Historical Landscape Perspectives on Grasslands in Sweden and the Baltic Region

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

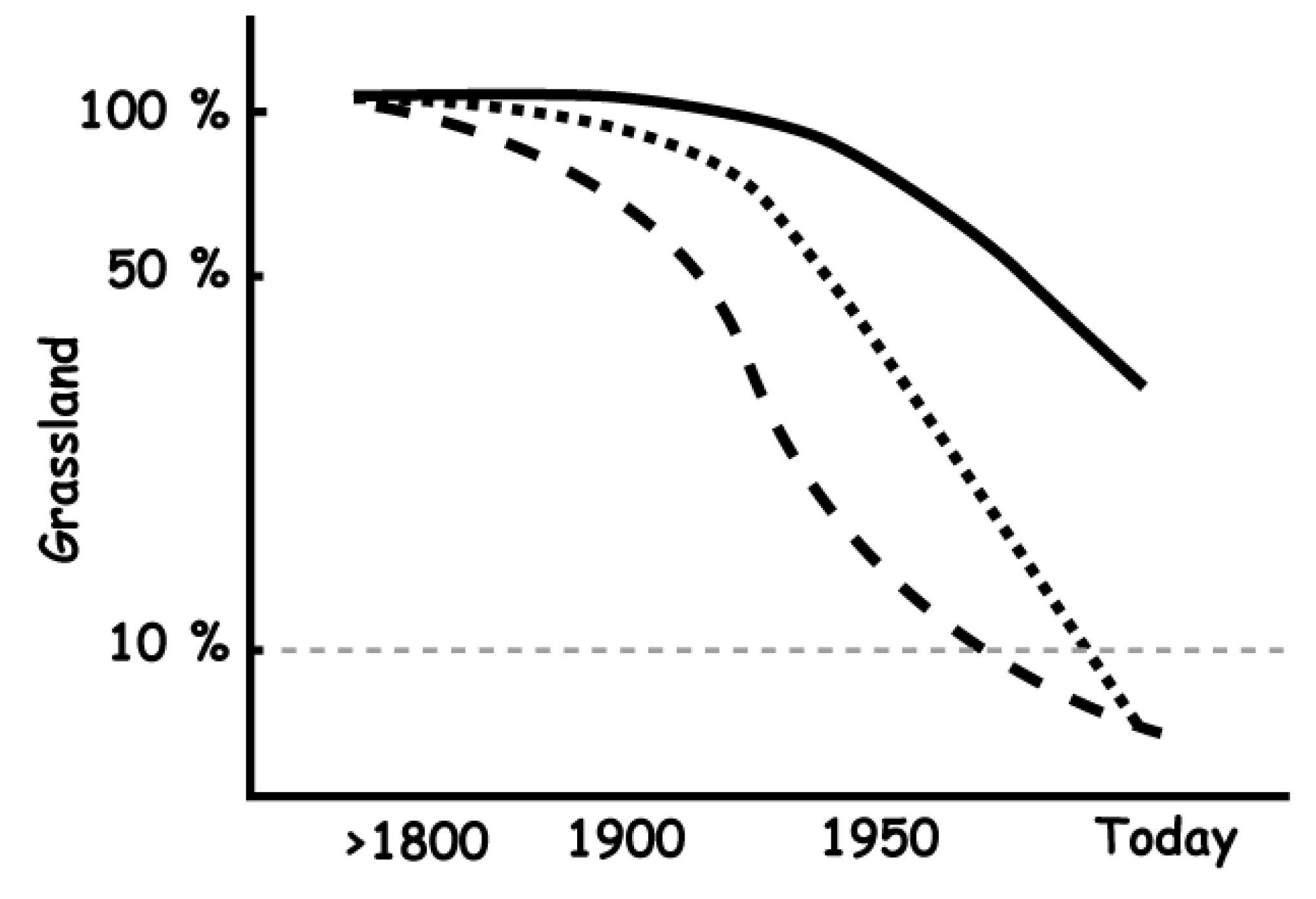
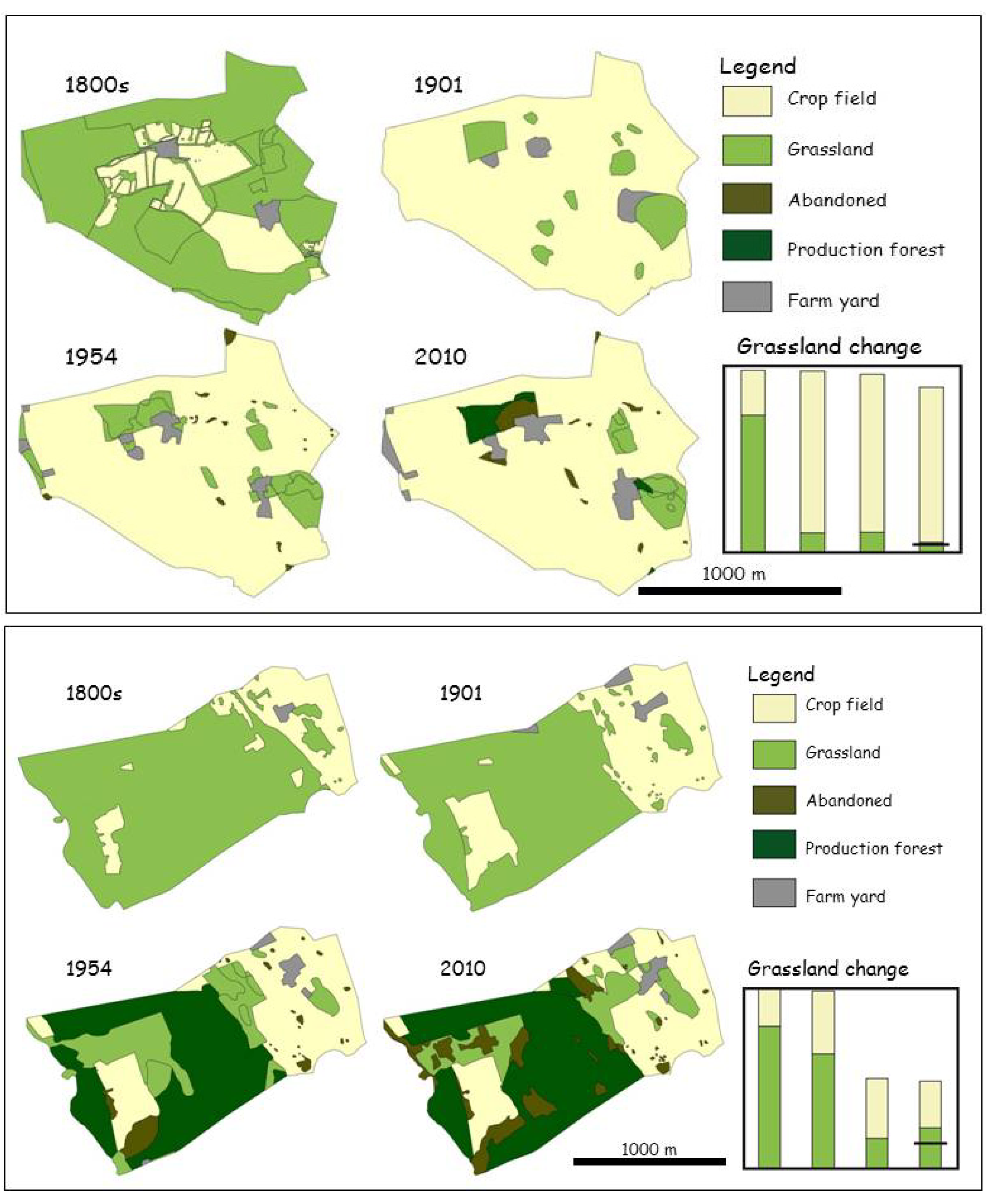
2. The Historical Context
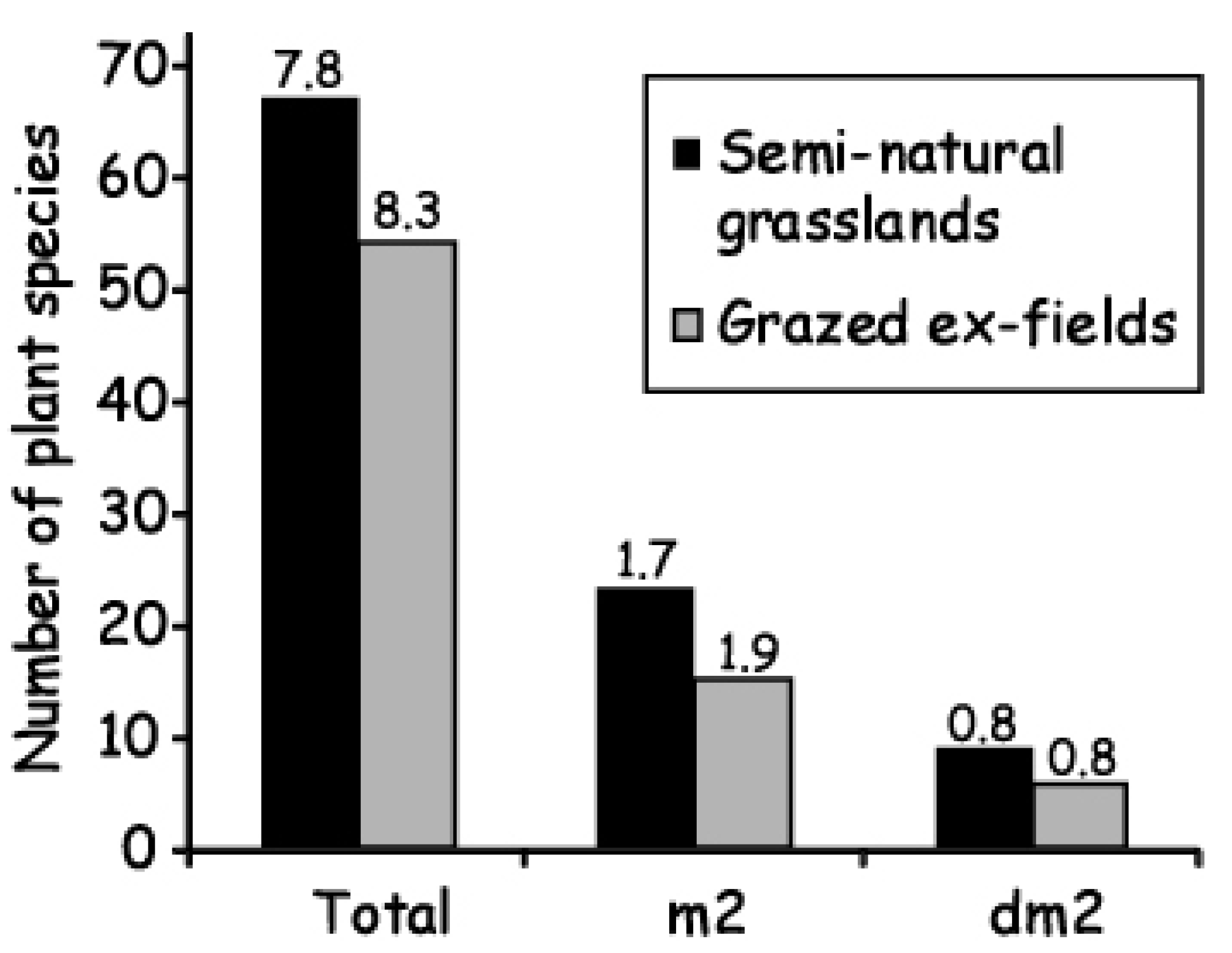
3. Biodiversity in Semi-Natural Grasslands

4. Effects of Delayed Species Response
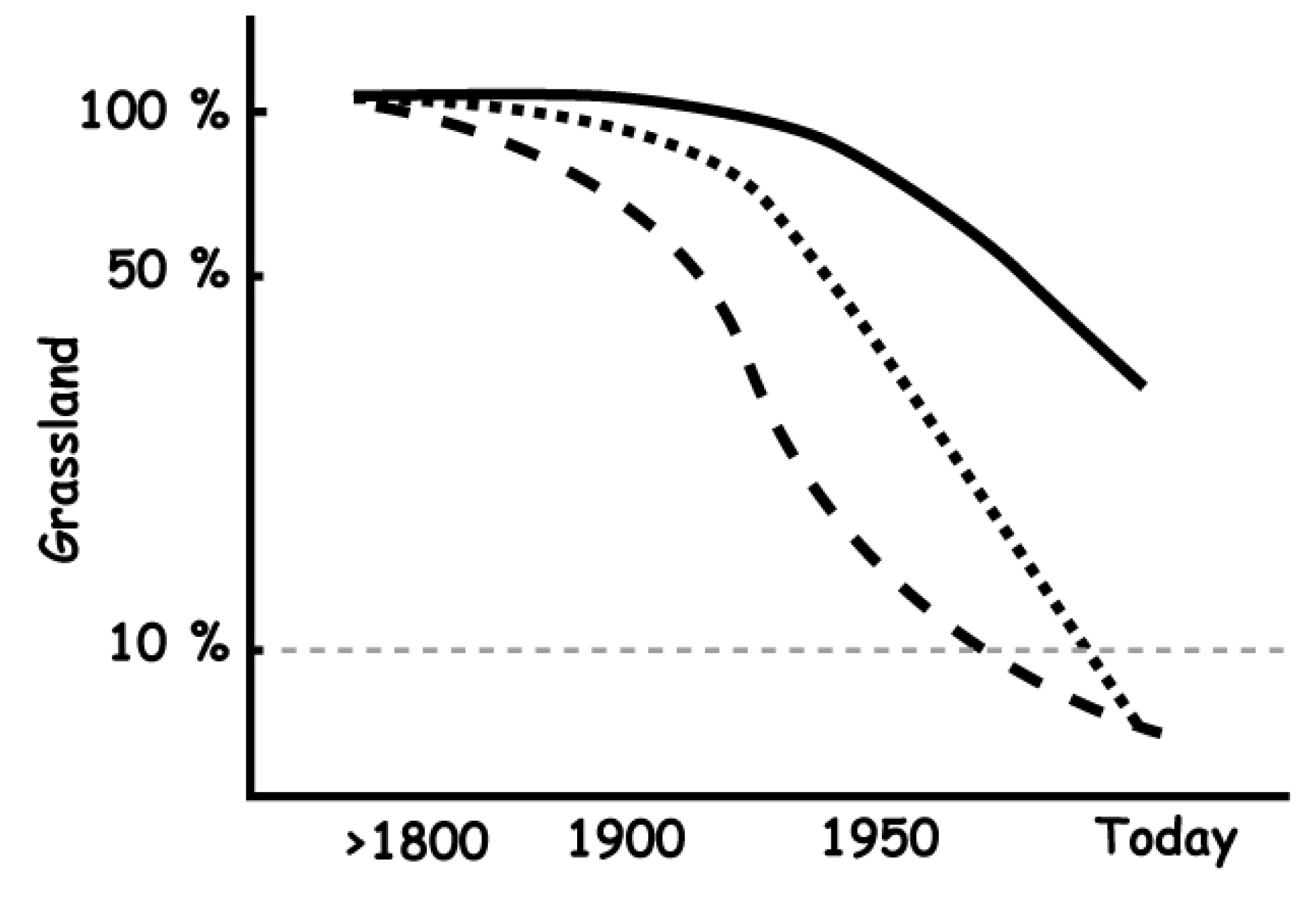
5. Perception and Valuation of Semi-Natural Grasslands
6. Conservation Models for Semi-Natural Grasslands
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lindborg, R.; Bengtsson, J.; Berg, Å.; Cousins, S.A.O.; Eriksson, O.; Gustafsson, T.; Hasund, K.P.; Lenoir, L.; Pihlgren, A.; Sjödin, E.; et al. A landscape perspective on conservation of semi-natural grasslands. Agric. Ecosyst. Environ. 2008, 125, 213–222. [Google Scholar] [CrossRef]
- Tscharntke, T.; Tylianakis, J.M.; Rand, T.A.; Didham, R.K.; Fahrig, L.; Batáry, P.; Bengtsson, J.; Clough, Y.; Crist, T.O.; Dormann, C.F.; et al. Landscape moderation of biodiversity patterns and processes—Eight hypotheses. Biol. Rev. 2012, 87, 661–685. [Google Scholar] [CrossRef]
- Driscoll, D.A.; Banks, S.C.; Barton, P.S.; Lindemayer, D.B.; Smith, A.L. Conceptual domain of the matrix in fragmented landscapes. Trends Ecol. Evol. 2013, 28, 605–613. [Google Scholar] [CrossRef]
- Schama, S. Landscape and Memory; Vintage Books: New York, NY, USA, 1995. [Google Scholar]
- Antrop, M. Why landscapes of the past are important for the future. Landsc. Urban Plan. 2005, 70, 21–34. [Google Scholar] [CrossRef]
- Palang, H.; Spek, T.; Stenseke, M. Digging in the past: New conceptual models in landscape history and their relevance in peri-urban landscapes. Landsc. Urban Plan. 2011, 100, 344–346. [Google Scholar]
- Balée, W. The research program of historical ecology. Annu. Rev. Anthropol. 2006, 35, 75–98. [Google Scholar] [CrossRef]
- Ellis, E.C.; Ramankutty, N. Putting people in the map: Anthropogenic biomes of the world. Front. Ecol. Environ. 2008, 6, 439–447. [Google Scholar] [CrossRef]
- Foster, D.R. Thoreau’s country: A historical-ecological perspective on conservation in the New England landscape. J. Biogeogr. 2002, 29, 1537–1555. [Google Scholar] [CrossRef]
- Foster, D.R.; Swanson, F.; Aber, J.; Burke, I.; Brokaw, N.; Tilman, D.; Knapp, A. The importance of land-use legacies to ecology and conservation. Bioscience 2003, 53, 77–88. [Google Scholar] [CrossRef]
- Szabó, P. Why history matters in ecology: An interdisciplinary perspective. Environ. Conserv. 2010, 37, 380–387. [Google Scholar] [CrossRef]
- Vellend, M.; Brown, C.D.; Kharouba, H.M.; McCune, J.L.; Myers-Smith, I.H. Historical ecology: Using unconventional data sources to test for effects of global environmental change. Am. J. Bot. 2013, 100, 1294–1305. [Google Scholar] [CrossRef]
- Cousins, S.A.O.; Eriksson, O. The influence of management history and habitat on plant species richness in a rural hemiboreal landscape, Sweden. Landsc. Ecol. 2002, 17, 517–529. [Google Scholar] [CrossRef]
- Lindborg, R.; Eriksson, O. Historical landscape connectivity affects present plant species diversity. Ecology 2004, 85, 1840–1845. [Google Scholar] [CrossRef]
- Helm, A.; Hanski, I.; Pärtel, M. Slow response of plant species richness to habitat loss and fragmentation. Ecol. Lett. 2006, 9, 72–77. [Google Scholar]
- Willis, K.J.; Gillson, L.; Brncic, T.M. How “virgin” is virgin rainforest? Science 2004, 304, 402–403. [Google Scholar] [CrossRef]
- Josefsson, T.; Bergman, I.; Östlund, L. Quantifying Sami settlement and movement patterns in northern Sweden 1700–1900. Arctic 2010, 63, 141–154. [Google Scholar]
- Millennium Ecosystem Assessment. Ecosystems and Human Well-being: A Framework for Assessment; Island Press: Washington, DC, USA, 2003. [Google Scholar]
- Emanuelsson, U. The Rural Landscape of Europe. How Man has Shaped European Nature; The Swedish Research Council Formas: Stockholm, Sweden, 2009. [Google Scholar]
- Storm, A. Hope and Rust: Reinterpreting the Industrial Place in the Late 20th Century; Stockholm Papers in the History and Philosophy of Technology; Royal Institute of Technology KTH: Stockholm, Sweden, 2008. [Google Scholar]
- Riede, F. Adaptation and niche construction in human prehistory: A case study from the southern Scandinavian late glacial. Philos. Trans. R. Soc. B 2011, 366, 793–808. [Google Scholar] [CrossRef]
- Svenning, J.-C. A review of natural vegetation openness in north-western Europe. Biol. Conserv. 2002, 104, 133–148. [Google Scholar] [CrossRef]
- Lambeck, K. Shoreline displacement in southern-central Sweden and the evolution of the Baltic Sea since the last maximum glaciation. J. Geol. Soc. 1999, 156, 465–486. [Google Scholar] [CrossRef]
- Brandt, G.; Haak, W.; Adler, C.J.; Roth, C.; Szécsényi-Nagy, A.; Karimnia, S.; Möller-Rieker, S.; Meller, H.; Ganslmeier, R.; Friederich, S.; et al. Ancient DNA reveals key stages in the formation of central European mitochondrial genetic diversity. Science 2013, 342, 257–261. [Google Scholar] [CrossRef]
- Odgaard, B.V.; Rasmussen, P. Origin and temporal development of macro-scale vegetation patterns in the cultural landscape of Denmark. J. Ecol. 2000, 88, 733–748. [Google Scholar] [CrossRef]
- Welinder, S. Early farming households. In The Agrarian History of Sweden from 4000 BC to AD 2000; Myrdal, J., Morell, M., Eds.; Nordic Academic Press: Lund, Sweden, 2011; pp. 18–45. [Google Scholar]
- The Cultural Landscape during 6000 Years in Southern Sweden: The Ystad Project; Ecological Bulletins 41; Berglund, B.E. (Ed.) Munksgaard International Booksellers and Publishers: Copenhagen, Denmark, 1991.
- Nielsen, A.B.; Giesecke, T.; Theurekauf, M.; Feeser, I.; Behre, K.-E.; Beug, H.-J.; Chen, S.-H.; Christiansen, J.; Dörfler, W.; Endtmann, E.; et al. Quantitative reconstruction of changes in regional openness in North-Central Europe reveal new insights into old questions. Quat. Sci. Rev. 2012, 47, 131–149. [Google Scholar] [CrossRef]
- Hannon, G.E.; Bradshaw, R.H.W.; Nord, J.; Gustafsson, M. The Bronze Age landscape of the Bjäre peninsula, southern Sweden, and its relationship to burial mounds. J. Archaeol. Sci. 2008, 35, 623–632. [Google Scholar] [CrossRef]
- Bradshaw, E.G.; Rasmussen, P.; Odgaard, B.V. Mid- to late Holocene land-use change and lake development at Dallund Sø, Denmark: Synthesis of multiproxy data, linking land and lake. Holocene 2005, 15, 1152–1162. [Google Scholar] [CrossRef]
- Kaplan, J.O.; Krumhardt, K.M.; Zimmermann, N. The prehistoric and preindustrial deforestation of Europe. Quat. Sci. Rev. 2009, 28, 3016–3034. [Google Scholar] [CrossRef]
- Pedersen, E.A.; Widgren, M. Agriculture in Sweden, 800 BC–AD 1000. In The Agrarian History of Sweden from 4000 BC to AD 2000; Myrdal, J., Morell, M., Eds.; Nordic Academic Press: Lund, Sweden, 2011; pp. 46–71. [Google Scholar]
- Overland, A.; Hjelle, K.L. From forest to open pastures and fields: Cultural landscape development in western Norway inferred from two pollen records representing different spatial scales of vegetation. Veg. Hist. Archaeobot. 2009, 18, 459–476. [Google Scholar] [CrossRef]
- Poska, A.; Saarse, L.; Veski, S. Reflections of pre- and early-agrarian human impact in the pollen diagrams of Estonia. Palaeogeogr. Palaeoclim. Palaeoecol. 2004, 209, 37–50. [Google Scholar] [CrossRef]
- Reitalu, T.; Seppä, H.; Sugita, S.; Kangur, M.; Koff, T.; Avel, E.; Kihno, K.; Vassiljev, J.; Renssen, H.; Hammarlund, D.; et al. Long-term drivers of forest composition in a boreonemoral region: The relative importance of climate and human impact. J. Biogeogr. 2013, 40, 1524–1534. [Google Scholar] [CrossRef]
- Sköld, E.; Lagerås, P.; Berglund, B.E. Temporal cultural landscape dynamics in a marginal upland area: Agricultural expansions and contractions inferred from palynological evidence at Yttra Berg, southern Sweden. Veg. Hist. Archaeobot. 2010, 19, 121–136. [Google Scholar] [CrossRef]
- Alenius, T.; Mikkola, E.; Ojala, A.E.K. History of agriculture in Mikkeli Orijärvi, eastern Finland as reflected by palynological and archaeological data. Veg. Hist. Archaeobot. 2008, 17, 171–183. [Google Scholar] [CrossRef]
- Alenius, T.; Saarnosto, M.; Taavitsainen, J.-P.; Lunkka, J.-P. Records of land use and medieval settlement history in Vuonninen, Dvina Karelia, Northwest Russia. Geoarchaeology 2011, 26, 142–163. [Google Scholar] [CrossRef]
- Alenius, T.; Mökkönen, T.; Lahelma, A. Early farming in the northern boreal zone: Reassessing the history of land use in southeastern Finland through high-resolution pollen analysis. Geoarchaeology 2013, 28, 1–24. [Google Scholar] [CrossRef]
- Karlsson, H.; Emanuelsson, M.; Segerström, U. The history of a farm-shieling system in the Central Swedish forest region. Veg. Hist. Archaeobot. 2010, 19, 103–119. [Google Scholar] [CrossRef]
- Josefsson, T.; Gunnarsson, B.; Liedgren, L.; Bergman, I.; Östlund, L. Historical human influence on forest composition and structure in boreal Fennoscandia. Can. J. For. Res. 2010, 40, 872–884. [Google Scholar] [CrossRef]
- Austheim, G.; Eriksson, O. Plant species diversity and grazing in the Scandinavian mountains: Patterns and processes at different spatial scales. Ecography 2001, 24, 683–695. [Google Scholar]
- Widgren, M. Settlement and Farming Systems in the Early Iron Age: A Study of Fossil Agrarian Landscapes in Östergötland, Sweden. Stockholm Studies in Human Geography 3; Almquist & Wiksell: Stockholm, Sweden, 1983. [Google Scholar]
- Lagerås, P. The Ecology of Expansion and Abandonment: Medieval and Post-Medieval Land-Use and Settlement Dynamics in a Landscape Perspective; National Heritage Board: Stockholm, Sweden, 2007. [Google Scholar]
- Nielsen, A.B.; Odgaard, B.V. Quantitative landscape dynamics in Denmark through the last three millenia based on the landscape reconstruction algorithm approach. Veg. Hist. Archaeobot. 2010, 19, 375–387. [Google Scholar] [CrossRef]
- Gadd, C.-J. The agricultural revolution in Sweden, 1700–1870. In The Agrarian History of Sweden from 4000 BC to AD 2000; Myrdal, J., Morell, M., Eds.; Nordic Academic Press: Lund, Sweden, 2011; pp. 118–164. [Google Scholar]
- Cousins, S.A.O. Analysis of land-cover transitions based on 17th and 18th century cadastral maps and aerial photographs. Landsc. Ecol. 2001, 16, 41–54. [Google Scholar] [CrossRef]
- Flygare, I.A.; Isacson, M. The tension between modernity and reality. In The Agrarian History of Sweden from 4000 BC to AD 2000; Myrdal, J., Morell, M., Eds.; Nordic Academic Press: Lund, Sweden, 2011; pp. 214–256. [Google Scholar]
- Cousins, S.A.O. Landscape history and soil properties affect grassland decline and plant species richness in rural landscapes. Biol. Conserv. 2009, 142, 2752–2758. [Google Scholar] [CrossRef]
- Dahlström, A.; Cousins, S.A.O.; Eriksson, O. The history (1620–2003) of land use, people and livestock, and the relationships to present plant species diversity in a rural landscape in Sweden. Environ. Hist. 2006, 12, 191–212. [Google Scholar] [CrossRef]
- Cousins, S.A.O.; Eriksson, O. After the hotspots are gone: Land use history and grassland plant species diversity in a strongly transformed agricultural landscape. Appl. Veg. Sci. 2008, 11, 365–374. [Google Scholar] [CrossRef]
- Luoto, M.; Rekolainen, S.; Aakkula, J.; Pykälä, J. Loss of plant species richness and habitat connectivity in grasslands associated with agricultural change in Finland. AMBIO 2003, 32, 447–452. [Google Scholar]
- Pärtel, M.; Mändla, R.; Zobel, M. Landscape history of a calcareous (alvar) grassland in Hanila, western Estonia, during the last three hundred years. Landsc. Ecol. 1999, 14, 187–196. [Google Scholar] [CrossRef]
- Palmgren, E. Distribution of Semi-Natural Pastures in Sweden: A Comparison of Coverage Estimation Using Random Sampling and Total Registration Data Sets. M.Sc. Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2010. [Google Scholar]
- Vessby, K.; Söderström, B.; Glimskär, A.; Svensson, B. Species-richness correlations of six different taxa in Swedish semi-natural grasslands. Conserv. Biol. 2002, 16, 430–439. [Google Scholar] [CrossRef]
- The 2010 Red List of Swedish Species; Gärdenfors, U. (Ed.) ArtDatabanken, SLU: Uppsala, Sweden, 2010.
- Cousins, S.A.O.; Eriksson, O. Plant species occurrences in a rural hemiboreal landscape: Effects of remnant habitats, site history, topography and soil. Ecography 2001, 24, 461–469. [Google Scholar] [CrossRef]
- Gustavsson, E.; Lennartsson, T.; Emanuelsson, M. Land use more than 200 years ago explains current grassland plant diversity in a Swedish agricultural landscape. Biol. Conserv. 2007, 138, 47–59. [Google Scholar] [CrossRef]
- Johansson, L.J.; Hall, K.; Prentice, H.C.; Ihse, M.; Reitalu, T.; Sykes, M.T.; Kindström, M. Semi-natural grassland continuity, long-term land-use change and plant species richness in an agricultural landscape on Öland, Sweden. Landsc. Urban Plan. 2008, 84, 200–211. [Google Scholar] [CrossRef]
- Reitalu, T.; Johansson, L.J.; Sykes, M.T.; Hall, K.; Prentice, H.C. History matters: Village distances, grazing and grassland diversity. J. Appl. Ecol. 2010, 47, 1216–1224. [Google Scholar] [CrossRef]
- Austrheim, G.; Olsson, E.G.A.; Grøntvedt, E. Land-use impact on plant communities in semi-natural sub-alpine grasslands of Budalen, Central Norway. Biol. Conserv. 1999, 87, 369–379. [Google Scholar] [CrossRef]
- Aggemyr, E.; Cousins, S.A.O. Landscape structure and land use history influence changes in island plant composition after 100 years. J. Biogeogr. 2012, 39, 1645–1656. [Google Scholar] [CrossRef]
- Eriksson, O. Species pools in cultural landscapes: Niche construction, ecological opportunity and niche shifts. Ecography 2013, 36, 403–413. [Google Scholar] [CrossRef]
- Poschlod, P.; Bonn, S. Changing dispersal processes in the Central European landscape since the last ice age: An explanation for the actual decrease of plant species richness in different habitats? Acta Bot. Neerl. 1998, 47, 27–44. [Google Scholar]
- Auffret, A.G. Can seed dispersal by human activity play a useful role for the conservation of European grasslands? Appl. Veg. Sci. 2011, 14, 291–303. [Google Scholar] [CrossRef]
- Auffret, A.G.; Schmucki, R.; Reimark, J.; Cousins, S.A.O. Grazing networks provide useful functional connectivity for plants in fragmented systems. J. Veg. Sci. 2012, 23, 970–977. [Google Scholar] [CrossRef]
- Eriksson, O.; Cousins, S.A.O. Land-use history and fragmentation of traditionally managed grasslands in Scandinavia. J. Veg. Sci. 2002, 13, 743–748. [Google Scholar] [CrossRef]
- Eriksson, O.; Cousins, S.A.O.; Lindborg, R. Land use history and the build-up and decline of species richness in Scandinavian semi-natural grasslands. In Pastoral Systems in Marginal Environments; Milne, J.A., Ed.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2005; pp. 51–60. [Google Scholar]
- Öster, M.; Cousins, S.A.O.; Eriksson, O. Size and heterogeneity rather than landscape context determine plant species richness in semi-natural grasslands. J. Veg. Sci. 2007, 18, 859–868. [Google Scholar] [CrossRef]
- Wilson, J.B.; Peet, R.K.; Dengler, J.; Pärtel, M. Plant species richness: The world records. J. Veg. Sci. 2012, 23, 796–802. [Google Scholar] [CrossRef]
- Berglund, B.E.; Persson, T.; Björkman, L. Late Quaternary landscape and vegetation diversity in a North European perspective. Quat. Int. 2008, 184, 187–194. [Google Scholar] [CrossRef]
- Berglund, B.E.; Gaillard, M.-J.; Björkman, L.; Persson, T. Long-term changes in floristic diversity in southern Sweden: Palynological richness, vegetation dynamics and land use. Veg. Hist. Archaeobot. 2008, 17, 573–583. [Google Scholar] [CrossRef]
- Pärtel, M.; Helm, A.; Reitalu, T.; Liira, J.; Zobel, M. Grassland diversity related to the late Iron Age human population density. J. Ecol. 2007, 95, 574–582. [Google Scholar] [CrossRef]
- Berg, Å.; Ahrné, K.; Öckinger, E.; Svensson, R.; Söderström, B. Butterfly distribution and abundance is affected by variation in the Swedish forest-farmland landscape. Biol. Conserv. 2011, 144, 2819–2831. [Google Scholar] [CrossRef]
- Wretenberg, J.; Pärt, T.; Berg, Å. Changes in local species richness of farmland birds in relation to land-use changes and landscape structure. Biol. Conserv. 2000, 143, 375–381. [Google Scholar] [CrossRef]
- Van Vuure, C. Retracing the Aurochs: History, Morphology and Ecology of an Extinct Wild Ox; Pensoft Publishers: Sofia, Bulgaria, 2005. [Google Scholar]
- Eriksson, O. Regional dynamics of plants: A review of evidence for remnant, source-sink and metapopulations. Oikos 1996, 77, 248–258. [Google Scholar] [CrossRef]
- Johansson, V.A.; Cousins, S.A.O.; Eriksson, O. Remnant populations and plant functional traits in abandoned semi-natural grasslands. Folia Geobot. 2011, 46, 165–179. [Google Scholar] [CrossRef]
- Auffret, A.G.; Cousins, S.A.O. Past and present management influences the seed bank and seed rain in a rural landscape mosaic. J. Appl. Ecol. 2011, 48, 1278–1285. [Google Scholar] [CrossRef]
- Plue, J.; Cousins, S.A.O. Temporal dispersal in fragmented landscapes. Biol. Conserv. 2013, 160, 250–262. [Google Scholar] [CrossRef]
- Cousins, S.A.O. Extinction debt in fragmented grasslands: Paid or not? J. Veg. Sci. 2009, 20, 3–7. [Google Scholar] [CrossRef]
- Cousins, S.A.O.; Vanhoenacker, D. Detection of extinction debt depends on scale and specialization. Biol. Conserv. 2011, 144, 782–787. [Google Scholar] [CrossRef]
- Tilman, D.; May, R.M.; Lehman, C.L.; Nowak, M.A. Habitat destruction and the extinction debt. Nature 1994, 371, 65–66. [Google Scholar] [CrossRef]
- Cousins, S.A.O.; Ohlson, H.; Eriksson, O. Effects of historical and present fragmentation on plant species diversity in semi-natural grasslands in Swedish rural landscapes. Landsc. Ecol. 2007, 22, 723–730. [Google Scholar] [CrossRef]
- Eriksson, Å. Regional distribution of Thymus serpyllum: Management history and dispersal limitation. Ecography 1998, 21, 35–43. [Google Scholar] [CrossRef]
- Eriksson, O.; Kiviniemi, K. Site occupancy, recruitment and extinction thresholds in grassland plants: An experimental study. Biol. Conserv. 1999, 87, 319–325. [Google Scholar] [CrossRef]
- Purschke, O.; Sykes, M.T.; Reitalu, T.; Poschlod, P.; Prentice, H.C. Linking landscape history and dispersal traits in grassland plant communities. Oecologia 2012, 168, 773–783. [Google Scholar] [CrossRef]
- Öster, M.; Ask, K.; Cousins, S.A.O.; Eriksson, O. Dispersal and establishment limitation reduces the potential for successful restoration of semi-natural grassland communities on former arable fields. J. Appl. Ecol. 2009, 46, 1266–1274. [Google Scholar]
- Cousins, S.A.O.; Lindborg, R. Remnant grassland habitats as source communities for plant diversification in agricultural landscapes. Biol. Conserv. 2008, 141, 233–240. [Google Scholar] [CrossRef]
- Lindborg, R.; Eriksson, O. Effects of restoration on plant species richness and composition in Scandinavian semi-natural grasslands. Restor. Ecol. 2004, 12, 318–326. [Google Scholar] [CrossRef]
- Cousins, S.A.O.; Aggemyr, E. The influence of field shape, area and surrounding landscape on plant species richness in grazed ex-fields. Biol. Conserv. 2008, 141, 126–135. [Google Scholar] [CrossRef]
- Öster, M.; Ask, K.; Römermann, C.; Tackenberg, O.; Eriksson, O. Plant colonization of ex-arable fields from adjacent species-rich grasslands: The importance of dispersal vs. recruitment ability. Agric. Ecosyst. Environ. 2009, 130, 93–99. [Google Scholar] [CrossRef]
- The European Landscape Convention. Council of Europe. Available online: http://www.coe.int/t/dg4/cultureheritage/heritage/landscape (accessed on 10 March 2014).
- Sweden’s Environmental Objectives. Available online: http://www.miljomal.nu (accessed on 10 March 2014).
- Orians, G.H.; Heerwagen, J.H. Evolved responses to landscapes. In The Adapted Mind: Evolutionary Psychology and the Generation of Culture; Barkow, J.H., Cosmides, L., Tooby, J., Eds.; Oxford University Press: Oxford, UK, 1992; pp. 555–579. [Google Scholar]
- Kaplan, S. Aethetics, affect, and cognition: Environmental preference from an evolutionary perspective. Environ. Behav. 1987, 19, 3–32. [Google Scholar] [CrossRef]
- Pinker, S. The cognitive niche: Coevolution of intelligence, sociality, and language. Proc. Natl. Acad. Sci. USA 2010, 107, 8993–8999. [Google Scholar] [CrossRef]
- Boyd, R.; Richerson, P.J.; Henrich, J. The cultural niche: Why social learning is essential for human adaptation. Proc. Natl. Acad. Sci. USA 2011, 108, 10918–10925. [Google Scholar] [CrossRef]
- Whiten, A.; Erdal, D. The human socio-cognitive niche and its evolutionary origins. Philos. Trans. R. Soc. B 2012, 367, 2119–2129. [Google Scholar] [CrossRef]
- Stenseke, M. Biodiversity and the local context: Linking seminatural grasslands and their future use to social aspects. Environ. Sci. Policy 2006, 9, 350–359. [Google Scholar] [CrossRef]
- Buijs, A.E. Lay people’s images of nature: Comprehensive frameworks of values, beliefs, and value orientations. Soc. Nat. Resour. 2009, 22, 417–432. [Google Scholar] [CrossRef]
- Fischer, A. On the role of ideas of human nature in shaping attitudes towards environmental governance. Hum. Ecol. 2010, 38, 123–135. [Google Scholar] [CrossRef]
- Van der Windt, H.J.; Swart, J.A.A.; Keulartz, J. Nature and landscape planning: Exploring the dynamics of valuation, the case of the Netherlands. Landsc. Urban Plan. 2007, 79, 218–228. [Google Scholar] [CrossRef]
- Greider, T.; Garkovich, L. Landscapes: The social construction of nature and the environment. Rural Soc. 1994, 59, 1–24. [Google Scholar] [CrossRef]
- Nassauer, J.I. Culture and changing landscape structure. Landsc. Ecol. 1995, 10, 229–237. [Google Scholar] [CrossRef]
- Bang, M.; Medin, D.L.; Atran, S. Cultural mosaics and mental models of nature. Proc. Natl. Acad. Sci. USA 2007, 104, 13868–13874. [Google Scholar]
- Buijs, A.E.; Pedroli, B.; Luginbühl, Y. From hiking through farmland to farming in a leisure landscape: Changing social perceptions of the European landscape. Landsc. Ecol. 2006, 21, 375–389. [Google Scholar] [CrossRef]
- Lindborg, R.; Stenseke, M.; Cousins, S.A.O.; Bengtsson, J.; Berg, Å.; Gustafsson, T.; Sjödin, N.E.; Eriksson, O. Investigating biodiversity trajectories using scenarios—Lessons from two contrasting agricultural landscapes. J. Environ. Manag. 2009, 91, 499–508. [Google Scholar] [CrossRef]
- Saltzman, K.; Head, L.; Stenseke, M. Do cows belong in nature? The cultural basis of agriculture in Sweden and Australia. J. Rural Stud. 2011, 27, 54–62. [Google Scholar] [CrossRef]
- Stenseke, M. Local participation in cultural landscape maintenance: Lessons from Sweden. Land Use Policy 2009, 26, 214–223. [Google Scholar] [CrossRef]
- Pihlgren, A.; Lennartsson, T. Shrub effects on herbs and grasses in semi-natural grasslands: Positive, negative or neutral relationships? Grass For. Sci. 2008, 63, 9–21. [Google Scholar] [CrossRef]
- Wästfält, A.; Saltzman, K.; Gräslund Berg, E.; Dahlberg, A. Landscape care paradoxes: Swedish landscape care arrangements in a European context. Geoforum 2012, 43, 1171–1181. [Google Scholar] [CrossRef]
- Kearney, A.R.; Bradley, G.A.; Petrich, C.H.; Kaplan, R.; Kaplan, S.; Simpson-Colebank, D. Public perception as support for scenic quality regulation in a nationally treasured landscape. Landsc. Urban Plan. 2008, 87, 117–128. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Eriksson, O.; Cousins, S.A.O. Historical Landscape Perspectives on Grasslands in Sweden and the Baltic Region. Land 2014, 3, 300-321. https://doi.org/10.3390/land3010300
Eriksson O, Cousins SAO. Historical Landscape Perspectives on Grasslands in Sweden and the Baltic Region. Land. 2014; 3(1):300-321. https://doi.org/10.3390/land3010300
Chicago/Turabian StyleEriksson, Ove, and Sara A. O. Cousins. 2014. "Historical Landscape Perspectives on Grasslands in Sweden and the Baltic Region" Land 3, no. 1: 300-321. https://doi.org/10.3390/land3010300