The Role of Mechanical Stimulation in Recovery of Bone Loss—High versus Low Magnitude and Frequency of Force

Abstract
:1. Introduction
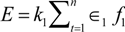
2. Mechanical Loading
2.1. Exercise: High Magnitude and Low Frequency
2.2. Muscle Contraction: Low Magnitude and High Frequency
3. Bone Loss Recovery Potential
4. Conclusions and Perspectives
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Turner, C.H. Three rules for bone adaptation to mechanical stimuli. Bone 1998, 23, 399–407. [Google Scholar] [CrossRef]
- Wolff, J. Das Gesetz der Transformation der Knochen; Verlag von August Hirschwald: Berlin, Germany, 1892. (In German) [Google Scholar]
- Frost, H.M. The Laws of Bone Structure; Charles C Thomas Publisher Ltd.: Springfield, IL, USA, 1964. [Google Scholar]
- Rubin, C.T.; Lanyon, L.E. Dynamic strain similarity in vertebrates; an alternative to allometric limb bone scaling. J Theor. Biol. 1984, 107, 321–327. [Google Scholar] [CrossRef]
- Marcus, R.; Feldman, D.; Kelsey, J.L. Osteoporosis, 2nd ed.; Academic Press: San Diego, CA, USA, 2001. [Google Scholar]
- Rubin, C.T.; Lanyon, L.E. Regulation of bone formation by applied dynamic loads. J. Bone Joint Surg. Am. 1984, 66, 397–402. [Google Scholar]
- Rubin, C.T.; Sommerfeldt, D.W.; Judex, S.; Qin, Y. Inhibition of osteopenia by low magnitude, high-frequency mechanical stimuli. Drug Discov. Today 2001, 6, 848–858. [Google Scholar] [CrossRef]
- Rubin, C.T.; Lanyon, L.E. Regulation of bone mass by mechanical strain magnitude. Calcif. Tissue Int. 1985, 37, 411–417. [Google Scholar] [CrossRef]
- Marcus, R. Exercise: Moving in the right direction. J. Bone Miner. Res. 1998, 13, 1793–1796. [Google Scholar] [CrossRef]
- Vuori, I. Exercise and physical health: Musculoskeletal health and functional capabilities. Res. Q. Exerc. Sport 1995, 66, 276–285. [Google Scholar] [CrossRef]
- Snow-Harter, C.; Bouxsein, M.L.; Lewis, B.T.; Carter, D.R.; Marcus, R. Effects of resistance and endurance exercise on bone mineral status of young women: A randomized exercise intervention trial. J. Bone Miner. Res. 1992, 7, 761–769. [Google Scholar]
- Galloway, M.T.; Jokl, P. Aging successfully: The importance of physical activity in maintaining health and function. J. Am. Acad. Orthop. Surg. 2000, 8, 37–44. [Google Scholar]
- Friedlander, A.L.; Genant, H.K.; Sadowsky, S.; Byl, N.N.; Gluer, C.C. A two-year program of aerobics and weight training enhances bone mineral density of young women. J. Bone Miner. Res. 1995, 10, 574–585. [Google Scholar]
- Kerr, D.; Morton, A.; Dick, I.; Prince, R. Exercise effects on bone mass in postmenopausal women are site-specific and load-dependent. J. Bone Miner. Res. 1996, 11, 218–225. [Google Scholar]
- Lohman, T.; Going, S.; Pamenter, R.; Hall, M.; Boyden, T.; Houtkooper, L.; Ritenbaugh, C.; Bare, L.; Hill, A.; Aickin, M. Effects of resistance training on regional and total bone mineral density in premenopausal women: A randomized prospective study. J. Bone Miner. Res. 1995, 10, 1015–1024. [Google Scholar]
- Bassey, E.J.; Rothwell, M.C.; Littlewood, J.J.; Pye, D.W. Pre- and postmenopausal women have different bone mineral density responses to the same high-impact exercise. J. Bone Miner. Res. 1998, 13, 1805–1813. [Google Scholar] [CrossRef]
- Shackelford, L.C.; LeBlanc, A.D.; Driscoll, T.B.; Evans, H.J.; Rianon, N.J.; Smith, S.M.; Spector, E.; Feeback, D.L.; Lai, D. Resistance exercise as a countermeasure to disuse-induced bone loss. J. Appl. Physiol. 2004, 97, 119–129. [Google Scholar] [CrossRef]
- Leblanc, A.; Matsumoto, T.; Jones, J.; Shapiro, J.; Lang, T.; Shackelford, L.; Smith, S.M.; Evans, H.; Spector, E.; Ploutz-Snyder, R.; et al. Bisphosphonates as a supplement to exercise to protect bone during long-duration spaceflight. Osteoporos. Int. 2013, 24, 2105–2114. [Google Scholar] [CrossRef]
- Smith, S.M.; Heer, M.A.; Shackelford, L.C.; Sibonga, J.D.; Ploutz-Snyder, L.; Zwart, S.R. Benefits for bone from resistance exercise and nutrition in long-duration spaceflight: Evidence from biochemistry and densitometry. J. Bone Miner. Res. 2012, 27, 1896–1906. [Google Scholar] [CrossRef]
- Sibonga, J.D.; Iwaniec, U.T.; Shogren, K.L.; Rosen, C.J.; Turner, R.T. Effects of parathyroid hormone (1–34) on tibia in an adult rat model for chronic alcohol abuse. Bone 2007, 40, 1013–1020. [Google Scholar] [CrossRef]
- Robinson, T.L.; Snow-Harter, C.; Taaffe, D.R.; Gillis, D.; Shaw, J.; Marcus, R. Gymnasts exhibit higher bone mass than runners despite similar prevalence of amenorrhea and oligomenorrhea. J. Bone Miner. Res. 1995, 10, 26–35. [Google Scholar]
- Haapasalo, H.; Sievanen, H.; Kannus, P.; Heinonen, A.; Oja, P.; Vuori, I. Dimensions and estimated mechanical characteristics of the humerus after long-term tennis loading. J. Bone Miner. Res. 1996, 11, 864–872. [Google Scholar]
- Bradney, M.; Pearce, G.; Naughton, G.; Sullivan, C.; Bass, S.; Beck, T.; Carlson, J.; Seeman, E. Moderate exercise during growth in prepubertal boys: Changes in bone mass, size, volumetric density, and bone strength: A controlled prospective study. J. Bone Miner. Res. 1998, 13, 1814–1821. [Google Scholar] [CrossRef]
- Rubin, C.; Turner, A.S.; Bain, S.; Mallinckrodt, C.; McLeod, K. Anabolism. Low mechanical signals strengthen long bones. Nature 2001, 412, 603–604. [Google Scholar]
- Gray, B.; Hsu, J.D.; Furumasu, J. Fractures caused by falling from a wheelchair in patients with neuromuscular disease. Dev. Med. Child Neurol. 1992, 34, 589–592. [Google Scholar] [CrossRef]
- Ward, K.; Alsop, C.; Caulton, J.; Rubin, C.; Adams, J.; Mughal, Z. Low magnitude mechanical loading is osteogenic in children with disabling conditions. J. Bone Miner. Res. 2004, 19, 360–369. [Google Scholar] [CrossRef]
- Asselin, P.; Spungen, A.M.; Muir, J.W.; Rubin, C.T.; Bauman, W.A. Transmission of low-intensity vibration through the axial skeleton of persons with spinal cord injury as a potential intervention for preservation of bone quantity and quality. J. Spinal Cord Med. 2011, 34, 52–59. [Google Scholar] [CrossRef]
- Kiel, D.P.; Hannan, M.T.; Barton, B.A.; Bouxsein, M.L.; Lang, T.F.; Brown, K.M.; Shane, E.; Magaziner, J.; Zimmerman, S.; Rubin, C.T. Insights from the conduct of a device trial in older persons: Low magnitude mechanical stimulation for musculoskeletal health. Clin. Trials 2010, 7, 354–367. [Google Scholar] [CrossRef]
- Rubin, C.; Recker, R.; Cullen, D.; Ryaby, J.; McCabe, J.; McLeod, K. Pevention of postmenopausal bone loss by a low-magnitude, high-frequency mechanical stimuli: A clinical trial assessing compliance; efficacy; and safety. J. Bone Miner. Res. 2004, 19, 343–351. [Google Scholar] [CrossRef]
- Rubin, C.; Turner, A.S.; Mallinckrodt, C.; Jerome, C.; McLeod, K.; Bain, S. Mechanical strain, induced noninvasively in the high-frequency domain, is anabolic to cancellous bone, but not cortical bone. Bone 2002, 30, 445–452. [Google Scholar] [CrossRef]
- Rubin, C.; Turner, A.S.; Muller, R.; Mittra, E.; McLeod, K.; Lin, W.; Qin, Y.X. Quantity and quality of trabecular bone in the femur are enhanced by a strongly anabolic, noninvasive mechanical intervention. J. Bone Miner. Res. 2002, 17, 349–357. [Google Scholar] [CrossRef]
- Rubin, C.; Xu, G.; Judex, S. The anabolic activity of bone tissue, suppressed by disuse, is normalized by brief exposure to extremely low-magnitude mechanical stimuli. FASEB J. 2001, 15, 2225–2229. [Google Scholar] [CrossRef]
- Judex, S.; Zhong, N.; Squire, M.E.; Ye, K.; Donahue, L.R.; Hadjiargyrou, M.; Rubin, C.T. Mechanical modulation of molecular signals which regulate anabolic and catabolic activity in bone tissue. J. Cell. Biochem. 2005, 94, 982–994. [Google Scholar] [CrossRef]
- Chan, M.E.; Adler, B.J.; Green, D.E.; Rubin, C.T. Bone structure and B-cell populations, crippled by obesity, are partially rescued by brief daily exposure to low-magnitude mechanical signals. FASEB J. 2012, 26, 4855–4863. [Google Scholar] [CrossRef]
- Fritton, S.P.; McLeod, K.J.; Rubin, C.T. Quantifying the strain history of bone: Spatial uniformity and self-similarity of low-magnitude strains. J. Biomech. 2000, 33, 317–325. [Google Scholar] [CrossRef]
- Rubin, C.T.; Lanyon, L.E. Limb mechanics as a function of speed and gait: A study of functional strains in the radius and tibia of horse and dog. J. Exp. Biol. 1982, 101, 187–211. [Google Scholar]
- Lanyon, L.E.; Smith, R.N. Bone strain in the tibia during normal quadrupedal locomotion. Acta Orthop. Scand. 1970, 41, 238–248. [Google Scholar] [CrossRef]
- Carter, D.R.; Smith, D.J.; Spengler, D.M.; Daly, C.H.; Frankel, V.H. Measurement and analysis of in vivo bone strains on the canine radius and ulna. J. Biomech. 1980, 13, 27–38. [Google Scholar] [CrossRef]
- Adams, D.J.; Spirt, A.A.; Brown, T.D.; Fritton, S.P.; Rubin, C.T.; Brand, R.A. Testing the daily stress stimulus theory of bone adaptation with natural and experimentally controlled strain histories. J. Biomech. 1997, 30, 671–678. [Google Scholar] [CrossRef]
- Issekutz, B., Jr.; Blizzard, J.J.; Birkhead, N.C.; Rodahl, K. Effect of prolonged bed rest on urinary calcium output. J. Appl. Physiol. 1966, 21, 1013–1020. [Google Scholar]
- Xie, L.; Jacobson, J.M.; Choi, E.S.; Busa, B.; Donahue, L.R.; Miller, L.M.; Rubin, C.T.; Judex, S. Low-level mechanical vibrations can influence bone resorption and bone formation in the growing skeleton. Bone 2006, 39, 1059–1066. [Google Scholar] [CrossRef]
- Garman, R.; Gaudette, G.; Donahue, L.R.; Rubin, C.; Judex, S. Low-level accelerations applied in the absence of weight bearing can enhance trabecular bone formation. J. Orthop. Res. 2007, 25, 732–740. [Google Scholar] [CrossRef]
- Gilsanz, V.; Wren, T.A.; Sanchez, M.; Dorey, F.; Judex, S.; Rubin, C. Low-level, high-frequency mechanical signals enhance musculoskeletal development of young women with low BMD. J. Bone Miner. Res. 2006, 21, 1464–1474. [Google Scholar] [CrossRef]
- Patel, M.J.; Chang, K.H.; Sykes, M.C.; Talish, R.; Rubin, C.; Jo, H. Low magnitude and high frequency mechanical loading prevents decreased bone formation responses of 2T3 preosteoblasts. J. Cell. Biochem. 2009, 106, 306–316. [Google Scholar] [CrossRef]
- Murfee, W.L.; Hammett, L.A.; Evans, C.; Xie, L.; Squire, M.; Rubin, C.; Judex, S.; Skalak, T.C. High-frequency, low-magnitude vibrations suppress the number of blood vessels per muscle fiber in mouse soleus muscle. J. Appl. Physiol. 2005, 98, 2376–2380. [Google Scholar] [CrossRef]
- Lau, E.; Al-Dujaili, S.; Guenther, A.; Liu, D.; Wang, L.; You, L. Effect of low-magnitude, high-frequency vibration on osteocytes in the regulation of osteoclasts. Bone 2010, 46, 1508–1515. [Google Scholar] [CrossRef]
- Uzer, G.; Manske, S.L.; Chan, M.E.; Chiang, F.P.; Rubin, C.T.; Frame, M.D.; Judex, S. Separating Fluid Shear Stress from Acceleration during Vibrations in Vitro: Identification of Mechanical Signals Modulating the Cellular Response. Cell. Mol. Bioeng. 2012, 5, 266–276. [Google Scholar]
- Uzer, G.; Pongkitwitoon, S.; Ete Chan, M.; Judex, S. Vibration induced osteogenic commitment of mesenchymal stem cells is enhanced by cytoskeletal remodeling but not fluid shear. J. Biomech. 2013, 46, 2296–2302. [Google Scholar] [CrossRef]
- Chan, M.E.; Uzer, G.; Rubin, C.T. The potential benefits and inherent risks of vibration as a non-drug therapy for the prevention and treatment of osteoporosis. Curr. Osteoporos. Rep. 2013, 11, 36–44. [Google Scholar] [CrossRef]
- Huijser, R.H. Desktop RPM: New Small Size Microgravity Simulator for the Bioscience Laboratory; Fokker Space: Leiden, The Netherlands, 2000; pp. 1–5. Available online: http://www.descsite.nl/Publications/Other/RPM-FS-MG-R00-017.pdf (accessed on 19 February 2014).
- Pardo, S.J.; Patel, M.J.; Sykes, M.C.; Platt, M.O.; Boyd, N.L.; Sorescu, G.P.; Xu, M.; van Loon, J.J.W.A.; Wang, M.D.; Jo, H. Simulated microgravity using the Random Positioning Machine inhibits differentiation and alters gene expression profiles of 2T3 preosteoblasts. Am. J. Physiol. Cell Physiol. 2005, 288, C1211–C1221. [Google Scholar] [CrossRef]
- Lian, J.B.; Stein, G.S. Development of the osteoblast phenotype: Molecular mechanisms mediating osteoblast growth and differentiation. Iowa Orthop. J. 1995, 15, 118–140. [Google Scholar]
- Bacabac, R.G.; Smit, T.H.; van Loon, J.J.; Doulabi, B.Z.; Helder, M.; Klein-Nulend, J. Bone cell responses to high-frequency vibration stress: Does the nucleus oscillate within the cytoplasm? FASEB J. 2006, 20, 858–864. [Google Scholar] [CrossRef]
- Holguin, N.; Muir, J.; Rubin, C.; Judex, S. Short applications of very low-magnitude vibrations attenuate expansion of the intervertebral disc during extended bed rest. Spine J. 2009, 9, 470–477. [Google Scholar] [CrossRef]
- Holguin, N.; Uzer, G.; Chiang, F.P.; Rubin, C.; Judex, S. Brief daily exposure to low-intensity vibration mitigates the degradation of the intervertebral disc in a frequency-specific manner. J. Appl. Physiol. 2011, 111, 1846–1853. [Google Scholar] [CrossRef]
- Iqbal, J.; Zaidi, M. Molecular regulation of mechanotransduction. Biochem. Biophys. Res. Commun. 2005, 328, 751–755. [Google Scholar] [CrossRef]
- Ingber, D. How cells (might) sense microgravity. FASEB J. 1999, 13, S3–S15. [Google Scholar]
- Ingber, D.E. Cellular mechanotransduction: Putting all the pieces together again. FASEB J. 2006, 20, 811–827. [Google Scholar] [CrossRef]
- Thompson, W.R.; Rubin, C.T.; Rubin, J. Mechanical regulation of signaling pathways in bone. Gene 2012, 503, 179–193. [Google Scholar] [CrossRef]
- Ozcivici, E.; Luu, Y.K.; Adler, B.; Qin, Y.X.; Rubin, J.; Judex, S.; Rubin, C.T. Mechanical signals as anabolic agents in bone. Nat. Rev. Rheumatol. 2010, 6, 50–59. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Nagaraja, M.P.; Jo, H. The Role of Mechanical Stimulation in Recovery of Bone Loss—High versus Low Magnitude and Frequency of Force. Life 2014, 4, 117-130. https://doi.org/10.3390/life4020117
Nagaraja MP, Jo H. The Role of Mechanical Stimulation in Recovery of Bone Loss—High versus Low Magnitude and Frequency of Force. Life. 2014; 4(2):117-130. https://doi.org/10.3390/life4020117
Chicago/Turabian StyleNagaraja, Mamta Patel, and Hanjoong Jo. 2014. "The Role of Mechanical Stimulation in Recovery of Bone Loss—High versus Low Magnitude and Frequency of Force" Life 4, no. 2: 117-130. https://doi.org/10.3390/life4020117
APA StyleNagaraja, M. P., & Jo, H. (2014). The Role of Mechanical Stimulation in Recovery of Bone Loss—High versus Low Magnitude and Frequency of Force. Life, 4(2), 117-130. https://doi.org/10.3390/life4020117