hand, de novo differentiation of a single heterocyst was observed in the middle of a string of vegetative cells (black arrows in Figure 5A). Therefore, the Mch phenotype in the pknH mutant was not formed by simultaneous differentiation of contiguous vegetative cells but by sequential differentiation of vegetative cells adjacent to preexisting heterocysts. We quantified the GFP fluorescence level of each cell and analyzed the temporal changes in hetR expression in cells adjacent to heterocysts. In the WT background, an increase in hetR expression was not observed in cells adjacent to heterocysts (Figure 4B). However, in the pknH mutant, hetR expression was induced in cells adjacent to the preexisting heterocysts in 23 out of the 30 heterocysts analyzed (Figure 5B). These results suggest that vegetative cells adjacent to heterocysts would have a tendency to increase hetR expression and that PknH, which is exclusively expressed in heterocysts [16], suppresses the induction of hetR in neighboring vegetative cells.
4. Discussion
In the present study, we demonstrated that the pknH mutant exhibited a Mch phenotype by statically inducing heterocyst differentiation, indicating that pknH is involved in heterocyst pattern formation. We have previously shown that heterocysts of the pknH mutant are easily detached from the filaments and proposed that pknH is involved in maintaining the connection between heterocysts and vegetative cells [16]. However, detachment of the heterocysts from the filaments in the pknH mutant would be initiated by breaking the connection between contiguous heterocysts, not by breaking the connection between heterocysts and vegetative cells. Heterocysts have a thick envelop outside the cell wall and are connected to adjacent cells by the narrowed septum at their poles [25]. The fragile connection between contiguous heterocysts would be responsible for fragmentation of the filaments in the pknH mutant, although the possibility that differences in culture conditions, such as bubbling with air containing 1% CO2 in the previous study or incubation on an agar medium containing 5 mM NaHCO3 under atmospheric CO2 levels in this study, affect the phenotype of the pknH mutant cannot be ruled out.
The pknH mutant initially exhibited the normal heterocyst pattern, but 48 h after nitrogen deprivation, one-half of the heterocysts were contiguous to another heterocyst (Figure 2). Thus, pknH is necessary for maintaining the heterocyst pattern during diazotrophic growth. Time-lapse analysis indicated that the expression of hetR increased in vegetative cells adjacent to preexisting heterocysts in the pknH mutant (Figure 5). Suppression of heterocyst differentiation of vegetative cells adjacent to preexisting heterocysts is likely to be stronger than that of vegetative cells separated from heterocysts because of the nitrogen-fixing products supplied by the heterocysts. During heterocyst differentiation, the expression of hetR is initially upregulated in groups of filament cells and then becomes restricted to a single cell [26,27]. An elevated signal that upregulates the expression of hetR might be maintained in cells adjacent to heterocysts.
The molecular mechanisms by which PknH suppresses hetR expression in cells adjacent to heterocysts, where the pknH gene is not expressed, remains to be elucidated; however, it was shown that Asp-184 of PknH, which is conserved in the catalytic domain of Ser/Thr kinases, was essential for suppression of Mch (Figure 2). The phosphorylation activity of PknH could be related to the hetN-dependent downregulation of HetR because the delayed Mch phenotype was also observed in the hetN mutant [13]. Since the hetN transcript levels were not affected by pknH disruption (Figure 3), PknH would not be involved in the transcriptional regulation of hetN. The hetN-dependent signal is transferred to adjacent vegetative cells [28], and the septal protein SepJ and predicted ABC transporter HetC play important roles in the intercellular transfer of the hetN-dependent signal [29,30]. PknH could enhance the intercellular movement of the hetN-dependent signal through interaction with these proteins. Alternatively, PknH might be related to processing the HetN protein. It has been reported that there were PatS-processing activities within cell extracts of Anabaena [31]. patS encodes a protein with 17 amino acids [11], but the PatS C-terminal hexapeptide (ERGSGR) has been proposed to be an active form of PatS [9]. Because the full-length HetN peptide is not intercellularly transferred, HetN should be cleaved before it is transferred [28]. HetN-processing activities might be activated by PknH. The pknH/hetN double mutant would be useful for understanding the relationship between PknH and HetN. Moreover, identification of proteins that are phosphorylated by PknH is crucial for elucidation of PknH’s function in maintaining the heterocyst pattern.
Author Contributions
S.-i.F. and S.E. designed the research, performed the experiments, analyzed the data and wrote the paper. Both authors have read and approved the final manuscript.
Funding
This work was supported in part by the Japan Science and Technology Agency [Core Research for Evolutional Science and Technology] and the Japan Society for the Promotion of Science [Grant-in-aid for Young Scientists (B) 26870472] to Shigeki Ehira.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Flores, E.; Herrero, A. Compartmentalized function through cell differentiation in filamentous cyanobacteria. Nat. Rev. Microbiol. 2010, 8, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.; Mella-Herrera, R.A.; Golden, J.W. Cyanobacterial heterocysts. Cold Spring Harb. Perspect. Biol. 2010, 2, a000315. [Google Scholar] [CrossRef] [PubMed]
- Buikema, W.J.; Haselkorn, R. Characterization of a gene controlling heterocyst differentiation in the cyanobacterium Anabaena 7120. Genes Dev. 1991, 5, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Buikema, W.J.; Haselkorn, R. Expression of the Anabaena hetR gene from a copper-regulated promoter leads to heterocyst differentiation under repressing conditions. Proc. Natl. Acad. Sci. USA 2001, 98, 2729–2734. [Google Scholar] [CrossRef] [PubMed]
- Asai, H.; Iwamori, S.; Kawai, K.; Ehira, S.; Ishihara, J.; Aihara, K.; Shoji, S.; Iwasaki, H. Cyanobacterial cell lineage analysis of the spatiotemporal hetR expression profile during heterocyst pattern formation in Anabaena sp. PCC 7120. PLoS ONE 2009, 4, e7371. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.S.; Golden, J.W. PatS and products of nitrogen fixation control heterocyst pattern. J. Bacteriol. 2001, 183, 2605–2613. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.; Golden, J.W. Heterocyst pattern formation controlled by a diffusible peptide. Science 1998, 282, 935–938. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Dong, Y.; Zhao, J. HetR homodimer is a DNA-binding protein required for heterocyst differentiation, and the DNA-binding activity is inhibited by PatS. Proc. Natl. Acad. Sci. USA 2004, 101, 4848–4853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldmann, E.A.; Ni, S.; Sahu, I.D.; Mishler, C.H.; Levengood, J.D.; Kushnir, Y.; McCarrick, R.M.; Lorigan, G.A.; Tolbert, B.S.; Callahan, S.M.; et al. Differential binding between PatS C-terminal peptide fragments and HetR from Anabaena sp. PCC 7120. Biochemistry 2012, 51, 2436–2442. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.X.; Jiang, Y.L.; Zhao, M.X.; Cai, K.; Liu, S.; Wen, B.; Lv, P.; Zhang, Y.; Peng, J.; Zhong, H.; et al. Structural insights into HetR–PatS interaction involved in cyanobacterial pattern formation. Sci. Rep. 2015, 5, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Corrales-Guerrero, L.; Mariscal, V.; Flores, E.; Herrero, A. Functional dissection and evidence for intercellular transfer of the heterocyst-differentiation PatS morphogen. Mol. Microbiol. 2013, 88, 1093–1105. [Google Scholar] [CrossRef] [PubMed]
- Bauer, C.C.; Ramaswamy, K.S.; Endley, S.; Scappino, L.A.; Golden, J.W.; Haselkorn, R. Suppression of heterocyst differentiation in Anabaena PCC 7120 by a cosmid carrying wild-type genes encoding enzymes for fatty acid synthesis. FEMS Microbiol. Lett. 1997, 151, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Callahan, S.M.; Buikema, W.J. The role of HetN in maintenance of the heterocyst pattern in Anabaena sp. PCC 7120. Mol. Microbiol. 2001, 40, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Corrales-Guerrero, L.; Mariscal, V.; Nurnberg, D.J.; Elhai, J.; Mullineaux, C.W.; Flores, E.; Herrero, A. Subcellular localization and clues for the function of the HetN factor influencing heterocyst distribution in Anabaena sp. strain PCC 7120. J. Bacteriol. 2014, 196, 3452–3460. [Google Scholar] [CrossRef] [PubMed]
- Higa, K.C.; Rajagopalan, R.; Risser, D.D.; Rivers, O.S.; Tom, S.K.; Videau, P.; Callahan, S.M. The RGSGR amino acid motif of the intercellular signalling protein, HetN, is required for patterning of heterocysts in Anabaena sp. strain PCC 7120. Mol. Microbiol. 2012, 83, 682–693. [Google Scholar] [CrossRef] [PubMed]
- Ehira, S.; Ohmori, M. The pknH gene restrictively expressed in heterocysts is required for diazotrophic growth in the cyanobacterium Anabaena sp. strain PCC 7120. Microbiology 2012, 158, 1437–1443. [Google Scholar] [CrossRef] [PubMed]
- Ehira, S.; Ohmori, M. NrrA, a nitrogen-responsive response regulator facilitates heterocyst development in the cyanobacterium Anabaena sp. strain PCC 7120. Mol. Microbiol. 2006, 59, 1692–1703. [Google Scholar] [CrossRef] [PubMed]
- Ehira, S.; Shimmori, Y.; Watanabe, S.; Kato, H.; Yoshikawa, H.; Ohmori, M. The nitrogen-regulated response regulator NrrA is a conserved regulator of glycogen catabolism in β-cyanobacteria. Microbiology 2017, 163, 1711–1719. [Google Scholar] [CrossRef] [PubMed]
- Elhai, J.; Vepritskiy, A.; Muro-Pastor, A.M.; Flores, E.; Wolk, C.P. Reduction of conjugal transfer efficiency by three restriction activities of Anabaena sp. strain PCC 7120. J. Bacteriol. 1997, 179, 1998–2005. [Google Scholar] [CrossRef] [PubMed]
- Pinto, F.L.; Thapper, A.; Sontheim, W.; Lindblad, P. Analysis of current and alternative phenol based RNA extraction methodologies for cyanobacteria. BMC Mol. Biol. 2009, 10, 79. [Google Scholar] [CrossRef] [PubMed]
- Ehira, S.; Ohmori, M. NrrA, a nitrogen-regulated response regulator protein, controls glycogen catabolism in the nitrogen-fixing cyanobacterium Anabaena sp. strain PCC 7120. J. Biol. Chem. 2011, 286, 38109–38114. [Google Scholar] [CrossRef] [PubMed]
- Aldea, M.R.; Mella-Herrera, R.A.; Golden, J.W. Sigma factor genes sigC, sigE, and sigG are upregulated in heterocysts of the cyanobacterium Anabaena sp. strain PCC 7120. J. Bacteriol. 2007, 189, 8392–8396. [Google Scholar] [CrossRef] [PubMed]
- Huse, M.; Kuriyan, J. The conformational plasticity of protein kinases. Cell 2002, 109, 275–282. [Google Scholar] [CrossRef]
- Yoshida, S.; Parniske, M. Regulation of plant symbiosis receptor kinase through serine and threonine phosphorylation. J. Biol. Chem. 2005, 280, 9203–9209. [Google Scholar] [CrossRef] [PubMed]
- Herrero, A.; Stavans, J.; Flores, E. The multicellular nature of filamentous heterocyst-forming cyanobacteria. FEMS Microbiol. Rev. 2016, 40, 831–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corrales-Guerrero, L.; Tal, A.; Arbel-Goren, R.; Mariscal, V.; Flores, E.; Herrero, A.; Stavans, J. Spatial fluctuations in expression of the heterocyst differentiation regulatory gene hetR in Anabaena filaments. PLoS Genet. 2015, 11, e1005031. [Google Scholar] [CrossRef] [PubMed]
- Black, T.A.; Cai, Y.; Wolk, C.P. Spatial expression and autoregulation of hetR, a gene involved in the control of heterocyst development in Anabaena. Mol. Microbiol. 1993, 9, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Rivers, O.S.; Videau, P.; Callahan, S.M. Mutation of sepJ reduces the intercellular signal range of a hetN-dependent paracrine signal, but not of a patS-dependent signal, in the filamentous cyanobacterium Anabaena sp. strain PCC 7120. Mol. Microbiol. 2014, 94, 1260–1271. [Google Scholar] [CrossRef] [PubMed]
- Mariscal, V.; Nürnberg, D.J.; Herrero, A.; Mullineaux, C.W.; Flores, E. Overexpression of SepJ alters septal morphology and heterocyst pattern regulated by diffusible signals in Anabaena. Mol. Microbiol. 2016, 101, 968–981. [Google Scholar] [CrossRef] [PubMed]
- Videau, P.; Rivers, O.S.; Higa, K.C.; Callahan, S.M. ABC transporter required for intercellular transfer of developmental signals in a heterocystous cyanobacterium. J. Bacteriol. 2015, 197, 2685–2693. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhou, F.; Wang, S.; Xu, X. Processing of PatS, a morphogen precursor, in cell extracts of Anabaena sp. PCC 7120. FEBS Lett. 2017, 591, 751–759. [Google Scholar] [CrossRef] [PubMed]
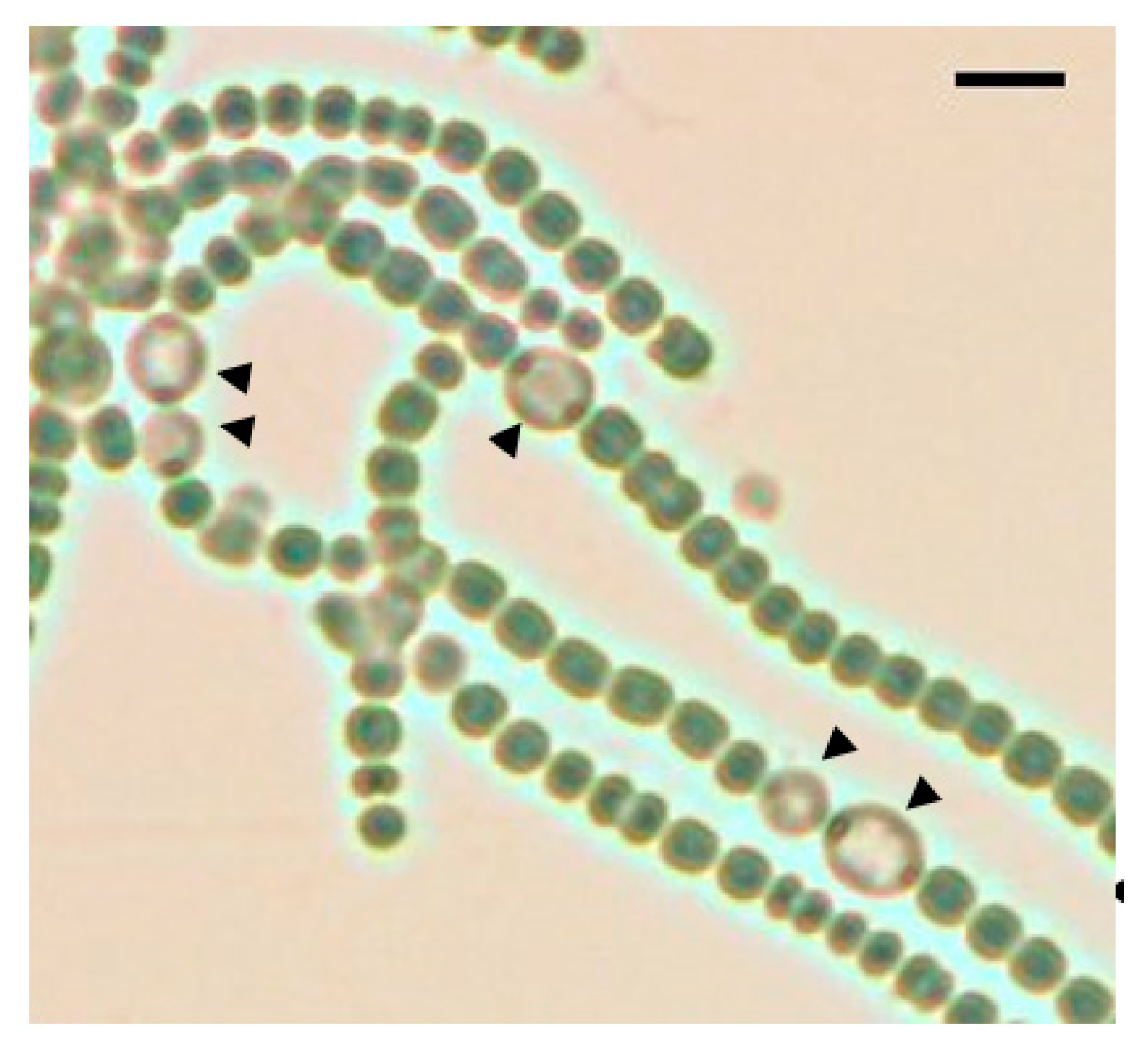
Figure 1.
A bright field image of the pknH mutant after 48 h of nitrogen deprivation. Black arrowheads indicate heterocysts. Scale bar, 10 μm.
Figure 1.
A bright field image of the pknH mutant after 48 h of nitrogen deprivation. Black arrowheads indicate heterocysts. Scale bar, 10 μm.
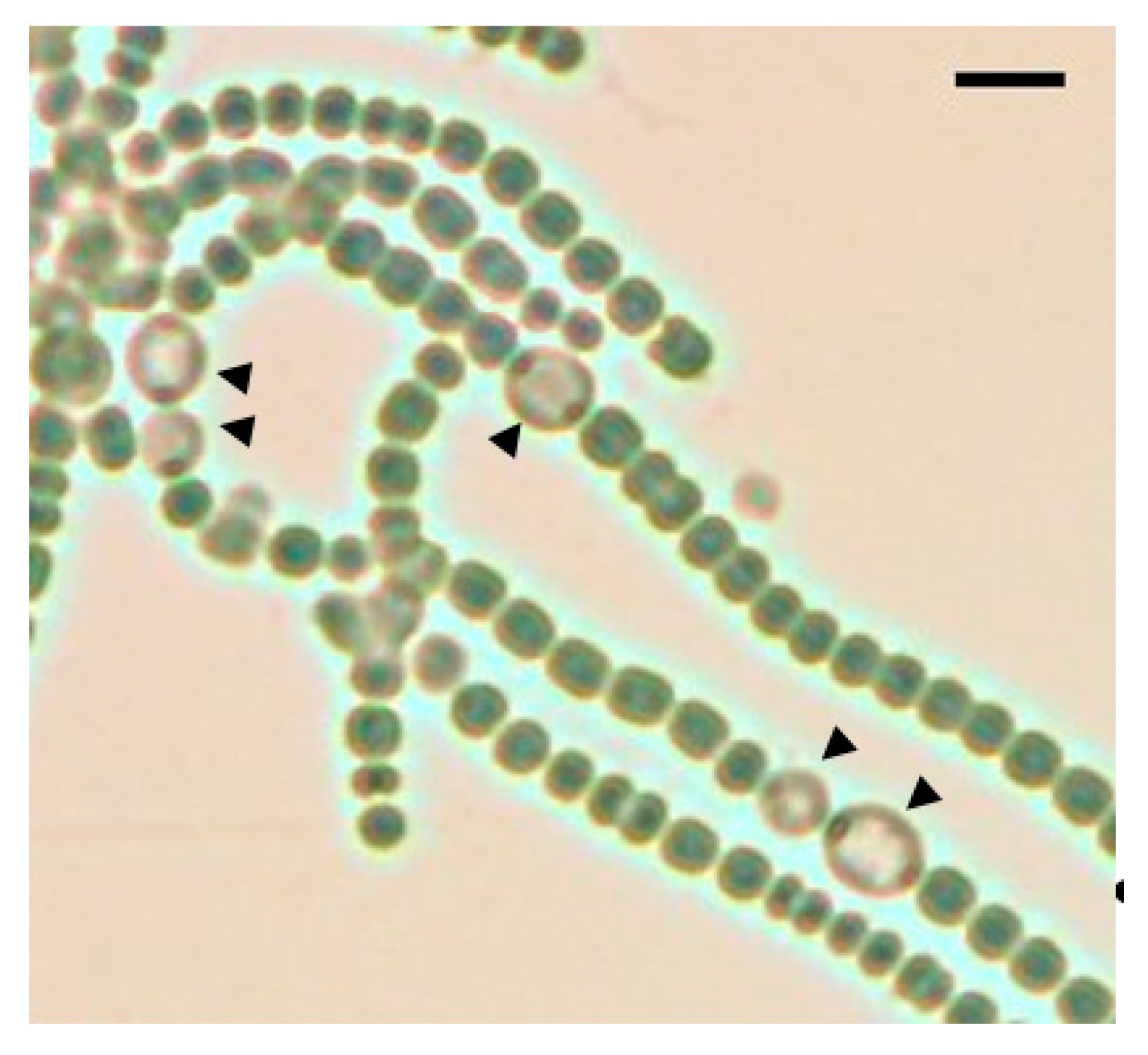
Figure 2.
Heterocyst distribution in the wild type, the pknH mutant (DR1336S), DR1336S complemented with pknH (DR1336S/pknH), and DR1336S complemented with the pknHD184N allele (DR1336S/pknHD184N). Filaments of each strain grown in the presence of nitrate were shifted to nitrogen-free medium and incubated for 24 or 48 h. More than 500 vegetative cells were counted for each sample.
Figure 2.
Heterocyst distribution in the wild type, the pknH mutant (DR1336S), DR1336S complemented with pknH (DR1336S/pknH), and DR1336S complemented with the pknHD184N allele (DR1336S/pknHD184N). Filaments of each strain grown in the presence of nitrate were shifted to nitrogen-free medium and incubated for 24 or 48 h. More than 500 vegetative cells were counted for each sample.
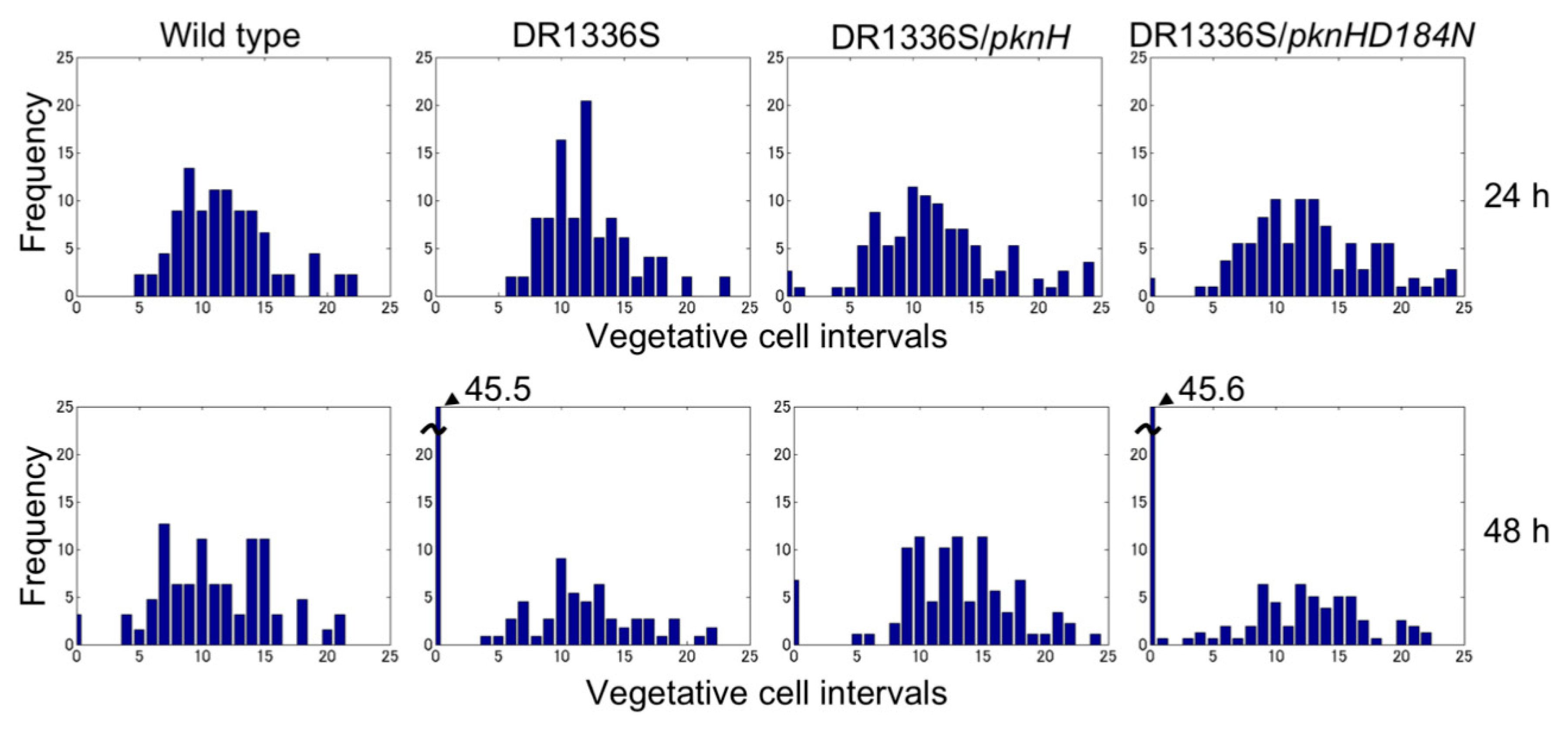
Figure 3.
Changes in the hetN transcript level after nitrogen deprivation. The relative transcript levels of hetN were determined by qRT-PCR in the wild-type (WT) strain (open circles) and the pknH mutant (closed circles). RNA samples were prepared from three independently grown cultures. The transcript level at 0 h of the WT strain was designated as 1.
Figure 3.
Changes in the hetN transcript level after nitrogen deprivation. The relative transcript levels of hetN were determined by qRT-PCR in the wild-type (WT) strain (open circles) and the pknH mutant (closed circles). RNA samples were prepared from three independently grown cultures. The transcript level at 0 h of the WT strain was designated as 1.

Figure 4.
Spatiotemporal expression of the hetR-gfp transcriptional fusion gene in the wild type. (A) Left panels in each column are green fluorescent protein (GFP) fluorescence images and right panels are corresponding auto-fluorescence images. Sequential micrographs were taken at 3 h intervals from 24 to 48 h after nitrogen deprivation. White arrows, cells with high GFP fluorescence at 24 h; black arrows, high GFP fluorescence cells located in the middle of a string of vegetative cells. Scale bar, 5 μm. (B) Changes in the GFP fluorescence levels of heterocysts (cell position 0) and adjacent vegetative cells (cell positions 1 to 5 and −1 to −5). The fluorescence intensity of a heterocyst and five vegetative cells located on each side of the heterocyst was measured. We analyzed 30 groups of cells and a typical cell profile, including the heterocyst marked with an asterisk in the 24 h image of Figure 4A, is indicated.
Figure 4.
Spatiotemporal expression of the hetR-gfp transcriptional fusion gene in the wild type. (A) Left panels in each column are green fluorescent protein (GFP) fluorescence images and right panels are corresponding auto-fluorescence images. Sequential micrographs were taken at 3 h intervals from 24 to 48 h after nitrogen deprivation. White arrows, cells with high GFP fluorescence at 24 h; black arrows, high GFP fluorescence cells located in the middle of a string of vegetative cells. Scale bar, 5 μm. (B) Changes in the GFP fluorescence levels of heterocysts (cell position 0) and adjacent vegetative cells (cell positions 1 to 5 and −1 to −5). The fluorescence intensity of a heterocyst and five vegetative cells located on each side of the heterocyst was measured. We analyzed 30 groups of cells and a typical cell profile, including the heterocyst marked with an asterisk in the 24 h image of Figure 4A, is indicated.
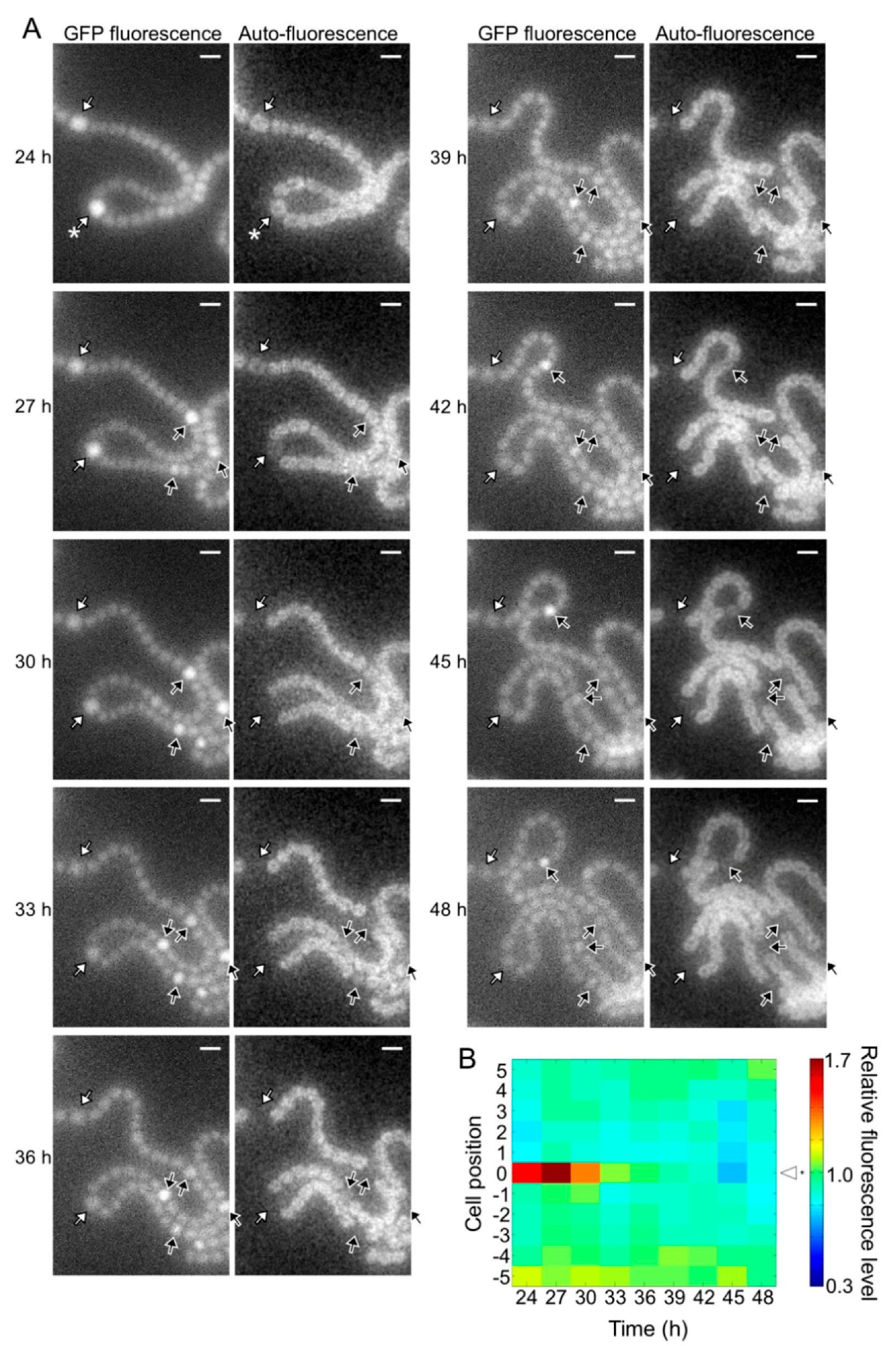
Figure 5.
Spatiotemporal expression of the hetR-gfp transcriptional fusion gene in the pknH mutant. (A) Left panels in each column are GFP fluorescence images and right panels are corresponding auto-fluorescence images. White arrows, cells with high GFP fluorescence at 24 h; yellow and red arrows, high GFP fluorescence cells adjacent to the preexisting heterocysts; black arrows, high GFP fluorescence cells located in the middle of a string of vegetative cells. Scale bar, 5 μm. (B) Changes in the GFP fluorescence levels of heterocysts (cell position 0) and adjacent vegetative cells (cell positions 1 to 5 and −1 to −5). The fluorescence intensity of a heterocyst and five vegetative cells located on each side of the heterocyst was measured. We analyzed 30 groups of cells and a typical cell profile, including the heterocyst marked with an asterisk in the 24 h image of Figure 5A, is indicated.
Figure 5.
Spatiotemporal expression of the hetR-gfp transcriptional fusion gene in the pknH mutant. (A) Left panels in each column are GFP fluorescence images and right panels are corresponding auto-fluorescence images. White arrows, cells with high GFP fluorescence at 24 h; yellow and red arrows, high GFP fluorescence cells adjacent to the preexisting heterocysts; black arrows, high GFP fluorescence cells located in the middle of a string of vegetative cells. Scale bar, 5 μm. (B) Changes in the GFP fluorescence levels of heterocysts (cell position 0) and adjacent vegetative cells (cell positions 1 to 5 and −1 to −5). The fluorescence intensity of a heterocyst and five vegetative cells located on each side of the heterocyst was measured. We analyzed 30 groups of cells and a typical cell profile, including the heterocyst marked with an asterisk in the 24 h image of Figure 5A, is indicated.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primers used in this study.
| Primer | Sequence (5′-3′) |
|---|---|
| 1336-5F | ATGAGCTCTTTACTGGTTGCCTGCTGTG |
| PknH-R | AAGTCGACGCATCACCGACACATAAAC |
| RTrrn16S-F2 | GCAAGTCGAACGGTCTCTTC |
| RTrrn16S-R2 | GGTATTAGCCACCGTTTCCA |
| RThetN-F | CATGATGGAACGCGGTAGTG |
| RThetN-R | AATTCCTGACGCATCGCATC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Fukushima, S.-i.; Ehira, S. The Ser/Thr Kinase PknH Is Essential for Maintaining Heterocyst Pattern in the Cyanobacterium Anabaena sp. Strain PCC 7120. Life 2018, 8, 34. https://doi.org/10.3390/life8030034
AMA Style
Fukushima S-i, Ehira S. The Ser/Thr Kinase PknH Is Essential for Maintaining Heterocyst Pattern in the Cyanobacterium Anabaena sp. Strain PCC 7120. Life. 2018; 8(3):34. https://doi.org/10.3390/life8030034
Chicago/Turabian StyleFukushima, Shun-ichi, and Shigeki Ehira. 2018. "The Ser/Thr Kinase PknH Is Essential for Maintaining Heterocyst Pattern in the Cyanobacterium Anabaena sp. Strain PCC 7120" Life 8, no. 3: 34. https://doi.org/10.3390/life8030034
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.